
Identification of Candidate Ice Nucleation Activity (INA) Genes in Fusarium avenaceum by Combining Phenotypic Characterization with Comparative Genomics and Transcriptomics



Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. INA Measurements
2.3. Characterization of INPs
2.4. Investigating the Effect of Growth Conditions on INA
2.5. Genome and Transcriptome Sequencing and Assembly
2.6. Phylogenetic Analyses
2.7. Gene Prediction and Genome Annotation
2.8. Functional Annotation, Prediction of the Signal Peptide and Prediction of Secondary Metabolite Genes
2.9. Pan-Genome Analysis and Prediction of Orthologs
2.10. Gene Expression Analysis
2.11. DNAseq Analysis
3. Results and Discussion
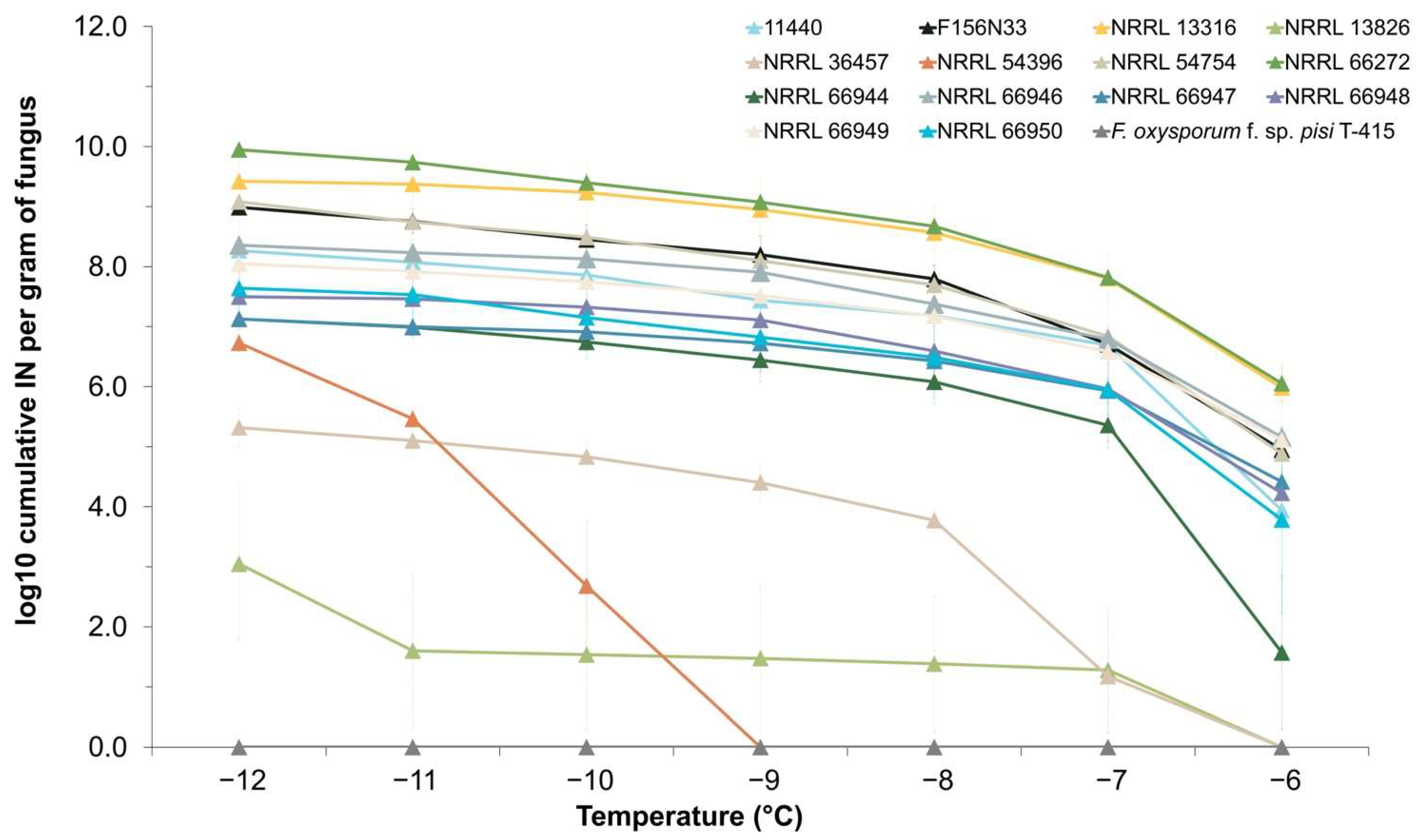
3.1. INA Varies between Strains of F. avenaceum
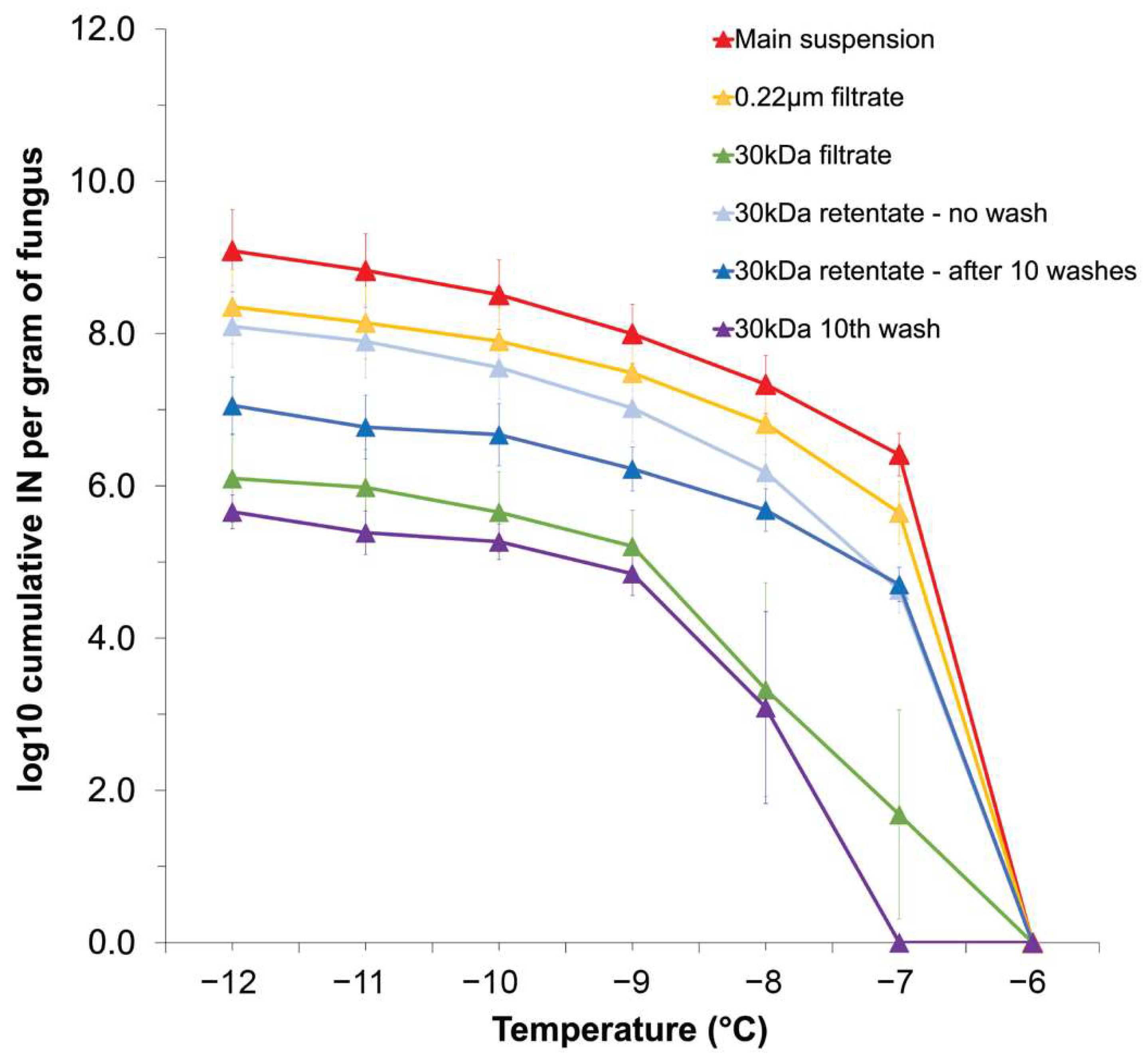
3.2. F. avenaceum INPs Are Secreted Aggregates Prone to Separation by Centrifugation and Washing
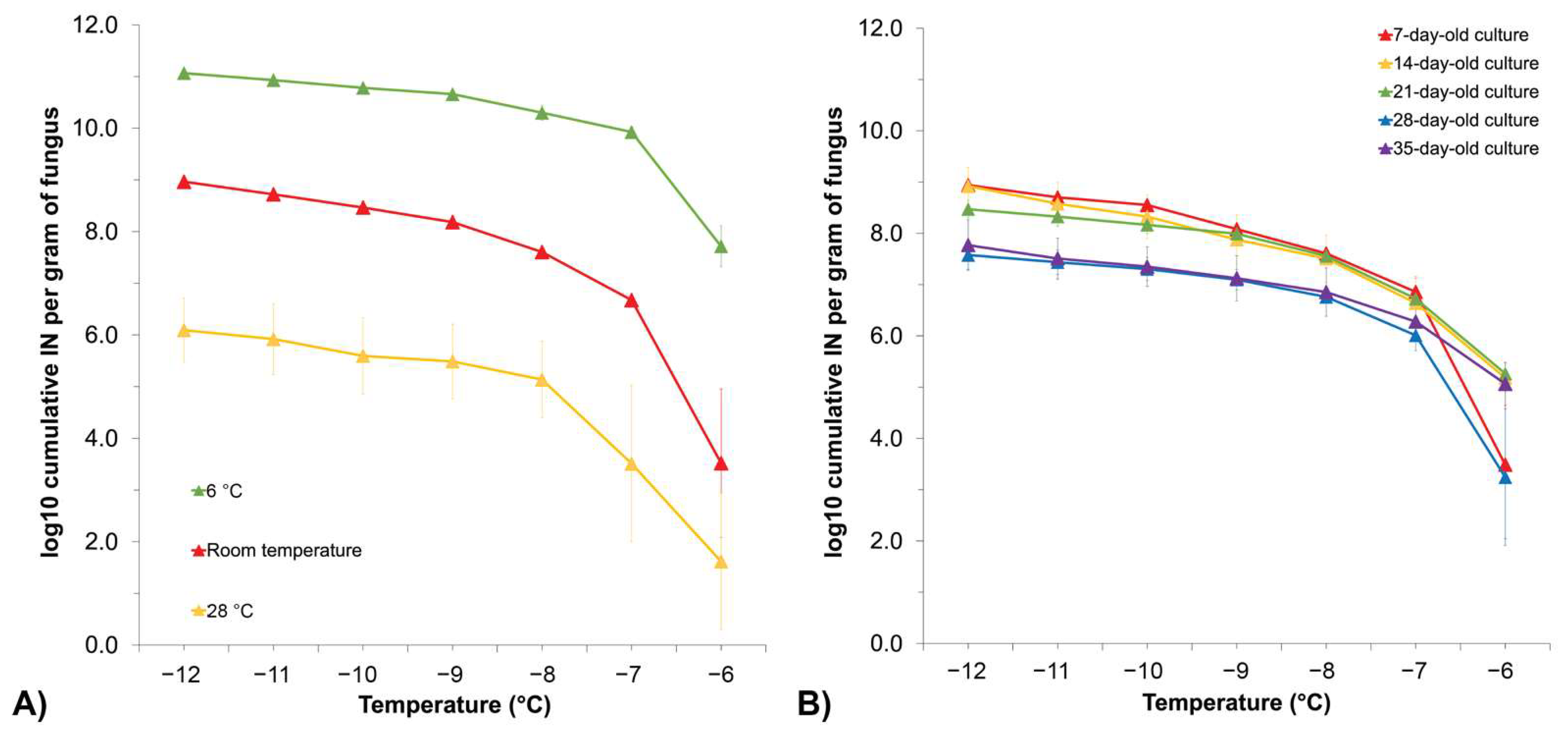
3.3. Growth Temperature Affects Fusarium INA, while the Length of Growth Time Does Not Have a Significant Impact
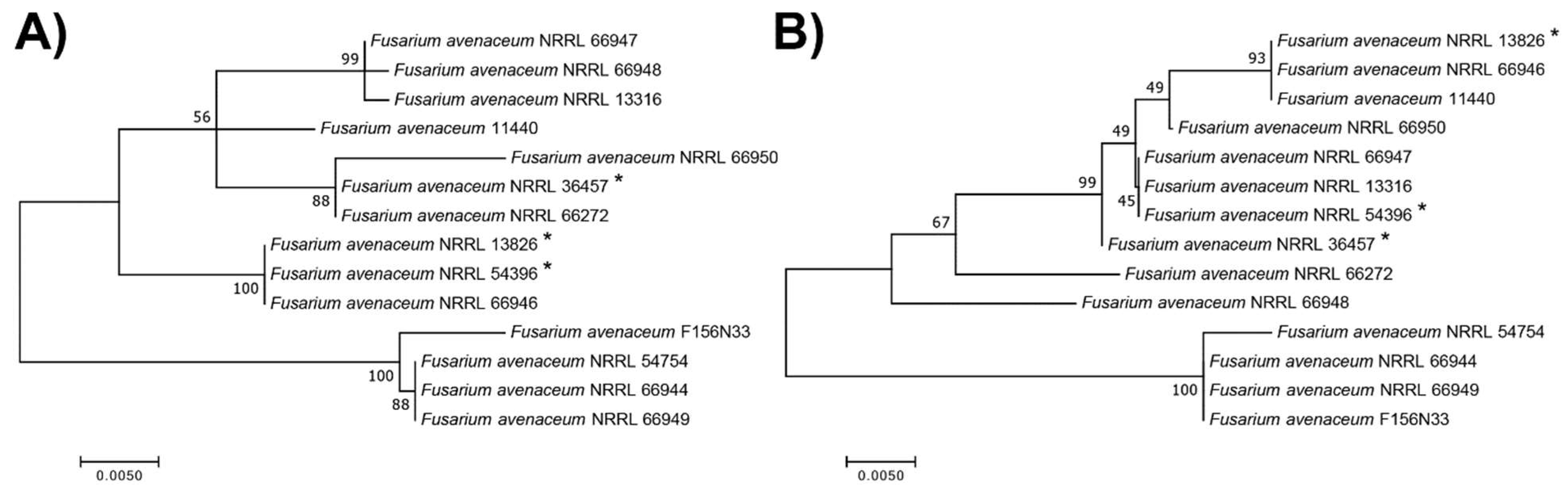
3.4. Phylogenetic Analyses Reveal That F. avenaceum Strains Form Several Within-Species Clusters and That the Strength of INA Does Not Correlate with Phylogeny
3.5. A Comparative Genomics Approach to Identifying Candidate INA Genes in F. avenaceum
