
Nanoclay/Polymer-Based Hydrogels and Enzyme-Loaded Nanostructures for Wound Healing Applications


 ,
,  , and
, and 
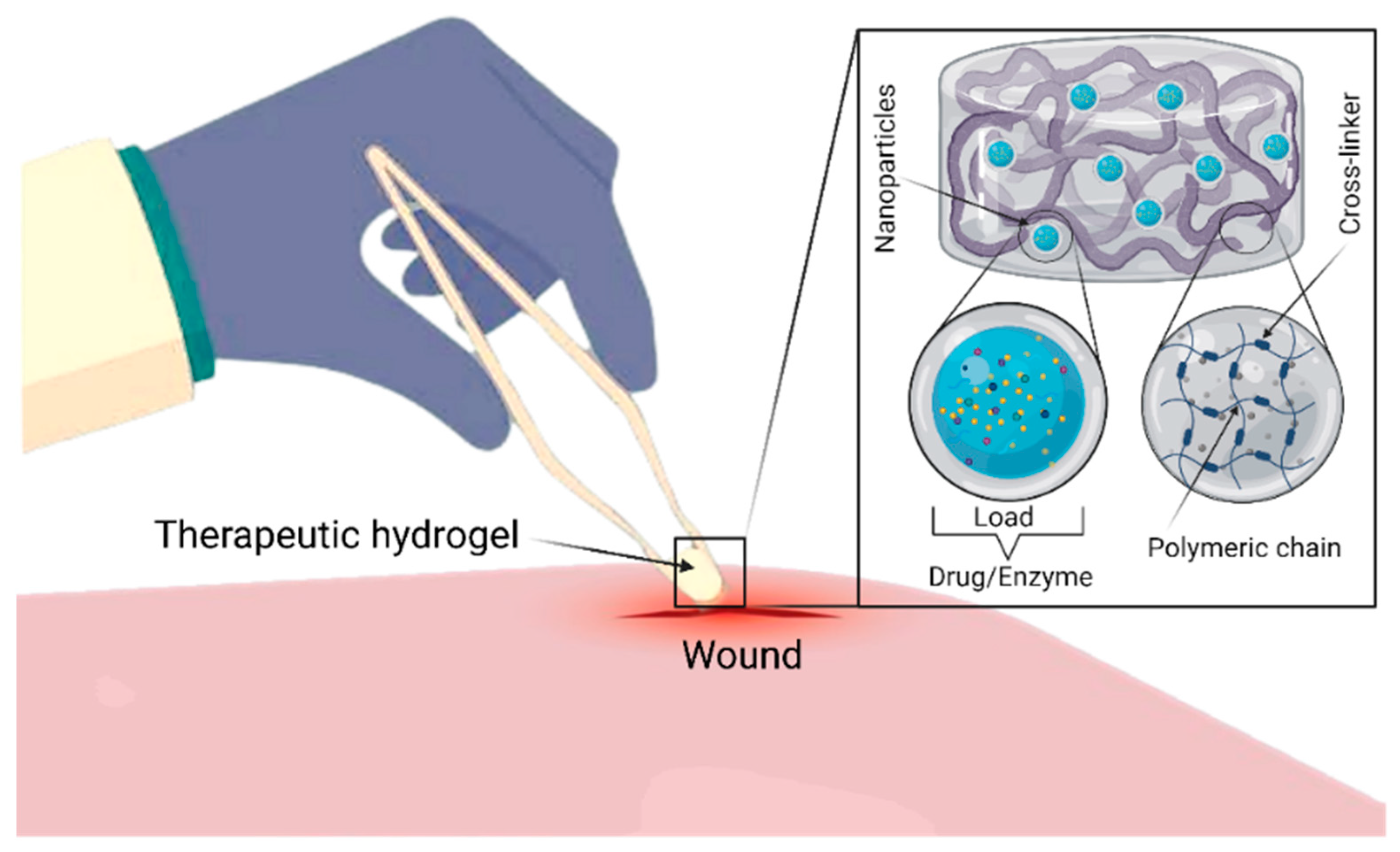{kind=link}
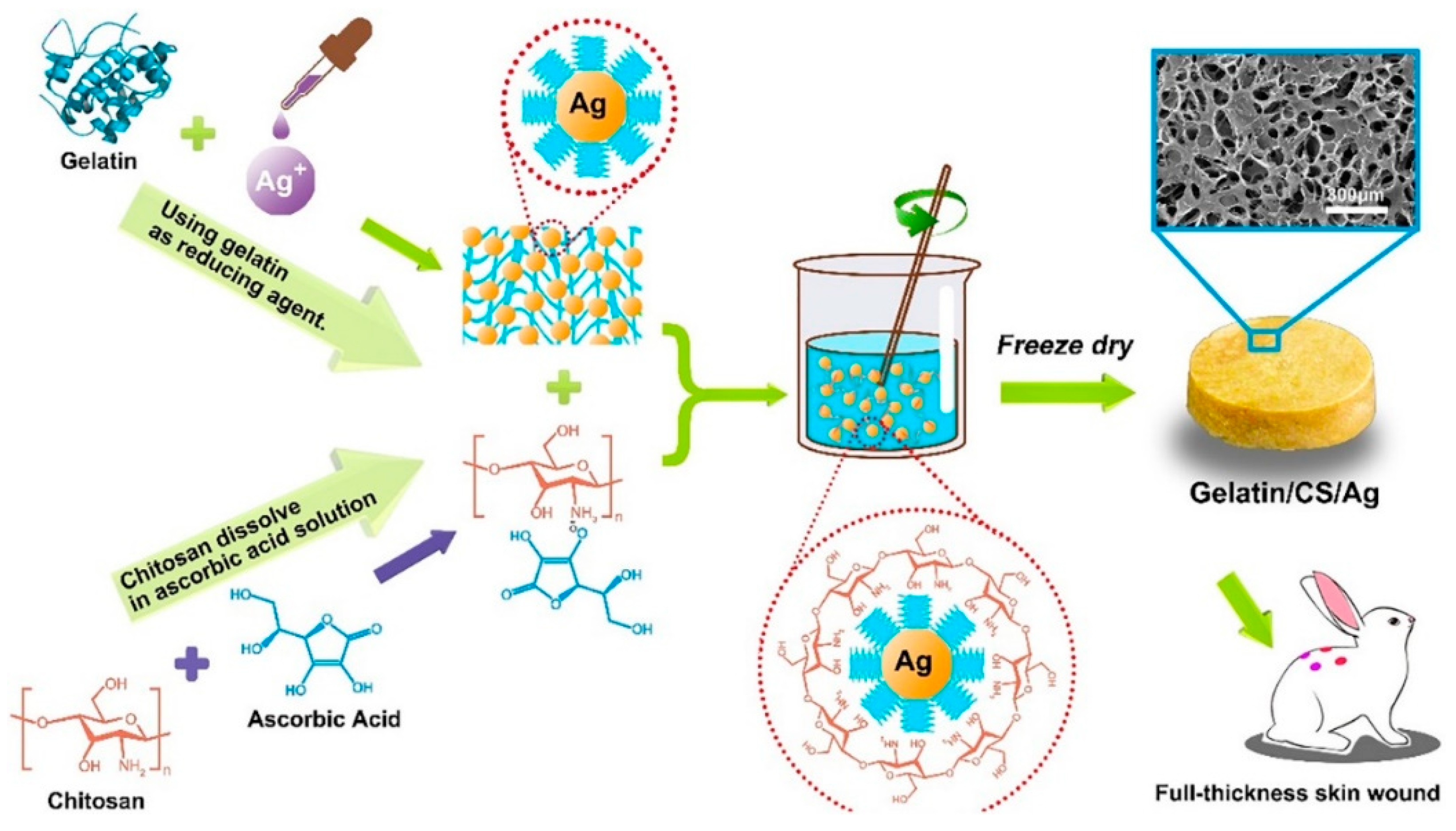{kind=link}
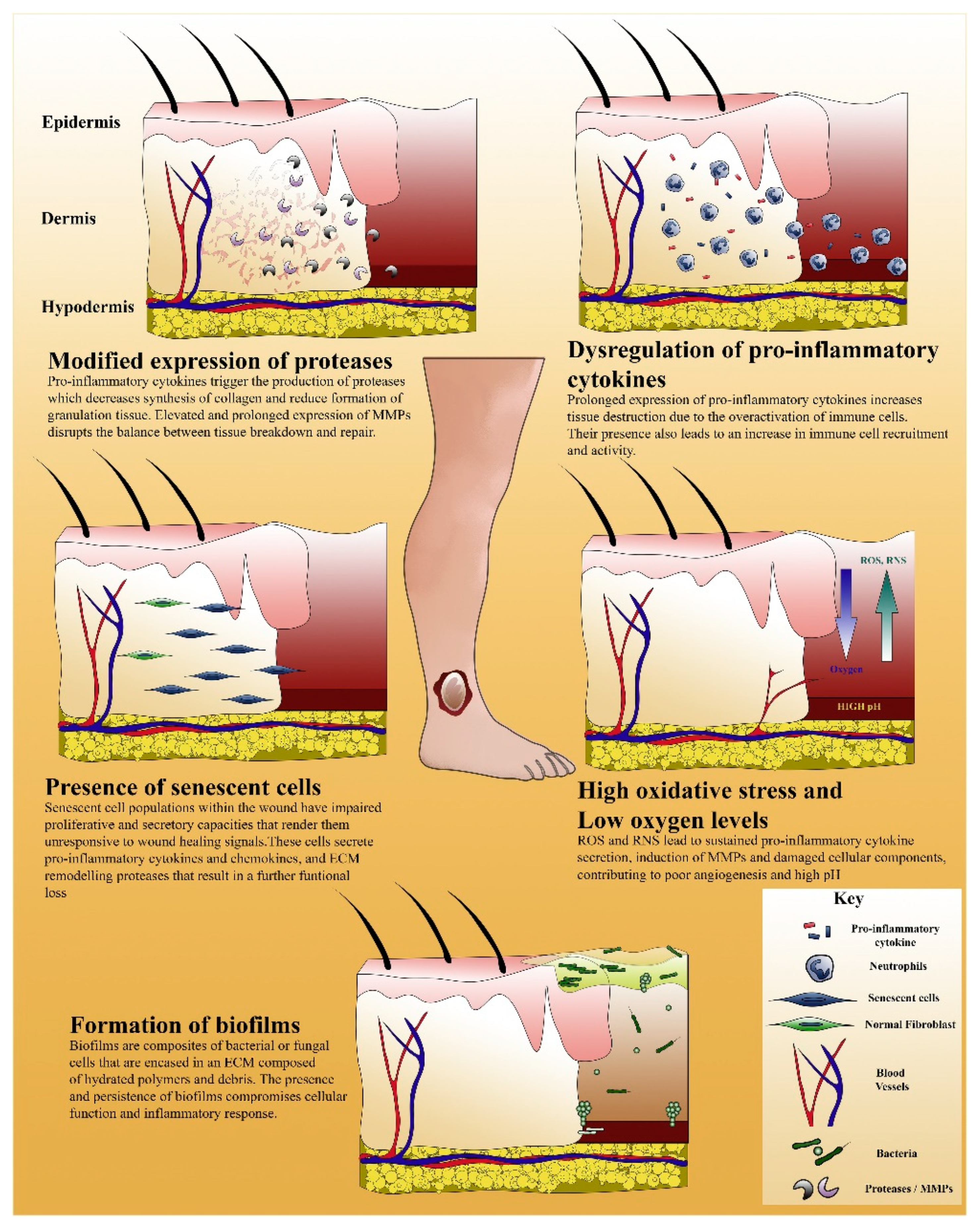{kind=link}
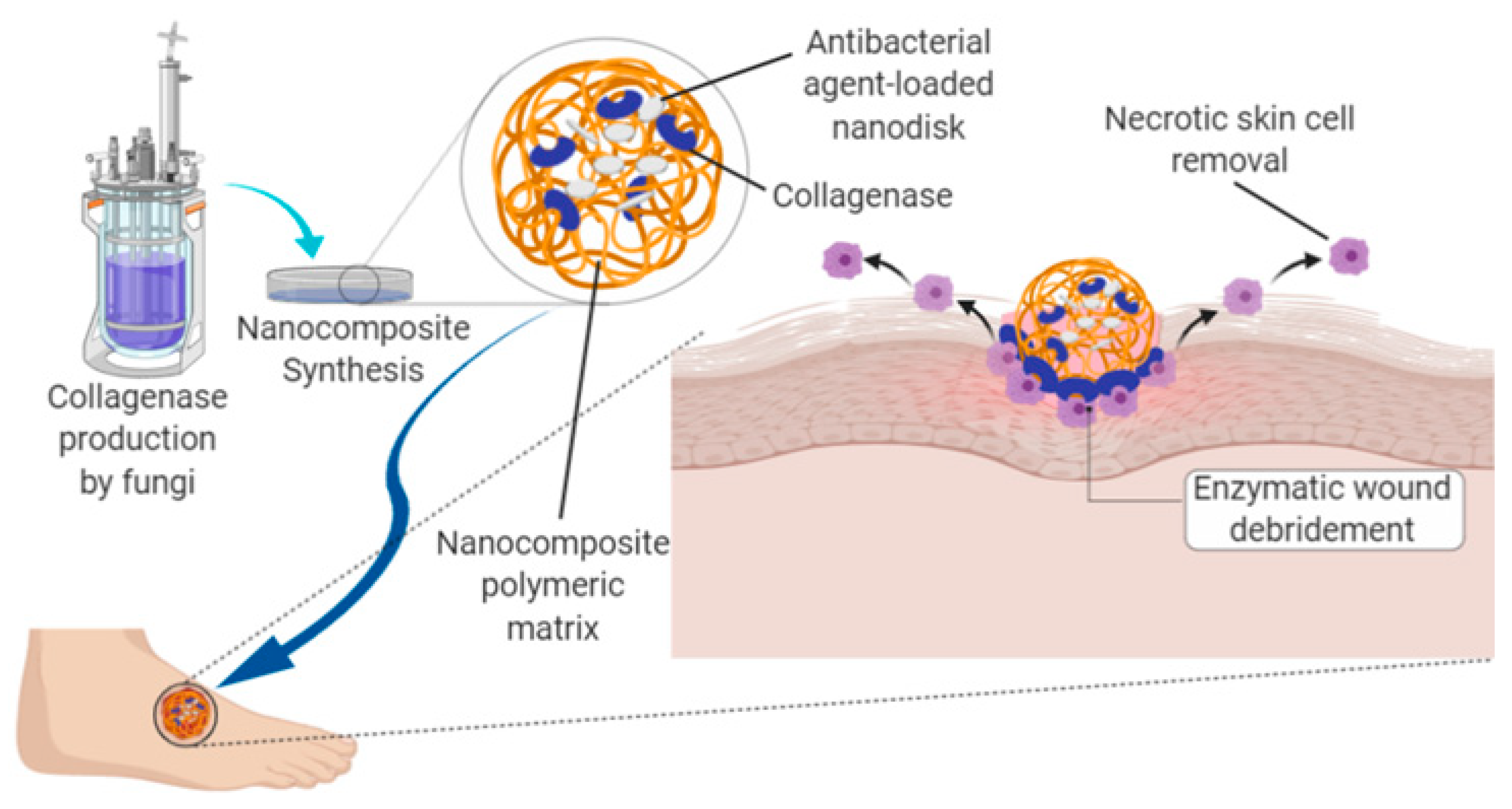{kind=link}
{kind=link}
Abstract
1. Introduction
2. Chitosan-Based Composite Materials
3. Nanoclay-Based Materials
4. Trypsin Delivery via Nanostructured Polymeric Materials
4.1. Nanoparticle-Immobilized Trypsin
4.2. Nanofiber-Immobilized Trypsin
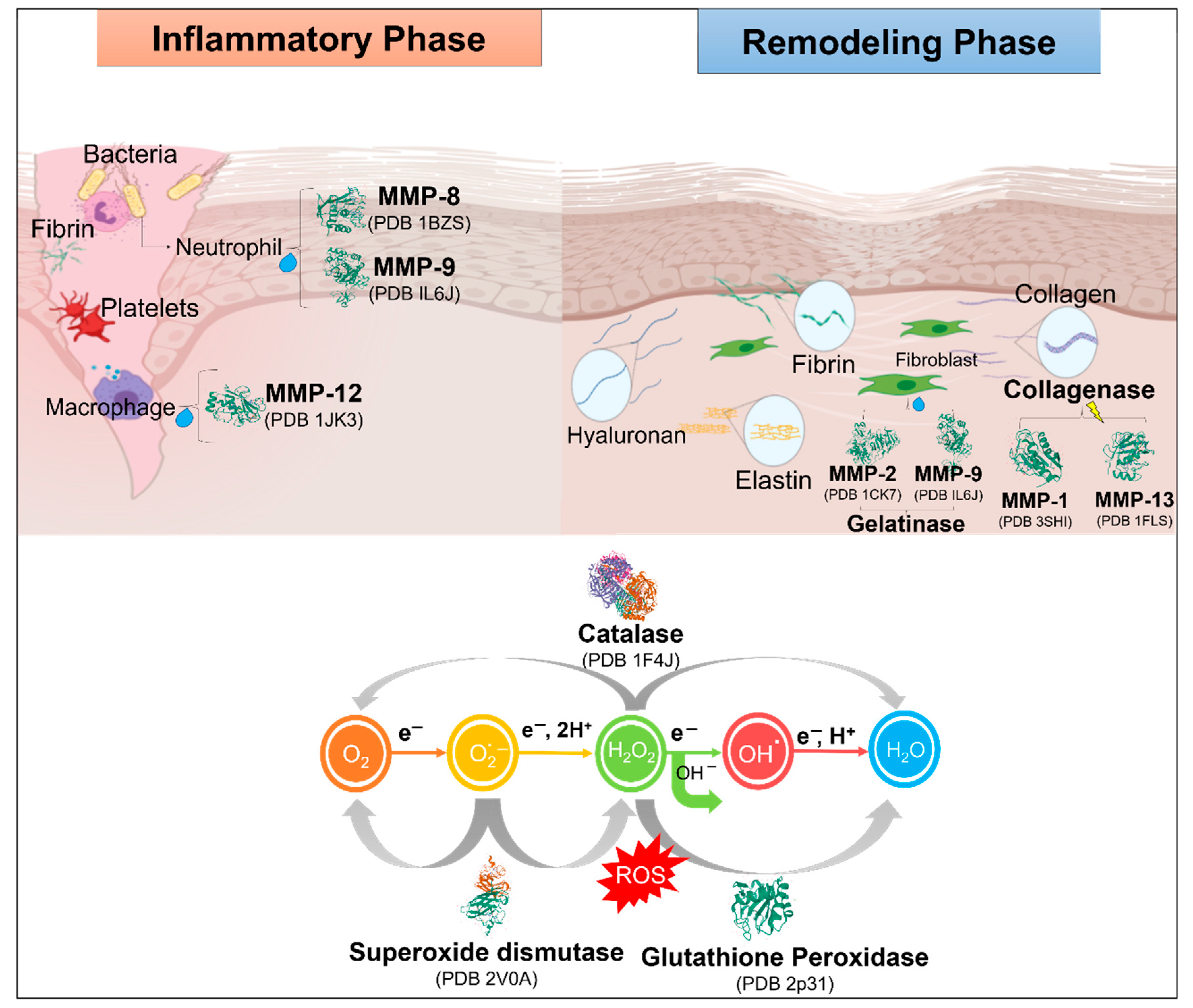
5. Complications in Wound Healing
6. Role of Enzymes in Topical Wound Healing
Role of Proteases in the Wound Healing Process
- ✓
