
Acquired Triazole Resistance Alters Pathogenicity-Associated Features in Candida auris in an Isolate-Dependent Manner
 , ,
, ,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culturing Conditions
2.2. Generation of Evolved Strains and Microevolution Process
2.3. Antifungal Susceptibility Testing
2.4. General Growth Kinetics
2.5. Determining the Abiotic Stress Tolerance of the Evolved Strains by Spotting Assay
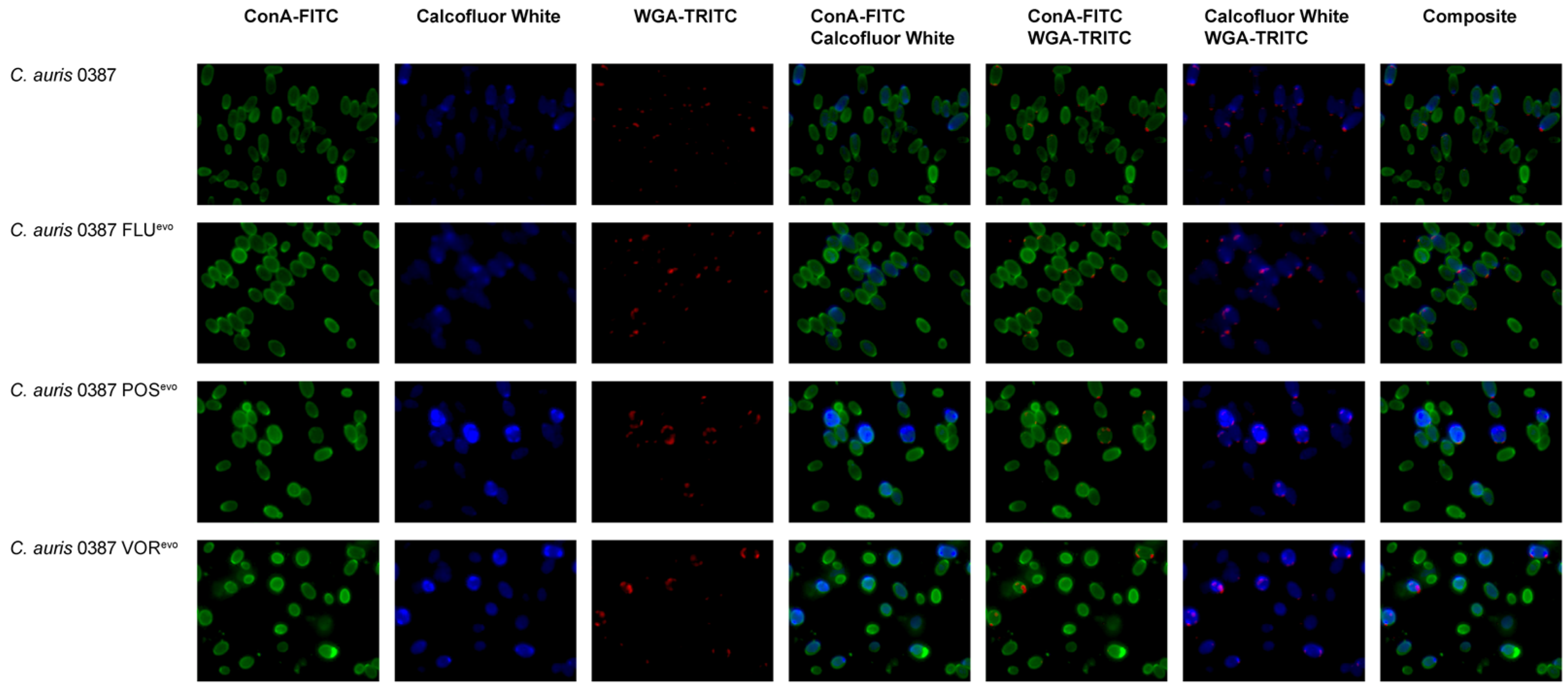
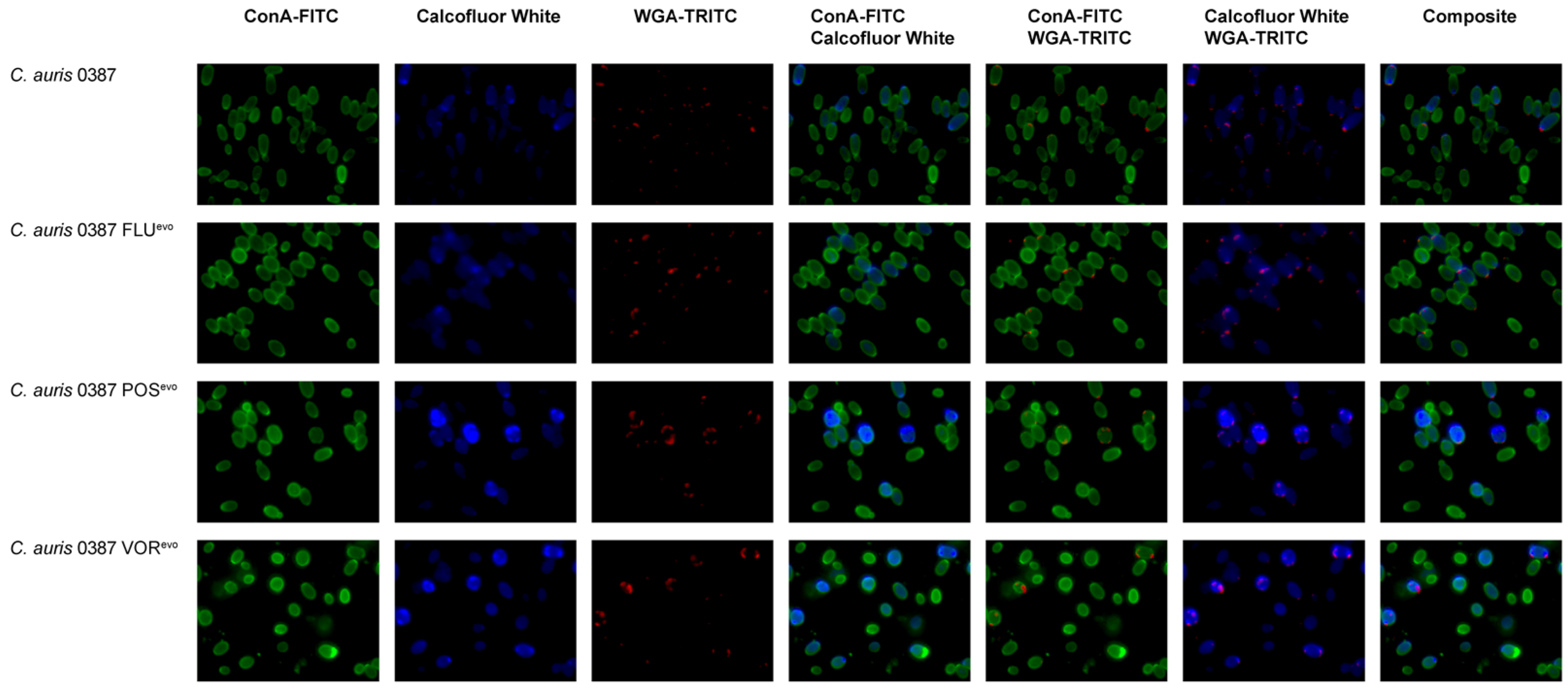
2.6. Cell Wall Staining
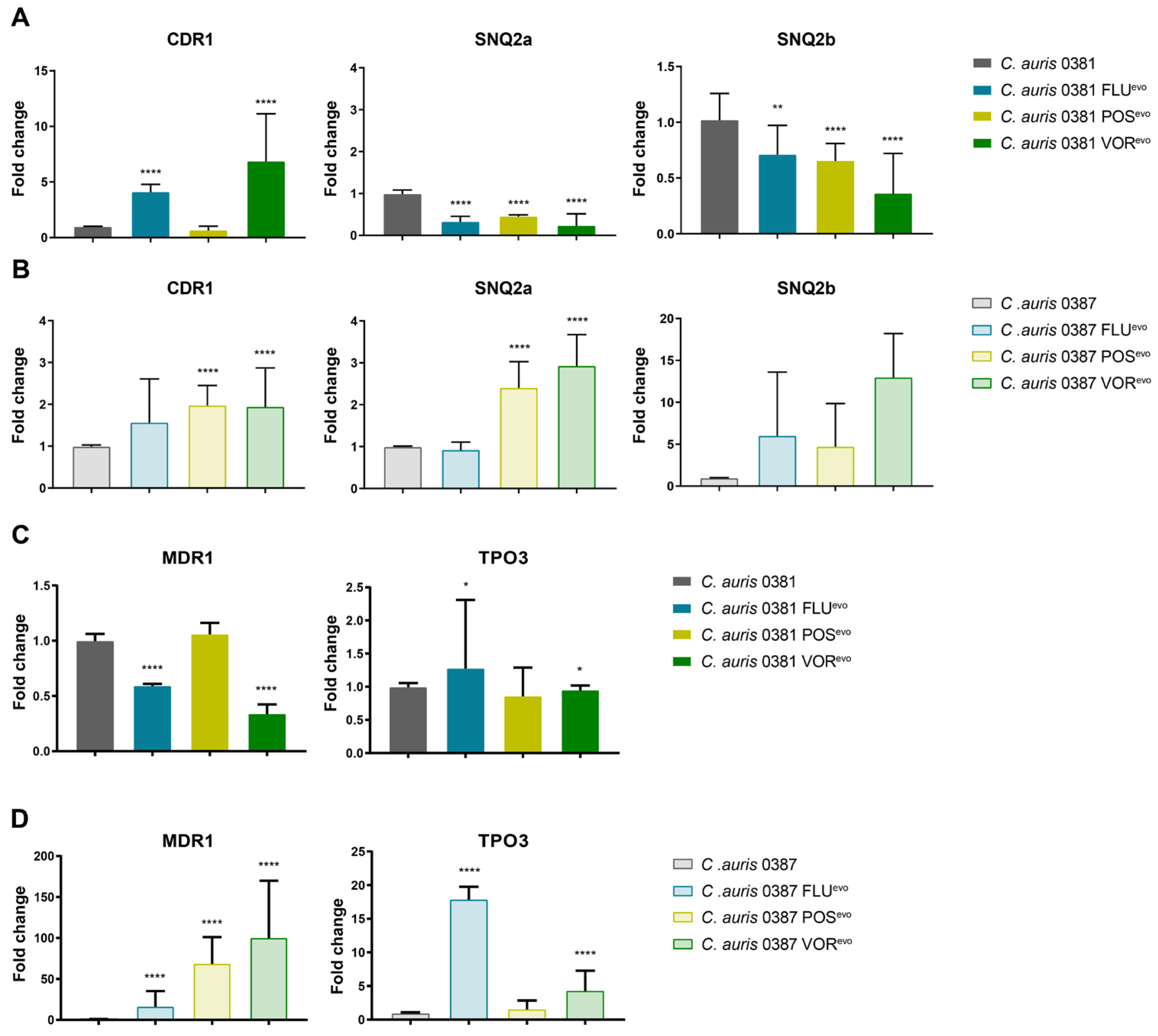
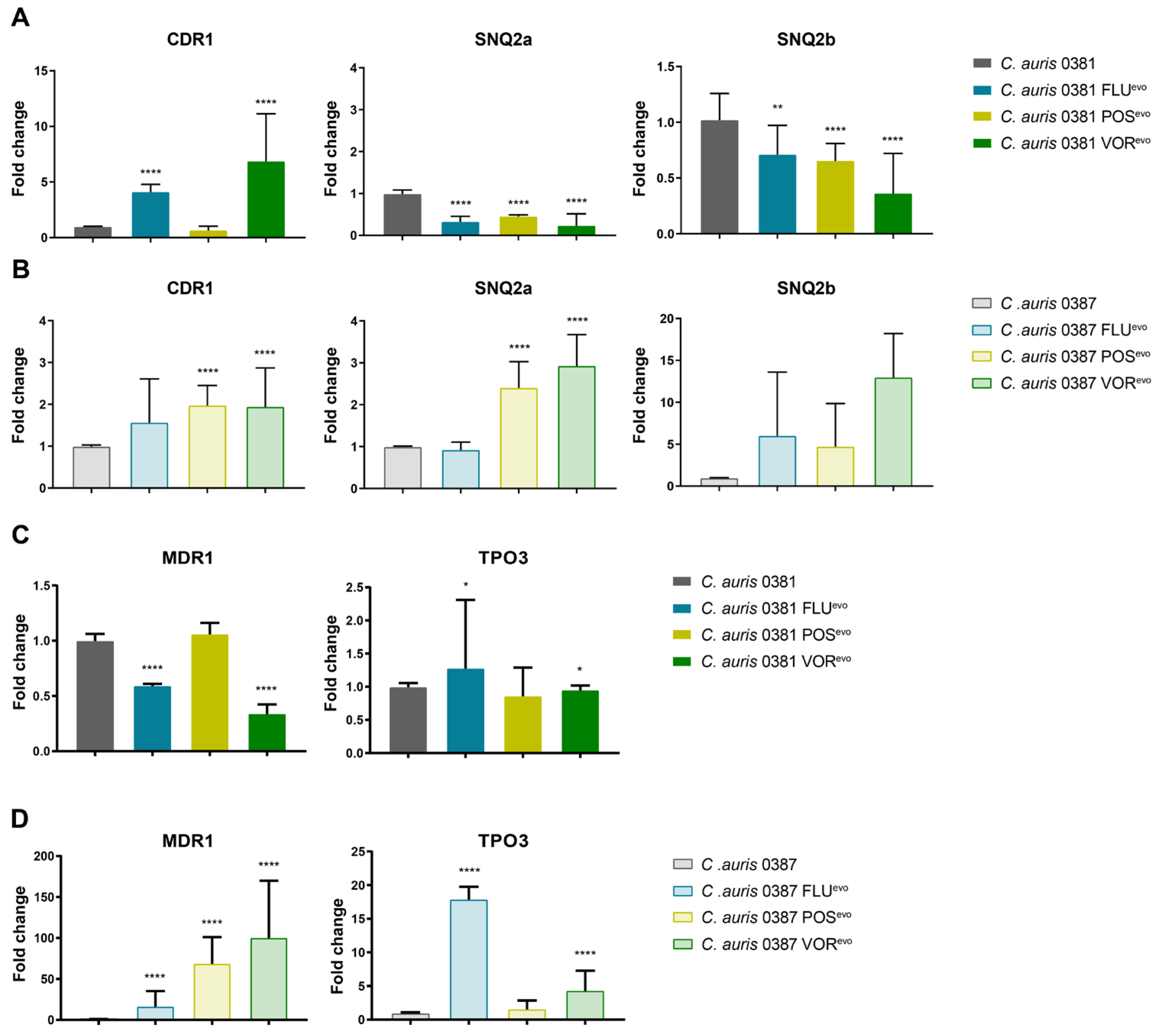
2.7. Efflux Pump Expression
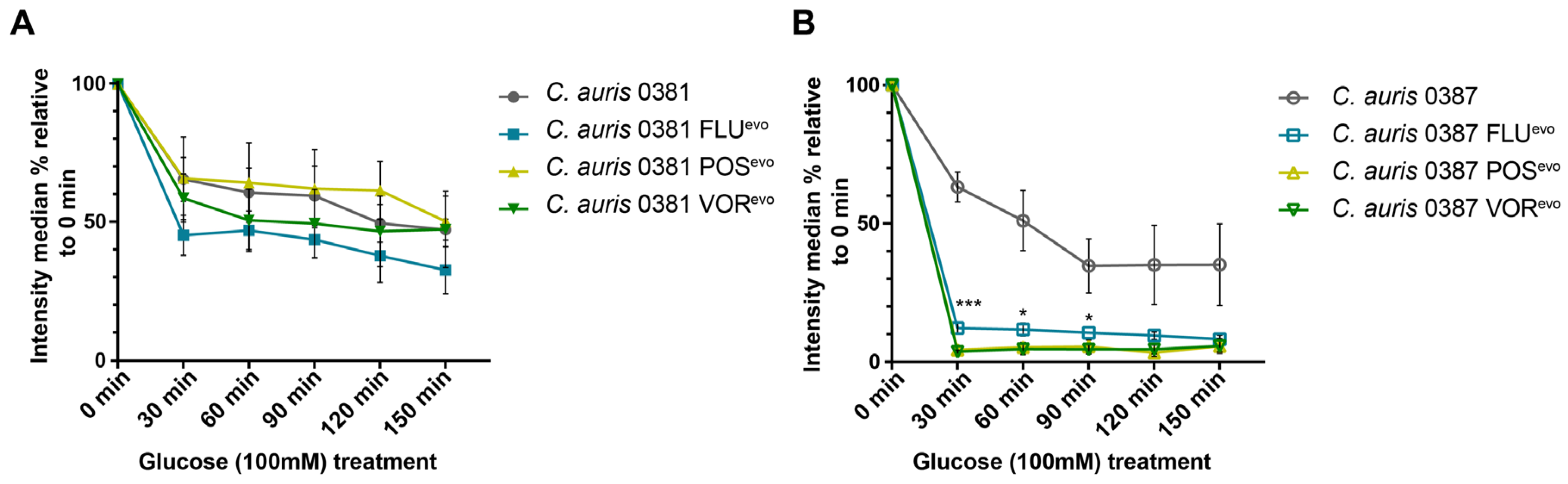
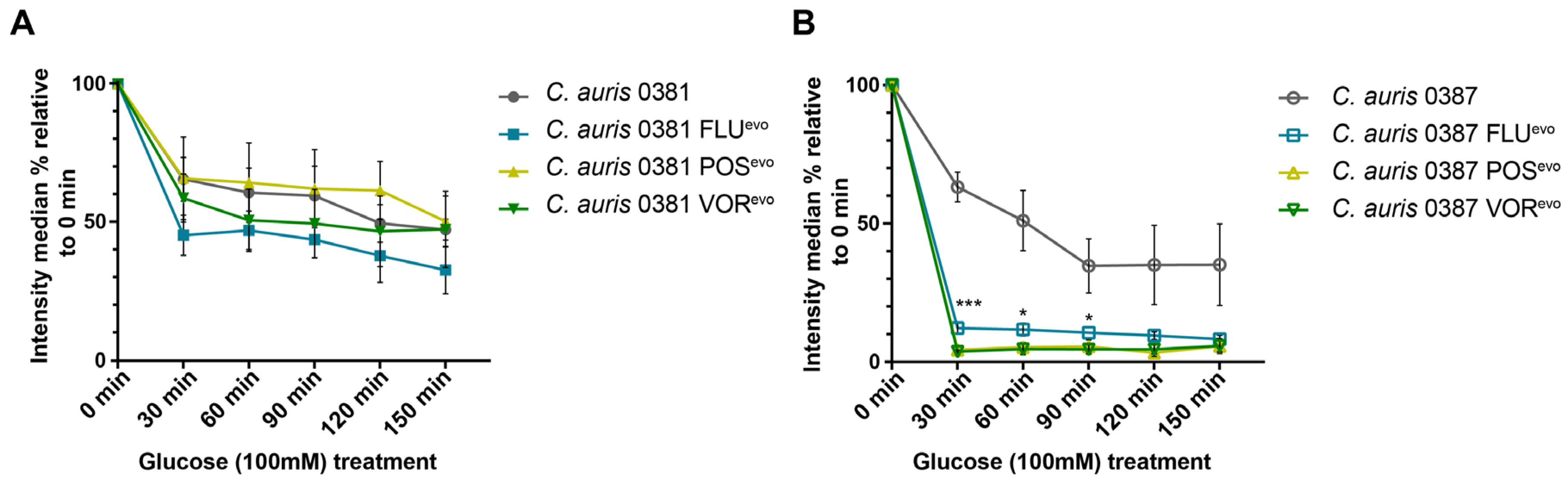
2.8. Efflux Activity Assay
2.9. XTT Reduction Assay
2.10. Analysis of Sterol Composition by LC-HRMS
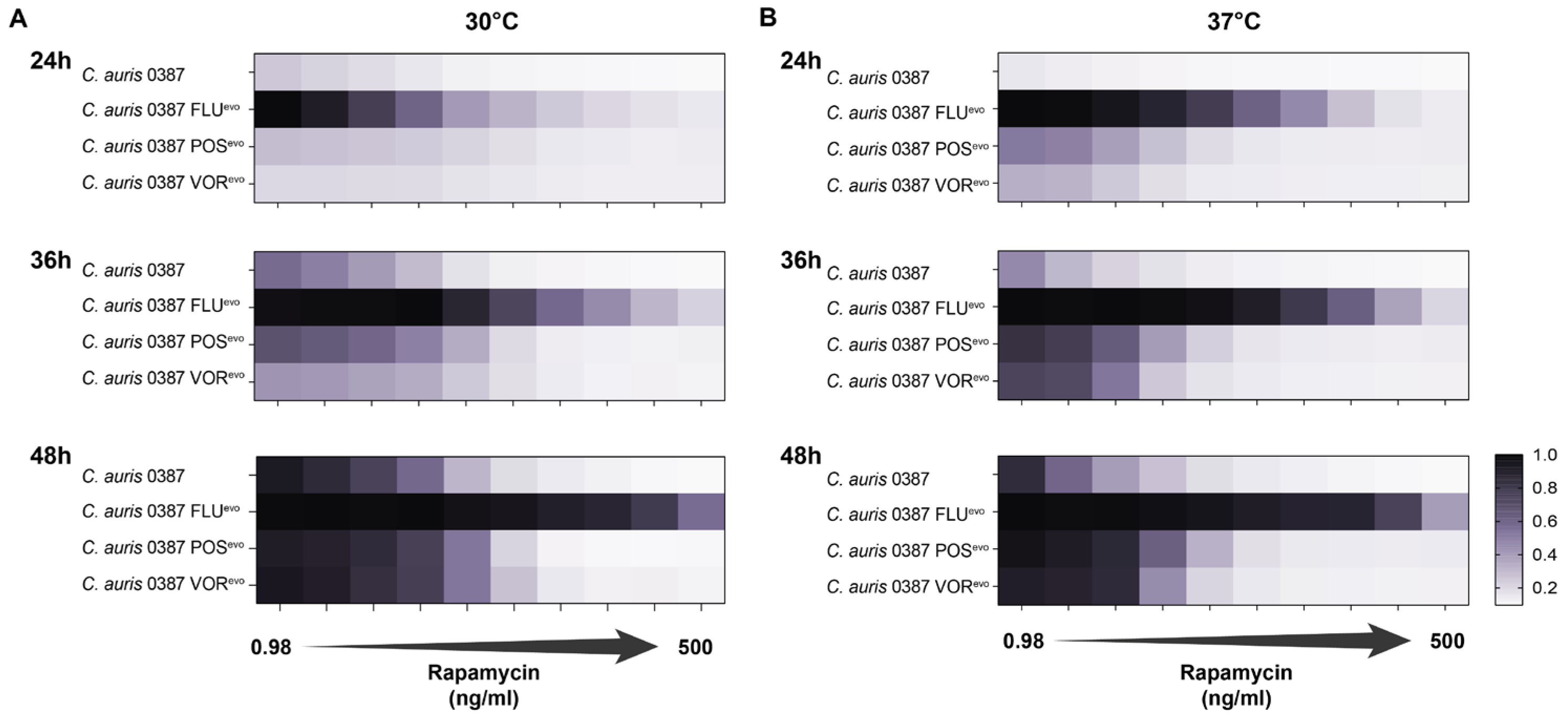
2.11. Rapamycin Susceptibility Assay in Liquid Media
2.12. DNA Extraction for Whole-Genome Sequencing
2.13. Genome Sequence Analysis
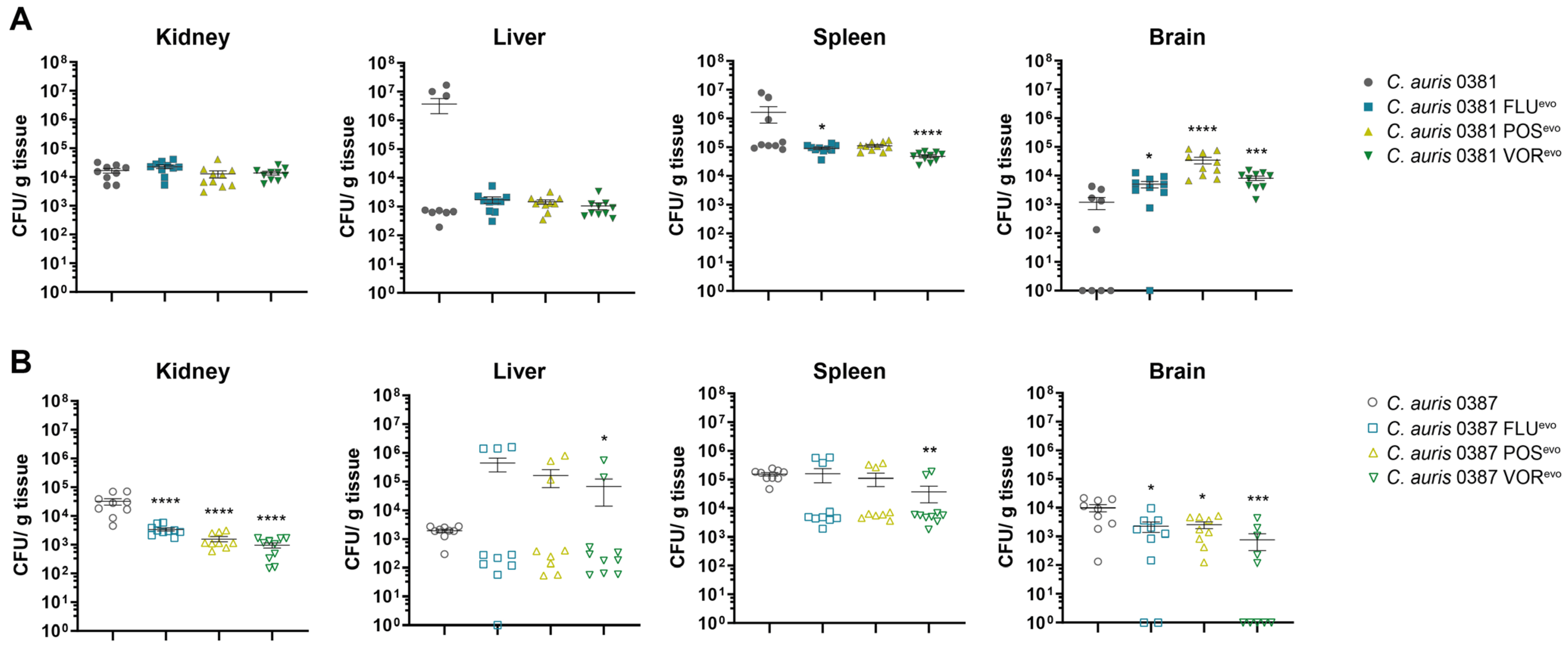
2.14. In Vivo Murine Infection and Determining of Fungal Burden
2.15. Statistical Tests
3. Results
3.1. Generation of C. auris Azole-Evolved Strains
3.2. Antifungal Susceptibility of C. auris Azole-Evolved Strains
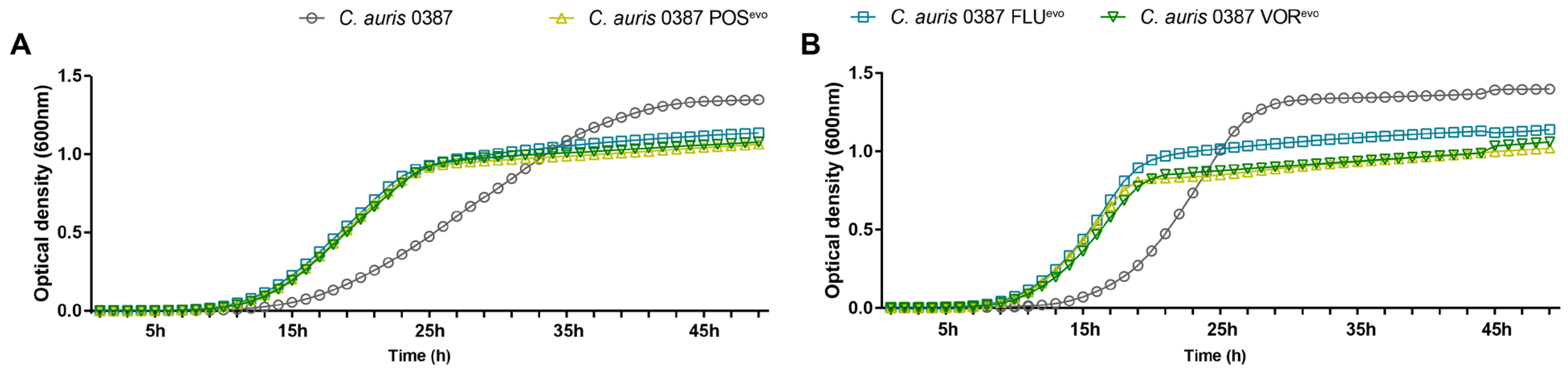
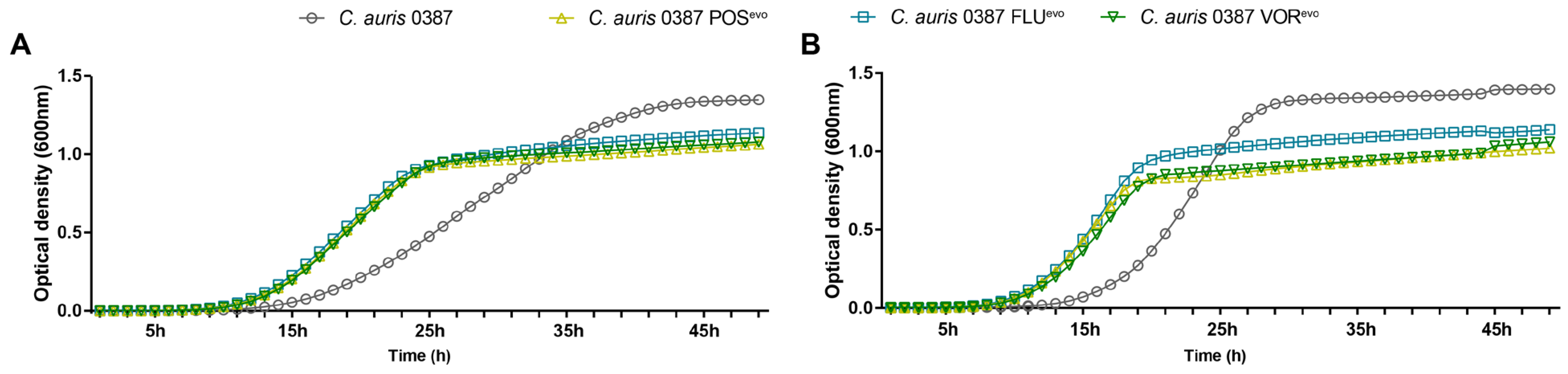
3.3. Growth Kinetics of the Evolved Strains in Complex Media
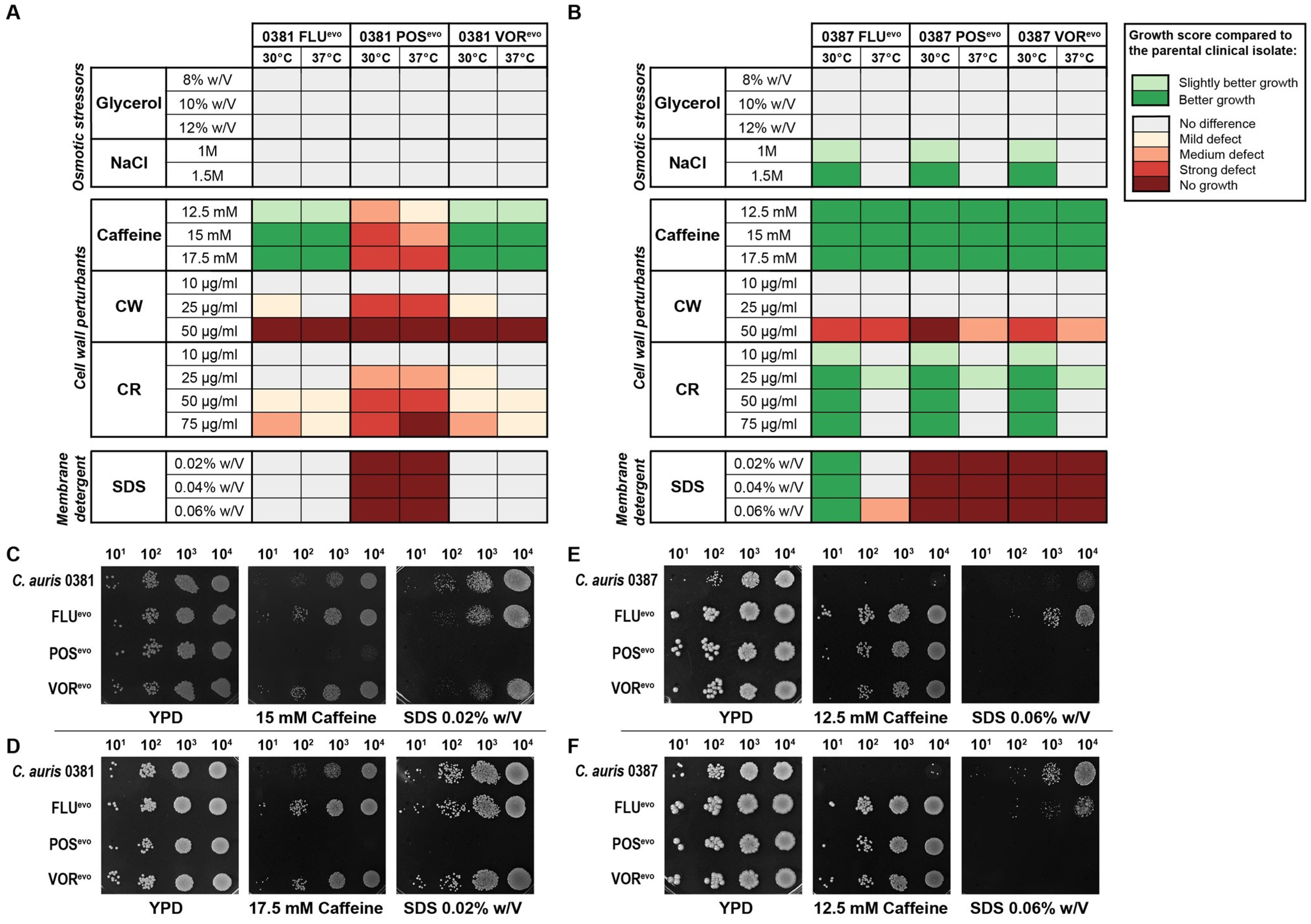
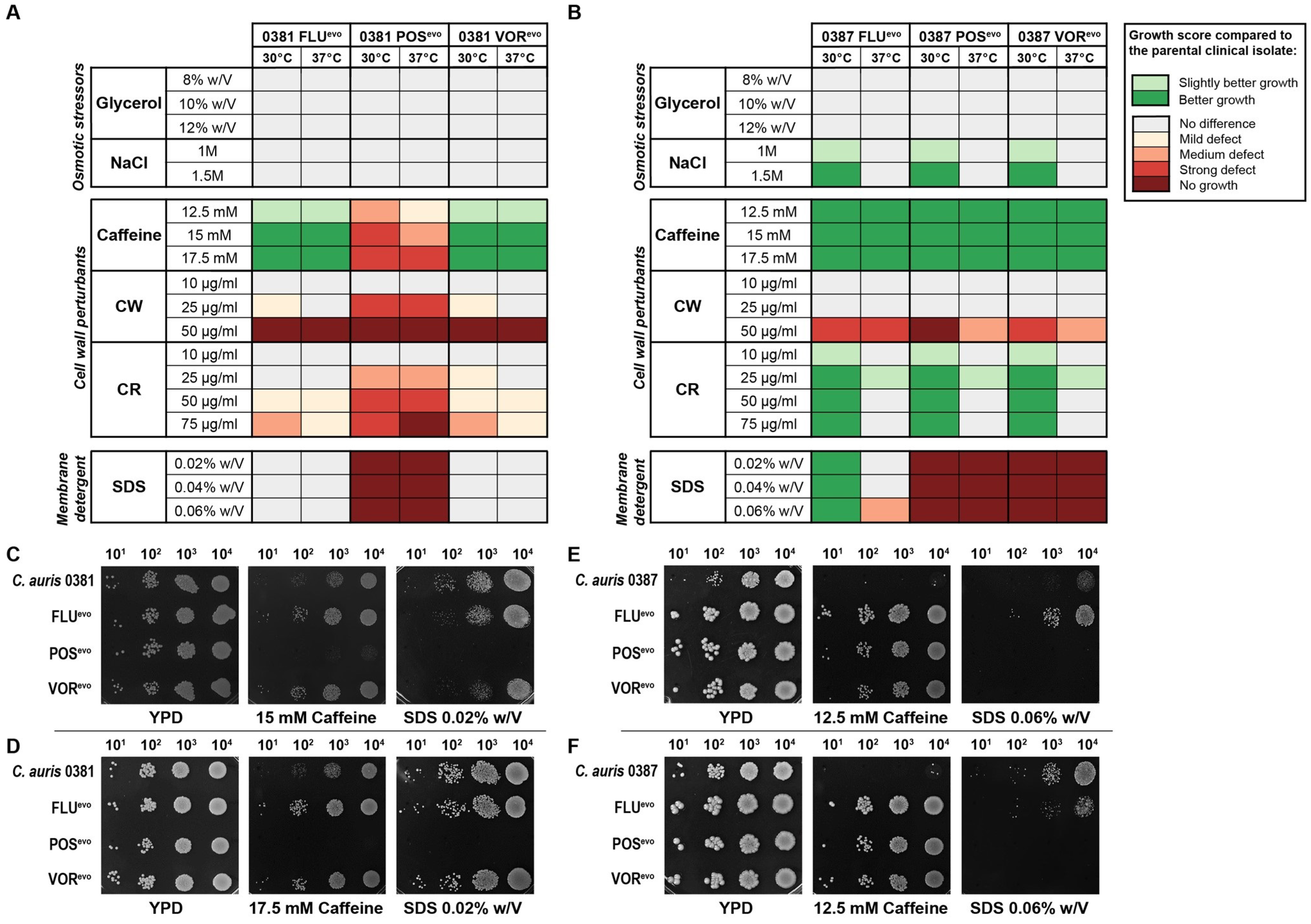
3.4. Triazole Resistance Development Alters Stress Tolerance in C. auris
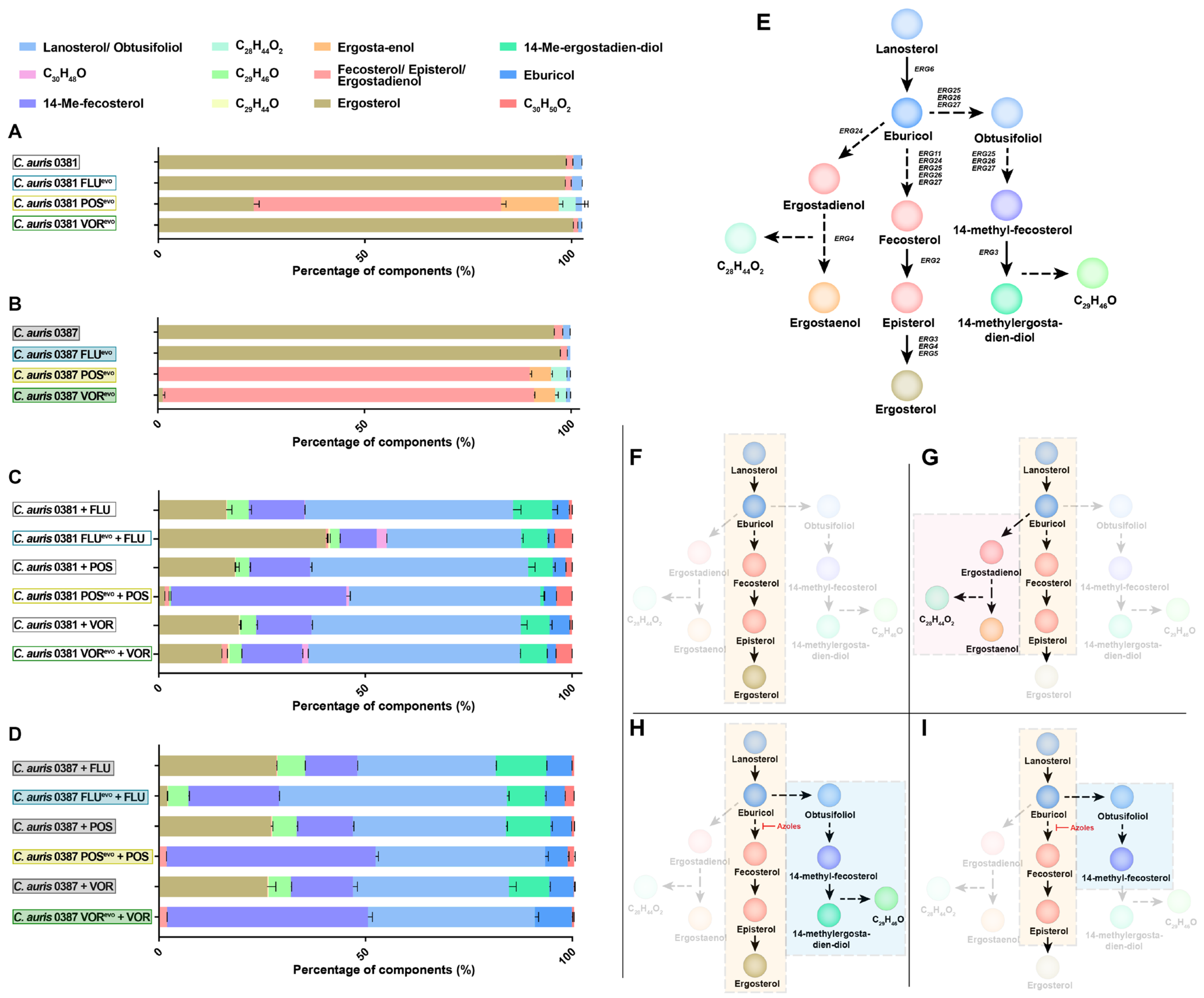
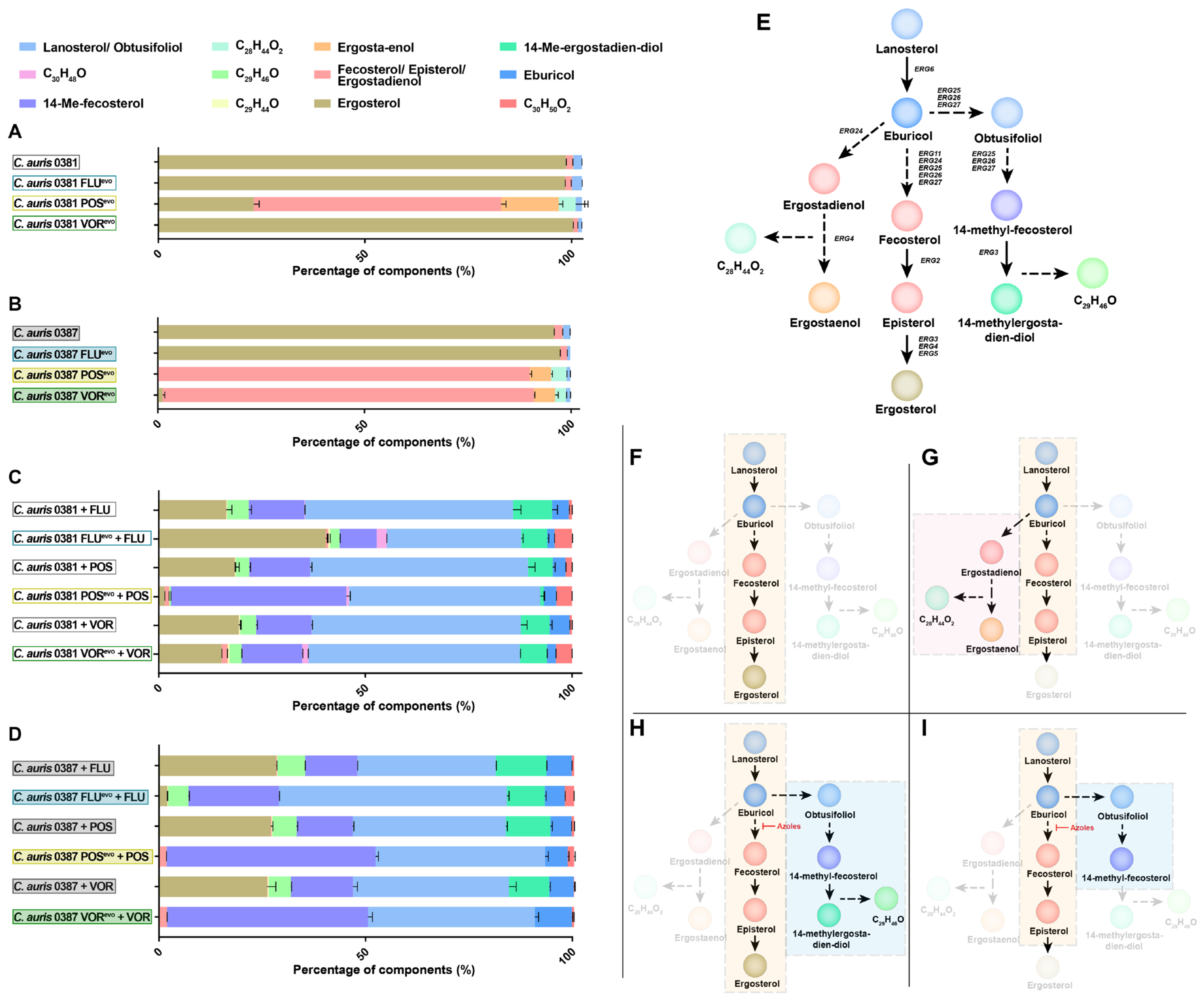
3.5. Azole Resistance Profoundly Alters the Ergosterol Content of C. auris Cell Membranes
3.6. Upon Antifungal Challenge, the Activation of the Alternative Pathway Promotes the Appearance of a Yet Uncharacterized Sterol Product, C29H46O
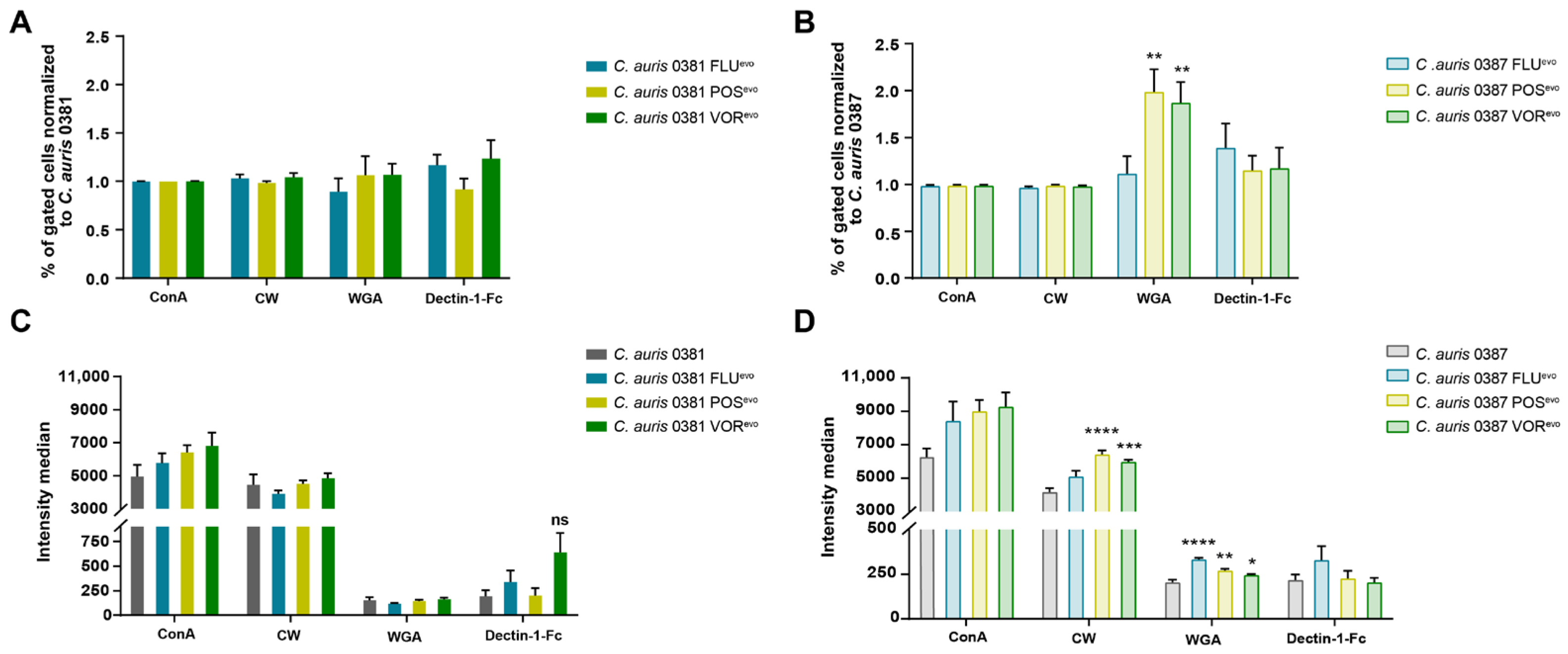
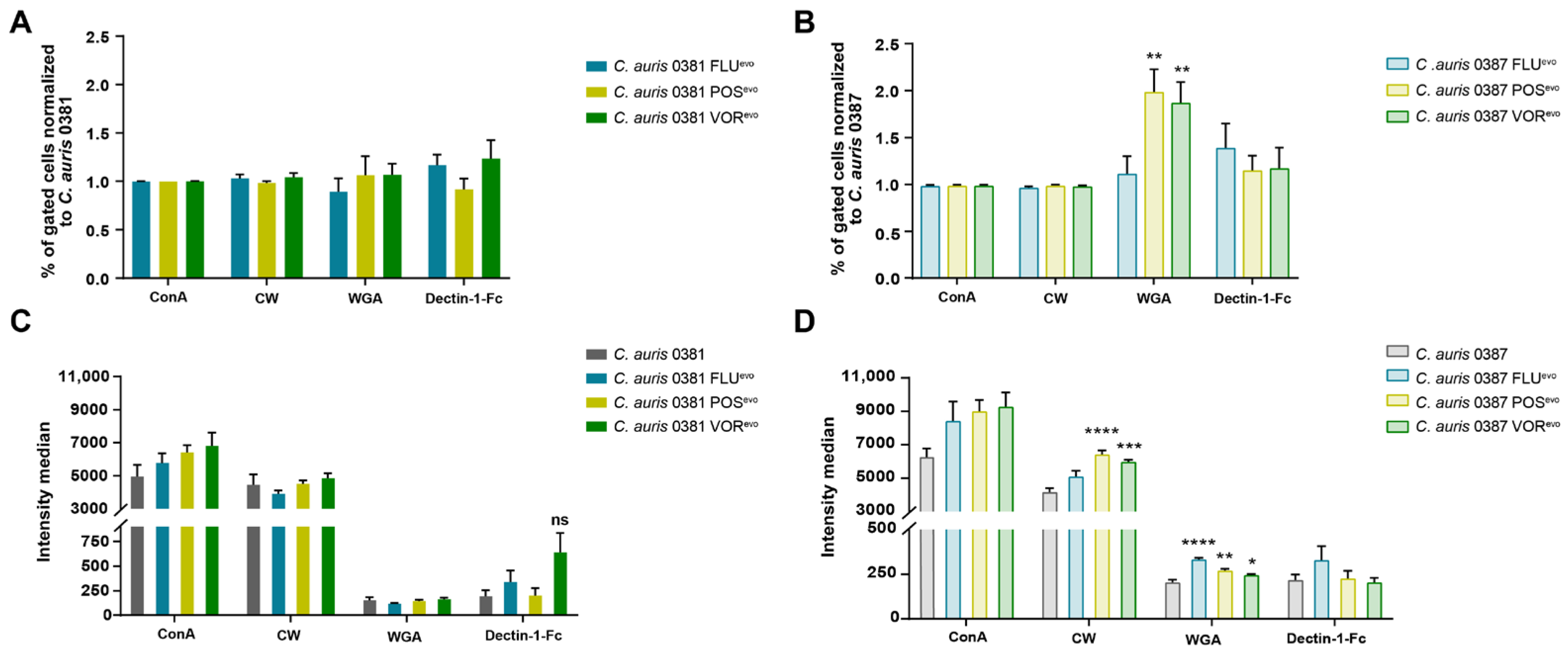
3.7. Flow Cytometry Analysis Reveals Modifications in the Cell Wall of Evolved Strains
3.8. Sequence Analysis of the Evolved Strains Suggests That Multiple Distinct Resistance Mechanisms Can Develop during Antifungal Treatment
3.9. Efflux Activity of the Triazole-Evolved Strains
3.10. Expression of Efflux Pump Genes in Evolved Strains
3.11. Function Loss of the Protein Kinase Inhibitor BCY1 Plays a Role in Rapamycin Resistance of C. auris
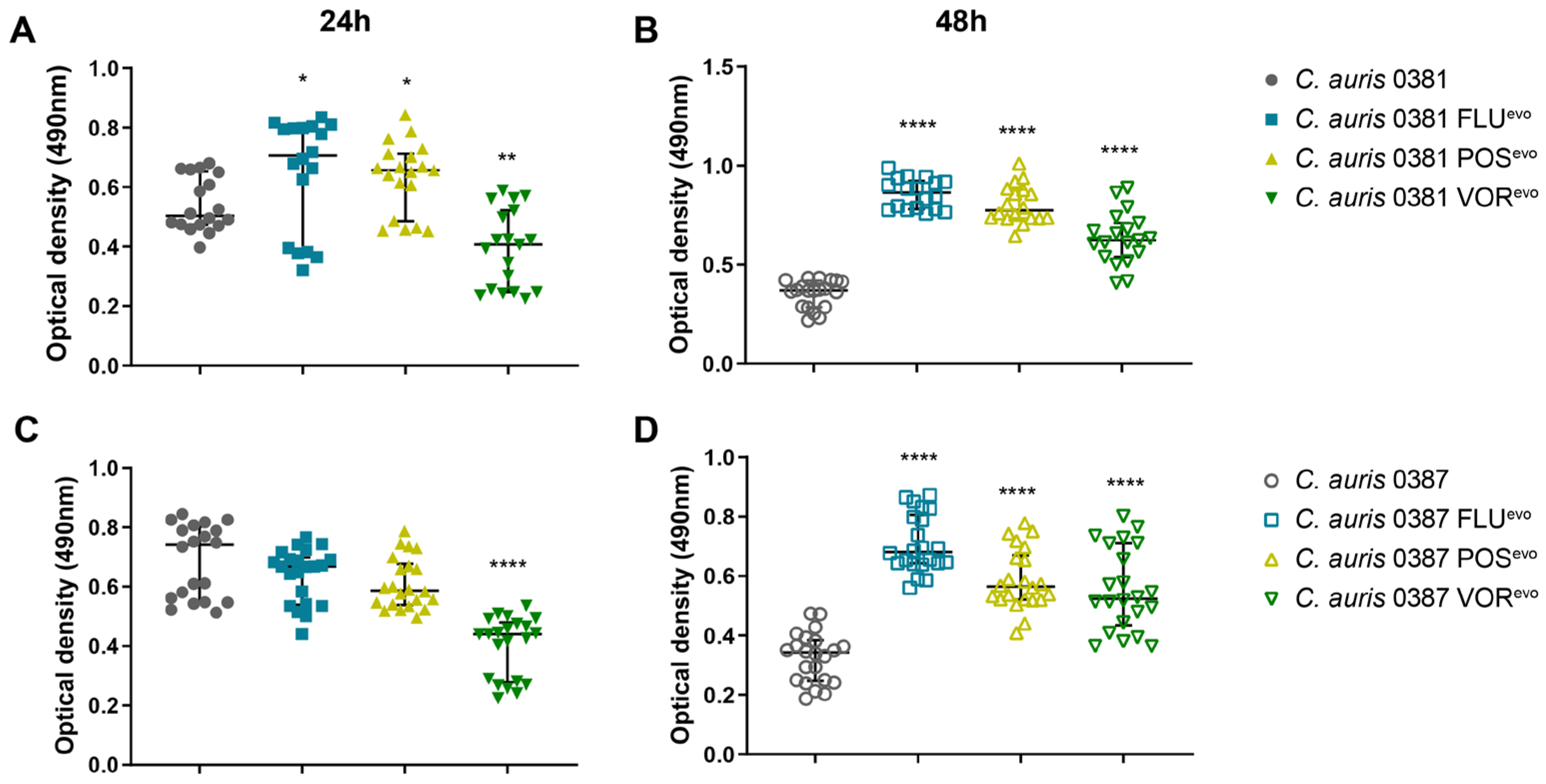
3.12. XTT Assay Indicates an Association between Biofilm Formation and Acquired Azole Resistance
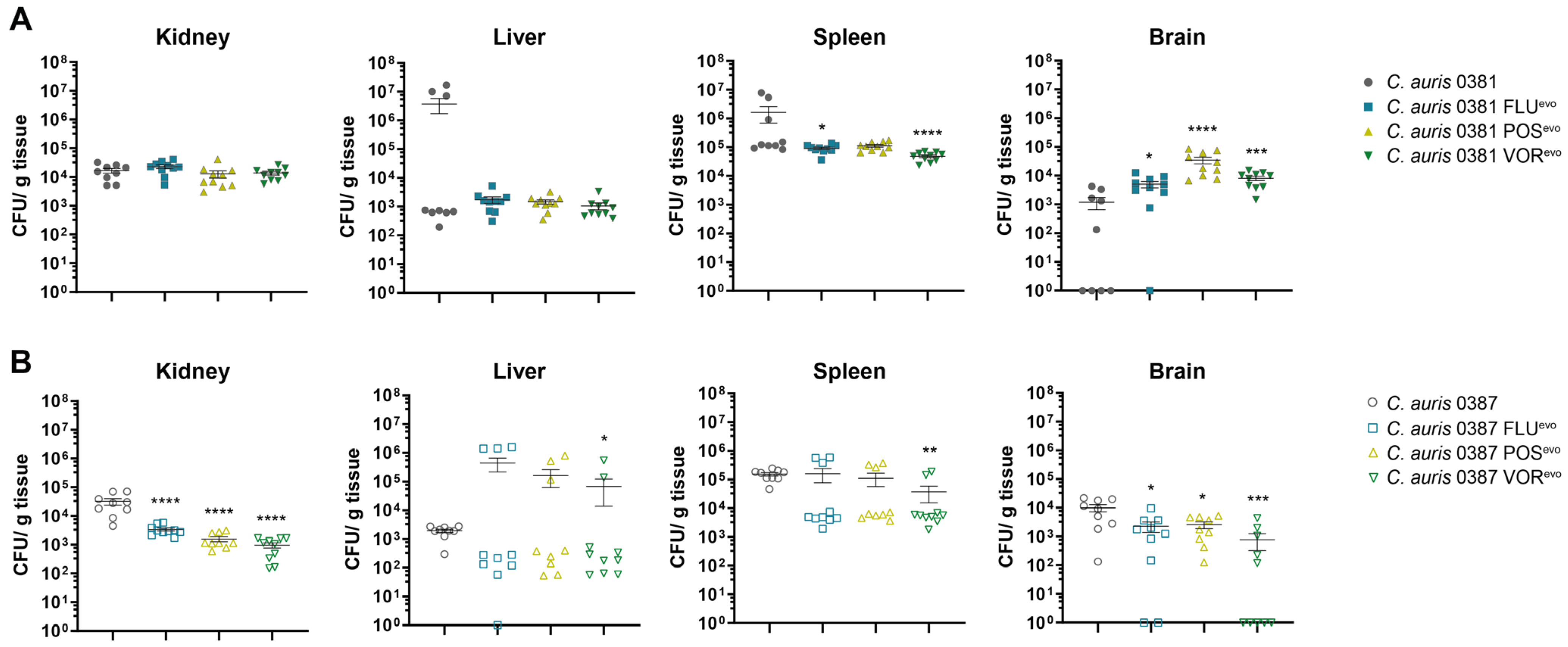
3.13. Acquired Triazole Resistance Significantly Alters the Virulence of Evolved Strains in Mice
4. Discussion
4.1. Experimental Evolution Can Induce Pan-Azole Resistance as Well as Caspofungin Cross-Resistance
4.2. Spotting Assay Indicated Changes in the Cell Wall and Membrane Homeostasis of the Evolved Strains
4.3. Posaconazole and Voriconazole Evolution Induced Unique Sterol Composition
4.4. Azole Treatment Promoted the Accumulation of a Yet Unclassified Sterol Product in C. auris
4.5. Microevolution Significantly Changes Efflux Processes
4.6. All 0387-Microevolved Strains Carry a Universal SNP in the PKA Pathway Inhibitor, BCY1
4.7. Upon the Acquisition of Antifungal Resistance, C. auris Strains Can Increase Their Pathogenic Potential
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris Sp. Nov., a Novel Ascomycetous Yeast Isolated from the External Ear Canal of an Inpatient in a Japanese Hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.G.; Shin, J.H.; Uh, Y.; Kang, M.G.; Kim, S.H.; Park, K.H.; Jang, H.C. First Three Reported Cases of Nosocomial Fungemia Caused by Candida auris. J. Clin. Microbiol. 2011, 49, 3139–3142. [Google Scholar] [CrossRef] [PubMed]