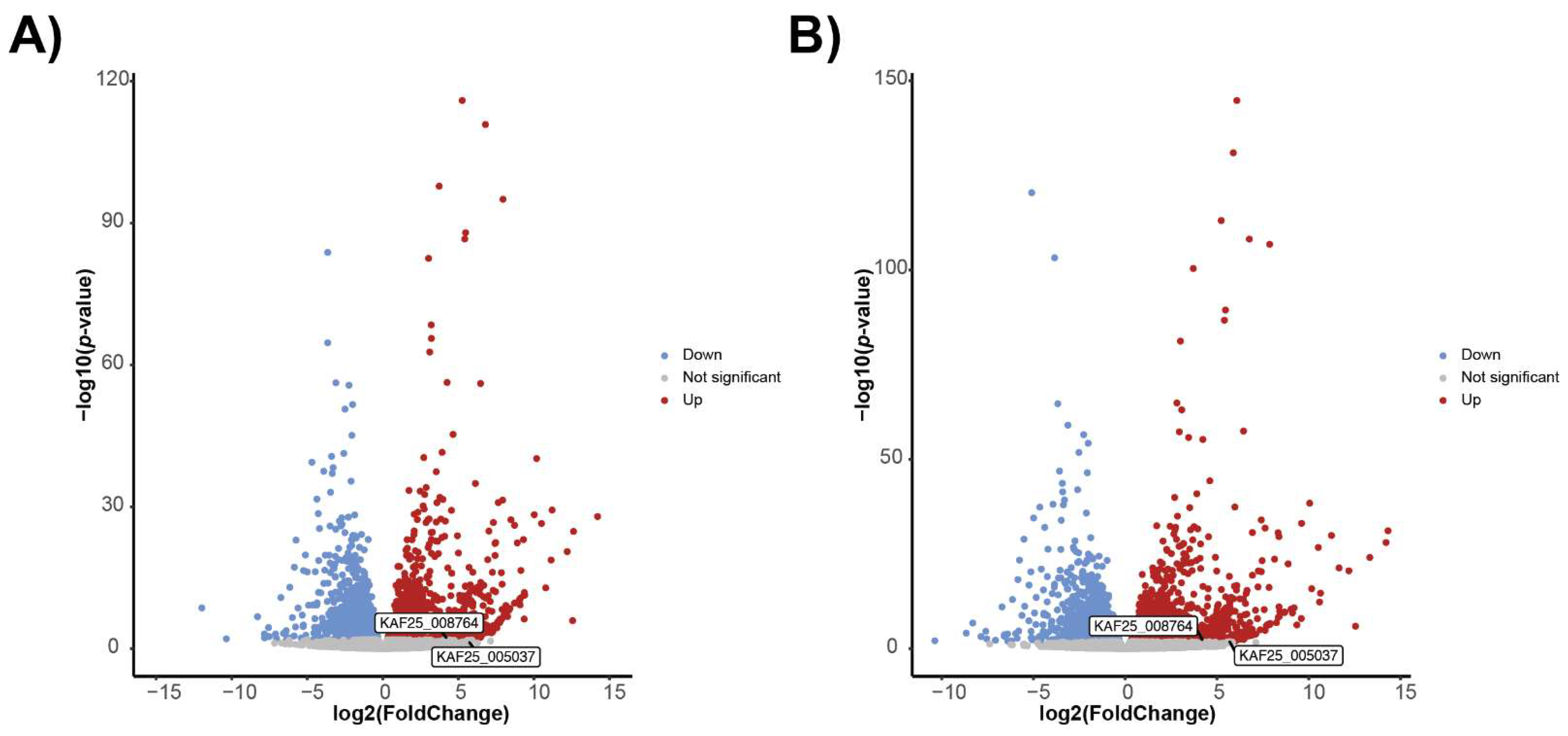
3.6. Transcriptomics Approach to Identify Candidate INA Genes in F. avenaceum
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koop, T.; Luo, B.; Tsias, A.; Peter, T. Water activity as the determinant for homogeneous ice nucleation in aqueous solutions. Nature 2000, 406, 611–614. [Google Scholar] [CrossRef]
- Santachiara, G.; Belosi, F. Does the homogeneous ice nucleation initiate in the bulk volume or at the surface of super-cooled water droplets? A review. Atmos. Clim. Sci. 2014, 4, 653. [Google Scholar] [CrossRef][Green Version]
- Matus, A.V.; L’Ecuyer, T.S. The role of cloud phase in Earth’s radiation budget. J. Geophys. Res. Atmos. 2017, 122, 2559–2578. [Google Scholar] [CrossRef]
- Murray, B.J.; O’Sullivan, D.; Atkinson, J.D.; Webb, M.E. Ice nucleation by particles immersed in supercooled cloud droplets. Chem. Soc. Rev. 2012, 41, 6519–6554. [Google Scholar] [CrossRef] [PubMed]
- Lundheim, R.; Zachariassen, K. Applications of biological ice nucleators. In Biotechnological Applications of Cold-Adapted Organisms; Springer: Berlin/Heidelberg, Germany, 1999; pp. 309–317. [Google Scholar]
- Saxena, V.K. Evidence of the biogenic nuclei involvement in Antarctic coastal clouds. J. Phys. Chem. 1983, 87, 4130–4134. [Google Scholar] [CrossRef]
