
An Evolved Strain of the Oleaginous Yeast Rhodotorula toruloides, Multi-Tolerant to the Major Inhibitors Present in Lignocellulosic Hydrolysates, Exhibits an Altered Cell Envelope

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Culture Media and Culture Conditions
2.3. Isolation of a Multi-Tolerant Strain, Derived from R. toruloides IST536, by Adaptive Laboratory Evolution (ALE)
2.4. Determination of the Concentration of Sugars and Organic Acids
2.5. Assessment of Lipid Production by Nile Red Staining
2.6. Cell Lipid Content and Lipid Profile by Gas Chromatography
2.7. Comparison of Yeast Tolerance to Inhibitors Present in the Lignocellulosic Hydrolysates
2.8. Yeast Cell Wall Susceptibility to Zymolyase
2.9. Assessment of Yeast Plasma Membrane Permeability Based on Propidium Iodide Fluorescent Probe
3. Results
3.1. Adaptive Laboratory Evolution (ALE) of IST 536
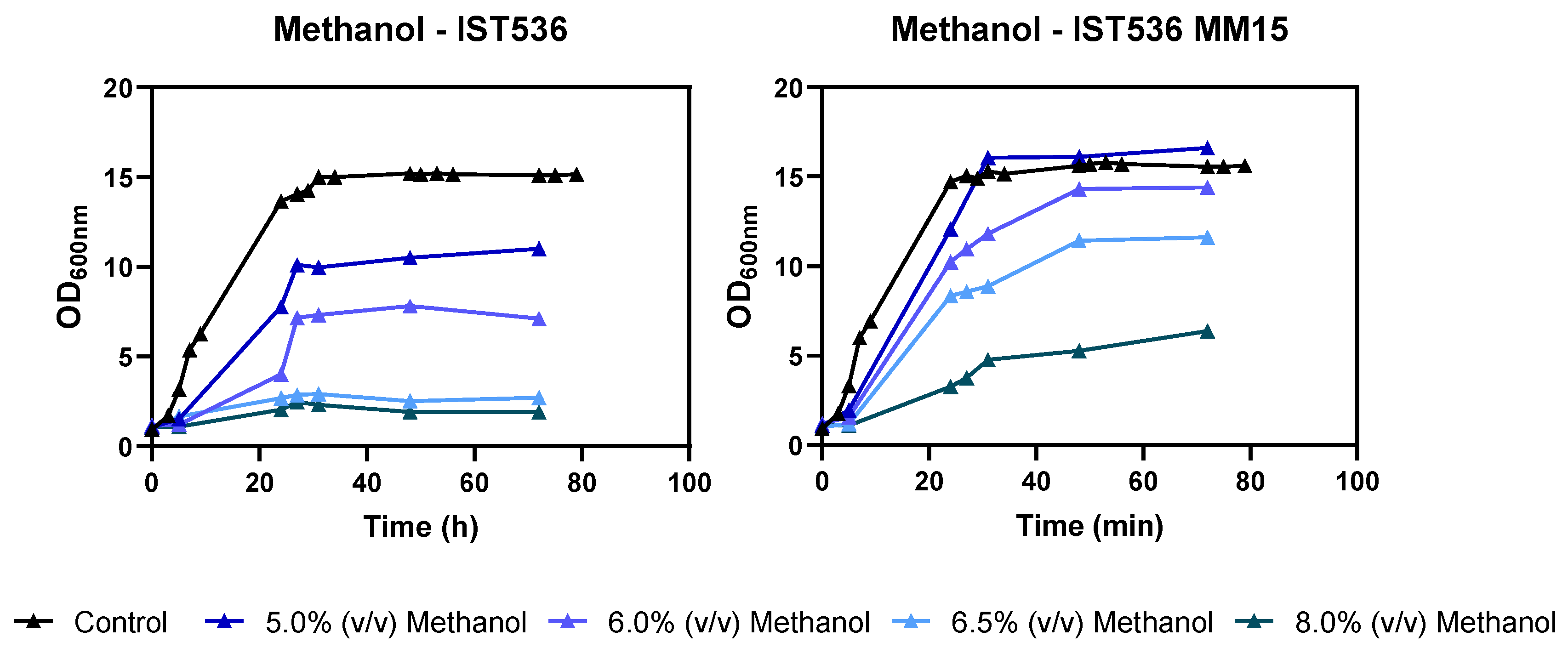
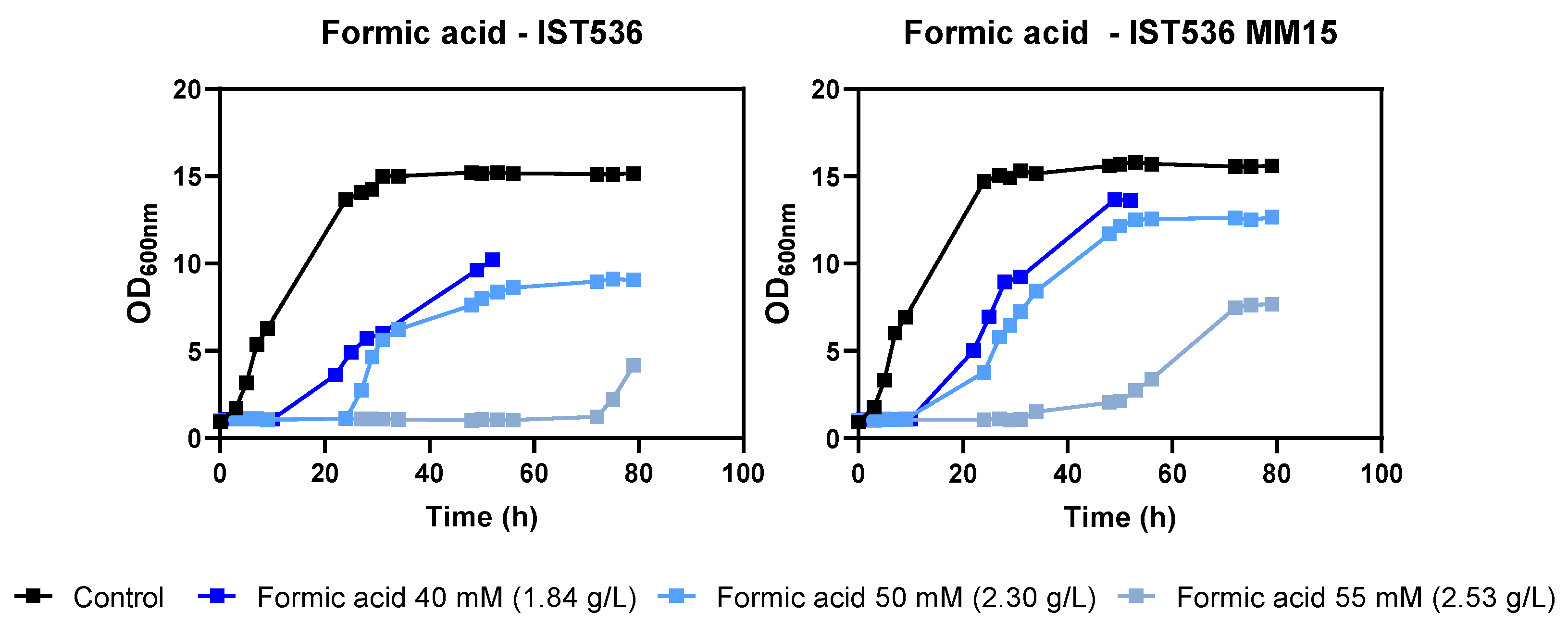
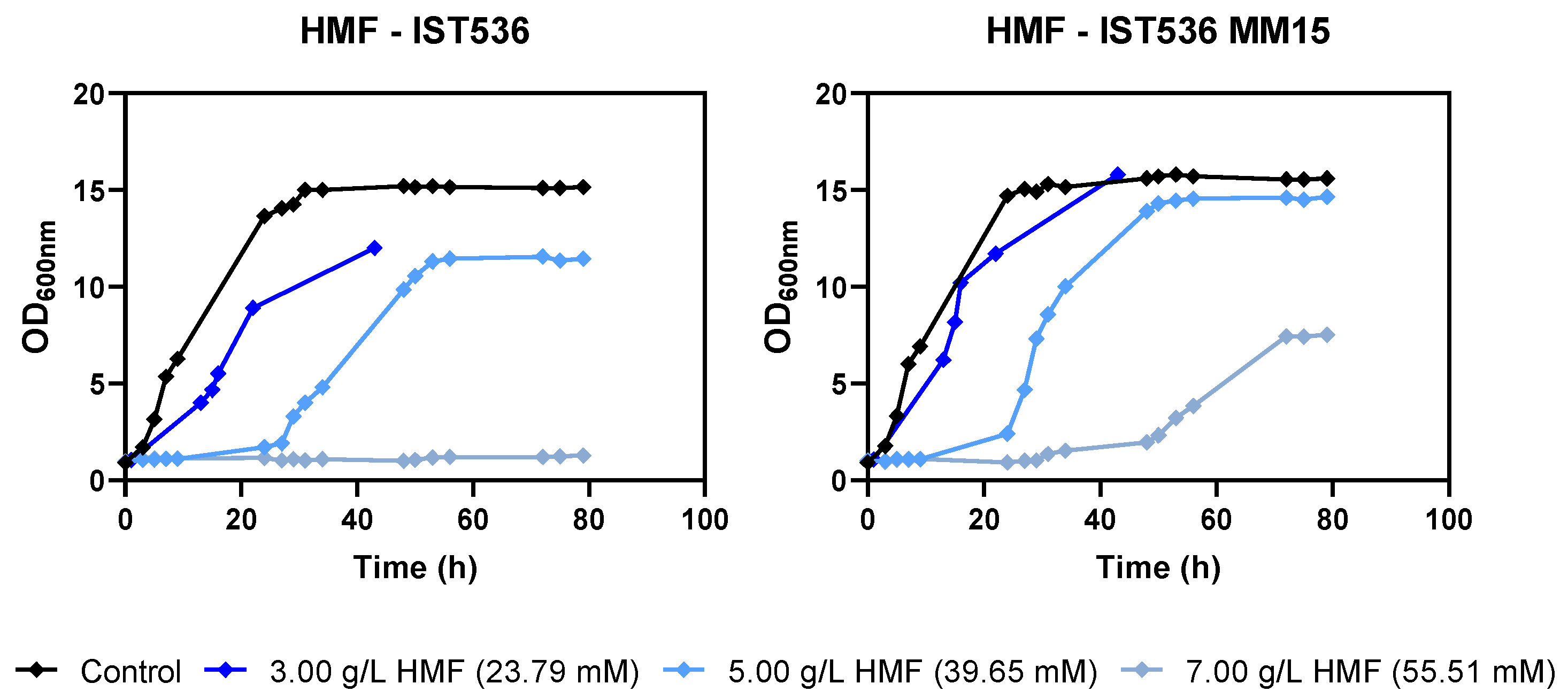
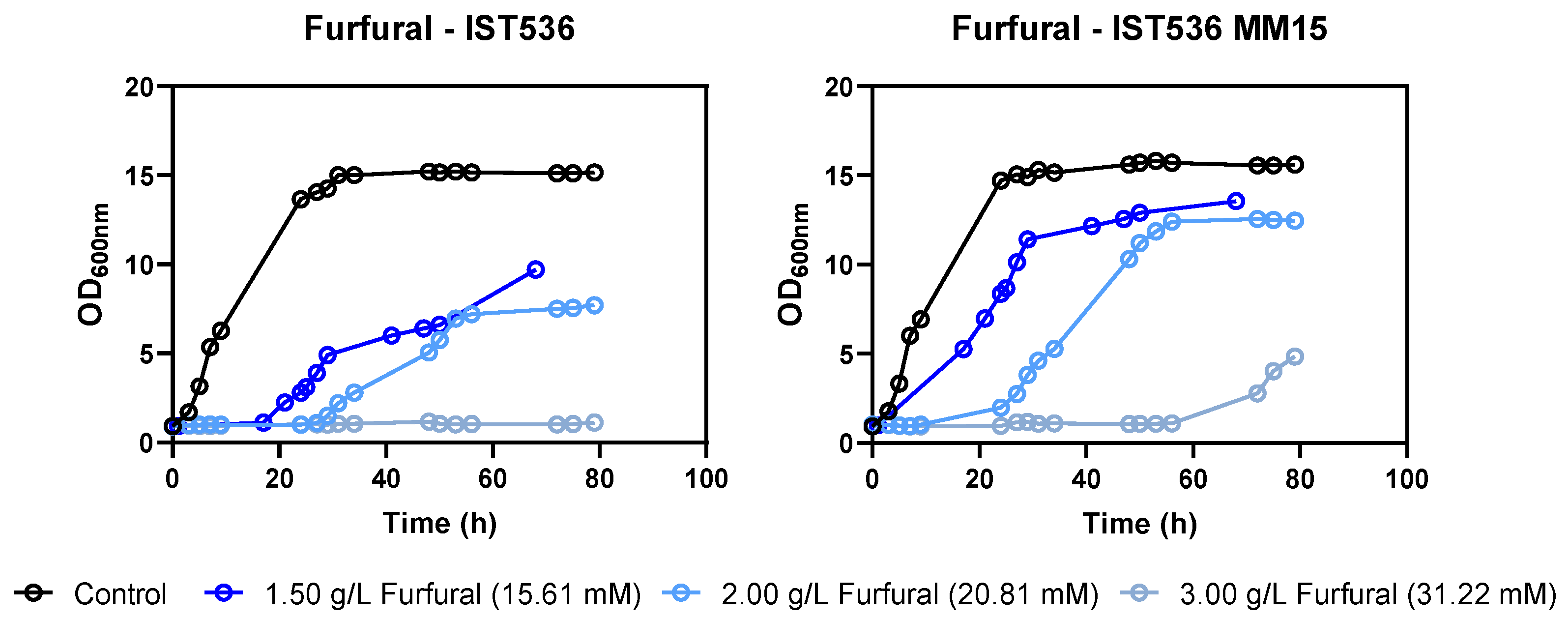
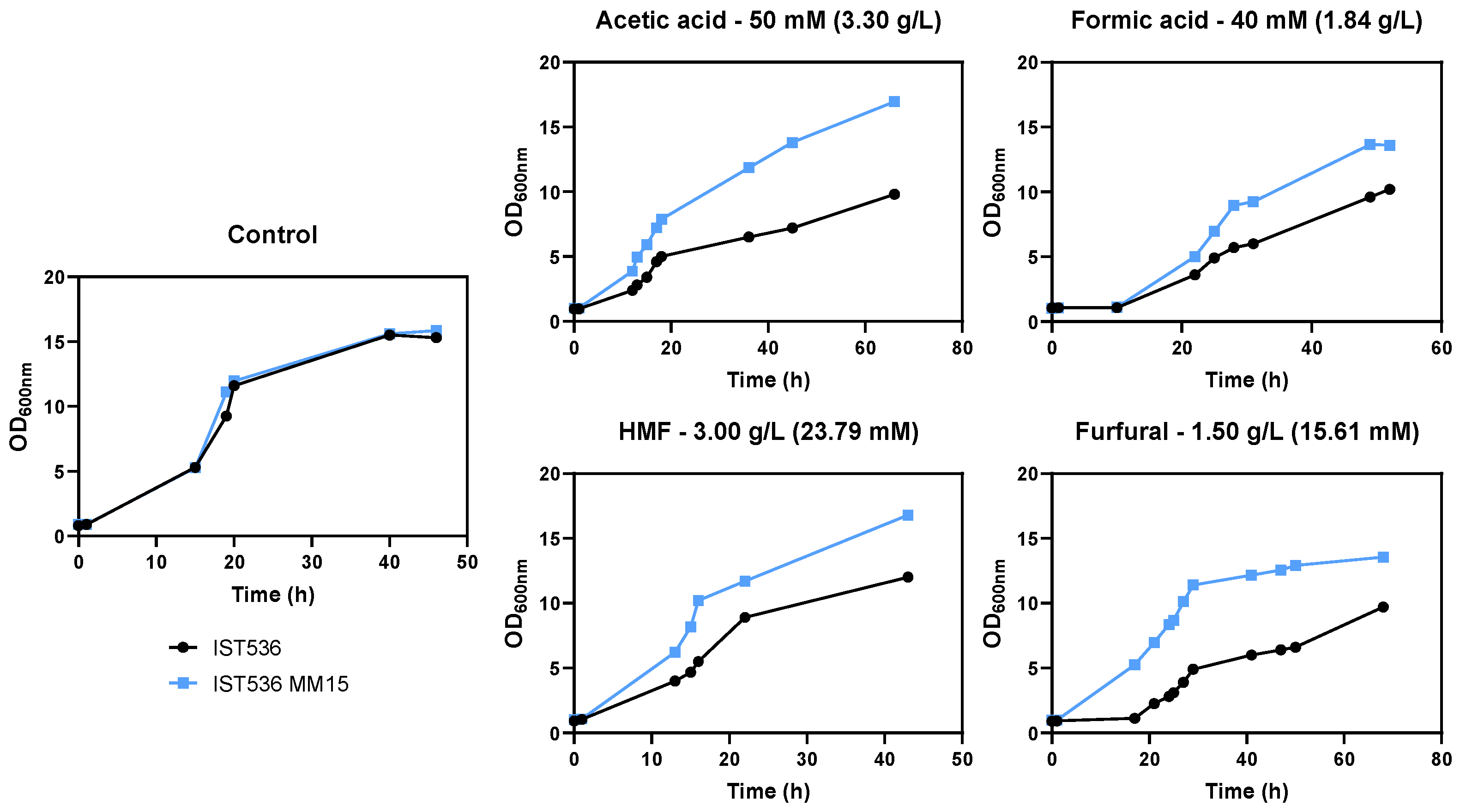
3.2. The Evolved Strain R. toruloides IST536 MM15 Is More Tolerant to Multiple Inhibitors, Compared with the Original Strain
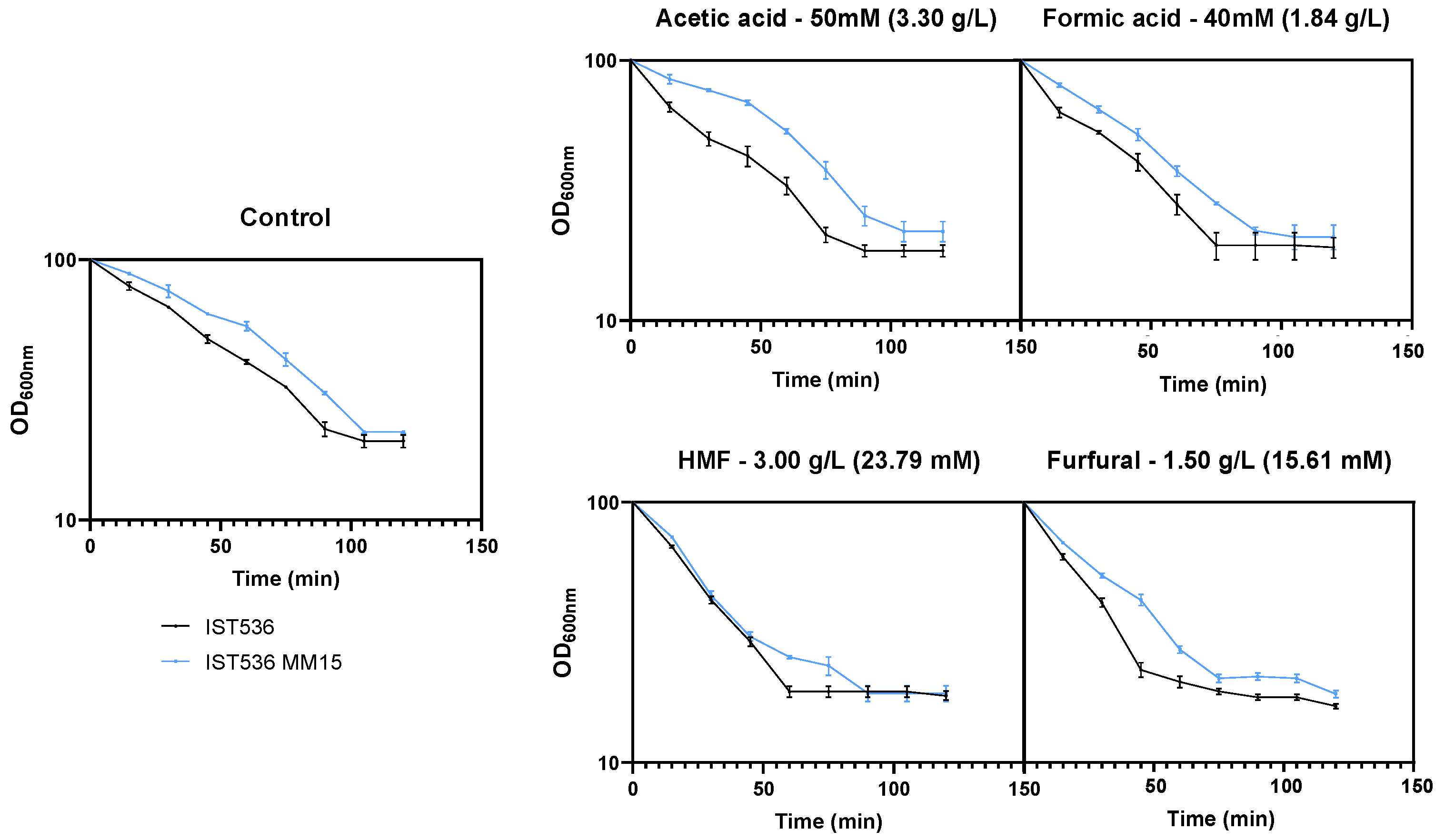
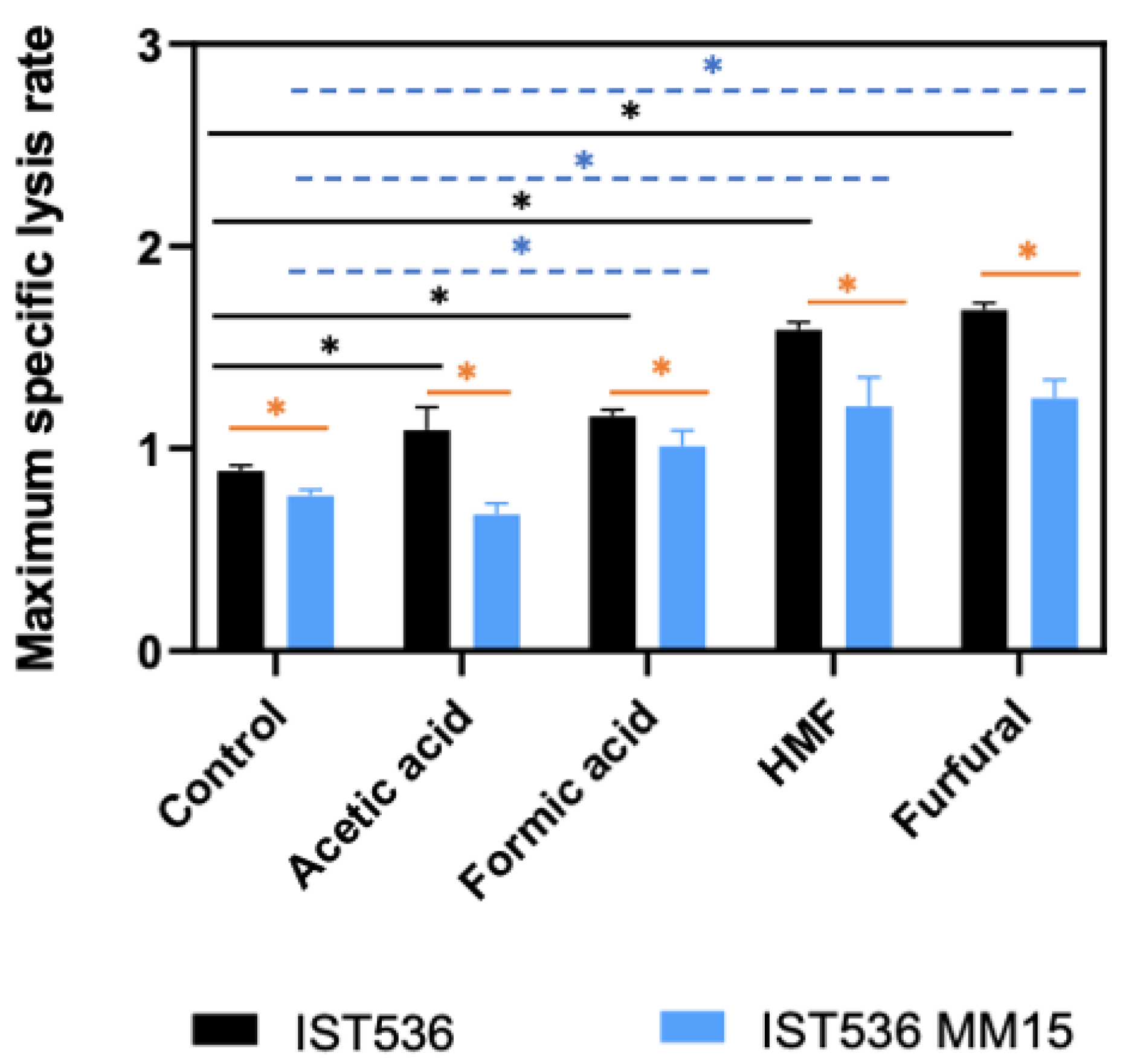
3.3. The Susceptibility of the Evolved Multi-Tolerant R. toruloides Strain to Zymolyase Activity Is below the Original Strain Susceptibility in Absence or Presence of Inhibitors
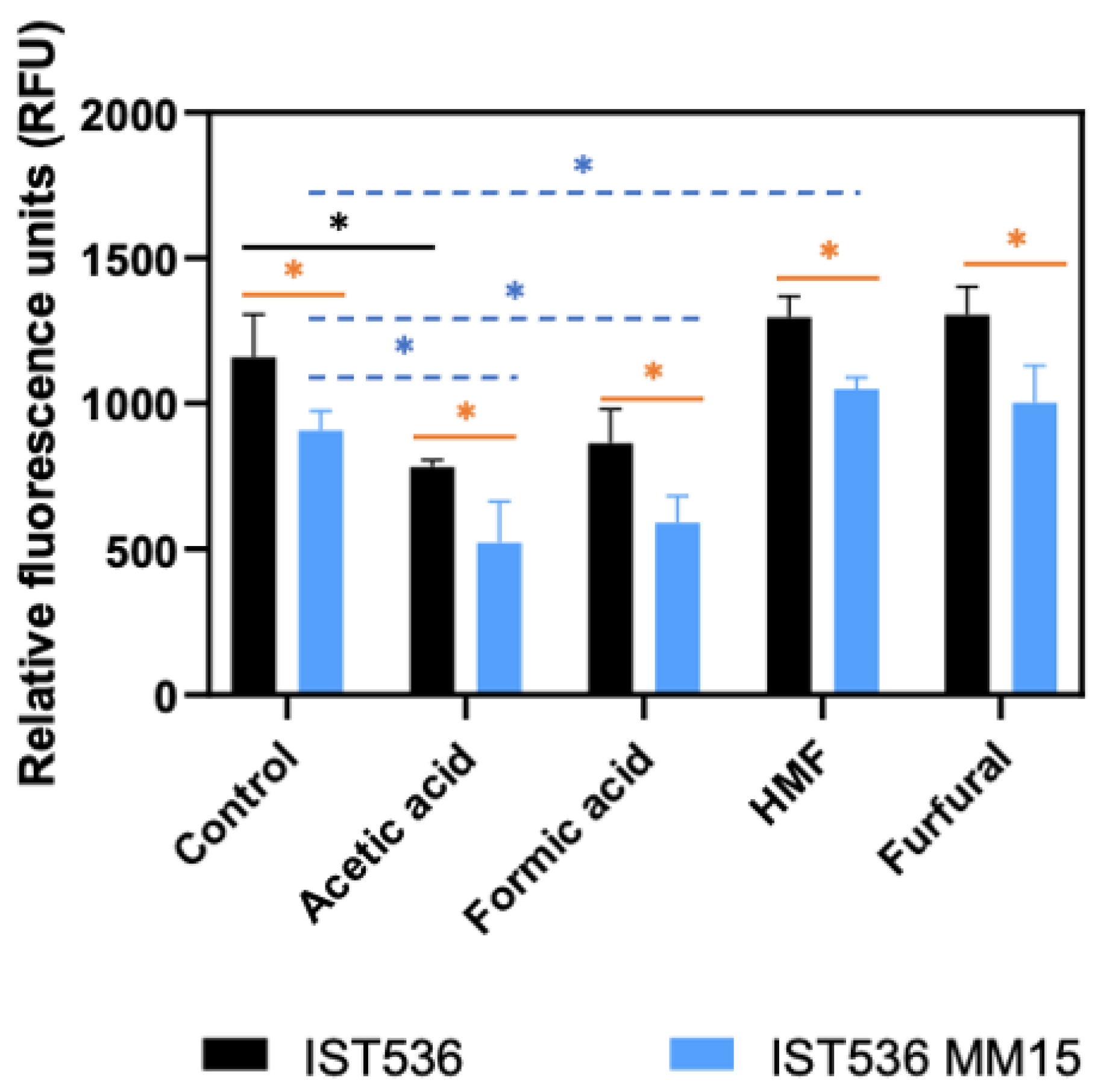
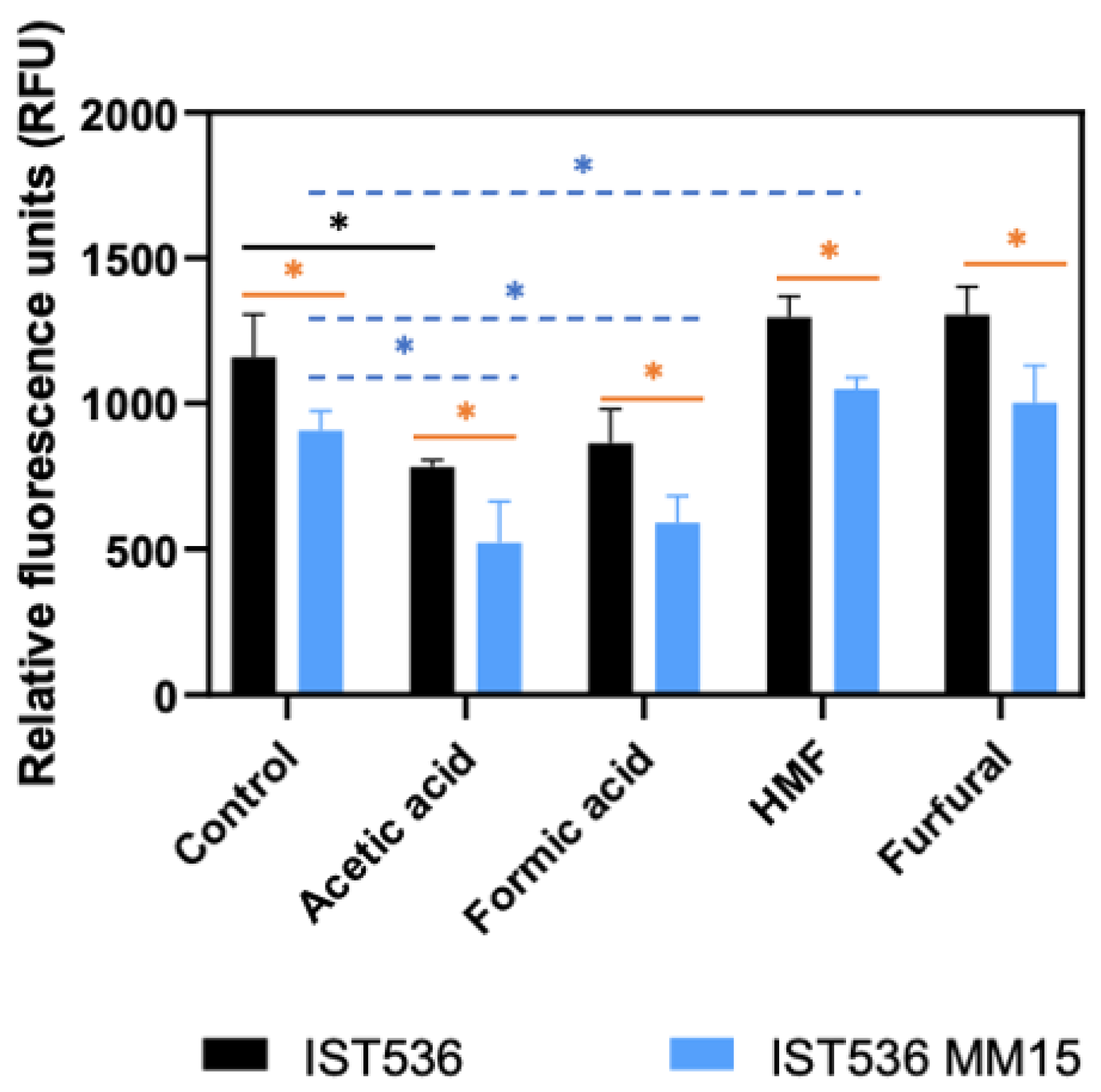
3.4. The Evolved Multi-Tolerant R. toruloides Strain Permeability to Propidium Iodide, Is below the Original Strain Permeability in Absence or Presence of Inhibitors
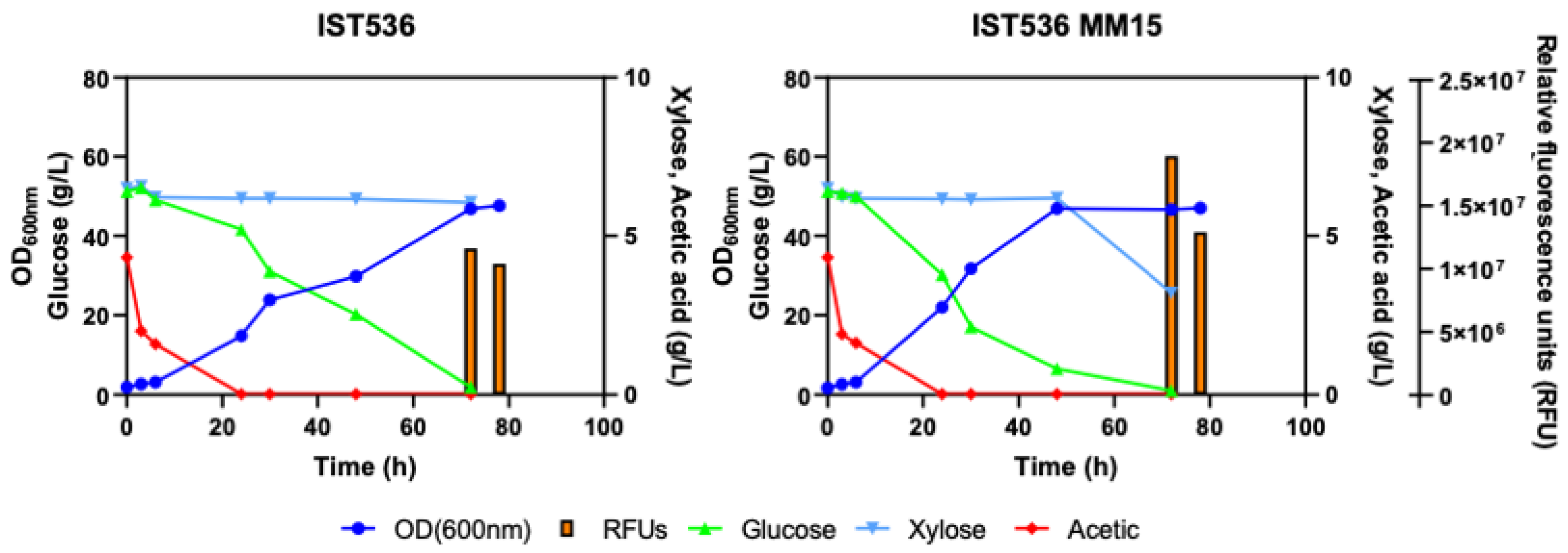
3.5. The Evolved R. toruloides Strain Performed Better in Synthetic Medium Simulating a Lignocellulosic Hydrolysate, Compared with the Original Strain
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cunha, J.T.; Romaní, A.; Costa, C.E.; Sá-Correia, I.; Domingues, L. Molecular and Physiological Basis of Saccharomyces cerevisiae Tolerance to Adverse Lignocellulose-Based Process Conditions. Appl. Microbiol. Biotechnol. 2019, 103, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Fels, M.; Dragone, G.; Mussatto, S.I. Effects of Inhibitory Compounds Derived from Lignocellulosic Biomass on the Growth of the Wild-Type and Evolved Oleaginous Yeast Rhodosporidium toruloides. Ind. Crops Prod. 2021, 170, 113799. [Google Scholar] [CrossRef]
