


Re-Examination of the Holotype of Ganoderma sichuanense (Ganodermataceae, Polyporales) and a Clarification of the Identity of Chinese Cultivated Lingzhi



Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. DNA Samples
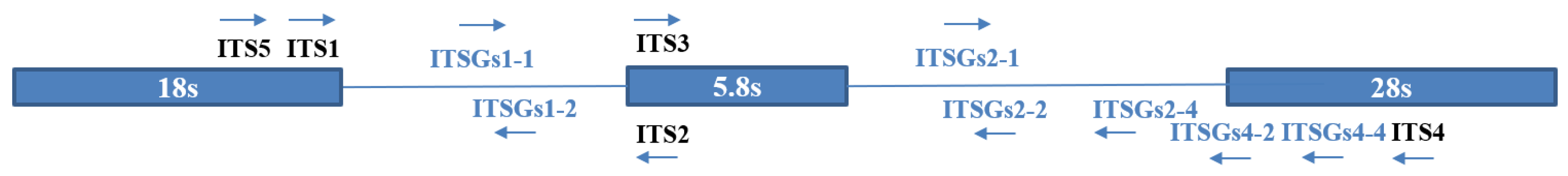
2.3. Specific Primer Design, Amplification, and Sequencing
- Directed PCR: the following 4 primer combinations were used: ITS1/ITSGs1-2, ITSGs1-1/ITS2, ITS3/ITSGs2-2, and ITSGs2-1/ITS4.
- Nested PCR: ITS5/ITS4 was followed by 6 different primer pairs as internal primers (Table 2).
2.4. Genome Sequencing
2.5. Phylogenetic Analyses
3. Results
3.1. Results of PCR, Sequencing Using Different Primers
3.2. Results of Genome Sequencing and Assembly
3.3. Results of Phylogenetic Analyses
3.4. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karsten, P.A. Enumeratio boletinearum et polyporearum fennicarum, systemate novo dispositarum. Rev. Mycol. 1881, 3, 16–19. [Google Scholar]
- Curtis, W. Flora Londinensis: Or Plates and Descriptions of Such Plants as Grow Wild in the Environs of London; Curtis, W.: London, UK, 1781; p. 530. [Google Scholar]
- Yuan, Y.; Wang, Y.J.; Sun, G.P.; Wang, Y.R.; Cao, L.J.; Shen, Y.M.; Yuan, B.; Han, D.; Huang, L.Q. Archaeological evidence suggests earlier use of Ganoderma in Neolithic China. Chin. Sci. Bull. 2018, 63, 1180–1188. (In Chinese) [Google Scholar]
- Pegler, D.N. Useful fungi of the world: The Lingzhi—The mushroom of immortality. Mycologist 2002, 16, 100–101. [Google Scholar] [CrossRef]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979; p. 1527. [Google Scholar]
- Pegler, D.N.; Yao, Y.J. Oriental species of Ganoderma section Ganoderma. In Botany and Mycology for the Next Millenium: Collection of Scientific Articles Devoted to the 70th Anniversary of Academician Sytnik, K.M; Wasser, S.P., Ed.; National Academy of Sciences of Ukraine: Kiev, Ukraine, 1996; pp. 336–347. [Google Scholar]
- Moncalvo, J.M.; Wang, H.F.; Hseu, R.S. Gene phylogeny of the Ganoderma lucidum complex based on ribosomal DNA sequences. Comparison with traditional taxonomic characters. Mycol. Res. 1995, 99, 1489–1499. [Google Scholar] [CrossRef]
- Moncalvo, J.M.; Wang, H.F.; Wang, H.H.; Hseu, R.S. The use of ribosomal DNA sequence data for species identification and phylogeny in the Ganodermataceae. In Proceedings of the Contributed Symposium 59A, B, 5th International Mycological Congress, Vancouver, BC, Canada, 14–21 August 1994. [Google Scholar]
- Smith, B.J.; Sivasithamparam, K. Internal transcribed spacer ribosomal DNA sequence of five species of Ganoderma from Australia. Mycol. Res. 2000, 104, 943–951. [Google Scholar] [CrossRef]
- Hong, S.G.; Jung, H.S. Phylogenetic analysis of Ganoderma based on nearly complete mitochondrial small-subunit ribosomal DNA sequences. Mycologia 2004, 96, 742–755. [Google Scholar] [CrossRef]
- Wang, D.M.; Wu, S.H.; Su, C.H.; Peng, J.T.; Shih, Y.H.; Chen, L.Z. Ganoderma multipileum, the correct name for ‘G. lucidum’ in tropical Asia. Bot. Stud. 2009, 50, 451–458. [Google Scholar]