- Pratt, K.A.; DeMott, P.J.; French, J.R.; Wang, Z.; Westphal, D.L.; Heymsfield, A.J.; Twohy, C.H.; Prenni, A.J.; Prather, K.A. In situ detection of biological particles in cloud ice-crystals. Nat. Geosci. 2009, 2, 398–401. [Google Scholar] [CrossRef]
- Uhlig, S.; Jestoi, M.; Parikka, P. Fusarium avenaceum-the North European situation. Int. J. Food Microbiol. 2007, 119, 17–24. [Google Scholar] [CrossRef]
- Christner, B.C.; Cai, R.; Morris, C.E.; McCarter, K.S.; Foreman, C.M.; Skidmore, M.L.; Montross, S.N.; Sands, D.C. Geographic, seasonal, and precipitation chemistry influence on the abundance and activity of biological ice nucleators in rain and snow. Proc. Natl. Acad. Sci. USA 2008, 105, 18854–18859. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.C.; Vinatzer, B.A.; Glaux, C.; Guilbaud, C.; Buffière, A.; Yan, S.; Dominguez, H.; Thompson, B. The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle. ISME J. 2008, 2, 321–334. [Google Scholar] [CrossRef]
- Christner, B.C.; Morris, C.E.; Foreman, C.M.; Cai, R.; Sands, D.C. Ubiquity of biological ice nucleators in snowfall. Science 2008, 319, 1214. [Google Scholar] [CrossRef]