- Mota, M.N.; Múgica, P.; Sá-Correia, I. Exploring Yeast Diversity to Produce Lipid-Based Biofuels from Agro-Forestry and Industrial Organic Residues. J. Fungi 2022, 8, 687. [Google Scholar] [CrossRef] [PubMed]
- de Melo e Silva, J.; da Silva Martins, L.H.; Moreira, D.K.T.; do Prado Silva, L.; de Paula Menezes Barbosa, P.; Komesu, A.; Ferreira, N.R.; de Oliveira, J.A.R. Microbial Lipid Based Biorefinery Concepts: A Review of Status and Prospects. Foods 2023, 12, 2074. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, Z.; Bai, F. High-Density Cultivation of Oleaginous Yeast Rhodosporidium toruloides Y4 in Fed-Batch Culture. Enzyme Microb. Technol. 2007, 41, 312–317. [Google Scholar] [CrossRef]
- Hu, C.; Zhao, X.; Zhao, J.; Wu, S.; Zhao, Z.K. Effects of Biomass Hydrolysis By-Products on Oleaginous Yeast Rhodosporidium toruloides. Bioresour. Technol. 2009, 100, 4843–4847. [Google Scholar] [CrossRef] [PubMed]
- Chaiyaso, T.; Manowattana, A.; Techapun, C.; Watanabe, M. Efficient Bioconversion of Enzymatic Corncob Hydrolysate into Biomass and Lipids by Oleaginous Yeast Rhodosporidium paludigenum KM281510. Prep. Biochem. Biotechnol. 2019, 49, 545–556. [Google Scholar] [CrossRef]
- Beopoulos, A.; Nicaud, J.-M.; Gaillardin, C. An Overview of Lipid Metabolism in Yeasts and Its Impact on Biotechnological Processes. Appl. Microbiol. Biotechnol. 2011, 90, 1193–1206. [Google Scholar] [CrossRef]
- Caporusso, A.; Capece, A.; De Bari, I. Oleaginous Yeasts as Cell Factories for the Sustainable Production of Microbial Lipids by the Valorization of Agri-Food Wastes. Fermentation 2021, 7, 50. [Google Scholar] [CrossRef]
- Saha, R.; Mukhopadhyay, M. Prospect of Metabolic Engineering in Enhanced Microbial Lipid Production: Review. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Santek, M.I.; Beluhan, S.; Santek, B. Production of Microbial Lipids from Lignocellulosic Biomass. In Advances in Biofuels and Bioenergy; InTech: London, UK, 2018; pp. 137–164. Available online: https://www.intechopen.com/chapters/59357 (accessed on 30 October 2023). [CrossRef]
- Sjulander, N.; Kikas, T. Origin, Impact and Control of Lignocellulosic Inhibitors in Bioethanol Production—A Review. Energies 2020, 13, 4751. [Google Scholar] [CrossRef]