- Yu, Y.N.; Shen, M.Z. The history of ling-zhi (Ganoderma spp.) cultivation. In Proceedings of the 3rd National Congress of Mycological Society of China and the 6th National Symposium on Mycology, Beijing, China, 22–25 September 2003. [Google Scholar]
- Jong, S.C.; Birmingham, J.M. Medicinal benefits of the mushroom Ganoderma. Adv. Appl. Microbiol. 1992, 37, 101–134. [Google Scholar] [PubMed]
- Wasser, S.P.; Weis, A.L. Medicinal properties of substances occurring in higher basidiomycetes mushrooms: Current perspectives (Review). Int. J. Med. Mushrooms 1999, 1, 31–62. [Google Scholar] [CrossRef]
- Tan, B.K.H.; Vanitha, J. Immunomodulatory and antimicrobial effects of some traditional Chinese medicinal herbs: A review. Curr. Med. Chem. 2004, 11, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R.M. Ganoderma—A therapeutic fungal biofactory. Phytochemistry 2006, 67, 1985–2001. [Google Scholar] [CrossRef]
- Rathore, H.; Prasad, S.; Sharma, S. Mushroom nutraceuticals for improved nutrition and better human health: A review. PharmaNutrition 2017, 5, 35–46. [Google Scholar] [CrossRef]
- Wang, X.C.; Xi, R.J.; Li, Y.; Wang, D.M.; Yao, Y.J. The species identity of the widely cultivated Ganoderma, ‘G. lucidum’ (Ling-zhi), in China. PLoS ONE 2012, 7, e40857. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.D.; Xu, L.W.; Zhang, X.Q. Taxonomic studies on the family Ganodermataceae of China II. Acta Mycol. Sin. 1983, 2, 159–167. (In Chinese) [Google Scholar]
- Cao, Y.; Wu, S.H.; Dai, Y.C. Species clarifcation of the prize medicinal Ganoderma mushroom “lingzhi”. Fungal. Divers. 2012, 56, 49–62. [Google Scholar] [CrossRef]
- Yao, Y.J.; Wang, X.C.; Wang, B. Epitypifcation of Ganoderma sichuanense J.D. Zhao & X.Q. Zhang (Ganodermataceae). Taxon 2013, 62, 1025–1031. [Google Scholar]
- Zhou, L.W.; Cao, Y.; Wu, S.H.; Vlasák, J.; Li, D.E.; Li, M.J.; Dai, Y.C. Global diversity of the Ganoderma lucidum complex (Ganodermataceae, Polyporales) inferred from morphology and multilocus phylogeny. Phytochemistry 2015, 114, 7–15. [Google Scholar] [CrossRef]
- Dai, Y.C.; Zhou, L.W.; Hattori, T.; Cao, Y.; Stalpers, J.A.; Ryvarden, L.; Buchanan, P.; Oberwinkler, F.; Hallenberg, N.; Liu, P.G.; et al. Ganoderma lingzhi (Polyporales, Basidiomycota): The scientifc binomial for the widely cultivated medicinal fungus lingzhi. Mycol. Prog. 2017, 16, 1051–1055. [Google Scholar] [CrossRef]
- Cui, B.K.; Wu, S.H. The scientific name of the widely cultivated Ganoderma species. Mycosystema 2020, 39, 7–12. (In Chinese) [Google Scholar]
- Yao, Y.J.; Li, Y.; Du, Z.; Wang, K.; Wang, X.C.; Spooner, B.M. On the typification of Ganoderma sichuanense (Agaricomycetes)the widely cultivated lingzhi medicinal mushroom. Int. J. Med. Mushrooms 2020, 22, 45–54. [Google Scholar] [CrossRef]
