
Sporothrix brasiliensis Infection Modulates Antimicrobial Peptides and Stress Management Gene Expression in the Invertebrate Biomodel Galleria mellonella

 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Galleria mellonella Survival Assays
2.3. Determination of Fungal Load during the Survival Curve
2.4. Determination of Hemocyte Density in Hemolymph
2.5. Analysis of Gene Expression
2.5.1. RNA Extraction and cDNA Synthesis
2.5.2. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
2.6. Statistical Analyses
3. Results
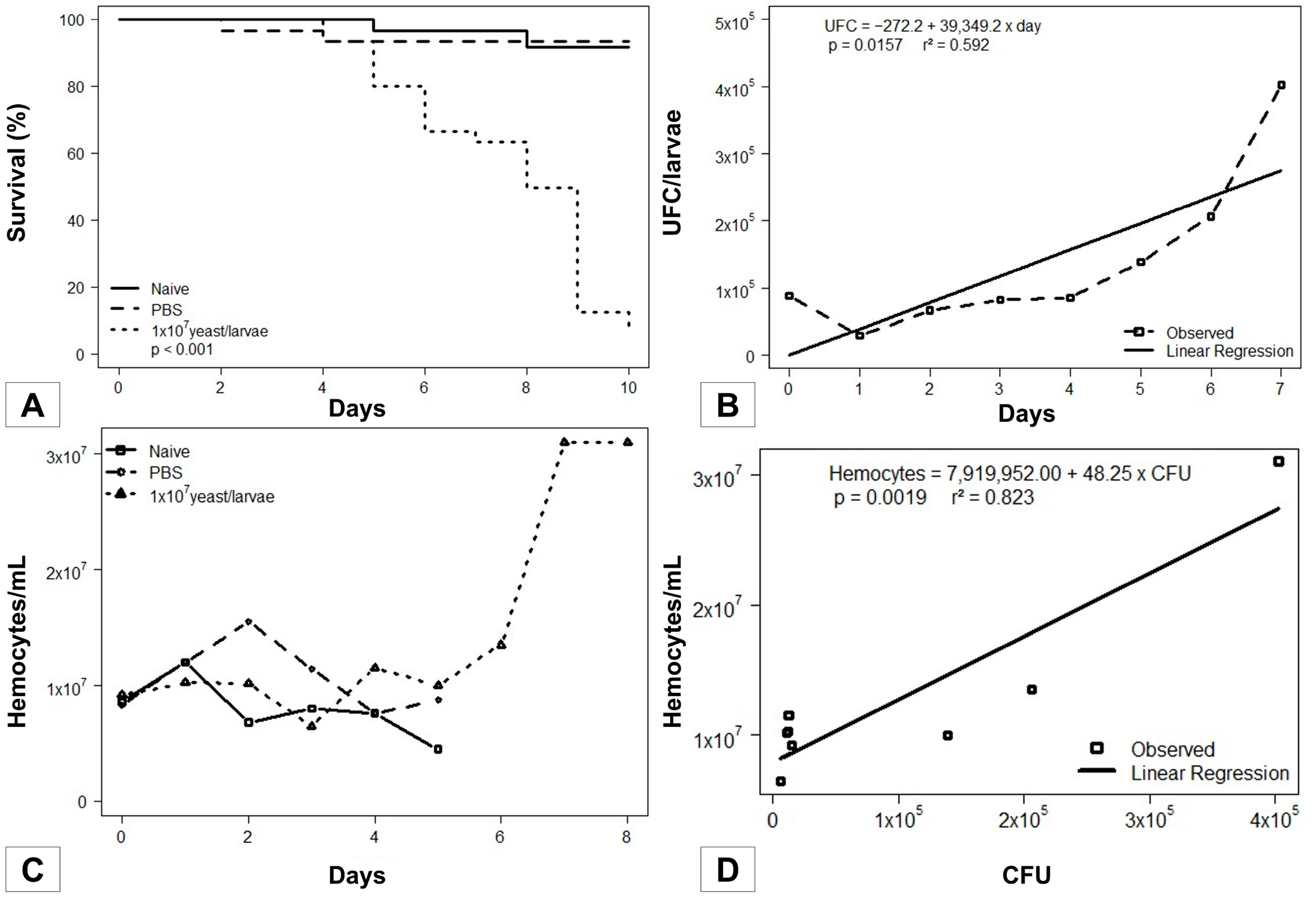
3.1. Galleria mellonella Survival Assays
3.2. Fungal Load during the Infection
3.3. Sporothrix brasiliensis–Galleria mellonella Interaction
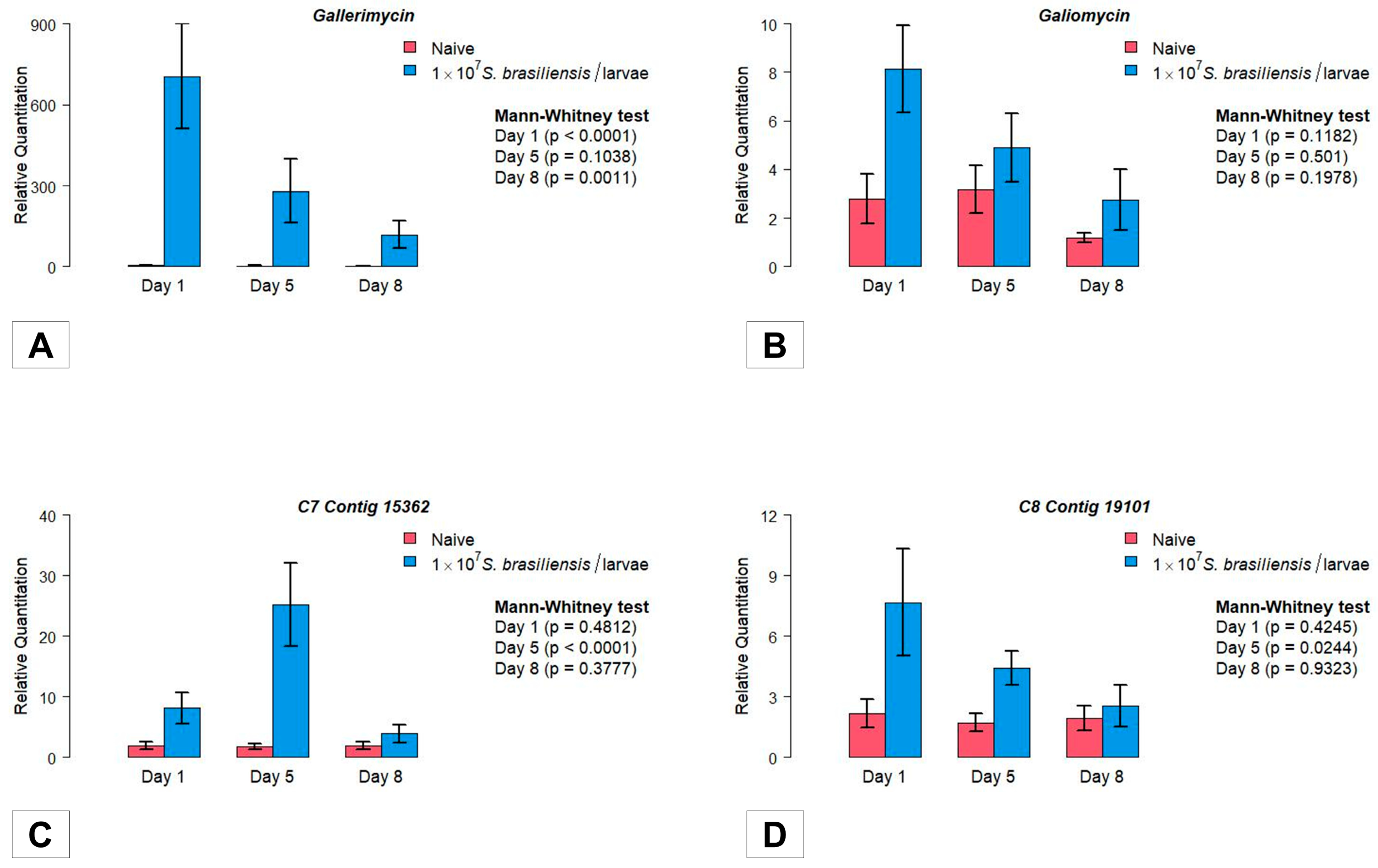
3.4. Gene Expression
3.4.1. Antimicrobial Peptides
3.4.2. Stress Managing Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gremião, I.D.F.; Miranda, L.H.M.; Reis, E.G.; Rodrigues, A.M.; Pereira, S.A. Zoonotic Epidemic of Sporotrichosis: Cat to Human Transmission. PLoS Pathog. 2017, 13, e1006077. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; Della Terra, P.P.; Gremião, I.D.; Pereira, S.A.; Orofino-Costa, R.; de Camargo, Z.P. The Threat of Emerging and Re-Emerging Pathogenic Sporothrix Species. Mycopathologia 2020, 185, 813–842. [Google Scholar] [CrossRef] [PubMed]
- Della Terra, P.P.; Rodrigues, A.M.; Fernandes, G.F.; Nishikaku, A.S.; Burger, E.; de Camargo, Z.P. Exploring Virulence and Immunogenicity in the Emerging Pathogen Sporothrix brasiliensis. PLoS Negl. Trop. Dis. 2017, 11, e0005903. [Google Scholar] [CrossRef]
- Nakasu, C.C.T.; Waller, S.B.; Ripoll, M.K.; Ferreira, M.R.A.; Conceição, F.R.; Dos Reis Gomes, A.; da Gama Osório, L.; de Faria, R.O.; Cleff, M.B. Feline Sporotrichosis: A Case Series of Itraconazole-Resistant Sporothrix brasiliensis Infection. Braz. J. Microbiol. 2021, 52, 163–171. [Google Scholar] [CrossRef]
- Gremião, I.D.F.; Oliveira, M.M.E.; Monteiro de Miranda, L.H.; Saraiva Freitas, D.F.; Pereira, S.A. Geographic Expansion of Sporotrichosis, Brazil. Emerg. Infect. Dis. 2020, 26, 621–624. [Google Scholar] [CrossRef]
- Schechtman, R.C.; Falcão, E.M.M.; Carard, M.; García, M.S.C.; Mercado, D.S.; Hay, R.J. Sporotrichosis: Hyperendemic by Zoonotic Transmission, with Atypical Presentations, Hypersensitivity Reactions and Greater Severity. An. Bras. De Dermatol. 2022, 97, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Etchecopaz, A.; Toscanini, M.A.; Gisbert, A.; Mas, J.; Scarpa, M.; Iovannitti, C.A.; Bendezú, K.; Nusblat, A.D.; Iachini, R.; Cuestas, M.L. Sporothrix brasiliensis: A Review of an Emerging South American Fungal Pathogen, Its Related Disease, Presentation and Spread in Argentina. J. Fungi 2021, 7, 170. [Google Scholar] [CrossRef] [PubMed]