- Sanchez-Marroquin, A.; West, J.S.; Burke, I.T.; McQuaid, J.B.; Murray, B.J. Mineral and biological ice-nucleating particles above the South East of the British Isles. Environ. Sci. Atmos. 2021, 1, 176–191. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.P.; Atanasova, N.S.; Sofieva, S.; Ravantti, J.; Heikkinen, A.; Brasseur, Z.; Duplissy, J.; Bamford, D.H.; Murray, B.J. Ice nucleation by viruses and their potential for cloud glaciation. Biogeosciences 2021, 18, 4431–4444. [Google Scholar] [CrossRef]
- Maki, L.R.; Willoughby, K.J. Bacteria as biogenic sources of freezing nuclei. J. Appl. Meteorol. 1978, 17, 1049–1053. [Google Scholar] [CrossRef]
- Anderson, J.A.; Ashworth, E.N. The effects of streptomycin, desiccation, and UV radiation on ice nucleation by Pseudomonas viridiflava. Plant Physiol. 1986, 80, 956–960. [Google Scholar] [CrossRef]
- Maki, L.R.; Galyan, E.L.; Chang-Chien, M.M.; Caldwell, D.R. Ice nucleation induced by Pseudomonas syringae. Appl. Microbiol. 1974, 28, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.; Arny, D.; Upper, C. Erwinia herbicola: A bacterial ice nucleus active in increasing frost injury to corn. Phytopathology 1978, 68, 523–527. [Google Scholar] [CrossRef]
- Michigami, Y.; Watabe, S.; Abe, K.; Obata, H.; Arai, S. Cloning and sequencing of an ice nucleation active gene of Erwinia uredovora. Biosci. Biotechnol. Biochem. 1994, 58, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Phelps, P.; Giddings, T.H.; Prochoda, M.; Fall, R. Release of cell-free ice nuclei by Erwinia herbicola. J. Bacteriol. 1986, 167, 496–502. [Google Scholar] [CrossRef]
