
Wood Coloration and Decay Capabilities of Mycoparasite Scytalidium ganodermophthorum



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Block Decay Test
2.2. Microscopic Examination of Decay
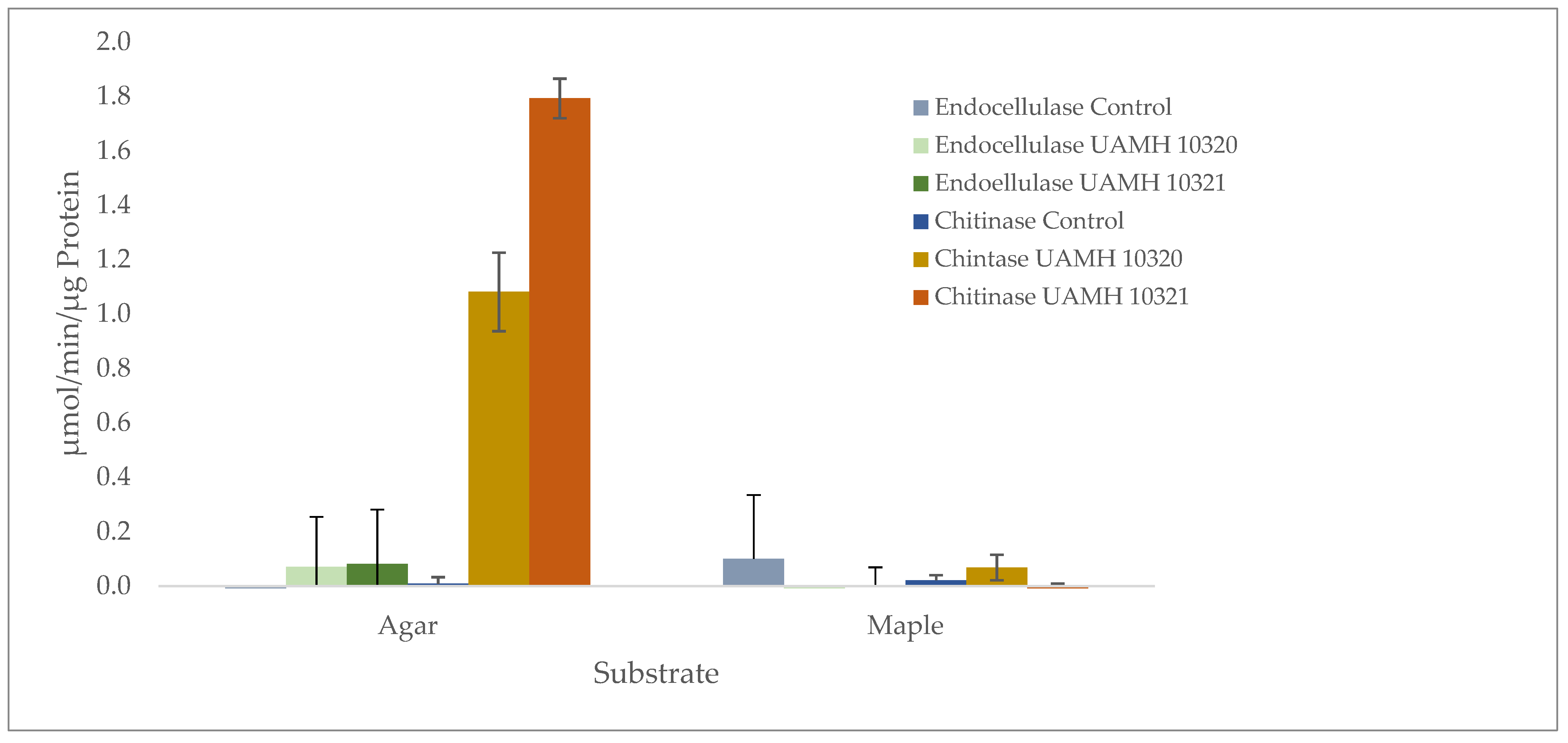
2.3. Enzymatic Analysis
3. Results
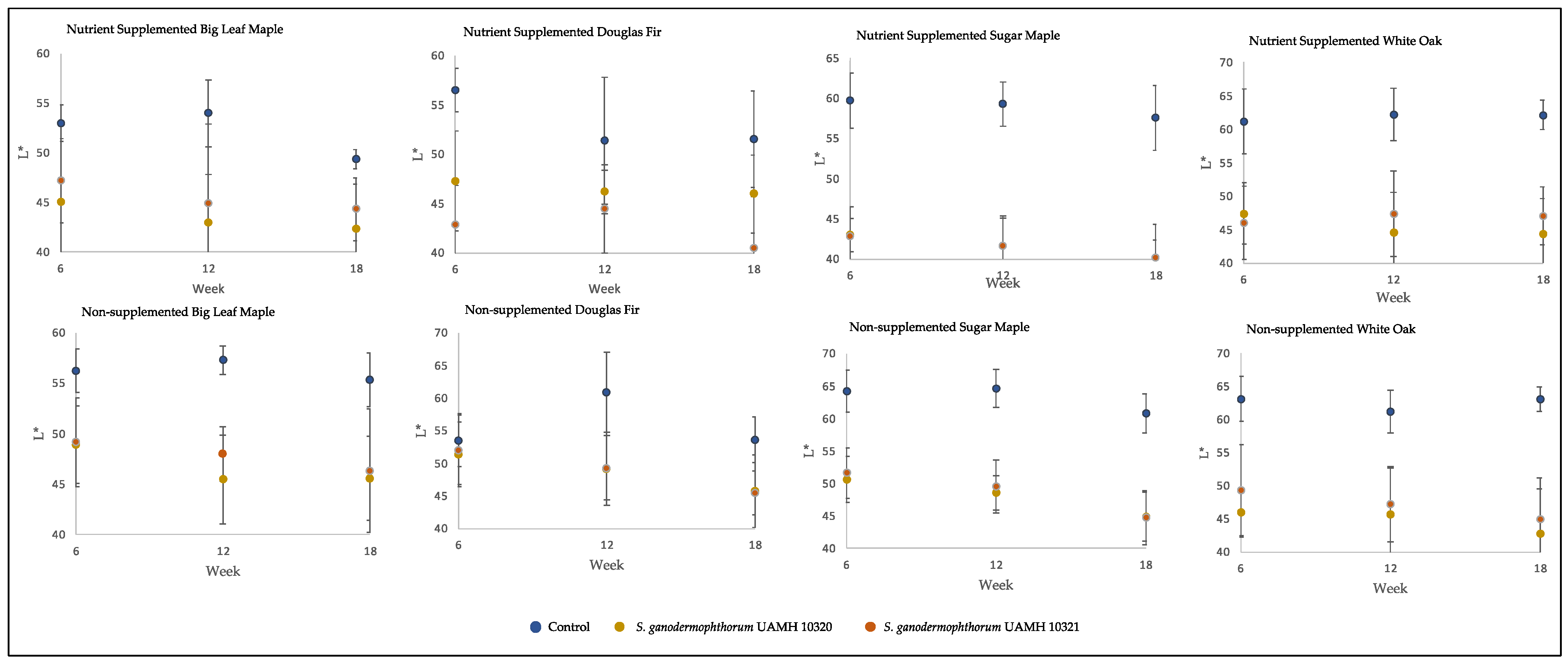
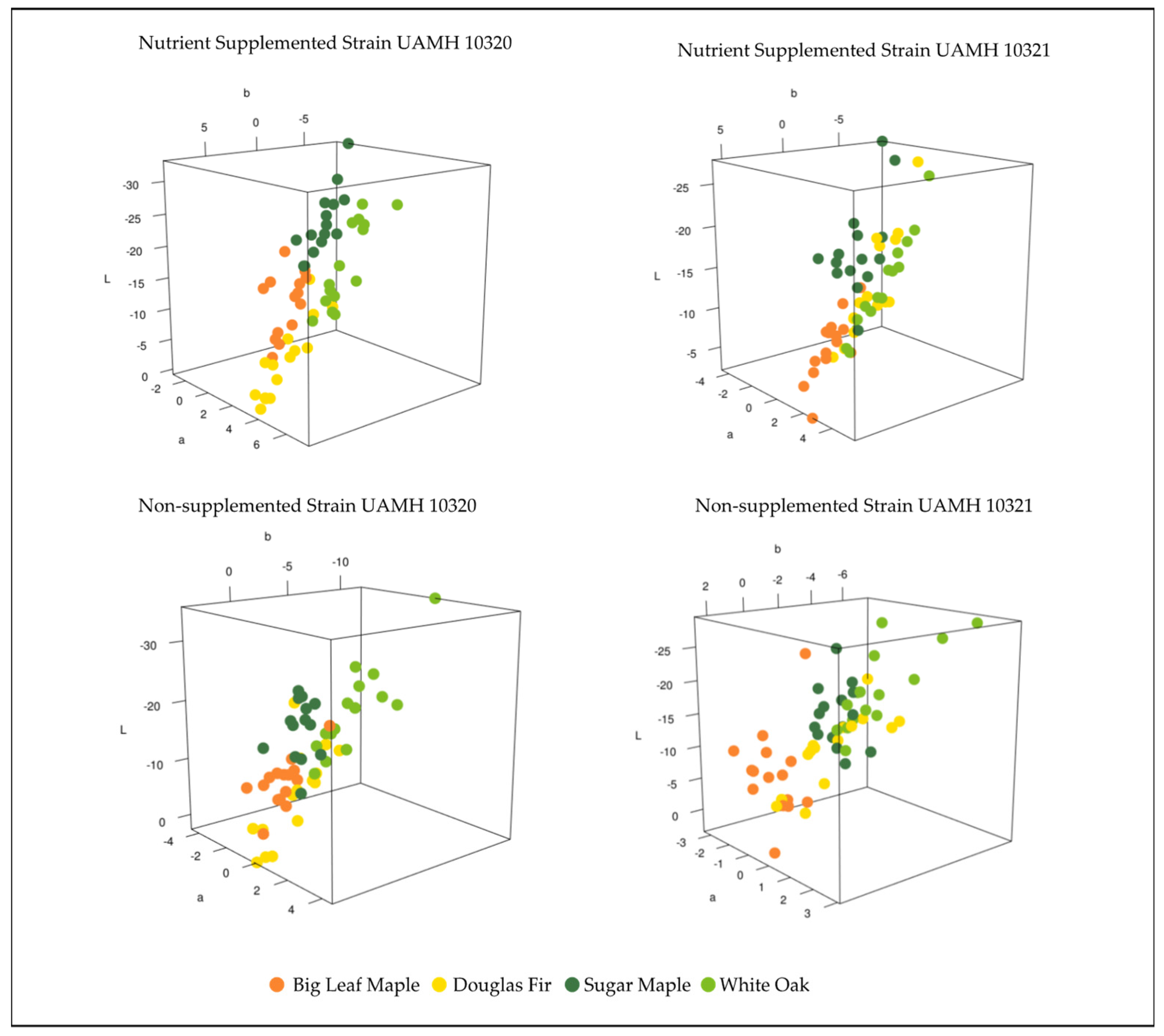
3.1. Block Color Change Analysis
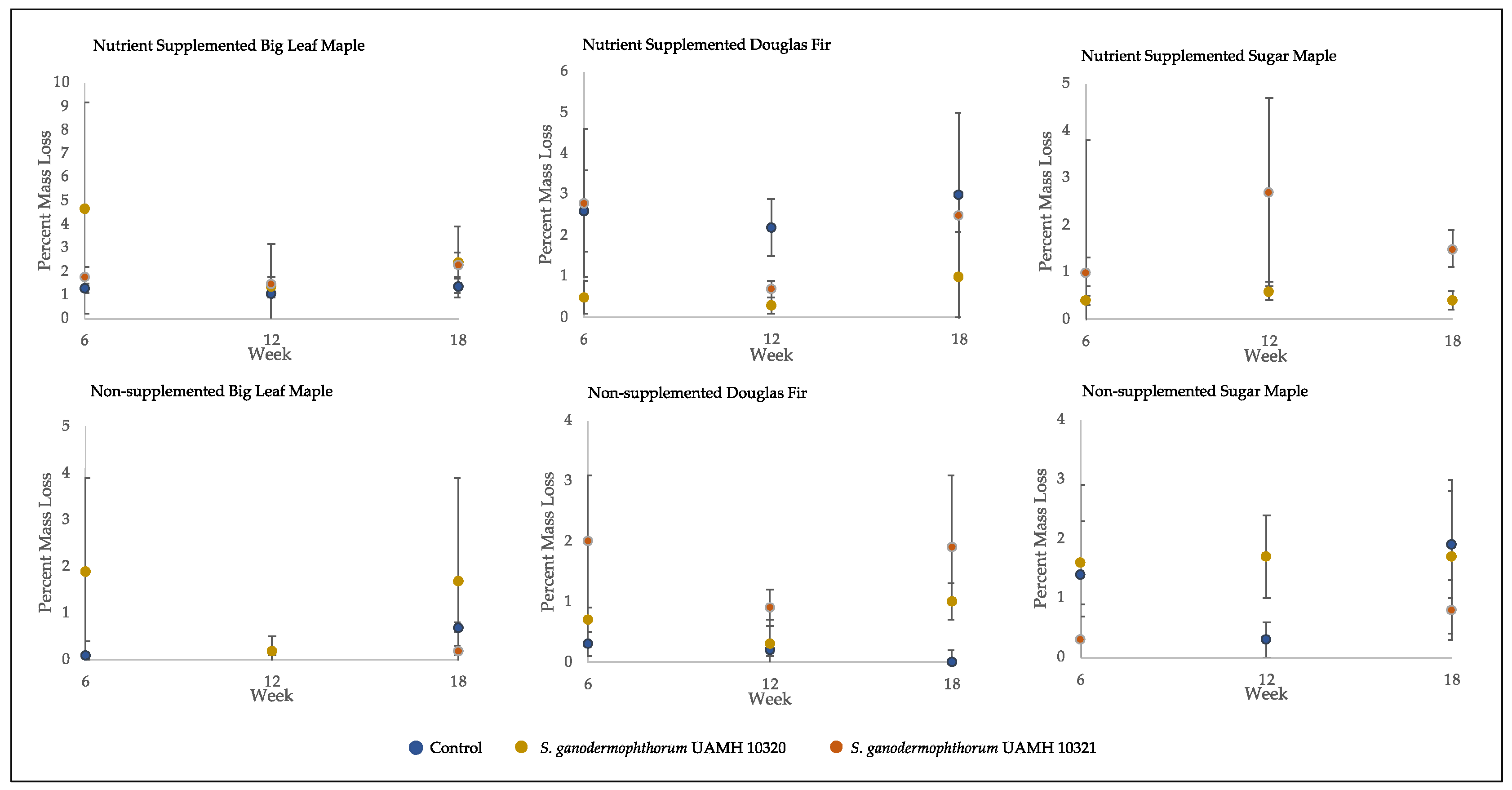
3.2. Assessment of Wood Decay Capabilities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kang, H.; Sigler, L.; Lee, J.; Gibas, C.; Yun, S.; Lee, Y. Xylogone ganodermophthora sp. nov., an ascomycetous pathogen causing yellow rot on cultivated mushroom Ganoderma lucidum in Korea. Mycologia 2010, 102, 1167–1184. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.C.; Hinsch, E.; Weber, G.; Leipus, K.; Cerney, D. Wood colorization through pressure treating: The potential of extracted colorants from spalting fungi as a replacement for woodworkers’ aniline dyes. Materials 2014, 7, 5427–5437. [Google Scholar] [CrossRef]
- Robinson, S.C.; Weber, G.; Hinsch, E.; Vega Gutierrez, S.M.; Pittis, L.; Freitas, S. Utilizing extracted fungal pigments for wood spalting: A comparison of induced fungal pigmentation to fungal dyeing. J. Coat. 2014, 2014, 759073. [Google Scholar] [CrossRef]
- Robinson, S.C.; Vega Gutierrez, S.; Garcia, R.A.C.; Iroume, N.; Vorland, N.R.; McClelland, A.; Huber, M.; Stanton, S. Potential for carrying dyes derived from spalting fungi in natural oils. J. Coat. Technol. Res. 2017, 14, 1107–1113. [Google Scholar] [CrossRef]
- Rui, H.; Robinson, C.S.; Gutierrez, P.V.; Stanton, S. Spalting colorants as dyes for wood stablizers. J. Coat. Technol. Res. 2019, 16, 905–911. [Google Scholar] [CrossRef]