- They are of high importance in wound management, and maintaining a balance between them prevents irregularities such as immoderate ECM degradation and depositing, thus resulting in impaired healing. An unbalanced process usually leads to abnormal scarring [55]. Furthermore, when conjugated to other complications, such as diabetes, it leads to continuous inflammation and non-healing [56].
- ✓
- Enzymes that catalyze protein hydrolysis into minor portions/particles such as peptides. They can be grouped based on their protein substrate, optimal pH, their cut specificity (the amino acid peptide bond hydrolyzing), and their catalytic site configuration.
- ✓
- Proteases are divided into two main categories: endopeptidases and exopeptidases.
- ✓
- Exopeptidases focus on the N- and C-terminations of peptide bonds, while endopeptidases separate peptide linkages apart from the endpoint of the protein substrate.
- ✓
- Based upon the active site configuration, proteases are classified as serine, aspartic, cysteine and metalloproteases.
- ✓
- Other proteases that do not fall into the conventional classifications are the ATP-dependent proteases.
- ✓
- A characteristic major function of proteolytic enzymes is to regulate the ratio between tissue regeneration and tissue degeneration.
- ✓
- They have an essential role in the transfer and stimulation of fibroblasts, ECM restoration and growth factor activation.
- ✓
- During the different stages of inflammation, they take part in damaged tissue removal (debridement) and influence bacterial load in the wound area.
- ✓
- During the stage of cell proliferation, they are quickly found at the formation of blood vessels to facilitate vascularization during angiogenesis.
- ✓
- In the ending phase of growth and restructuring, they absorb the ECM and assist in tissue regeneration. It has been estimated that over 100 enzymes are involved in this phase.
- ✓
- The proteases that are involved the most in the wound healing process are MMPs and metalloproteinases of the thrombospondin domain (ADAM-TS, tolloids, serine proteases, pappalysins, and meprins [57].
7. Classification of Proteases Involved in the Wound Healing Process
- ✓
- Catalytic mechanism: Enzymes that exhibit proteolytic activity are grouped as glutamic, cysteine, threonine, serine, asparagine, aspartic, or metalloproteases. Stimulation of many trypsin-like proteases of the serine group requires proteolytic processing of an idle zymogen precursor. Practically all PA Clan proteases utilize the canonical catalytic triad and hydrolyze the peptide bond via two tetrahedral intermediates [72].