- Valdés, G.; Mendonça, R.T.; Aggelis, G. Lignocellulosic Biomass as a Substrate for Oleaginous Microorganisms: A Review. Appl. Sci. 2020, 10, 7698. [Google Scholar] [CrossRef]
- Guo, H.; Zhao, Y.; Chang, J.S.; Lee, D.J. Inhibitor Formation and Detoxification during Lignocellulose Biorefinery: A Review. Bioresour. Technol. 2022, 361, 127666. [Google Scholar] [CrossRef] [PubMed]
- Vanmarcke, G.; Demeke, M.M.; Foulquié-Moreno, M.R.; Thevelein, J.M. Identification of the Major Fermentation Inhibitors of Recombinant 2G Yeasts in Diverse Lignocellulose Hydrolysates. Biotechnol. Biofuels 2021, 14, 92. [Google Scholar] [CrossRef]
- Li, B.; Liu, N.; Zhao, X. Response Mechanisms of Saccharomyces cerevisiae to the Stress Factors Present in Lignocellulose Hydrolysate and Strategies for Constructing Robust Strains. Biotechnol. Biofuels Bioprod. 2022, 15, 28. [Google Scholar] [CrossRef]
- Du, C.; Li, Y.; Xiang, R.; Yuan, W. Formate Dehydrogenase Improves the Resistance to Formic Acid and Acetic Acid Simultaneously in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2022, 23, 3406. [Google Scholar] [CrossRef]
- Sharma, H.K.; Xu, C.; Qin, W. Biological Pretreatment of Lignocellulosic Biomass for Biofuels and Bioproducts: An Overview. Waste Biomass Valorization 2019, 10, 235–251. [Google Scholar] [CrossRef]
- Piotrowski, J.S.; Zhang, Y.; Bates, D.M.; Keating, D.H.; Sato, T.K.; Ong, I.M.; Landick, R. Death by a Thousand Cuts: The Challenges and Diverse Landscape of Lignocellulosic Hydrolysate Inhibitors. Front. Microbiol. 2014, 5, 90. [Google Scholar] [CrossRef]
- Park, G.W.; Shin, S.; Kim, S.J.; Lee, J.-S.; Moon, M.; Min, K. Rice Straw-Derived Lipid Production by HMF/Furfural-Tolerant Oleaginous Yeast Generated by Adaptive Laboratory Evolution. Bioresour. Technol. 2023, 367, 128220. [Google Scholar] [CrossRef]
- Brandenburg, J.; Blomqvist, J.; Shapaval, V.; Kohler, A.; Sampels, S.; Sandgren, M.; Passoth, V. Oleaginous Yeasts Respond Differently to Carbon Sources Present in Lignocellulose Hydrolysate. Biotechnol. Biofuels 2021, 14, 124. [Google Scholar] [CrossRef]