- Jiang, Y.; Yao, Y.J. ITS sequence analysis and ascomatal development of Pseudogymnoascus roseus. Mycotaxon 2005, 94, 55–73. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide. Methods. Appl. 1990, 18, 315–322. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Li, D.H.; Liu, C.M.; Luo, R.B.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Yang, Z.L.; Feng, B. What is the Chinese “lingzhi”?—A taxonomic mini-review. Mycology 2013, 4, 1–4. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code). In Proceedings of the 19th International Botanical Congress, Shenzhen, China, 23–29 July 2017. [Google Scholar]
- Hillis, D.M.; Moritz, C.; Porter, C.A.; Baker, R.J. Evidence for biased gene conversion in concerted evolution of ribosomal DNA. Science 1991, 251, 308. [Google Scholar] [CrossRef]
- Coen, E.S.; Dover, G.A. Unequal exchanges and the coevolution of X and Y rDNA arrays in Drosophila melanogaster. Cell 1983, 33, 849–855. [Google Scholar] [CrossRef]
- Chen, J.; Parra, L.A.; De, K.A.; Khalid, A.N.; Qasim, T.; Ashraf, A.; Bahkali, A.H.; Hyde, K.D.; Zhao, R.L.; Callac, P. Inter-and intra-specific diversity in Agaricus endoxanthus and allied species reveals a new taxon, A. punjabensis. Phytotaxa 2016, 252, 1–16. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Lindner, D.L.; Banik, M.T. Intragenomic variation in the ITS rDNA region obscures phylogenetic relationships and inflates estimates of operational taxonomic units in genus Laetiporus. Mycologia 2011, 103, 731–740. [Google Scholar] [CrossRef]
- Li, Y.; Jiao, L.; Yao, Y.J. Non-concerted ITS evolution in fungi, as revealed from the important medicinal fungus Ophiocordyceps sinensis. Mol. Phylogenet. Evol. 2013, 68, 373–379. [Google Scholar] [CrossRef]
- Hijri, M.; Hosny, M.; van Tuinen, D.; Dulieu, H. Intraspecific ITS polymorphism in Scutellospora castanea (Glomales, Zygomycota) is structured within multinucleate spores. Fungal. Genet. Biol. 1999, 26, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Fell, J.W.; Scorzetti, G.; Statzell, T.A.; Boundy, M.K. Molecular diversity and intragenomic variability in the yeast genus Xanthophyllomyces: The origin of Phaffia rhodozyma? FEMS Yeast Res. 2007, 7, 1399–1408. [Google Scholar] [CrossRef]
- Wang, D.M.; Yao, Y.J. Intrastrain internal transcribed spacer heterogeneity in Ganoderma species. Can. J. Microbiol. 2005, 51, 113–121. [Google Scholar] [CrossRef]
- Sun, Y.F.; Xing, J.H.; He, X.L.; Wu, D.M.; Song, C.G.; Liu, S.; Vlasák, J.; Gates, G.; Gibertoni, T.B.; Cui, B.K. Species diversity, systematic revision and molecular phylogeny of Ganodermataceae (Polyporales, Basidiomycota) with an emphasis on Chinese collections. Stud. Mycol. 2022, 101, 287–415. [Google Scholar] [CrossRef]
- Cabarroi-Hernández, M.; Villalobos-Arámbula, A.R.; Torres-Torres, M.G.; Decock, C.; GuzmánDávalos, L. The Ganoderma weberianum-resinaceum lineage: Multilocus phylogenetic analysis and morphology confirm G. mexicanum and G. parvulum in the Neotropics. MycoKeys 2019, 59, 95–131. [Google Scholar] [CrossRef]
- Hapuarachchi, K.K. Ganodermataceae (Polyporales): Diversity in Greater Mekong Subregion countries (China, Laos, Myanmar, Thailand and Vietnam). Mycosphere 2019, 10, 221–309. [Google Scholar] [CrossRef]