- ✓
- PA Clan Proteases: The largest family of serine proteases is the PA proteases clan that is present in the trypsin fold and is possibly the best-studied group of enzymes currently [73]. Most proteases of the PA clan have specificity for substrates similar to trypsin and prefer the Lys and/or Arg chains at the P1 position. Additionally, trypsin and chymotrypsin are known to be digestive enzymes that break polypeptide chains of positively charged or large hydrophobic residues, respectively. This type of proteases relies on several crucial biological processes such as blood clotting and immune response, which involve torrents of sequential zymogen activation [74].
- ✓
- E*form: The critical serine protease in recent kinetic studies on thrombin showed that the blood coagulation pathway asserts for unpredictable plasticity of the trypsin fold [69]. Thrombin exists in three forms at equilibrium, such as Na+-free form E, Na+-bound form E, and E* [75]. Where Na+ are the low and high activity configurations of the enzyme, Na+-bound being the cause of the procoagulant, prothrombotic, and signaling activities. Another form, E*, is in balance with E and is idle toward the substrate and, therefore, it is unable to link Na+ [69].
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Feynman, R.P. There’s plenty of room at the bottom. California Institute of Technology. Eng. Sci. Mag. 1960, 4, 23–36. [Google Scholar]
- Binnig, G.; Rohrer, H. Scanning Tunneling Microscope. U.S. Patent 4,343,993A, 10 August 1982. [Google Scholar]
- Vithun, S.N.; Reddy, T.N.; Vinod, P. Investigation on Nanoscale Imaging of Gold Sputtered Sample by Scanning Tunneling Microscope. Mater. Today Proc. 2020, 22, 2439–2445. [Google Scholar] [CrossRef]
- Gleiter, H. Nanostructured materials: Basic concepts and microstructure. Acta Mater. 2020, 48, 1–29. [Google Scholar] [CrossRef]
- Van Assenbergh, P.; Meinders, E.; Geraedts, J.; Dodou, D. Nanostructure and microstructure fabrication: From desired properties to suitable processes. Small 2018, 14, 1703401. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H. New Insights on Unique Features and Role of Nanostructured Materials in Cosmetics. Cosmetics 2020, 7, 24. [Google Scholar] [CrossRef]
- Lata, S.; Sharma, G.; Joshi, M.; Kanwar, P.; Mishra, T. Role of nanotechnology in drug delivery. Int. J. Nanotechnol. Nanosci. 2017, 5, 1–29. [Google Scholar]
- Pelegrino, M.T.; Seabra, A.B. Chitosan-based nanomaterials for skin regeneration. Med. Sci. 2017, 4, 352–381. [Google Scholar] [CrossRef]
- Paumo, H.K.; Das, R.; Bhaumik, M.; Maity, A. Visible-Light-Responsive Nanostructured Materials for Photocatalytic Degradation of Persistent Organic Pollutants in Water. In Green Methods for Wastewater Treatment; Springer: Cham, Switzerland, 2020; pp. 1–29. [Google Scholar]
- Jacob, J.; Haponiuk, J.T.; Thomas, S.; Gopi, S. Biopolymer based nanomaterials in drug delivery systems: A review. Mater. Today Chem. 2018, 9, 43–55. [Google Scholar] [CrossRef]
- Aithal, P.S. Nanotechnology Innovations & Business Opportunities: A Review. Int. J. Manag. It Eng. 2016, 6, 182–204. [Google Scholar]
- Peña-Parás, L.; Sánchez-Fernández, J.A.; Vidaltamayo, R. Nanoclays for biomedical applications. In Handbook of Ecomaterials; Springer: Cham, Switzerland, 2018; pp. 1–19. [Google Scholar]
- Dhama, K.; Munjal, A.; Iqbal, H.M. Recent advances and novel strategies for the development of biomedical therapeutics: State-of-the-art and future perspectives. Int. J. Pharmacol. 2017, 13, 929–933. [Google Scholar] [CrossRef]
- Hamdan, S.; Pastar, I.; Drakulich, S. Nanotechnology-Driven Therapeutic Interventions in Wound Healing: Potential Uses and Applications. ACS Cent. Sci. 2017, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Negut, I.; Grumezescu, V.; Grumezescu, A.M. Treatment strategies for infected wounds. Molecules 2018, 23, 2392. [Google Scholar] [CrossRef] [PubMed]
- Elgadir, M.A.; Uddin, M.S.; Ferdosh, S.; Adam, A.; Chowdhury, A.J.K.; Sarker, M.Z.I. Impact of chitosan composites and chitosan nanoparticle composites on various drug delivery systems: A review. J. Food Drug Anal. 2015, 23, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Izumi, R.; Osaki, T.; Ifuku, S.; Morimoto, M.; Saimoto, H.; Minami, S.; Okamoto, Y. Chitin, chitosan, and its derivatives for wound healing: Old and new materials. J. Funct. Biomater. 2015, 6, 104–142, Retraction on 2018, 9, 38. [Google Scholar] [CrossRef]