- Wolber, P.K.; Deininger, C.A.; Southworth, M.W.; Vandekerckhove, J.; van Montagu, M.; Warren, G.J. Identification and purification of a bacterial ice-nucleation protein. Proc. Natl. Acad. Sci. USA 1986, 83, 7256–7260. [Google Scholar] [CrossRef]
- Kim, H.K.; Orser, C.; Lindow, S.E.; Sands, D.C. Xanthomonas campestris pv. translucens strains active in ice nucleation. Plant Dis. 1987, 71, 994–997. [Google Scholar] [CrossRef]
- Michigami, Y.; Abe, K.; Iwabuchi, K.; Obata, H.; Arai, S. Formation of ice nucleation-active vesicles in Erwinia uredovora at low temperature and transport of InaU molecules into shed vesicles. Biosci. Biotechnol. Biochem. 1995, 59, 1996–1998. [Google Scholar] [CrossRef]
- Morris, C.E.; Georgakopoulos, D.G.; Sands, D.C. Ice nucleation active bacteria and their potential role in precipitation. J. Phys. IV Fr. 2004, 121, 87–103. [Google Scholar] [CrossRef]
- Šantl-Temkiv, T.; Sahyoun, M.; Finster, K.; Hartmann, S.; Augustin-Bauditz, S.; Stratmann, F.; Wex, H.; Clauss, T.; Nielsen, N.W.; Sørensen, J.H.; et al. Characterization of airborne ice-nucleation-active bacteria and bacterial fragments. Atmos. Environ. 2015, 109, 105–117. [Google Scholar] [CrossRef]