- Xing, J.H.; Sun, Y.F.; Han, Y.L.; Cui, B.K.; Dai, Y.C. Morphological and molecular identification of two new Ganoderma species on Casuarina equisetifolia from China. MycoKeys 2018, 34, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.H.; Song, J.; Decock, C.; Cui, B.K. Morphological characters and phylogenetic analysis reveal a new species within the Ganoderma lucidum complex from South Africa. Phytotaxa 2016, 266, 115–124. [Google Scholar] [CrossRef]
- Luangharn, T.; Karunarathna, S.C.; Mortimer, P.E.; Hyde, K.D.; Xu, J.C. Morphology, phylogeny and culture characteristics of Ganoderma gibbosum collected from Kunming, Yunnan Province, China. Phyton Int. J. Exp. Bot. 2020, 89, 743–764. [Google Scholar] [CrossRef]
- Luangharn, T.; Karunarathna, S.C.; Dutta, A.K.; Paloi, S.; Mortimer, P.E. Ganoderma (Ganodermataceae, Basidiomycota) species from the Greater Mekong Subregion. J. Fungi. 2021, 7, 819. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Han, X.; Luo, Z.L.; Li, E.X.; Tang, S.M.; Luo, H.M.; Niu, K.Y.; Sun, X.J.; Li, S.H. Species diversity of Ganoderma (Ganodermataceae, Polyporales) with three new species and a key to Ganoderma in Yunnan Province, China. Front. Microbiol. 2022, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence 5′-3′ | Tm (°C) | GC% | Length | Location |
|---|---|---|---|---|---|
| ITSGs1-1 | TAC TGT GGG CTT CAG ATT GC | 56.0 | 55.0 | 20 | ITS1 |
| ITSGs1-2 | GTG CCT CGC AAT CTG AAG C | 58.9 | 57.9 | 19 | ITS1 |
| ITSGs2-1 | TTA TCG GTC GGC TCC TCT TA | 57.7 | 50.0 | 20 | ITS2 |
| ITSGs2-2 | AAG AGG AGC CGA CCG ATA AC | 58.4 | 55.0 | 20 | ITS2 |
| ITSGs2-4 | AGC TGT CTT ATA AGA CGG T | 51.6 | 36.4 | 22 | ITS2 |
| ITSGs4-2 | CAG GTC ATA AAG CTG TCT TAT | 48.4 | 38.1 | 21 | ITS2 & 28s |
| ITSGs4-4 | GTC CTA CCT GAT TTG AGG TCA | 54 | 47.6 | 21 | 28s |
| Primers for the First Round PCR | Primers for the Second Round PCR |
|---|---|
| ITS5/ITS4 | ITS1/ITS4 |
| ITS1/ITSGs2-4 | |
| ITS1/ITSGs4-2 | |
| ITS1/ITSGs4-4 | |
| ITSGs1-1/ITSGs2-4 | |
| ITSGs1-1/ITSGs4-4 |
| Species | Voucher/Strain | Origin | Accession Number (ITS) |
|---|---|---|---|
| Ganoderma adspersum (Schulzer) Donk | SFC 20141001-16 | Korea | KY364251 |
| G. adspersum | SFC 20141001-22 | Korea | KY364252 |
| G. angustisporum J.H. Xing, B.K. Cui & Y.C. Dai | Cui 13817 (holotype) | China | MG279170 |
| G. angustisporum | Cui 14578 | China | MG279171 |
| G. applanatum (Pers.) Pat. | SFC 20150930-02 | Korea | KY364258 |
| G. applanatum | XC 14080601 | China | MK345426 |
| G. aridicola J.H. Xing & B.K. Cui | Dai 12588 (holotype) | South Africa | KU572491 |
| G. australe (Fr.) Pat. | GDGM 25344 | China | JX195198 |
| G. australe | GACP 14061914 | China | MK345428 |
| G. austroafricanum M.P.A. Coetzee, M.J. Wingf., Marinc. & Blanchette | CBS 138724 (ex-type) | South Africa | KM507324 |
| G. bambusicola Sheng H. Wu, C.L. Chern & T. Hatt. | Wu 1207-151 (holotype) | China | MN957781 |
| G. bambusicola | Wu 1207-153 (paratype) | China | MN957783 |
| G. boninense Pat. | WD 2028 | Japan | KJ143905 |
| G. boninense | WD 2085 | Japan | KJ143906 |