- Thomson, P.; González, C.; Blank, O.; Ramírez, V.; del Río, C.; Santibáñez, S.; Pena, P. Sporotrichosis Outbreak Due to Sporothrix brasiliensis in Domestic Cats in Magallanes, Chile: A One-Health-Approach Study. J. Fungi 2023, 9, 226. [Google Scholar] [CrossRef]
- Barnacle, J.R.; Chow, Y.J.; Borman, A.M.; Wyllie, S.; Dominguez, V.; Russell, K.; Roberts, H.; Armstrong-James, D.; Whittington, A.M. The First Three Reported Cases of Sporothrix brasiliensis Cat-Transmitted Sporotrichosis Outside South America. Med. Mycol. Case Rep. 2023, 39, 14–17. [Google Scholar] [CrossRef]
- Orofino-Costa, R.; de Macedo, P.M.; Rodrigues, A.M.; Bernardes-Engemann, A.R. Sporotrichosis: An Update on Epidemiology, Etiopathogenesis, Laboratory and Clinical Therapeutics. An. Bras. Dermatol. 2017, 92, 606–620. [Google Scholar] [CrossRef]
- Gremião, I.D.F.; Martins da Silva da Rocha, E.; Montenegro, H.; Carneiro, A.J.B.; Xavier, M.O.; de Farias, M.R.; Monti, F.; Mansho, W.; de Macedo Assunção Pereira, R.H.; Pereira, S.A.; et al. Guideline for the Management of Feline Sporotrichosis Caused by Sporothrix brasiliensis and Literature Revision. Braz. J. Microbiol. 2021, 52, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Dekkerová-Chupáčová, J.; Borghi, E.; Morace, G.; Bujdáková, H. Up-Regulation of Antimicrobial Peptides Gallerimycin and Galiomicin in Galleria mellonella Infected with Candida Yeasts Displaying Different Virulence Traits. Mycopathologia 2018, 183, 935–940. [Google Scholar] [CrossRef] [PubMed]
- García-Carnero, L.C.; Clavijo-Giraldo, D.M.; Gómez-Gaviria, M.; Lozoya-Pérez, N.E.; Tamez-Castrellón, A.K.; López-Ramírez, L.A.; Mora-Montes, H.M. Early Virulence Predictors during the Candida Species–Galleria mellonella Interaction. J. Fungi 2020, 6, 152. [Google Scholar] [CrossRef] [PubMed]
- Trevijano-Contador, N.; Herrero-Fernández, I.; García-Barbazán, I.; Scorzoni, L.; Rueda, C.; Rossi, S.A.; García-Rodas, R.; Zaragoza, O. Cryptococcus neoformans Induces Antimicrobial Responses and Behaves as a Facultative Intracellular Pathogen in the Non Mammalian Model Galleria mellonella. Virulence 2015, 6, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, I. Animal Models to Study Mucormycosis. J. Fungi 2019, 5, 27. [Google Scholar] [CrossRef] [PubMed]