- Weber, G.; Chen, H.-L.; Hinsch, E.; Freitas, S.; Robinson, S. Pigments extracted from the wood-staining fungi Chlorociboria aeruginosa, Scytalidium cuboideum, and S. ganodermophthorum show potential for use as textile dyes. Color. Technol. 2014, 130, 445–452. [Google Scholar] [CrossRef]
- Hinsch, E.M.; Weber, G.; Chen, H.-L.; Robinson, S.C. Colorfastness of extracted wood-staining fungal pigments on fabrics: A new potential for textile dyes. J. Text. Appar. Technol. Manag. 2015, 9. [Google Scholar]
- Hinsch, E.; Robinson, S. Comparing Colorfastness to Light of Wood-Staining Fungal Pigments and Commercial Dyes: An Alternative Light Test Method for Color Fastness. Coatings 2018, 8, 189. [Google Scholar] [CrossRef]
- Robinson, S.C.; Vega Gutierrez, S.M.; Garcia, R.A.C.; Iroume, N.; Vorland, N.R.; Andersen, C.; de Oliveira Xaxa, I.D.; Kramer, O.E.; Huber, M.E. Potential for fungal dyes as colorants in oil and acrylic paints. J. Coat. Technol. Res. 2018, 15, 845–849. [Google Scholar] [CrossRef]
- Harrison, R.; Quinn, A.; Weber, G.; Johnson, B.; Rath, J.; Remcho, V.; Robinson, S.; Ostroverkhova, O. Fungi-derived pigments as sustainable organic (opto)electronic materials. In Organic Photonic Materials and Devices XIX, Proceedings of SPIE OPTO, San Francisco, CA, USA, 28 January–2 February 2017; SPIE: Bellingham, WA, USA, 2017; Volume 10101, p. 101010U. [Google Scholar]
- Daub, M.E.; Herrero, S.; Chung, K.-R. Photoactivated perylenequinone toxins in fungal pathogenesis of plants. FEMS Microbiol. Lett. 2005, 252, 197–206. [Google Scholar] [CrossRef]
- Unagul, P.; Wongsa, P.; Kittakoop, P.; Intamas, S.; Srikitikulchai, P.; Tanticharoen, M. Production of red pigments by the insect pathogenic fungus Cordyceps unilateralis BCC 1869. J. Ind. Microbiol. Biotechnol. 2005, 32, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.H.; Bell, A.A. Melanins and their importance in pathogenic fungi. In Current Topics in Medical Mycology; Springer: New York, NY, USA, 1988; Volume 2, pp. 338–387. [Google Scholar]