| G. calidophilum J.D. Zhao, L.W. Hsu & X.Q. Zhang | MFLU 19-2174 | China | MN398337 |
| G. calidophilum | H36 | China | MW750241 |
| G. carnosum Pat. | JV 8709/8 | Czech R | KU572493 |
| G. carnosum | MJ 21/08 | Czech R | KU572492 |
| G. carocalcareum Douanla-Meli | DMC 322 (holotype) | Cameroon | EU089969 |
| G. carocalcareum | DMC 513 | Cameroon | EU089970 |
| G. casuarinicola J.H. Xing, B.K. Cui & Y.C. Dai | Dai 16336 (holotype) | China | MG279173 |
| G. casuarinicola | HKAS 104639 | Thailand | MK817650 |
| G. curtisii (Berk.) Murrill | CBS 100131 | USA | JQ781848 |
| G. curtisii | CBS 100132 | USA | JQ781849 |
| G. destructans M.P.A. Coetzee, Marinc. & M.J.Wingf. | CMW 43670 (ex-type) | South Africa | KR183856 |
| G. destructans | CMW 43671 | South Africa | KR183857 |
| G. dianzhongense J. He, H.Y. Su & S.H. Li | L4331 (holotype) | China | MW750237 |
| G. dianzhongense | L4230 | China | MW750236 |
| G. dunense Tchotet, Rajchenb. & Jol. Roux | CMW 42157 (holotype) | South Africa | MG020255 |
| G. dunense | CMW 42150 | South Africa | MG020249 |
| G. ecuadoriense A. Salazar, C.W. Barnes & Ordoñez | ASL 799 (holotype) | Ecuador | KU128524 |
| G. ecuadoriense | PMC 126 | Ecuador | KU128525 |
| G. eickeri Tchotet, M.P.A. Coetzee, Rajchenb. & Jol. Roux | CMW 49692 (holotype) | South Africa | MH571690 |
| G. eickeri | CMW 50325 | South Africa | MH571689 |
| G. ellipsoideum Hapuar., T.C. Wen & K.D. Hyde | GACP 1408966 (holotype) | China | MH106867 |
| G. ellipsoideum | GACP 14081215 (paratype) | China | MH106886 |
| G. enigmaticum M.P.A. Coetzee, Marinc. & M.J.Wingf. | CMW 43669 (ex-type) | South Africa | KR183855 |
| G. enigmaticum | Dai 15970 | South Africa | KU572486 |
| G. esculentum J. He & S.H. Li | L4935 (holotype) | China | MW750242 |
| G. esculentum | L4946 | China | MW750243 |
| G. flexipes Pat. | VT 17102301 | Vietnam | MK345430 |
| G. flexipes | Wei 5200 | China | JN383978 |
| G. gibbosum (Cooke) Pat. | SFC 20150630-23 | Korea | KY364264 |
| G. gibbosum | GZ 14070501 | China | MK345432 |
| G. heohnelianum Bres. | Yuan 6337 | China | MG279160 |
| G. heohnelianum | Dai 11995 | China | KU219988 |
| G. hochiminhense Karunarathna, Mortimer, Huyen & Luangharn | MFLU 19-2224 (holotype) | Vietnam | MN398324 |
| G. hochiminhense | MFLU 19-2225 (paratype) | Vietnam | MN396662 |
| G. knysnamense Tchotet, M.P.A. Coetzee, Rajchenb. & Jol. Roux | CMW 47755 (ex-type) | South Africa | MH571681 |
| G. knysnamense | CMW 47756 | South Africa | MH571684 |
| G. leucocontextum T.H. Li, W.Q. Deng, Sheng H. Wu, Dong M. Wang & H.P. Hu | GDGM 40200 (holotype) | China | KF011548 |
| G. leucocontextum | GDGM 44303(paratype) | China | KJ027607 |
| G. lingzhi Sheng H. Wu, Y. Cao & Y.C. Dai | Wu 1006-38 (holotype) | China | JQ781858 |
| G. lingzhi | Cui 4018 | China | JQ781856 |
| G. lingzhi | Cui 10165 | China | JQ781857 |
| G. lingzhi | Cui 9164 | China | JQ781859 |
| G. lingzhi | Dai 10631 | China | JQ781860 |
| G. lingzhi | Dai 12438 | China | JQ781861 |
| G. lingzhi | Dai 12479 | China | JQ781864 |
| G. lingzhi | IFP 01021 | China | JQ781865 |