- Maurer, E.; Hörtnagl, C.; Lackner, M.; Grässle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a Model System to Study Virulence Potential of Mucormycetes and Evaluation of Antifungal Treatment. Med. Mycol. 2019, 57, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Stączek, S.; Zdybicka-Barabas, A.; Wojda, I.; Wiater, A.; Mak, P.; Suder, P.; Skrzypiec, K.; Cytryńska, M. Fungal α-1,3-Glucan as a New Pathogen-Associated Molecular Pattern in the Insect Model Host Galleria mellonella. Molecules 2021, 26, 5097. [Google Scholar] [CrossRef]
- Thomaz, L.; García-Rodas, R.; Guimarães, A.J.; Taborda, C.P.; Zaragoza, O.; Nosanchuk, J.D. Galleria mellonella as a Model Host to Study Paracoccidioides lutzii and Histoplasma capsulatum. Virulence 2013, 4, 139–146. [Google Scholar] [CrossRef]
- Singulani, J.L.; Scorzoni, L.; de Oliveira, H.C.; Marcos, C.M.; Assato, P.A.; Fusco-Almeida, A.; Mendes-Giannini, M. Applications of Invertebrate Animal Models to Dimorphic Fungal Infections. J. Fungi 2018, 4, 118. [Google Scholar] [CrossRef]
- Freitas, D.F.; Santos, S.S.; Almeida-Paes, R.; de Oliveira, M.M.; do Valle, A.C.; Gutierrez-Galhardo, M.C.; Zancopé-Oliveira, R.M.; Nosanchuk, J.D. Increase in Virulence of Sporothrix brasiliensis over Five Years in a Patient with Chronic Disseminated Sporotrichosis. Virulence 2015, 6, 112–120. [Google Scholar] [CrossRef]
- Clavijo-Giraldo, D.M.; Matínez-Alvarez, J.A.; Lopes-Bezerra, L.M.; Ponce-Noyola, P.; Franco, B.; Almeida, R.S.; Mora-Montes, H.M. Analysis of Sporothrix schenckii sensu stricto and Sporothrix brasiliensis Virulence in Galleria mellonella. J. Microbiol. Methods 2016, 122, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Lozoya-Pérez, N.E.; Casas-Flores, S.; Martínez-Álvarez, J.A.; López-Ramírez, L.A.; Lopes-Bezerra, L.M.; Franco, B.; Mora-Montes, H.M. Generation of Sporothrix schenckii Mutants Expressing the Green Fluorescent Protein Suitable for the Study of Host-Fungus Interactions. Fungal Biol. 2018, 122, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Borba-Santos, L.P.; Barreto, T.L.; Vila, T.; Chi, K.D.; dos Santos Monti, F.; de Farias, M.R.; Alviano, D.S.; Alviano, C.S.; Futuro, D.O.; Ferreira, V.; et al. In Vitro and In Vivo Antifungal Activity of Buparvaquone against Sporothrix brasiliensis. Antimicrob. Agents Chemother. 2021, 65, e00699-21. [Google Scholar] [CrossRef]
- Browne, N.; Heelan, M.; Kavanagh, K. An Analysis of the Structural and Functional Similarities of Insect Hemocytes and Mammalian Phagocytes. Virulence 2013, 4, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Garvey, A.; Croke, M.; Kavanagh, K. Innate Humoral Immune Defences in Mammals and Insects: The Same, with Differences? Virulence 2018, 9, 1625–1639. [Google Scholar] [CrossRef] [PubMed]