- Hosoe, T.; Nozawa, K.; Kawahara, N.; Fukushima, K.; Nishimura, K.; Miyaji, M.; Kawai, K.-I. Isolation of a new potent cytotoxic pigment along with indigotin from the pathogenic basidiomycetous fungus Schizophyllum commune. Mycopathologia 1999, 146, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-J.; Chun, C.-S.; Lee, J.-K.; Kim, H.-K. Occurence and identification of the fungus causing yellow rot on Ganoderma lucidum. Korean J. Mycol. 1998, 26, 31–38. [Google Scholar]
- Kang, H.-J.; Kim, Y.-S.; Kim, T.-I.; Jeong, T.-G.; Han, J.-W.; Nam, S.-Y.; Kim, I.-J. Suppression of powdery mildew using the water extract of Xylogone ganodermophthora and aqueous potassium phosphonate solution on watermelon under greenhouse conditions. Res. Plant Dis. 2015, 21, 309–314. [Google Scholar] [CrossRef]
- Robinson, S.; Hinsch, E.; Weber, G.; Freitas, S. Method of extraction and resolubilisation of pigments from Chlorociboria aeruginosa and Scytalidium cuboideum, two prolific spalting fungi. Color. Technol. 2014, 130, 221–225. [Google Scholar] [CrossRef]
- Worrall, J.T.; Anagnost, S.E.; Zabel, R.A. Comparison of wood decay among diverse lignicolous fungi. Mycologia 1997, 89, 199–219. [Google Scholar] [CrossRef]
- Richter, D.L.; Glaeser, J.A. Wood decay by Chlorociboria aeruginascens (Nyl.) Kanouse (Helotiales, Leotiaceae) and associated basidiomycete fungi. Int. Biodeterior. Biodegrad. 2015, 105, 239–244. [Google Scholar] [CrossRef]
- Uchida, Y.; Kuida, K.; Uchiyama, S.; Udagawa, S. Arthrographic cuboidea isolated as a causal fungus from disease wood logs for cultivation of shiitake mushroom (Lentinus edodes). Trans. Mycol. Soc. Jpn. 1993, 34, 275–281. [Google Scholar]