| G. lingzhi | Dai 12443 | China | JQ781866 |
| G. lingzhi | Dai 12374 | China | JQ781867 |
| G. lingzhi | Dai 3583 | China | JQ781868 |
| G. lingzhi | Dai 12441 | China | JQ781869 |
| G. lingzhi | Dai 12426 | China | JQ781870 |
| G. lingzhi | Dai 12425 | China | JQ781871 |
| G. lingzhi | Dai 12447 | China | JQ781872 |
| G. lingzhi | Dai 12449 | China | JQ781873 |
| G. lingzhi | Cui 6982 | China | JQ781862 |
| G. lingzhi | Dai 12573 | China | JQ781855 |
| G. lingzhi | Li245 | China | JQ781863 |
| G. lingzhi | LPDR 18011910 | Laos | MK345437 |
| G. lingzhi | LPDR 18011911 | Laos | MK345438 |
| G. lobatum (Cooke) G.F. Atk. | JV1212/10J | USA | KF605676 |
| G. lobatum | JV0409/13J | USA | KF605675 |
| G. lucidum (Curtis) P. Karst. | HMAS 86597 | UK | AY884176 |
| G. lucidum | G1T 099 | Italy | AM269773 |
| G. martinicense Welti & Courtec | LIP SW-Mart08-44 | France | KF963257 |
| G. martinicense | LIP SW-Mart08-55 | France | KF963256 |
| G. mbrekobenum E.C. Otto, Blanchette, Held, C.W. Barnes & Obodai | UMN7-3 GHA (holotype) | Ghana | KX000896 |
| G. mbrekobenum | UMN7-4 GHA (paratype) | Ghana | KX000898 |
| G. mexicanum Pat. | MUCL 49453 | Martinique | MK531811 |
| G. mexicanum | MUCL 55832 | Martinique | MK531815 |
| G. mizoramense Zothanz., Blanchette, Held & C.W. Barnes | UMN-MZ4 (holotype) | India | KY643750 |
| G. mizoramense | UMN-MZ5 | India | KY643751 |
| G. multipileum Ding Hou | CWN 04670 | China | KJ143913 |
| G. multipileum | Dai 9447 | China | KJ143914 |
| G. multiplicatum (Mont.) Pat. | Dai 13122 | China | KU572488 |
| G. multiplicatum | MN 14091108 | Myanmar | MK345440 |
| G. mutabile Y. Cao & H.S. Yuan | Yuan 2289 (holotype) | China | JN383977 |
| G. mutabile | CLZhao 982 | China | MG231527 |
| G. myanmarense Karunarathna, Mortimer & Luangharn | MFLU 19-2167 ((holotype) | Myanmar | MN396330 |
| G. myanmarense | MFLU 19-2211 (paratype) | Myanmar | MN396329 |
| G. nasalanense Hapuar., Pheng., & K.D. Hyde. | LPDR 17060211 (holotype) | Laos | MK345441 |
| G. nasalanense | LPDR 17060212 (paratype) | Laos | MK345442 |
| G. neojaponicum Imazeki | FFPRI WD-1285 | Japan | MN957784 |
| G. neojaponicum | FFPRI WD-1532 | Japan | MN957785 |
| G. orbiforme (Fr.) Ryvarden | JFL 14081202 | China | MK345445 |
| G. orbiforme | GACP 14061414 | Laos | MK345446 |
| G. oregonense Murrill | CBS 265.88 | USA | JQ781875 |
| G. oregonense | CBS 266.88 | USA | JQ781876 |
| G. parvulum Murrill | MUCL 47096 | Cuba | MK554783 |
| G. parvulum | MUCL 52655 | French Guiana | MK554770 |
| G. philippii (Bres. & Henn. ex Sacc.) Bres. | E7098 | Malaysia | AJ536662 |
| G. philippii | E7425 | Malaysia | AJ608713 |
| G. podocarpense J.A. Flores, C.W. Barnes & Ordoñez | QCAM 6422 (holotype) | Ecuador | MF796661 |
| G. resinaceum Boud. | BCRC 36147 | Netherlands | KJ143916 |
| G. resinaceum | BR 4150 | France | KJ143915 |
| G. ryvardenii Tonjock & Mih | HKAS 58053 (holotype) | Cameroon | HM138671 |
| G. ryvardenii | GanoTK32 | Cameroon | JN105698 |
| G. sandunense Hapuar., T.C. Wen & K.D. Hyde. | SA 18012501 (holotype) | China | MK345450 |