- Wen, Z.; Zhang, S.; Odoh, C.K.; Jin, M.; Zhao, Z.K. Rhodosporidium toruloides—A Potential Red Yeast Chassis for Lipids and Beyond. FEMS Yeast Res. 2020, 20, foaa038. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.C.; Palma, M.; Angelov, A.; Nevoigt, E.; Liebl, W.; Sá-Correia, I. Complete Utilization of the Major Carbon Sources Present in Sugar Beet Pulp Hydrolysates by the Oleaginous Red Yeasts Rhodotorula toruloides and R. mucilaginosa. J. Fungi 2021, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-C.; Honda, K.; Kazuhito, F. Current Advances in Alteration of Fatty Acid Profile in Rhodotorula toruloides: A Mini-Review. World J. Microbiol. Biotechnol. 2023, 39, 234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Peng, J.; Zhao, H.; Shi, S. Engineering Oleaginous Yeast Rhodotorula toruloides for Overproduction of Fatty Acid Ethyl Esters. Biotechnol. Biofuels 2021, 14, 115. [Google Scholar] [CrossRef]
- Sandberg, T.E.; Salazar, M.J.; Weng, L.L.; Palsson, B.O.; Feist, A.M. The Emergence of Adaptive Laboratory Evolution as an Efficient Tool for Biological Discovery and Industrial Biotechnology. Metab. Eng. 2019, 56, 1–16. [Google Scholar] [CrossRef]
- Arora, N.; Yen, H.W.; Philippidis, G.P. Harnessing the Power of Mutagenesis and Adaptive Laboratory Evolution for High Lipid Production by Oleaginous Microalgae and Yeasts. Sustainability 2020, 12, 5125. [Google Scholar] [CrossRef]
- Morales-Palomo, S.; Liras, M.; González-Fernández, C.; Tomás-Pejó, E. Key Role of Fluorescence Quantum Yield in Nile Red Staining Method for Determining Intracellular Lipids in Yeast Strains. Biotechnol. Biofuels Bioprod. 2022, 15, 37. [Google Scholar] [CrossRef]
- Ribeiro, R.A.; Godinho, C.P.; Vitorino, M.V.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Crosstalk between Yeast Cell Plasma Membrane Ergosterol Content and Cell Wall Stiffness under Acetic Acid Stress Involving Pdr18. J. Fungi 2022, 8, 103. [Google Scholar] [CrossRef]
- Sah, S.K.; Bhattacharya, S.; Yadav, A.; Husain, F.; Ndiaye, A.B.K.T.; Kruppa, M.D.; Hayes, J.J.; Rustchenko, E. Multiple Genes of Candida albicans Influencing Echinocandin Susceptibility in Caspofungin-Adapted Mutants. Antimicrob. Agents Chemother. 2022, 66, e00977-22. [Google Scholar] [CrossRef]
- Davey, H.M.; Hexley, P. Red but Not Dead? Membranes of Stressed Saccharomyces cerevisiae Are Permeable to Propidium Iodide. Environ. Microbiol. 2011, 13, 163–171. [Google Scholar] [CrossRef]