- Highley, T.L. Laboratory studies on antagonism of Scytalidium lignicola to wood decay fungi. Mater. Org. 1990, 25, 181–192. [Google Scholar]
- Savory, J.G. Breakdown of timber by ascomycetes and fungi imperfecti. Ann. Appl. Biol. 1954, 41, 336–347. [Google Scholar] [CrossRef]
- Nilsson, T. Studies on wood degradation and cellulolytic activity of microfungi. Stud. For. Suec. 1974, 104, 5–40. [Google Scholar]
- Hale, M.D.; Eaton, R.A. The ultrastructure of soft rot fungi. II. Cavity-forming hyphae in wood cell walls. Mycologia 1985, 77, 594–605. [Google Scholar] [CrossRef]
- Worrall, J.J.; Wang, C.J.K. Importance and mobilization of nurtients in soft rot of wood. Can. J. Microbiol. 1991, 37, 864–868. [Google Scholar] [CrossRef]
- Van Court, R.C.; Giesbers, G.; Ostroverkhova, O.; Robinson, C.S. Optimizing xylindein from Chlorociboria spp. for (opto)electronic applications. Processes 2020, 8, 477. [Google Scholar] [CrossRef]
- Robinson, S.; Tudor, D.; Zhang, W.; Ng, S.; Cooper, P. Ability of three yellow pigment producing fungi to colour wood under controlled conditions. Int. Wood Prod. J. 2014, 5, 103–107. [Google Scholar] [CrossRef]
- Standard method of testing wood preservatives by laboratory soil-block culture. In American Wood Preservers Association Book of Standards; AWPA: Granbury, TX, USA, 2003.
- Duncan, C.G. Wood-Attacking Capacities and Physiology of Soft-Rot Fungi; Report No. 2173; Forest Products Laboratory: Madison, WI, USA, 1960. [Google Scholar]
- Vega Gutierrez, S.; Vega Gutierrez, P.; Godinez, A.; Pittis, L.; Huber, M.; Stanton, S.; Robinson, S. Feasibility of coloring bamboo with the application of natural and extracted fungal pigments. Coatings 2016, 6, 37. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Kassambara, A. rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R Package Version 0.7.2. 2023. Available online: https://rpkgs.datanovia.com/rstatix/ (accessed on 1 February 2023).