- Kathuria, S.; Singh, P.K.; Sharma, C.; Prakash, A.; Masih, A.; Kumar, A.; Meis, J.F.; Chowdhary, A. Multidrug-Resistant Candida auris Misidentified as Candida haemulonii: Characterization by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry and DNA Sequencing and Its Antifungal Susceptibility Profile Variability by Vitek 2, CL. J. Clin. Microbiol. 2015, 53, 1823–1830. [Google Scholar] [CrossRef]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First Hospital Outbreak of the Globally Emerging Candida auris in a European Hospital. Antimicrob. Resist. Infect. Control 2016, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.; Fisher, M.C. Global Epidemiology of Emerging Candida auris. Curr. Opin. Microbiol. 2019, 52, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R. Candida auris and Multidrug Resistance: Defining the New Normal. Fungal Genet. Biol. 2019, 131, 103243. [Google Scholar] [CrossRef] [PubMed]
- Jeffery-Smith, A.; Taori, S.K.; Schelenz, S.; Jeffery, K.; Johnson, E.M.; Borman, A.; Manuel, R.; Browna, C.S. Candida auris: A Review of the Literature. Clin. Microbiol. Rev. 2018, 31, e00029-17. [Google Scholar] [CrossRef] [PubMed]
- Chow, N.A.; De Groot, T.; Badali, H.; Abastabar, M.; Chiller, T.M.; Meis, J.F. Potential Fifth Clade of Candida auris, Iran, 2018. Emerg. Infect. Dis. 2019, 25, 1780–1781. [Google Scholar] [CrossRef]
- Prakash, A.; Sharma, C.; Singh, A.; Kumar Singh, P.; Kumar, A.; Hagen, F.; Govender, N.P.; Colombo, A.L.; Meis, J.F.; Chowdhary, A. Evidence of Genotypic Diversity among Candida auris Isolates by Multilocus Sequence Typing, Matrix-Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry and Amplified Fragment Length Polymorphism. Clin. Microbiol. Infect. 2016, 22, 277.e1–277.e9. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef]
- Welsh, R.M.; Sexton, D.J.; Forsberg, K.; Vallabhaneni, S.; Litvintseva, A. Insights into the Unique Nature of the East Asian Clade of the Emerging Pathogenic Yeast Candida auris. J. Clin. Microbiol. 2019, 57, e00007-19. [Google Scholar] [CrossRef] [PubMed]
- Ksiezopolska, E.; Gabaldón, T. Evolutionary Emergence of Drug Resistance in Candida Opportunistic Pathogens. Genes 2018, 9, 461. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Bing, J.; Hu, T.; Ennis, C.L.; Nobile, C.J.; Huang, G. Candida auris: Epidemiology, Biology, Antifungal Resistance, and Virulence. PLoS Pathog. 2020, 16, e1008921. [Google Scholar] [CrossRef] [PubMed]
- Parums, D.V. Editorial: The World Health Organization (WHO) Fungal Priority Pathogens List in Response to Emerging Fungal Pathogens during the COVID-19 Pandemic. Med. Sci. Monit. 2022, 28, e939088-1–e939088-3. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Alfouzan, W. Candida auris: Epidemiology, Diagnosis, Pathogenesis, Antifungal Susceptibility, and Infection Control Measures to Combat the Spread of Infections in Healthcare Facilities. Microorganisms 2021, 9, 807. [Google Scholar] [CrossRef] [PubMed]
- Chow, N.A.; Gade, L.; Tsay, S.V.; Forsberg, K.; Greenko, J.A.; Southwick, K.L.; Barrett, P.M.; Kerins, J.L.; Lockhart, S.R.; Chiller, T.M.; et al. Multiple Introductions and Subsequent Transmission of Multidrug-Resistant Candida auris in the USA: A Molecular Epidemiological Survey. Lancet Infect. Dis. 2018, 18, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Wickes, B.L. Analysis of a Candida auris Outbreak Provides New Insights into an Emerging Pathogen. J. Clin. Microbiol. 2020, 58, e02083-19. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Chakrabarti, A. Strategies to Reduce Mortality in Adult and Neonatal Candidemia in Developing Countries. J. Fungi 2017, 3, 41. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Patterson, T.F. Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef]