- Konzock, O.; Zaghen, S.; Norbeck, J. Tolerance of Yarrowia lipolytica to Inhibitors Commonly Found in Lignocellulosic Hydrolysates. BMC Microbiol. 2021, 21, 77. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Bourbon-Melo, N.; Sá-Correia, I. The Cell Wall and the Response and Tolerance to Stresses of Biotechnological Relevance in Yeasts. Front. Microbiol. 2022, 13, 953479. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Li, N.; Lv, S. Iron Oxide Magnetic Nanoparticles Exhibiting Zymolyase-like Lytic Activity. Chem. Eng. J. 2020, 394, 125000. [Google Scholar] [CrossRef]
- Rodríguez-Peña, J.M.; Díez-Muñiz, S.; Bermejo, C.; Nombela, C.; Arroyo, J. Activation of the Yeast Cell Wall Integrity MAPK Pathway by Zymolyase Depends on Protease and Glucanase Activities and Requires the Mucin-like Protein Hkr1 but Not Msb2. FEBS Lett. 2013, 587, 3675–3680. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Wu, S.; Liu, L. Enzymatic Saccharification of Dilute Acid Pretreated Eucalyptus Chips for Fermentable Sugar Production. Bioresour. Technol. 2012, 110, 302–307. [Google Scholar] [CrossRef]
- Rigual, V.; Santos, T.M.; Domínguez, J.C.; Alonso, M.V.; Oliet, M.; Rodriguez, F. Evaluation of Hardwood and Softwood Fractionation Using Autohydrolysis and Ionic Liquid Microwave Pretreatment. Biomass Bioenergy 2018, 117, 190–197. [Google Scholar] [CrossRef]
- Kraisintu, P.; Yongmanitchai, W.; Limtong, S. Selection and Optimization for Lipid Production of a Newly Isolated Oleaginous Yeast, Rhodosporidium toruloides DMKU3-TK16. Agric. Nat. Resour. 2010, 44, 436–445. [Google Scholar]
- Palma, M.; Guerreiro, J.F.; Sá-Correia, I. Adaptive Response and Tolerance to Acetic Acid in Saccharomyces cerevisiae and Zygosaccharomyces bailii: A Physiological Genomics Perspective. Front. Microbiol. 2018, 9, 274. [Google Scholar] [CrossRef]
- Sargeant, L.A.; Mardell, M.; Saad-Allah, K.M.; Hussein, A.H.; Whiffin, F.; Santomauro, F.; Chuck, C.J. Production of Lipid from Depolymerised Lignocellulose Using the Biocontrol Yeast, Rhodotorula minuta: The Fatty Acid Profile Remains Stable Irrespective of Environmental Conditions. Eur. J. Lipid Sci. Technol. 2016, 118, 777–787. [Google Scholar] [CrossRef]
- Zhang, G.; French, W.T.; Hernandez, R.; Alley, E.; Paraschivescu, M. Effects of Furfural and Acetic Acid on Growth and Lipid Production from Glucose and Xylose by Rhodotorula glutinis. Biomass Bioenergy 2011, 35, 734–740. [Google Scholar] [CrossRef]