- Wojda, I. Immunity of the Greater Wax Moth Galleria mellonella: Galleria mellonella Immunity. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, N.A.; Gagen, S.J. Studies on the in vivo Cellular Reactions of Insects: An Ultrastructural Analysis of Nodule Formation in Galleria mellonella. Tissue Cell 1977, 9, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Lavine, M.D.; Strand, M.R. Insect Hemocytes and Their Role in Immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Pereira, T.; de Barros, P.; Fugisaki, L.; Rossoni, R.; Ribeiro, F.; de Menezes, R.; Junqueira, J.; Scorzoni, L. Recent Advances in the Use of Galleria mellonella Model to Study Immune Responses against Human Pathogens. J. Fungi 2018, 4, 128. [Google Scholar] [CrossRef]
- de Melo, N.R.; Abdrahman, A.; Greig, C.; Mukherjee, K.; Thornton, C.; Ratcliffe, N.A.; Vilcinskas, A.; Butt, T.M. Myriocin Significantly Increases the Mortality of a Non-Mammalian Model Host during Candida Pathogenesis. PLoS ONE 2013, 8, e78905. [Google Scholar] [CrossRef]
- Gandra, R.M.; McCarron, P.; Viganor, L.; Fernandes, M.F.; Kavanagh, K.; McCann, M.; Branquinha, M.H.; Santos, A.L.S.; Howe, O.; Devereux, M. in vivo Activity of Copper(II), Manganese(II), and Silver(I) 1,10-Phenanthroline Chelates Against Candida Haemulonii Using the Galleria mellonella Model. Front. Microbiol. 2020, 11, 470. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Altincicek, B.; Glöckner, G.; Vilcinskas, A. A Comprehensive Transcriptome and Immune-Gene Repertoire of the Lepidopteran Model Host Galleria mellonella. BMC Genom. 2011, 12, 308. [Google Scholar] [CrossRef] [PubMed]
- Nale, J.Y.; Chutia, M.; Cheng, J.K.J.; Clokie, M.R.J. Refining the Galleria mellonella Model by Using Stress Marker Genes to Assess Clostridioides difficile Infection and Recuperation during Phage Therapy. Microorganisms 2020, 8, 1306. [Google Scholar] [CrossRef]
- Champion, O.; Titball, R.; Bates, S. Standardization of G. mellonella Larvae to Provide Reliable and Reproducible Results in the Study of Fungal Pathogens. J. Fungi 2018, 4, 108. [Google Scholar] [CrossRef]