| G. sandunense | SA 18012502 | China | MK345451 |
| G. sessile Murrill | JV 1209/27 | USA | KF605630 |
| G. sessile | 165MO | USA | MG654312 |
| G. shandongense J.D. Zhao & L.W. Xu | Dai 15785 | China | MG279190 |
| G. shandongense | Dai 15787 | China | MG279191 |
| G. shanxiense L. Fan & H. Liu | BJTC FM423 (holotype) | China | MK764268 |
| G. shanxiense | HSA 539 (paratype) | China | MK764269 |
| G. sichuanense J.D. Zhao & X.Q. Zhang | HMAS 42798-2 (holotype) | China | OP805615 |
| G. sichuanense | HMAS 42798-3 (holotype) | China | OP805616 |
| G. sichuanense | HMAS 42798-4 (holotype) | China | OP805617 |
| G. sichuanense | HMAS 42798-5 (holotype) | China | OP805618 |
| G. sichuanense | HMAS 42798-6 (holotype) | China | OP805619 |
| G. sichuanense | HMAS 42798-8 (holotype) | China | OP805620 |
| G. sichuanense | HMAS 42798-19 (holotype) | China | OP805621 |
| G. sichuanense | HMAS 42798-23 (holotype) | China | OP805622 |
| G. sichuanense | HMAS 42798-d (holotype) | China | OP805623 |
| G. sichuanense | HMAS 244431-1 | China | OP805624 |
| G. sichuanense | HMAS 244431-2 | China | OP805625 |
| G. sichuanense | HMAS 244431-3 | China | OP805626 |
| G. sichuanense | HMAS 244431-4 | China | OP805627 |
| G. sichuanense | HMAS 25103 | China | OP805628 |
| G. sichuanense | HMAS 252081 (epitype) | China | KC662402 |
| G. sichuanense | HMAS 25066 | China | JN197275 |
| G. sichuanense | HMAS 25067 | China | JN197276 |
| G. sichuanense | HMAS 42605 | China | JN197277 |
| G. sichuanense | HMAS 42745 | China | JN197278 |
| G. sichuanense | HMAS 47337 | China | JN197279 |
| G. sichuanense | HMAS 59482 | China | JN197280 |
| G. sichuanense | HMAS 60537 | China | JN197281 |
| G. sichuanense | HMAS 62503 | China | JF915405 |
| G. sichuanense | HMAS 76566 | China | JF915406 |
| G. sichuanense | HMAS 99391 | China | JF915407 |
| G. sichuanense | HMAS 130131 | China | JF915408 |
| G. sichuanense | HMAS 240175 | China | JF915393 |
| G. sichuanense | HMAS 240176 | China | JF915394 |
| G. sichuanense | HMAS 240177 | China | JF915395 |
| G. sichuanense | HMAS 240178 | China | JF915396 |
| G. sichuanense | HMAS 240187 | China | JF915397 |
| G. sichuanense | HMAS 250672 | China | JF915398 |
| G. sichuanense | HMAS 250677 | China | JF915399 |
| G. sichuanense | HMAS 251145 | China | JF915400 |
| G. sichuanense | HMAS 251146 | China | JF915401 |
| G. sichuanense | HMAS 251147 | China | JF915402 |
| G. sichuanense | HMAS 251148 | China | JF915403 |
| G. sichuanense | HMAS130128 | China | JF915404 |
| G. sichuanense | CGMCC 5.75 | China | JN197282 |
| G. sichuanense | CGMCC 5.425 | China | JN197283 |
| G. sichuanense | CGMCC 5.533 | China | JN197284 |
| G. sichuanense | Cui 7691 | China | JQ781878 |
| G. sichuanense | HMAS 42798 (holotype) | China | JQ781877 |
| G. sinense J.D. Zhao, L.W. Hsu & X.Q. Zhang | SA 17092559 | China | MK345452 |
| G. sinense | SA 17092539 | China | MK345453 |
| G. steyaertanum B.J. Sm. & Sivasith. | MEL:2382783 | Australia | KP012964 |