- Rubentheren, V.; Ward, T.A.; Chee, C.Y.; Tang, C.K. Processing and analysis of chitosan nanocomposites reinforced with chitin whiskers and tannic acid as a crosslinker. Carbohydr. Polym. 2015, 115, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Lavertu, M.; Filion, D.; Buschmann, M.D. Heat-induced transfer of protons from chitosan to glycerol phosphate produces chitosan precipitation and gelation. Biomacromolecules 2008, 9, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Cheng, J.; Yu, K. In situ reduction of silver nanoparticles by gelatin to obtain porous silver nanoparticle/chitosan composites with enhanced antimicrobial and wound-healing activity. Int. J. Biol. Macromol. 2019, 121, 633–642. [Google Scholar] [CrossRef]
- Mahdavinia, G.R.; Shokri, E. Synthesis and characterization of magnetic amidoximated chitosan-g poly (polyacrylonitrile)/laponite RD nanocomposites with enhanced adsorption capacity for Cu 2+. Turk. J. Chem. 2017, 41, 135–152. [Google Scholar] [CrossRef]
- Sheikhi, A.; Afewerki, S.; Oklu, R.; Gaharwar, A.K.; Khademhosseini, A. Effect of ionic strength on shear-thinning nanoclay–polymer composite hydrogels. Biomater. Sci. 2018, 6, 2073–2083. [Google Scholar] [CrossRef]
- Maeda, T.; Kitagawa, M.; Hotta, A.; Koizumi, S. Thermo-responsive nanocomposite hydrogels based on PEG-b-PLGA diblock copolymer and laponite. Polymers 2019, 11, 250. [Google Scholar] [CrossRef]
- Cidonio, G.; Alcala-Orozco, C.R.; Lim, K.S.; Glinka, M.; Mutreja, I.; Kim, Y.-H.; I Dawson, J.; Woodfield, T.B.F.; Oreffo, R.O.C. Osteogenic and angiogenic tissue formation in high fidelity nanocomposite Laponite-gelatin bioinks. Biofabrication 2019, 11, 035027. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.E.; Sawvel, A.M.; Zheng, N.; Stucky, G.D. Controlling bioprocesses with inorganic surfaces: Layered clay hemostatic agents. Chem. Mater. 2007, 19, 4390–4392. [Google Scholar] [CrossRef]
- Sun, J.; Yang, L.; Jiang, M.; Xu, B. Stability and activity of immobilized trypsin on carboxymethyl chitosan-functionalized magnetic nanoparticles cross-linked with carbodiimide and glutaraldehyde. J. Chromatogr. B 2017, 1054, 57–63. [Google Scholar] [CrossRef]
- Cacicedo, M.L.; Manzo, R.M.; Municoy, S.; Bonazza, H.L.; Islan, G.A.; Desimone, M.; Bellino, M.; Mammarella, E.J.; Castro, G.R. Immobilized enzymes and their applications. In Advances in Enzyme Technology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 169–200. [Google Scholar]
- De Queiroz, J.L.C.; Costa, R.O.D.A.; Matias, L.L.R.; De Medeiros, A.F.; Gomes, A.F.T.; Pais, T.D.S.; Passos, T.S.; Maciel, B.L.L.; Dos Santos, E.A.; Morais, A.H.D.A. Chitosan-whey protein nanoparticles improve encapsulation efficiency and stability of a trypsin inhibitor isolated from Tamarindus indica L. Food Hydrocoll. 2018, 84, 247–256. [Google Scholar] [CrossRef]
- Sun, J.; Xu, B.; Shi, Y.; Yang, L.; Ma, H.L. Activity and stability of trypsin immobilized onto chitosan magnetic nanoparticles. Adv. Mater. Sci. Eng. 2017, 1457072. [Google Scholar] [CrossRef]
- Atacan, K.; Çakıroğlu, B.; Özacar, M. Covalent immobilization of trypsin onto modified magnetite nanoparticles and its application for casein digestion. Int. J. Biol. Macromol. 2017, 97, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.K.; Kim, B.C.; Jun, S.H.; Chang, M.S.; Lopez-Ferrer, D.; Smith, R.D.; Gu, M.B.; Lee, S.-W.; Kim, B.S.; Kim, J. Robust trypsin coating on electrospun polymer nanofibers in rigorous conditions and its uses for protein digestion. Biotechnol. Bioeng. 2010, 107, 917–923. [Google Scholar] [CrossRef]
- Silva, T.R.; Rodrigues, D.P.; Rocha, J.M.; Gil, M.H.; Pinto, S.C.; Lopes-da-Silva, J.A.; Guiomar, A.J. Immobilization of trypsin onto poly (ethylene terephthalate)/poly (lactic acid) nonwoven nanofiber mats. Biochem. Eng. J. 2015, 104, 48–56. [Google Scholar] [CrossRef]
- Jasti, L.S.; Dola, S.R.; Kumaraguru, T.; Bajja, S.; Fadnavis, N.W.; Addepally, U.; Rajdeo, K.; Ponrathnam, S.; Deokar, S. Protein-coated polymer as a matrix for enzyme immobilization: Immobilization of trypsin on bovine serum albumin-coated allyl glycidyl ether–ethylene glycol dimethacrylate copolymer. Biotechnol. Prog. 2014, 30, 317–323. [Google Scholar] [CrossRef]
- Franz, M.G.; Steed, D.L.; Robson, M.C. Optimizing Healing of the Acute Wound by Minimizing Complications. Curr. Probl. Surg. 2007, 44, 691–763. [Google Scholar] [CrossRef]
- Stojadinovic, A.; Carlson, J.W.; Schultz, G.S.; Davis, T.A.; Elster, E.A. Topical advances in wound care. Gynecol. Oncol. 2008, 111, S70–S80. [Google Scholar] [CrossRef]
- Guo, S.; DiPietro, L.A. Factors Affecting Wound Healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef]