- Fernandes, T.; Osório, C.; Sousa, M.J.; Franco-Duarte, R. Contributions of Adaptive Laboratory Evolution towards the Enhancement of the Biotechnological Potential of Non-Conventional Yeast Species. J. Fungi 2023, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Menegon, Y.A.; Gross, J.; Jacobus, A.P. How Adaptive Laboratory Evolution Can Boost Yeast Tolerance to Lignocellulosic Hydrolyses. Curr. Genet. 2022, 68, 319–342. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Zhang, J.; Zhou, J.; Fang, Z.; Zhu, Z.; Li, J.; Du, G. Stress Tolerance Phenotype of Industrial Yeast: Industrial Cases, Cellular Changes, and Improvement Strategies. Appl. Microbiol. Biotechnol. 2019, 103, 6449–6462. [Google Scholar] [CrossRef] [PubMed]
- Ndukwe, J.K.; Aliyu, G.O.; Onwosi, C.O.; Chukwu, K.O.; Ezugworie, F.N. Mechanisms of Weak Acid-Induced Stress Tolerance in Yeasts: Prospects for Improved Bioethanol Production from Lignocellulosic Biomass. Process Biochem. 2020, 90, 118–130. [Google Scholar] [CrossRef]
- Dague, E.; Bitar, R.; Ranchon, H.; Durand, F.; Yken, H.M.; François, J.M. An Atomic Force Microscopy Analysis of Yeast Mutants Defective in Cell Wall Architecture. Yeast 2010, 27, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Vitorino, M.V.; Godinho, C.P.; Bourbon-Melo, N.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Yeast Adaptive Response to Acetic Acid Stress Involves Structural Alterations and Increased Stiffness of the Cell Wall. Sci. Rep. 2021, 11, 12652. [Google Scholar] [CrossRef]
- Schiavone, M.; Formosa-Dague, C.; Elsztein, C.; Teste, M.A.; Martin-Yken, H.; De Morais, M.A.; Dague, E.; François, J.M. Evidence for a Role for the Plasma Membrane in the Nanomechanical Properties of the Cell Wall as Revealed by an Atomic Force Microscopy Study of the Response of Saccharomyces cerevisiae to Ethanol Stress. Appl. Environ. Microbiol. 2016, 82, 4789–4801. [Google Scholar] [CrossRef]
- Pillet, F.; Lemonier, S.; Schiavone, M.; Formosa, C.; Martin-Yken, H.; Francois, J.M.; Dague, E. Uncovering by Atomic Force Microscopy of an Original Circular Structure at the Yeast Cell Surface in Response to Heat Shock. BMC Biol. 2014, 12, 6. [Google Scholar] [CrossRef]
- Kitichantaropas, Y.; Boonchird, C.; Sugiyama, M.; Kaneko, Y.; Harashima, S.; Auesukaree, C. Cellular Mechanisms Contributing to Multiple Stress Tolerance in Saccharomyces cerevisiae Strains with Potential Use in High-Temperature Ethanol Fermentation. AMB Express 2016, 6, 107. [Google Scholar] [CrossRef]