- Murdoch, D.; Adler, D. rgl: 3D Visualization Using OpenGL. R Package Version 0.110.2. 2022. Available online: https://github.com/dmurdoch/rgl (accessed on 1 February 2023).
- Leightley, L.E. A rapid screening method for determining soft-rot decay ability. Mycologia 1980, 72, 632–637. [Google Scholar] [CrossRef]
- Scheffer, T.C.; Morrell, J.J. Natural Durability of Wood: A Worldwide Checklist of Species; Oregon State University, Forest Research Laboratory: Corvallis, OR, USA, 1998. [Google Scholar]
- McCleary, B.V.; McKie, V.; Draga, A. Measurement of endo-1,4-β-glucanase. Methods Enzymol. 2012, 510, 1–17. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Anagnost, S.E. Light microscopic diagnosis of wood decay. Iawa J. 1998, 19, 141–167. [Google Scholar] [CrossRef]
- Van Court, R.C.; Gutierrez, P.T.V.; Robinson, S.C. Exploration of pigment production by spalting fungi Scytalidium ganodermophthorum and potential for industry. In Proceedings of the IRG51 Scientific Conference on Wood Protection, Bled, Slovenia, 7–11 June 2020. [Google Scholar]
- Keilich, G.; Bailey, P.; Liese, W. Enzymatic degradation of cellulose, cellulose derivatives and hemicelluloses in relation to the fungal decay of wood. Wood Sci. Technol. 1970, 4, 273–283. [Google Scholar] [CrossRef]
- Berg, B. Cellulose degradation and cellulase formation by Phialophora malorum. Arch. Microbiol. 1978, 118, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Green, N. The biochemical basis of wood decay micro-morphology. J. Inst. Wood Sci. 1980, 8, 221–228. [Google Scholar]
- Hale, M.; Eaton, R. Soft rot cavity widening: A kinetic approach. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1986, 227, 217–226. [Google Scholar]
- Eslyn, W.; Kirk, T.; Effland, M. Changes in the chemical composition of wood caused by six soft-rot fungi. Phytopathology 1975, 65, 473–476. [Google Scholar] [CrossRef]