- Nguyen, M.H.; Yu, C.Y. Voriconazole against Fluconazole-Susceptible and Resistant Candida Isolates: In-Vitro Efficacy Compared with That of Itraconazole and Ketoconazole. J. Antimicrob. Chemother. 1998, 42, 253–256. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Twenty Years of the SENTRY Antifungal Surveillance Program: Results for Candida Species from 1997–2016. Open Forum Infect. Dis. 2019, 6, S79–S94. [Google Scholar] [CrossRef] [PubMed]
- Mayr, E.-M.; Ramírez-Zavala, B.; Krüger, I.; Morschhäuser, J. A Zinc Cluster Transcription Factor Contributes to the Intrinsic Fluconazole Resistance of Candida auris. mSphere 2020, 5, e00279-20. [Google Scholar] [CrossRef] [PubMed]
- Page, A.V.; Liles, W.C. Posaconazole: A New Agent for the Prevention and Management of Severe, Refractory or Invasive Fungal Infections. Can. J. Infect. Dis. Med. Microbiol. 2008, 19, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Pandya, N.; Cag, Y.; Pandak, N.; Pekok, A.U.; Poojary, A.; Ayoade, F.; Fasciana, T.; Giammanco, A.; Caskurlu, H.; Rajani, D.P.; et al. International Multicentre Study of Candida auris Infections. J. Fungi 2021, 7, 878. [Google Scholar] [CrossRef] [PubMed]
- Fakhim, H.; Chowdhary, A.; Prakash, A.; Vaezi, A.; Dannaoui, E.; Meis, J.F.; Badali, H. In Vitro Interactions of Echinocandins with Triazoles against Multidrug-Resistant Candida auris. Antimicrob. Agents Chemother. 2017, 61, e01056-17. [Google Scholar] [CrossRef]
- Kim, S.H.; Iyer, K.R.; Pardeshi, L.; Muñoz, J.F.; Robbins, N.; Cuomo, C.A.; Wong, K.H.; Cowen, L.E. Genetic Analysis of Candida auris Implicates Hsp90 in Morphogenesis and Azole Tolerance and Cdr1 in Azole Resistance. mBio 2019, 10, e02529-18. [Google Scholar] [CrossRef] [PubMed]
- Rybak, J.M.; Doorley, L.A.; Nishimoto, A.T.; Barker, K.S.; Palmer, G.E.; Rogers, P.D. Abrogation of Triazole Resistance upon Deletion of CDR1 in a Clinical Isolate of Candida auris. Antimicrob. Agents Chemother. 2019, 63, e00057-19. [Google Scholar] [CrossRef]
- Healey, K.R.; Kordalewska, M.; Ortigosa, C.J.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 Mutations Identified in Isolates of Candida auris Directly Contribute to Reduced Azole Susceptibility. Antimicrob. Agents Chemother. 2018, 62, e01427-18. [Google Scholar] [CrossRef]
- Dolande, M.; García, N.; Capote, A.M.; Panizo, M.M.; Ferrara, G.; Alarcón, V. Candida auris: Antifungal Multi-Resistant Emerging Yeast. Curr. Fungal Infect. Rep. 2017, 11, 197–202. [Google Scholar] [CrossRef]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic Insights into Multidrug-Resistance, Mating and Virulence in Candida auris and Related Emerging Species. Nat. Commun. 2018, 9, 5346. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Holowka, T.; Orner, E.P.; Fries, B.C. Gene Duplication Associated with Increased Fluconazole Tolerance in Candida auris Cells of Advanced Generational Age. Sci. Rep. 2019, 9, 5052. [Google Scholar] [CrossRef] [PubMed]
- Knorre, D.A.; Galkina, K.V.; Shirokovskikh, T.; Banerjee, A.; Prasad, R. Do Multiple Drug Resistance Transporters Interfere with Cell Functioning under Normal Conditions? Biochemistry 2020, 85, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Wasi, M.; Kumar Khandelwal, N.; Moorhouse, A.J.; Nair, R.; Vishwakarma, P.; Bravo Ruiz, G.; Ross, Z.K.; Lorenz, A.; Rudramurthy, S.M.; Chakrabarti, A.; et al. ABC Transporter Genes Show Upregulated Expression in Drug-Resistant Clinical Isolates of Candida auris: A Genome-Wide Characterization of Atp-Binding Cassette (Abc) Transporter Genes. Front. Microbiol. 2019, 10, 1445. [Google Scholar] [CrossRef]