- Chin, J.S.; Madden, L.; Chew, S.Y.; Becker, D.L. Drug therapies and delivery mechanisms to treat perturbed skin wound healing. Adv. Drug Deliv. Rev. 2019, 149, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. 2016, 58, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Demidova-Rice, T.N.; Durham, J.T.; Herman, I.M. Wound Healing Angiogenesis: Innovations and Challenges in Acute and Chronic Wound Healing. Adv. Wound Care 2012, 1, 17–22. [Google Scholar] [CrossRef]
- Kurahashi, T.; Fujii, J. Roles of Antioxidative Enzymes in Wound Healing. J. Dev. Biol. 2015, 3, 57–70. [Google Scholar] [CrossRef]
- Auf dem Keller, U.; Kümin, A.; Braun, S.; Werner, S. Reactive Oxygen Species and Their Detoxification in Healing Skin Wounds. J. Investig. Dermatol. Symp. Proc. 2006, 11, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Dunning, S.; ur Rehman, A.; Tiebosch, M.H.; Hannivoort, R.A.; Haijer, F.W.; Woudenberg, J.; van den Heuvel, F.A.J.; Buist-Homan, M.; Faber, K.N.; Moshage, H. Glutathione and antioxidant enzymes serve complementary roles in protecting activated hepatic stellate cells against hydrogen peroxide-induced cell death. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2013, 1832, 2027–2034. [Google Scholar] [CrossRef]
- Westby, M.J.; Dumville, J.C.; Stubbs, N.; Norman, G.; Wong, J.K.; Cullum, N.; Riley, R.D. Protease activity as a prognostic factor for wound healing in venous leg ulcers. Cochrane Database Syst. Rev. 2018, 9. [Google Scholar] [CrossRef]
- Boran, R.; Ugur, A.; Sarac, N. Investigation of Hyaluronidase, Collagenase and Elastase Inhibitory Potentials and Comparative Evaluation of the Antimicrobial, Antioxidant and Homeostatic Activities of Two Natural Polysaccharides. Süleyman Demirel Üniversitesi Fen Bilimleri Enstitüsü Derg. 2018, 22, 1182–1189. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive compounds from natural resources against skin aging. Phytomedicine 2011, 19, 64–73. [Google Scholar] [CrossRef] [PubMed]
- McCarty, S.M.; Percival, S.L. Proteases and Delayed Wound Healing. Adv. Wound Care 2013, 2, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Frykberg, R.G.; Banks, J. Challenges in the Treatment of Chronic Wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef]
- Eming, S.A.; Hammerschmidt, M.; Krieg, T.; Roers, A. Interrelation of immunity and tissue repair or regeneration. Semin. Cell Dev. Biol. 2009, 20, 517–527. [Google Scholar] [CrossRef]
- Olczyk, P.; Mencner, Ł.; Komosinska-Vassev, K. The role of the extracellular matrix components in cutaneous wound healing. Biomed. Res. Int. 2014, 2014, 1–8. [Google Scholar]
- Powers, J.G.; Higham, C.; Broussard, K.; Phillips, T.J. Wound healing and treating wounds: Chronic wound care and management. J. Am. Acad. Dermatol. 2016, 74, 607–625. [Google Scholar] [CrossRef]
- Singh, N.; Bhattacharyya, D. Proteases in wound healing and immunity. In Proteases in Human Diseases; Springer: Singapore, 2017; pp. 147–170. [Google Scholar]
- Thapa, R.K.; Diep, D.B.; Tønnesen, H.H. Topical antimicrobial peptide formulations for wound healing: Current developments and future prospects. Acta Biomater. 2020, 103, 52–67. [Google Scholar] [CrossRef]
- Xue, M.; Jackson, C.J. Extracellular matrix reorganization during wound healing and its impact on abnormal scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef]
- Pastar, I.; Wong, L.L.; Egger, A.N.; Tomic-Canic, M. Descriptive vs mechanistic scientific approach to study wound healing and its inhibition: Is there a value of translational research involving human subjects? Exp. Dermatol. 2015, 27, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Chakraborti, S.; Chakraborti, T.; Dhalla, N.S. (Eds.) Proteases in Human Diseases; Springer: Singapore, 2017. [Google Scholar]
- Puente, X.S.; Sánchez, L.M.; Overall, C.M.; López-Otín, C. Human and mouse proteases: A comparative genomic approach. Nat. Rev. Genet. 2003, 4, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Fanjul-Fernández, M.; Folgueras, A.R.; Fueyo, A.; Balbín, M.; Suárez, M.F.; Fernández-García, M.S.; Shapiro, S.D.; Freije, J.M.P.; López-Otín, C. Matrix metalloproteinase Mmp-1a is dispensable for normal growth and fertility in mice and promotes lung cancer progression by modulating inflammatory responses. J. Biol. Chem. 2013, 288, 14647–14656. [Google Scholar] [CrossRef]
- Panwar, P.; Butler, G.S.; Jamroz, A.; Azizi, P.; Overall, C.M.; Brömme, D. Aging-associated modifications of collagen affect its degradation by matrix metalloproteinases. Matrix Biol. 2018, 65, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Collier, I.E.; Legant, W.; Marmer, B.; Lubman, O.; Saffarian, S.; Wakatsuki, T.; Elson, E.; Goldberg, G.I. Diffusion of MMPs on the surface of collagen fibrils: The mobile cell surface–collagen substratum interface. PLoS ONE 2011, 6, e24029. [Google Scholar] [CrossRef]