| G. steyaertanum | 6-WN-20BL-B | Indonesia | KJ654462 |
| G. subresinosum (Murrill) C.J. Humphrey | 5-D-3-D-26 | Indonesia | KJ654467 |
| G. subresinosum | LPDR 18011907 | Laos | MK345455 |
| G. thailandicum Luangharn, P.E. Mortimer, Karun. & J.C. Xu | HKAS 104640 (holotype) | Thailand | MK848681 |
| G. thailandicum | HKAS 104641 (paratype) | Thailand | MK848682 |
| G. tropicum (Jungh.) Bres. | Dai 9724 | China | JQ781879 |
| G. tropicum | TH 15081610 | Thailand | MK345456 |
| G. tsugae Murrill | Dai 12760 | USA | KJ143920 |
| G. tsugae | AFTOL-ID 771 | -- | DQ206985 |
| G. tuberculosum Murrill | GVL-21 | Mexico | MT232639 |
| G. tuberculosum | GVL-40 | Mexico | MT232634 |
| G. weberianum (Bres. & Henn. ex Sacc.) Steyaert | CBS 219.36 | Philippines | JQ520219 |
| G. weberianum | CBS 128581 | Taiwan, China | MK603805 |
| G. wiiroense E.C. Otto, Blanchette, C.W. Barnes &Held | UMN-20-GHA (paratype) | Ghana | KT952361 |
| G. wiiroense | UMN-21-GHA | Ghana | KT952363 |
| G. williamsianum Murrill | Dai 16809 | China | MG279183 |
| G. williamsianum | Wei 5032 | China | KU219994 |
| G. zonatum Murrill | FL-02 | USA | KJ143921 |
| G. zonatum | FL-03 | USA | KJ143922 |
| Tomophagus colossus (Fr.) Murrill | CBS 216.36 | Philippines | Z37071&Z37091 |
| T.s colossus | CGMCC 5.763 | Philippines | JQ081068 |
| Method | Primer Pair | Results | Product Size (bp) |
|---|---|---|---|
| Directed PCR | ITS5/ITS4 | * | 654 |
| ITS1/ITS4 | * | 633 | |
| ITS1/ITS2 | * | 266 | |
| ITS3/ITS4 | * | 367 | |
| ITS1/ITSGs1-2 | + | 116 | |
| ITSGs1-1/ITS2 | + | 157 | |
| ITS3/ITSGs2-2 | + | 180 | |
| ITSGs2-1/ITS4 | + | 176 | |
| Nested PCR | First round PCR | ||
| ITS5/ITS4 | * | 654 | |
| Second round PCR | |||
| ITS1/ITS4 | * | 633 | |
| ITS1/ITSGs4-4 | * | 597 | |
| ITS1/ITSGs2-4 | + | 567 | |
| ITS1/ITSGs4-2 | + | 576 | |
| ITSGs1-1/ITSGs2-4 | + | 458 | |
| ITSGs1-1/ITSGs4-4 | + | 488 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Z.; Li, Y.; Wang, X.-C.; Wang, K.; Yao, Y.-J. Re-Examination of the Holotype of Ganoderma sichuanense (Ganodermataceae, Polyporales) and a Clarification of the Identity of Chinese Cultivated Lingzhi. J. Fungi 2023, 9, 323. https://doi.org/10.3390/jof9030323
Du Z, Li Y, Wang X-C, Wang K, Yao Y-J. Re-Examination of the Holotype of Ganoderma sichuanense (Ganodermataceae, Polyporales) and a Clarification of the Identity of Chinese Cultivated Lingzhi. Journal of Fungi. 2023; 9(3):323. https://doi.org/10.3390/jof9030323
Chicago/Turabian StyleDu, Zhuo, Yi Li, Xin-Cun Wang, Ke Wang, and Yi-Jian Yao. 2023. "Re-Examination of the Holotype of Ganoderma sichuanense (Ganodermataceae, Polyporales) and a Clarification of the Identity of Chinese Cultivated Lingzhi" Journal of Fungi 9, no. 3: 323. https://doi.org/10.3390/jof9030323
APA StyleDu, Z., Li, Y., Wang, X.-C., Wang, K., & Yao, Y.-J. (2023). Re-Examination of the Holotype of Ganoderma sichuanense (Ganodermataceae, Polyporales) and a Clarification of the Identity of Chinese Cultivated Lingzhi. Journal of Fungi, 9(3), 323. https://doi.org/10.3390/jof9030323