- Rybak, J.M.; Muñoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida auris. mBio 2020, 11, e00365-20. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Lee, K.T.; Lee, M.H.; Cheong, E.; Bahn, Y.S. Adenylyl Cyclase and Protein Kinase A Play Redundant and Distinct Roles in Growth, Differentiation, Antifungal Drug Resistance, and Pathogenicity of Candida auris. mBio 2021, 12, e0272921. [Google Scholar] [CrossRef] [PubMed]
- Elena, S.F.; Lenski, R.E. Evolution Experiments with Microorganisms: The Dynamics and Genetic Bases of Adaptation. Nat. Rev. Genet. 2003, 4, 457–469. [Google Scholar] [CrossRef]
- Beekman, C.N.; Ene, I.V. Short-Term Evolution Strategies for Host Adaptation and Drug Escape in Human Fungal Pathogens. PLoS Pathog. 2020, 16, e1008519. [Google Scholar] [CrossRef]
- Carolus, H.; Pierson, S.; Muñoz, J.F.; Subotić, A.; Cruz, R.B.; Cuomo, C.A.; Van Dijck, P. Genome-Wide Analysis of Experimentally Evolved Candida auris Reveals Multiple Novel Mechanisms of Multidrug Resistance. mBio 2021, 12, e03333-20. [Google Scholar] [CrossRef]
- Bing, J.; Hu, T.; Zheng, Q.; Muñoz, J.F.; Cuomo, C.A.; Huang, G. Experimental Evolution Identifies Adaptive Aneuploidy as a Mechanism of Fluconazole Resistance in Candida auris. Antimicrob. Agents Chemother. 2021, 65, 2001–2014. [Google Scholar] [CrossRef]
- Papp, C.; Bohner, F.; Kocsis, K.; Varga, M.; Szekeres, A.; Bodai, L.; Willis, J.R.; Gabaldón, T.; Tóth, R.; Nosanchuk, J.D.; et al. Triazole Evolution of Candida parapsilosis Results in Cross-Resistance to Other Antifungal Drugs, Influences Stress Responses, and Alters Virulence in an Antifungal Drug-Dependent Manner. mSphere 2020, 5, e00821-20. [Google Scholar] [CrossRef]
- Pais, P.; Galocha, M.; Viana, R.; Cavalheiro, M.; Pereira, D.; Teixeira, M.C. Microevolution of the Pathogenic Yeasts Candida albicans and Candida glabrata during Antifungal Therapy and Host Infection. Microb. Cell 2019, 6, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Ischer, F.; Calabrese, D.; Posteraro, B.; Sanguinetti, M.; Fadda, G.; Rohde, B.; Bauser, C.; Bader, O.; Sanglard, D. Gain of Function Mutations in CgPDR1 of Candida glabrata Not Only Mediate Antifungal Resistance but Also Enhance Virulence. PLoS Pathog. 2009, 5, e1000268. [Google Scholar] [CrossRef] [PubMed]
- Vale-Silva, L.A.; Sanglard, D. Tipping the Balance Both Ways: Drug Resistance and Virulence in Candida glabrata. FEMS Yeast Res. 2015, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Burrack, L.S.; Todd, R.T.; Soisangwan, N.; Wiederhold, N.P.; Selmecki, A. Genomic Diversity across Candida auris Clinical Isolates Shapes Rapid Development of Antifungal Resistance In Vitro and In Vivo. mBio 2022, 13, e00842-22. [Google Scholar] [CrossRef] [PubMed]
- Bravo Ruiz, G.; Ross, Z.K.; Holmes, E.; Schelenz, S.; Gow, N.A.R.; Lorenz, A. Rapid and Extensive Karyotype Diversification in Haploid Clinical Candida auris Isolates. Curr. Genet. 2019, 65, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, A.; Kumar, P.; Chauhan, A.; Kumar, M.; Yadav, K.; Banerjee, A.; Sharma, R.D.; Rudramurthy, S.M.; Chakrabarti, A.; Sanyal, K.; et al. Directed Evolution Detects Supernumerary Centric Chromosomes Conferring Resistance to Azoles in Candida auris. mBio 2022, 13, e03052-22. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Approved Standard. CLSI Document M27-A3; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008; ISBN 1-56238-666-2.
- Centers for Disease Control and Prevention. Recommendations for Identification of Candida auris. 2017. Available online: https://www.cdc.gov/fungal/diseases/candidiasis/recommendations.html (accessed on 29 November 2018).