- Senior, N.J.; Titball, R.W. Isolation and Primary Culture of Galleria mellonella Hemocytes for Infection Studies. F1000Research 2021, 9, 1392. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Lucas, M.P.; Mesa-Arango, A.C.; Fusco-Almeida, A.M.; Lozano, E.; Cuenca-Estrella, M.; Mendes-Giannini, M.J.; Zaragoza, O. Antifungal Efficacy during Candida krusei Infection in Non-Conventional Models Correlates with the Yeast In Vitro Susceptibility Profile. PLoS ONE 2013, 8, e60047. [Google Scholar] [CrossRef]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. A Peptidomics Study Reveals the Impressive Antimicrobial Peptide Arsenal of the Wax Moth Galleria mellonella. Insect Biochem. Mol. Biol. 2009, 39, 792–800. [Google Scholar] [CrossRef]
- Zhao, H.-X.; Xiao, W.-Y.; Ji, C.-H.; Ren, Q.; Xia, X.-S.; Zhang, X.-F.; Huang, W.-Z. Candidate Chemosensory Genes Identified from the Greater Wax Moth, Galleria mellonella, through a Transcriptomic Analysis. Sci. Rep. 2019, 9, 10032. [Google Scholar] [CrossRef]
- Miranda, L.H.M.; Santiago, M.d.A.; Schubach, T.M.P.; Morgado, F.N.; Pereira, S.A.; Oliveira, R.d.V.C.d.; Conceição-Silva, F. Severe Feline Sporotrichosis Associated with an Increased Population of CD8 low Cells and a Decrease in CD4 + Cells. Med. Mycol. 2015, 54, 29–39. [Google Scholar] [CrossRef]
- Legrand, N.; Weijer, K.; Spits, H. Experimental Models to Study Development and Function of the Human Immune System In Vivo. J. Immunol. 2006, 176, 2053–2058. [Google Scholar] [CrossRef]
- Arrillaga-Moncrieff, I.; Capilla, J.; Mayayo, E.; Marimon, R.; Marine, M.; Genis, J.; Cano, J.; Guarro, J. Different Virulence Levels of the Species of Sporothrix in a Murine Model. Clin. Microbiol. Infect. 2009, 15, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.M. Of Mice and Men and Larvae: Galleria mellonella to Model the Early Host-Pathogen Interactions after Fungal Infection. Virulence 2018, 9, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gaviria, M.; Martínez-Álvarez, J.A.; Mora-Montes, H.M. Current Progress in Sporothrix brasiliensis Basic Aspects. J. Fungi 2023, 9, 533. [Google Scholar] [CrossRef] [PubMed]
- Piatek, M.; Sheehan, G.; Kavanagh, K. Utilising Galleria mellonella Larvae for Studying in vivo Activity of Conventional and Novel Antimicrobial Agents. Pathog. Dis. 2020, 78, ftaa059. [Google Scholar] [CrossRef]
- Dinh, H.; Semenec, L.; Kumar, S.S.; Short, F.L.; Cain, A.K. Microbiology’s next Top Model: Galleria in the Molecular Age. Pathog. Dis. 2021, ftab006. [Google Scholar] [CrossRef]
- Binder, U.; Maurer, E.; Lass-Flörl, C. Galleria mellonella: An Invertebrate Model to Study Pathogenicity in Correctly Defined Fungal Species. Fungal Biol. 2016, 120, 288–295. [Google Scholar] [CrossRef]