- Tardáguila-García, A.; García-Morales, E.; García-Alamino, J.M.; Álvaro-Afonso, F.J.; Molines-Barroso, R.J.; Lázaro-Martínez, J.L. Metalloproteinases in chronic and acute wounds: A systematic review and meta-analysis. Wound Repair Regen. 2019, 27, 415–420. [Google Scholar] [CrossRef]
- Gomis-Rüth, F.X. Catalytic domain architecture of metzincin metalloproteases. J. Biol. Chem. 2009, 284, 15353–15357. [Google Scholar] [CrossRef]
- Mirastschijski, U.; Dinesh, N.; Baskaran, S.; Wedekind, D.; Gavrilovic, J.; Murray, M.Y.; Bevan, D.; Kelm, S. Novel specific human and mouse stromelysin-1 (MMP-3) and stromelysin-2 (MMP-10) antibodies for biochemical and immunohistochemical analyses. Wound Repair Regen. 2019, 27, 309–323. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Overall, C.M. Molecular determinants of metalloproteinase substrate specificity. Mol. Biotechnol. 2002, 22, 51–86. [Google Scholar] [CrossRef]
- McGuire, J.K.; Li, Q.; Parks, W.C. Matrilysin (matrix metalloproteinase-7) mediates E-cadherin ectodomain shedding in injured lung epithelium. Am. J. Pathol. 2003, 162, 1831–1843. [Google Scholar] [CrossRef]
- Souza, P.M.D.; Bittencourt, M.L.D.A.; Caprara, C.C.; Freitas, M.D.; Almeida, R.P.C.D.; Silveira, D.; Fonseca, Y.M.; Filho, E.X.F.; Junior, A.P.; Magalhães, P.O. A biotechnology perspective of fungal proteases. Braz. J. Microbiol. 2015, 201, 337–346. [Google Scholar] [CrossRef]
- Puente, X.S.; Sánchez, L.M.; Gutiérrez-Fernández, A.; Velasco, G.; López-Otín, C. A genomic view of the complexity of mammalian proteolytic systems. Biochem. Soc. Trans. 2005, 33, 331–334. [Google Scholar] [CrossRef]
- Di Cera, E. Thrombin. Mol. Asp. Med. 2008, 29, 203–254. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D. Twenty-five years of nomenclature and classification of proteolytic enzymes. Biochim. Biophys. Acta BBA Proteins Proteom. 2020, 1868, 140345. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Eckhard, U.; Dufour, A.; Solis, N.; Overall, C.M. Proteolytic Cleavage Mechanisms, Function, and “Omic” Approaches for a Near-Ubiquitous Posttranslational Modification. Chem. Rev. 2018, 118, 1137–1168. [Google Scholar] [CrossRef]
- Jones, J.C.; Rustagi, S.; Dempsey, P.J. ADAM proteases and gastrointestinal function. Annu. Rev. Physiol. 2016, 78, 243–276. [Google Scholar] [CrossRef] [PubMed]
- Flemmig, M.; Melzig, M.F. Serine-proteases as plasminogen activators in terms of fibrinolysis. J. Pharm. Pharmacol. 2012, 64, 1025–1039. [Google Scholar] [CrossRef]
- Pozzi, N.; Chen, R.; Chen, Z.; Bah, A.; Di Cera, E. Rigidification of the autolysis loop enhances Na+ binding to thrombin. Biophys. Chem. 2011, 159, 6–13. [Google Scholar] [CrossRef][Green Version]
- Shoba, E.; Lakra, R.; Kiran, M.S.; Korrapati, P.S. Design and development of papain–urea loaded PVA nanofibers for wound debridement. Rsc Adv. 2014, 4, 60209–60215. [Google Scholar] [CrossRef]
- Martins, V.L.; Caley, M.; O’Toole, E.A. Matrix metalloproteinases and epidermal wound repair. Cell Tissue Res. 2012, 351, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Avila-Rodríguez, M.I.; Meléndez-Martínez, D.; Licona-Cassani, C.; Aguilar-Yañez, J.M.; Benavides, J.; Sánchez, M.L. Practical context of enzymatic treatment for wound healing: A secreted protease approach. Biomed. Rep. 2020, 13, 3–14. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalba-Rodríguez, A.M.; Martínez-González, S.; Sosa-Hernández, J.E.; Parra-Saldívar, R.; Bilal, M.; Iqbal, H.M.N. Nanoclay/Polymer-Based Hydrogels and Enzyme-Loaded Nanostructures for Wound Healing Applications. Gels 2021, 7, 59. https://doi.org/10.3390/gels7020059
Villalba-Rodríguez AM, Martínez-González S, Sosa-Hernández JE, Parra-Saldívar R, Bilal M, Iqbal HMN. Nanoclay/Polymer-Based Hydrogels and Enzyme-Loaded Nanostructures for Wound Healing Applications. Gels. 2021; 7(2):59. https://doi.org/10.3390/gels7020059
Chicago/Turabian StyleVillalba-Rodríguez, Angel M., Sara Martínez-González, Juan Eduardo Sosa-Hernández, Roberto Parra-Saldívar, Muhammad Bilal, and Hafiz M. N. Iqbal. 2021. "Nanoclay/Polymer-Based Hydrogels and Enzyme-Loaded Nanostructures for Wound Healing Applications" Gels 7, no. 2: 59. https://doi.org/10.3390/gels7020059
APA StyleVillalba-Rodríguez, A. M., Martínez-González, S., Sosa-Hernández, J. E., Parra-Saldívar, R., Bilal, M., & Iqbal, H. M. N. (2021). Nanoclay/Polymer-Based Hydrogels and Enzyme-Loaded Nanostructures for Wound Healing Applications. Gels, 7(2), 59. https://doi.org/10.3390/gels7020059