- Arendrup, M.C.; Prakash, A.; Meletiadis, J.; Sharma, C.; Chowdhary, A. Comparison of EUCAST and CLSI Reference Microdilution Mics of Eight Antifungal Compounds for Candida auris and Associated Tentative Epidemiological Cutoff Values. Antimicrob. Agents Chemother. 2017, 61, e00485-17. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Sprouffske, K.; Wagner, A. Growthcurver: An R Package for Obtaining Interpretable Metrics from Microbial Growth Curves. BMC Bioinform. 2016, 17, 172. [Google Scholar] [CrossRef]
- Papp, C.; Kocsis, K.; Tóth, R.; Bodai, L.; Willis, J.R.; Ksiezopolska, E.; Lozoya-Pérez, N.E.; Vágvölgyi, C.; Mora Montes, H.; Gabaldón, T.; et al. Echinocandin-Induced Microevolution of Candida parapsilosis Influences Virulence and Abiotic Stress Tolerance. mSphere 2018, 3, e00547-18. [Google Scholar] [CrossRef]
- Varga, M.; Bartók, T.; Mesterházy, Á. Determination of Ergosterol in Fusarium-Infected Wheat by Liquid Chromatography-Atmospheric Pressure Photoionization Mass Spectrometry. J. Chromatogr. A 2006, 1103, 278–283. [Google Scholar] [CrossRef]
- Tong, Y.; Zhang, J.; Wang, L.; Wang, Q.; Huang, H.; Chen, X.; Zhang, Q.; Li, H.; Sun, N.; Liu, G.; et al. Hyper-Synergistic Antifungal Activity of Rapamycin and Peptide-Like Compounds against Candida albicans Orthogonally via Tor1 Kinase. ACS Infect. Dis. 2021, 7, 2826–2835. [Google Scholar] [CrossRef]
- Kadosh, D.; Johnson, A.D. Rfg1, a Protein Related to the Saccharomyces Cerevisiae Hypoxic Regulator Rox1, Controls Filamentous Growth and Virulence in Candida albicans. Mol. Cell. Biol. 2001, 21, 2496–2505. [Google Scholar] [CrossRef]
- Giacometti, R.; Kronberg, F.; Biondi, R.M.; Passeron, S. Catalytic Isoforms Tpk1 and Tpk2 of Candida albicans PKA Have Non-Redundant Roles in Stress Response and Glycogen Storage. Yeast 2009, 26, 273–285. [Google Scholar] [CrossRef]
- Lin, C.J.; Chen, Y.L. Conserved and Divergent Functions of the CAMP/PKA Signaling Pathway in Candida albicans and Candida tropicalis. J. Fungi 2018, 4, 68. [Google Scholar] [CrossRef]
- Hogan, D.A.; Sundstrom, P. The Ras/cAMP/PKA Signaling Pathway and Virulence in Candida albicans. Future Microbiol. 2009, 4, 1263–1270. [Google Scholar] [CrossRef]
- Brown, V.; Sexton, J.A.; Johnston, M. A Glucose Sensor in Candida albicans. Eukaryot. Cell 2006, 5, 1726–1737. [Google Scholar] [CrossRef]
- Ivnitski-Steele, I.; Holmes, A.R.; Lamping, E.; Monk, B.C.; Cannon, R.D.; Sklar, L.A. Identification of Nile Red as a Fluorescent Substrate of the Candida albicans ATP-Binding Cassette Transporters Cdr1p and Cdr2p and the Major Facilitator Superfamily Transporter Mdr1p. Anal. Biochem. 2009, 394, 87–91. [Google Scholar] [CrossRef]
- Soulard, A.; Cremonesi, A.; Moes, S.; Schütz, F.; Jenö, P.; Hall, M.N. The Rapamycin-Sensitive Phosphoproteome Reveals That TOR Controls Protein Kinase A toward Some but Not All Substrates. Mol. Biol. Cell 2010, 21, 3475–3486. [Google Scholar] [CrossRef]
- Kunkel, J.; Luo, X.; Capaldi, A.P. Integrated TORC1 and PKA Signaling Control the Temporal Activation of Glucose-Induced Gene Expression in Yeast. Nat. Commun. 2019, 10, 3558. [Google Scholar] [CrossRef]
- Schmelzle, T.; Beck, T.; Martin, D.E.; Hall, M.N. Activation of the RAS/Cyclic AMP Pathway Suppresses a TOR Deficiency in Yeast. Mol. Cell. Biol. 2004, 24, 338–351. [Google Scholar] [CrossRef]
- Chowdhury, T.; Köhler, J.R. Ribosomal Protein S6 Phosphorylation Is Controlled by TOR and Modulated by PKA in Candida albicans. Mol. Microbiol. 2015, 98, 384–402. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, Z.; Al-Sweih, N.; Alfouzan, W.; Joseph, L. Candida auris in Various Hospitals across Kuwait and Their Susceptibility and Molecular Basis of Resistance to Antifungal Drugs. Mycoses 2020, 63, 104–112. [Google Scholar] [CrossRef]