- Abrha, G.T.; Li, Q.; Kuang, X.; Xiao, D.; Ayepa, E.; Wu, J.; Chen, H.; Zhang, Z.; Liu, Y.; Yu, X.; et al. Contribution of YPRO15C Overexpression to the Resistance of Saccharomyces cerevisiae BY4742 Strain to Furfural Inhibitor. Polish J. Microbiol. 2023, 72, 177–186. [Google Scholar] [CrossRef]
- Godinho, C.P.; Prata, C.S.; Pinto, S.N.; Cardoso, C.; Bandarra, N.M.; Fernandes, F.; Sá-Correia, I. Pdr18 Is Involved in Yeast Response to Acetic Acid Stress Counteracting the Decrease of Plasma Membrane Ergosterol Content and Order. Sci. Rep. 2018, 8, 7860. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, L.; Genheden, S.; Eriksson, L.A.; Olsson, L.; Bettiga, M. Sphingolipids Contribute to Acetic Acid Resistance in Zygosaccharomyces bailii. Biotechnol. Bioeng. 2016, 113, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, L.; Santos, A.X.S.; Riezman, H.; Olsson, L.; Bettiga, M. Lipidomic Profiling of Saccharomyces cerevisiae and Zygosaccharomyces bailii Reveals Critical Changes in Lipid Composition in Response to Acetic Acid Stress. PLoS ONE 2013, 8, e73936. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Sampaio, J.L. Membrane Organization and Lipid Rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, a004697. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Zhu, Y.; Peng, Y.; Liang, X.; Liu, X.; Shao, L.; Xu, Y.; Xu, Z.; Liu, R.; Li, J. Physiological Mechanism of Improved Tolerance of Saccharomyces cerevisiae to Lignin-Derived Phenolic Acids in Lignocellulosic Ethanol Fermentation by Short-Term Adaptation. Biotechnol. Biofuels 2019, 12, 268. [Google Scholar] [CrossRef] [PubMed]
- Kłosowski, G.; Mikulski, D. Impact of Lignocellulose Pretreatment By-Products on S. cerevisiae Strain Ethanol Red Metabolism during Aerobic and an-Aerobic Growth. Molecules 2021, 26, 806. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation and Short Description | Composition |
|---|---|
| Minimal medium with glucose (MM) Glycerol (MMGly, for ALE experiments) | 6.7 g/L YNB (Difco, Michigan, MI, USA) with: 20.0 g/L glucose (NZYTech, Lisbon, Portugal) (MM) or 50.0 g/L of glycerol (Scharlau, Barcelona, Spain) (MMGly) pH adjusted to 4.5 |