- Pandey, R.; Usui, K.; Livingstone, R.A.; Fischer, S.A.; Pfaendtner, J.; Backus, E.H.G.; Nagata, Y.; Fröhlich-Nowoisky, J.; Schmüser, L.; Mauri, S.; et al. Ice-nucleating bacteria control the order and dynamics of interfacial water. Sci. Adv. 2016, 2, e1501630. [Google Scholar] [CrossRef]
- Roeters, S.J.; Golbek, T.W.; Bregnhøj, M.; Drace, T.; Alamdari, S.; Roseboom, W.; Kramer, G.; Šantl-Temkiv, T.; Finster, K.; Pfaendtner, J.; et al. Ice-nucleating proteins are activated by low temperatures to control the structure of interfacial water. Nat. Commun. 2021, 12, 1183. [Google Scholar] [CrossRef]
- Failor, K.C.; Schmale, D.G., III; Vinatzer, B.A.; Monteil, C.L. Ice nucleation active bacteria in precipitation are genetically diverse and nucleate ice by employing different mechanisms. ISME J. 2017, 11, 2740–2753. [Google Scholar] [CrossRef]
- Failor, K.C. Identification and Characterization of Ice Nucleation Active Bacteria Isolated from Precipitation. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 2018. [Google Scholar]
- Failor, K.C.; Liu, H.; Llontop, M.E.M.; LeBlanc, S.; Eckshtain-Levi, N.; Sharma, P.; Reed, A.; Yang, S.; Tian, L.; Lefevre, C.; et al. Ice nucleation in a Gram-positive bacterium isolated from precipitation depends on a polyketide synthase and non-ribosomal peptide synthetase. ISME J. 2021, 16, 890–897. [Google Scholar] [CrossRef]