- Hori, C.; Gaskell, J.; Igarashi, K.; Kersten, P.; Mozuch, M.; Samejima, M.; Cullen, D. Temporal alterations in the secretome of the selective ligninolytic fungus Ceriporiopsis subvermispora during growth on aspen wood reveal this organism’s strategy for degrading lignocellulose. Appl. Environ. Microbiol. 2014, 80, 2062–2070. [Google Scholar] [CrossRef]
- Ahmad, R.; Lim, C.K.; Marzuki, N.F.; Goh, Y.-K.; Azizan, K.A.; Goh, Y.K.; Goh, K.J.; Ramzi, A.B.; Baharum, S.N. Metabolic Profile of Scytalidium parasiticum-Ganoderma boninense Co-Cultures Revealed the Alkaloids, Flavonoids and Fatty Acids that Contribute to Anti-Ganoderma Activity. Molecules 2020, 25, 5965. [Google Scholar] [CrossRef]
- Goh, Y.K.; Marzuki, N.F.; Goh, T.K.; Tan, S.Y.; Goh, Y.K.; Goh, K.J. Mycoparasitic Scytalidium parasiticum as a potential biocontrol agent against Ganoderma boninense basal stem rot in oil palm. Biocontrol Sci. Technol. 2016, 26, 1352–1365. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Kumar, A.; de Los Santos-Villalobos, S.; Parra-Cota, F.I.; Orozco-Mosqueda, M.D.C.; Fadiji, A.E.; Hyder, S.; Babalola, O.O.; Santoyo, G. Trichoderma Species: Our Best Fungal Allies in the Biocontrol of Plant Diseases—A Review. Plants 2023, 12, 432. [Google Scholar] [CrossRef]
- Silva, F.D.A.; Liotti, R.G.; Boleti, A.P.D.A.; Reis, É.D.M.; Passos, M.B.S.; dos Santos, E.L.; Sampaio, O.M.; Januário, A.H.; Branco, C.L.B.; Silva, G.F.D.; et al. Diversity of cultivable fungal endophytes in Paullinia cupana (Mart.) Ducke and bioactivity of their secondary metabolites. PLoS ONE 2018, 13, e0195874. [Google Scholar] [CrossRef]
- Moore, M.K. The Infection of Human Skin and Nail by Scytalidium Species. In Current Topics in Medical Mycology; Borgers, M., Hay, R., Rinaldi, M.G., Eds.; Springer: New York, NY, USA, 1992; pp. 1–42. [Google Scholar] [CrossRef]