- You, L.; Qian, W.; Yang, Q.; Mao, L.; Zhu, L.; Huang, X.; Jin, J.; Meng, H. ERG11 Gene Mutations and MDR1 Upregulation Confer Pan-Azole Resistance in Candida tropicalis Causing Disseminated Candidiasis in an Acute Lymphoblastic Leukemia Patient on Posaconazole Prophylaxis. Antimicrob. Agents Chemother. 2017, 61, S54. [Google Scholar] [CrossRef]
- Ruckwardt, T.; Scott, A.; Scott, J.; Mikulecky, P.; Hartsel, S.C. Lipid and Stress Dependence of Amphotericin B Ion Selective Channels in Sterol-Free Membranes. Biochim. Biophys. Acta—Biomembr. 1998, 1372, 283–288. [Google Scholar] [CrossRef]
- Tsujimoto, Y.; Shimizu, Y.; Otake, K.; Nakamura, T.; Okada, R.; Miyazaki, T.; Watanabe, K. Multidrug Resistance Transporters Snq2p and Pdr5p Mediate Caffeine Efflux in Saccharomyces Cerevisiae. Biosci. Biotechnol. Biochem. 2015, 79, 1103–1110. [Google Scholar] [CrossRef]
- Ollinger, T.L.; Vu, B.; Murante, D.; Parker, J.E.; Simonicova, L.; Doorley, L.; Stamnes, M.A.; Kelly, S.L.; Rogers, P.D.; Moye-Rowley, W.S.; et al. Loss-of-Function ROX1 Mutations Suppress the Fluconazole Susceptibility of upc2AΔ Mutation in Candida glabrata, Implicating Additional Positive Regulators of Ergosterol Biosynthesis. mSphere 2021, 6, e00830-21. [Google Scholar] [CrossRef]
- Martel, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.G.S.; Kelly, D.E.; Kelly, S.L. A Clinical Isolate of Candida albicans with Mutations in ERG11 (Encoding Sterol 14α-Demethylase) and ERG5 (Encoding C22 Desaturase) Is Cross Resistant to Azoles and Amphotericin B. Antimicrob. Agents Chemother. 2010, 54, 3578–3583. [Google Scholar] [CrossRef]
- Kelly, S.L.; Lamb, D.C.; Kelly, D.E.; Manning, N.J.; Loeffler, J.; Hebart, H.; Schumacher, U.; Einsele, H. Resistance to Fluconazole and Cross-Resistance to Amphotericin B in Candida albicans from AIDS Patients Caused by Defective Sterol Δ5,6-Desaturation. FEBS Lett. 1997, 400, 80–82. [Google Scholar] [CrossRef]
- Nolte, F.S.; Parkinson, T.; Falconer, D.J.; Dix, S.; Williams, J.; Gilmore, C.; Geller, R.; Wingard, J.R. Isolation and Characterization of Fluconazole- and Amphotericin B-Resistant Candida albicans from Blood of Two Patients with Leukemia. Antimicrob. Agents Chemother. 1997, 41, 196–199. [Google Scholar] [CrossRef]
- Maphanga, T.G.; Naicker, S.D.; Kwenda, S.; Muñoz, J.F.; van Schalkwyk, E.; Wadula, J.; Nana, T.; Ismail, A.; Coetzee, J.; Govind, C.; et al. In Vitro Antifungal Resistance of Candida auris Isolates from Bloodstream Infections, South Africa. Antimicrob. Agents Chemother. 2021, 65, e00517-21. [Google Scholar] [CrossRef]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of Antifungal Drug Resistance. Cold Spring Harb. Perspect. Med. 2015, 5, a019752. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.E.; Soulard, A.; Hall, M.N. TOR Regulates Ribosomal Protein Gene Expression via PKA and the Forkhead Transcription Factor FHL1. Cell 2004, 119, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Alampalli, S.V.; Nageshan, R.K.; Chettiar, S.T.; Joshi, S.; Tatu, U.S. Draft Genome of a Commonly Misdiagnosed Multidrug Resistant Pathogen Candida auris. BMC Genom. 2015, 16, 686. [Google Scholar] [CrossRef] [PubMed]
- Zamith-Miranda, D.; Heyman, H.M.; Cleare, L.G.; Couvillion, S.P.; Clair, G.C.; Bredeweg, E.L.; Gacser, A.; Nimrichter, L.; Nakayasu, E.S.; Nosanchuk, J.D. Multi-Omics Signature of Candida auris, an Emerging and Multidrug-Resistant Pathogen. mSystems 2019, 4, e00257-19. [Google Scholar] [CrossRef]
- Kuranda, K.; Leberre, V.; Sokol, S.; Palamarczyk, G.; François, J. Investigating the Caffeine Effects in the Yeast Saccharomyces cerevisiae Brings New Insights into the Connection between TOR, PKC and Ras/cAMP Signalling Pathways. Mol. Microbiol. 2006, 61, 1147–1166. [Google Scholar] [CrossRef]