| Synthetic medium simulating lignocellulosic hydrolysate (SH) | 1.3 g/L of YNB without ammonium sulfate and amino acids (Difco); supplemented with: 1 g/L de (NH4)2SO4 (Panreac, Chicago, IL, USA); amino acids (20 mg/L of L-histidine, 40 mg/L DL-methionine and 40 mg/L DL-tryptophan, all from Sigma, Darmstadt, Germany); 60.0 g/L glucose (NZYTech, Lisbon, Portugal); 7.0 g/L xylose (Sigma-Aldrich, Darmstadt, Germany); 5.0 g/L of acetic acid (Fluka, Buchs, Switzerland); 0.7 g/L of formic acid (Fluka, Buchs, Switzerland); 0.4 g/L of furfural (Sigma-Aldrich, Darmstadt, Germany); 1.0 g/L of HMF (Sigma-Aldrich, Darmstadt, Germany). pH adjusted to 5.5 |
| Yeast peptone dextrose (YPD) | 20.0 g/L of glucose (NZYTech, Lisbon, Portugal); 20.0 g/L of bactopeptone (Gibco, Detroit, MI, USA); 10.0 g/L of yeast extract (VWR, Darmstadt, Germany). |
| Fatty Acid Content (%, w/w) | ||
|---|---|---|
| Fatty Acid Type | IST 536 | IST 536 MM15 |
| C14:0 | 2.3 | 1.8 |
| C16:0 | 31.7 | 24.6 |
| C16:1 | 0.8 | 0.9 |
| C18:0 | 24.2 | 16.6 |
| C18:1 | 41.0 | 56.1 |
| Total lipid content | 24.9 | 36.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, M.A.; Mota, M.N.; Faria, N.T.; Sá-Correia, I. An Evolved Strain of the Oleaginous Yeast Rhodotorula toruloides, Multi-Tolerant to the Major Inhibitors Present in Lignocellulosic Hydrolysates, Exhibits an Altered Cell Envelope. J. Fungi 2023, 9, 1073. https://doi.org/10.3390/jof9111073
Fernandes MA, Mota MN, Faria NT, Sá-Correia I. An Evolved Strain of the Oleaginous Yeast Rhodotorula toruloides, Multi-Tolerant to the Major Inhibitors Present in Lignocellulosic Hydrolysates, Exhibits an Altered Cell Envelope. Journal of Fungi. 2023; 9(11):1073. https://doi.org/10.3390/jof9111073
Chicago/Turabian StyleFernandes, Mónica A., Marta N. Mota, Nuno T. Faria, and Isabel Sá-Correia. 2023. "An Evolved Strain of the Oleaginous Yeast Rhodotorula toruloides, Multi-Tolerant to the Major Inhibitors Present in Lignocellulosic Hydrolysates, Exhibits an Altered Cell Envelope" Journal of Fungi 9, no. 11: 1073. https://doi.org/10.3390/jof9111073
APA StyleFernandes, M. A., Mota, M. N., Faria, N. T., & Sá-Correia, I. (2023). An Evolved Strain of the Oleaginous Yeast Rhodotorula toruloides, Multi-Tolerant to the Major Inhibitors Present in Lignocellulosic Hydrolysates, Exhibits an Altered Cell Envelope. Journal of Fungi, 9(11), 1073. https://doi.org/10.3390/jof9111073