- Machouart, M.; Menir, P.; Helenon, R.; Quist, D.; Desbois, N. Scytalidium and scytalidiosis: What’s new in 2012? J. Mycol. Méd. 2013, 23, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Oren, Y.; Sadowsky, A.; Gefen, D.; Solel, Z.; Kimchy, M. Scytalidium Wilt of Citrus. Eur. J. Plant Pathol. 2001, 107, 467–470. [Google Scholar] [CrossRef]
- de Medeiros, E.V.; Notaro, K.D.A.; de Barros, J.A.; Duda, G.P.; Moraes, M.D.C.H.D.S.; Ambrósio, M.M.D.Q.; Negreiros, A.M.P.; Sales Júnior, R. Soils from intercropped fields have a higher capacity to suppress black root rot in cassava, caused by Scytalidium lignicola. J. Phytopathol. 2019, 167, 209–217. [Google Scholar] [CrossRef]
- Almurshidi, B.H. Toxicity of Fungal Pigments from Chlorociboria spp. and Scytalidium spp. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2015. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Court, R.C.; Rogers, L.; Robinson, S.C.; Presley, G. Wood Coloration and Decay Capabilities of Mycoparasite Scytalidium ganodermophthorum. J. Fungi 2023, 9, 738. https://doi.org/10.3390/jof9070738
Van Court RC, Rogers L, Robinson SC, Presley G. Wood Coloration and Decay Capabilities of Mycoparasite Scytalidium ganodermophthorum. Journal of Fungi. 2023; 9(7):738. https://doi.org/10.3390/jof9070738
Chicago/Turabian StyleVan Court, Ray C., Leon Rogers, Seri C. Robinson, and Gerald Presley. 2023. "Wood Coloration and Decay Capabilities of Mycoparasite Scytalidium ganodermophthorum" Journal of Fungi 9, no. 7: 738. https://doi.org/10.3390/jof9070738
APA StyleVan Court, R. C., Rogers, L., Robinson, S. C., & Presley, G. (2023). Wood Coloration and Decay Capabilities of Mycoparasite Scytalidium ganodermophthorum. Journal of Fungi, 9(7), 738. https://doi.org/10.3390/jof9070738