- Estrada, A.F.; Muruganandam, G.; Prescianotto-Baschong, C.; Spang, A. The ArfGAP2/3 Glo3 and Ergosterol Collaborate in Transport of a Subset of Cargoes. Biol. Open 2015, 4, 792–802. [Google Scholar] [CrossRef]
- Souza, C.M.; Schwabe, T.M.E.; Pichler, H.; Ploier, B.; Leitner, E.; Guan, X.L.; Wenk, M.R.; Riezman, I.; Riezman, H. A Stable Yeast Strain Efficiently Producing Cholesterol Instead of Ergosterol Is Functional for Tryptophan Uptake, but Not Weak Organic Acid Resistance. Metab. Eng. 2011, 13, 555–569. [Google Scholar] [CrossRef]
- Johnston, E.J.; Moses, T.; Rosser, S.J. The Wide-Ranging Phenotypes of Ergosterol Biosynthesis Mutants, and Implications for Microbial Cell Factories. Yeast 2020, 37, 27–44. [Google Scholar] [CrossRef]
- Misas, E.; Escandón, P.; McEwen, J.G.; Clay, O.K. The LUFS Domain, Its Transcriptional Regulator Proteins, and Drug Resistance in the Fungal Pathogen Candida auris. Protein Sci. 2019, 28, 2024–2029. [Google Scholar] [CrossRef]
- Chung, H.; Lee, Y.-H. Hypoxia: A Double-Edged Sword During Fungal Pathogenesis? Front. Microbiol. 2020, 11, 1920. [Google Scholar] [CrossRef]
- Lionakis, M.S.; Lim, J.K.; Lee, C.-C.R.; Murphy, P.M. Organ-Specific Innate Immune Responses in a Mouse Model of Invasive Candidiasis. J. Innate Immun. 2011, 3, 180–199. [Google Scholar] [CrossRef]
- Bruno, M.; Kersten, S.; Bain, J.M.; Jaeger, M.; Rosati, D.; Kruppa, M.D.; Lowman, D.W.; Rice, P.J.; Graves, B.; Ma, Z.; et al. Transcriptional and Functional Insights into the Host Immune Response against the Emerging Fungal Pathogen Candida auris. Nat. Microbiol. 2020, 5, 1516–1531. [Google Scholar] [CrossRef]
- Navarro-Arias, M.J.; Hernández-Chávez, M.J.; García-Carnero, L.C.; Amezcua-Hernández, D.G.; Lozoya-Pérez, N.E.; Estrada-Mata, E.; Martínez-Duncker, I.; Franco, B.; Mora-Montes, H.M. Differential Recognition of Candida tropicalis, Candida guilliermondii, Candida krusei, and Candida auris by Human Innate Immune Cells. Infect. Drug Resist. 2019, 12, 783–794. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC (μg/mL) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strains | AMB | FLU | POS | VOR | AND | CAS | MICA | |||||||
| 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | |
| C. auris 0381 | 0.25 | 1 | 8 | 16 | 0.0625 | 0.0625 | 0.0625 | 0.0625 | 0.03125 | 0.125 | 0.25 | 1 | 0.03125 | 0.03125 |
| C. auris 0381 FLUevo | 0.5 | 1 | 256 * | 512 * | 0.0625 | 0.25 ** | 2 ** | 4 ** | 0.0625 | 0.5 | 0.5 | 0.5 | 0.03125 | 0.03125 |
| C. auris 0381 POSevo | 0.5 | 0.5 | >512 * | >512 * | >32 ** | >32 ** | >32 ** | >32 ** | 0.125 | 0.5 | 0.25 | 0.25 | 0.03125 | 0.03125 |
| C. auris 0381 VORevo | 0.5 | 1 | >512 * | >512 * | 0.25 ** | 0.5 ** | 8 ** | 16 ** | 0.0625 | 0.125 | 0.25 | 0.25 | 0.03125 | 0.03125 |
| MIC (μg/mL) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strains | AMB | FLU | POS | VOR | AND | CAS | MICA | |||||||
| 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | |
| C. auris 0387 | 0.25 | 0.5 | 2 | 8 | 0.0625 | 0.0625 | 0.0625 | 0.0625 | 0.0625 | 0.125 | 0.5 | 1 | 0.03125 | 0.03125 |
| C. auris 0387 FLUevo | 0.25 | 0.5 | >512 * | >512 * | >32 ** | >32 ** | 16 ** | >32 ** | 0.5 | 1 | >16 * | >16 * | 0.03125 | 0.0625 |
| C. auris 0387 POSevo | 0.5 | 0.5 | >512 * | >512 * | >32 ** | >32 ** | >32 ** | >32 ** | 0.125 | 0.5 | 2 * | 4 * | 0.03125 | 0.03125 |
| C. auris 0387 VORevo | 0.5 | 0.5 | >512 * | >512 * | >32 ** | >32 ** | >32 ** | >32 ** | 0.25 | 0.5 | 2 * | 2 * | 0.03125 | 0.03125 |
| C. auris 0381 FLUevo | |||||
| Type of mutation | Gene | C. albicans ortholog | S. cerevisiae ortholog | Amino Acid Substitution | Function |
| Nonsynonymous SNP | B9J08_004820 (TAC1b) | HAL9 | HAL9 | N690S | Putative transcriptional activator of the drug-responsive genes CDR1 and CDR2; gain-of-function mutations are associated with resistance to fluconazole |