- Fakhim, H.; Vaezi, A.; Dannaoui, E.; Chowdhary, A.; Nasiry, D.; Faeli, L.; Meis, J.F.; Badali, H. Comparative Virulence of Candida Auris with Candida haemulonii, Candida glabrata and Candida albicans in a Murine Model. Mycoses 2018, 61, 377–382. [Google Scholar] [CrossRef]
- Corrêa-Moreira, D.; Menezes, R.C.; Romeo, O.; Borba, C.M.; Oliveira, M.M.E. Clinical and Anatomopathological Evaluation of BALB/c Murine Models Infected with Isolates of Seven Pathogenic Sporothrix Species. Pathogens 2021, 10, 1647. [Google Scholar] [CrossRef]
- Trevijano-Contador, N.; Zaragoza, O. Immune Response of Galleria mellonella against Human Fungal Pathogens. J. Fungi 2018, 5, 3. [Google Scholar] [CrossRef]
- Tamez-Castrellón, A.K.; van der Beek, S.L.; López-Ramírez, L.A.; Martínez-Duncker, I.; Lozoya-Pérez, N.E.; van Sorge, N.M.; Mora-Montes, H.M. Disruption of Protein Rhamnosylation Affects the Sporothrix schenckii-Host Interaction. Cell Surf. 2021, 7, 100058. [Google Scholar] [CrossRef]
- Rossoni, R.D.; Fuchs, B.B.; de Barros, P.P.; Velloso, M.d.S.; Jorge, A.O.C.; Junqueira, J.C.; Mylonakis, E. Lactobacillus Paracasei Modulates the Immune System of Galleria mellonella and Protects against Candida albicans Infection. PLoS ONE 2017, 12, e0173332. [Google Scholar] [CrossRef] [PubMed]
- Mancera, E.; Nocedal, I.; Hammel, S.; Gulati, M.; Mitchell, K.F.; Andes, D.R.; Nobile, C.J.; Butler, G.; Johnson, A.D. Evolution of the complex transcription network controlling biofilm formation in Candida species. eLife 2021, 10, e64682. [Google Scholar] [CrossRef] [PubMed]
- de Beer, Z.W.; Duong, T.A.; Wingfield, M.J. The Divorce of Sporothrix and Ophiostoma: Solution to a Problematic Relationship. Stud. Mycol. 2016, 83, 165–191. [Google Scholar] [CrossRef]
- Meister, M.; Lemaitre, B.; Hoffmann, J.A. Antimicrobial Peptide Defense in Drosophila. Bioessays 1997, 19, 1019–1026. [Google Scholar] [CrossRef]
- Bulet, P.; Hetru, C.; Dimarcq, J.-L.; Hoffmann, D. Antimicrobial Peptides in Insects; Structure and Function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Lee, Y.S.; Yun, E.K.; Jang, W.S.; Kim, I.; Lee, J.H.; Park, S.Y.; Ryu, K.S.; Seo, S.J.; Kim, C.H.; Lee, I.H. Purification, CDNA Cloning and Expression of an Insect Defensin from the Great Wax Moth, Galleria mellonella. Insect Mol. Biol. 2004, 13, 65–72. [Google Scholar] [CrossRef]
- Mothé, G.B. Estudo Da Interação Entre Fagócitos Do Felis Catus (LINNAEUS,1758) e os Principais Agentes Etiológicos Da Esporotricose. Ph.D. thesis, Universidade Federal Fluminense, Rio de Janeiro, Brasil, 2021. Available online: https://app.uff.br/riuff/handle/1/24482 (accessed on 29 August 2023).