| C. auris 0381 POSevo | |||||
| Type of mutation | Gene | C. albicans ortholog | S. cerevisiae ortholog | Amino Acid Substitution | Function |
| Deletion (200bp) | B9J08_004089 | RFG1 | ROX1 | - | Ortholog(s) has DNA binding, bending, DNA-binding transcription repressor activity, RNA polymerase II-specific, cis-regulatory region sequence-specific DNA binding, sequence-specific DNA binding, transcription corepressor activity |
| C. auris 0381 VORevo | |||||
| Type of mutation | Gene | C. albicans ortholog | S. cerevisiae ortholog | Amino Acid Substitution | Function |
| Nonsynonymous SNP | B9J08_004820 (TAC1b) | HAL9 | HAL9 | S19I | Putative transcriptional activator of the drug-responsive genes CDR1 and CDR2; gain-of-function mutations are associated with resistance to fluconazole |
| Duplication | CJI96_0001194-CJI96_0001248 * | - | |||
| C. auris 0387 FLUevo | |||||
| Type of mutation | Gene | C. albicans ortholog | S. cerevisiae ortholog | Amino Acid Substitution | Function |
| Nonsynonymous SNP | B9J08_002818 | BCY1 | BCY1 | R160S | Ortholog(s) have cAMP binding, cAMP-dependent protein kinase inhibitor activity, cAMP-dependent protein kinase regulator activity |
| Nonsynonymous SNP | B9J08_002259 | HGT4 | SNF3 | P95T | Ortholog(s) have role in detection of glucose, fructose transmembrane transport, glucose-mediated signaling pathway, glucose transmembrane transport, mannose transmembrane transport, negative regulation of meiotic nuclear division |
| C. auris 0387 POSevo | |||||
| Type of mutation | Gene | C. albicans ortholog | S. cerevisiae ortholog | Amino Acid Substitution | Function |
| Nonsynonymous SNP | B9J08_002818 | BCY1 | BCY1 | R160S | Ortholog(s) have cAMP binding, cAMP-dependent protein kinase inhibitor activity, cAMP-dependent protein kinase regulator activity |
| Nonsynonymous SNP | B9J08_003737 | ERG3 | ERG3 | S316* | Ortholog(s) have C-5 sterol desaturase activity, role in ergosterol biosynthetic process and endoplasmic reticulum lumen localization |
| C. auris 0387 VORevo | |||||
| Type of mutation | Gene | C. albicans ortholog | S. cerevisiae ortholog | Amino Acid Substitution | Function |
| Nonsynonymous SNP | B9J08_002818 | BCY1 | BCY1 | R160S | Ortholog(s) have cAMP binding, cAMP-dependent protein kinase inhibitor activity, cAMP-dependent protein kinase regulator activity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bohner, F.; Papp, C.; Takacs, T.; Varga, M.; Szekeres, A.; Nosanchuk, J.D.; Vágvölgyi, C.; Tóth, R.; Gacser, A. Acquired Triazole Resistance Alters Pathogenicity-Associated Features in Candida auris in an Isolate-Dependent Manner. J. Fungi 2023, 9, 1148. https://doi.org/10.3390/jof9121148
Bohner F, Papp C, Takacs T, Varga M, Szekeres A, Nosanchuk JD, Vágvölgyi C, Tóth R, Gacser A. Acquired Triazole Resistance Alters Pathogenicity-Associated Features in Candida auris in an Isolate-Dependent Manner. Journal of Fungi. 2023; 9(12):1148. https://doi.org/10.3390/jof9121148
Chicago/Turabian StyleBohner, Flora, Csaba Papp, Tamas Takacs, Mónika Varga, András Szekeres, Joshua D. Nosanchuk, Csaba Vágvölgyi, Renáta Tóth, and Attila Gacser. 2023. "Acquired Triazole Resistance Alters Pathogenicity-Associated Features in Candida auris in an Isolate-Dependent Manner" Journal of Fungi 9, no. 12: 1148. https://doi.org/10.3390/jof9121148
APA StyleBohner, F., Papp, C., Takacs, T., Varga, M., Szekeres, A., Nosanchuk, J. D., Vágvölgyi, C., Tóth, R., & Gacser, A. (2023). Acquired Triazole Resistance Alters Pathogenicity-Associated Features in Candida auris in an Isolate-Dependent Manner. Journal of Fungi, 9(12), 1148. https://doi.org/10.3390/jof9121148