- Złotko, K.; Wiater, A.; Waśko, A.; Pleszczyńska, M.; Paduch, R.; Jaroszuk-Ściseł, J.; Bieganowski, A. A Report on Fungal (1→3)-α-d-Glucans: Properties, Functions and Application. Molecules 2019, 24, 3972. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | NCBI Genbank References | Sequence (5′–3′) | Annealing Temperature |
|---|---|---|---|
| Galiomycin | AY528421.1 | F-TCCAGTCCGTTTTGTTGTTG | 60 °C |
| [37] | R-CAGAGGTGTAATTCGTCGCA | ||
| Gallerimycin | AF453824.1 | F-GAAGATCGCTTTCATAGTCGC | 60 °C |
| [37] | R-TACTCCTGCAGTTAGCAATGC | ||
| C7 Contig 15362 | Contig 15362 | F-CGAGCTAAAGACAGGCGATT | 58 °C |
| [30] | R-TCACCTGCGGTTGAATCATA | ||
| C8 Contig 19101 | Contig 19101 | F-ATTGCTAGCCAGGTTCAGGA | 60 °C |
| [30] | R-AGCTATTTGGCGGAAACTCA | ||
| β-actin | [38] | F-GGACTTGTACGCCAACACAG R-CCACATCTGCTGGAATGTCG | 55 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, N.F.; de Jesus, M.C.S.; de Souza, L.C.d.S.V.; Alcântara, L.M.; Rodrigues, J.A.d.C.; Brito, S.C.P.; Penna, P.d.A.; Vieira, C.S.; Silva, J.R.S.; Penna, B.d.A.; et al. Sporothrix brasiliensis Infection Modulates Antimicrobial Peptides and Stress Management Gene Expression in the Invertebrate Biomodel Galleria mellonella. J. Fungi 2023, 9, 1053. https://doi.org/10.3390/jof9111053
Reis NF, de Jesus MCS, de Souza LCdSV, Alcântara LM, Rodrigues JAdC, Brito SCP, Penna PdA, Vieira CS, Silva JRS, Penna BdA, et al. Sporothrix brasiliensis Infection Modulates Antimicrobial Peptides and Stress Management Gene Expression in the Invertebrate Biomodel Galleria mellonella. Journal of Fungi. 2023; 9(11):1053. https://doi.org/10.3390/jof9111053
Chicago/Turabian StyleReis, Nathália Faria, Myrela Conceição Santos de Jesus, Lais Cavalcanti dos Santos Velasco de Souza, Lucas Martins Alcântara, Julia Andrade de Castro Rodrigues, Simone Cristina Pereira Brito, Patrícia de Azambuja Penna, Cecília Stahl Vieira, José Rodrigo Santos Silva, Bruno de Araújo Penna, and et al. 2023. "Sporothrix brasiliensis Infection Modulates Antimicrobial Peptides and Stress Management Gene Expression in the Invertebrate Biomodel Galleria mellonella" Journal of Fungi 9, no. 11: 1053. https://doi.org/10.3390/jof9111053
APA StyleReis, N. F., de Jesus, M. C. S., de Souza, L. C. d. S. V., Alcântara, L. M., Rodrigues, J. A. d. C., Brito, S. C. P., Penna, P. d. A., Vieira, C. S., Silva, J. R. S., Penna, B. d. A., Machado, R. L. D., Mora-Montes, H. M., & Baptista, A. R. d. S. (2023). Sporothrix brasiliensis Infection Modulates Antimicrobial Peptides and Stress Management Gene Expression in the Invertebrate Biomodel Galleria mellonella. Journal of Fungi, 9(11), 1053. https://doi.org/10.3390/jof9111053