- Arny, D.C.; Lindow, S.E.; Upper, C.D. Frost sensitivity of Zea mays increased by application of Pseudomonas syringae. Nature 1976, 262, 282–284. [Google Scholar] [CrossRef]
- Sule, S. The role of ice formation in the infection of sour cherry leaves by Pseudomonas syringae pv. syringae. Phytopathology 1987, 77, 173–177. [Google Scholar] [CrossRef]
- Morris, C.E.; Monteil, C.L.; Berge, O. The life history of Pseudomonas syringae: Linking agriculture to earth system processes. Annu. Rev. Phytopathol. 2013, 51, 85–104. [Google Scholar] [CrossRef]
- Pouleur, S.; Richard, C.; Martin, J.G.; Antoun, H. Ice nucleation activity in Fusarium acuminatum and Fusarium avenaceum. Appl. Environ. Microbiol. 1992, 58, 2960–2964. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.C.; Glaux, C.; Samsatly, J.; Asaad, S.; Moukahel, A.R.; Gonçalves, F.L.T.; Bigg, E.K. Urediospores of rust fungi are ice nucleation active at >−10 °C and harbor ice nucleation active bacteria. Atmos. Chem. Phys. 2013, 13, 4223–4233. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Hill, T.C.J.; Pummer, B.G.; Yordanova, P.; Franc, G.D.; Pöschl, U. Ice nucleation activity in the widespread soil fungus Mortierella alpina. Biogeosciences 2015, 12, 1057–1071. [Google Scholar] [CrossRef]
- Nelson, P.E.; Dignani, M.C.; Anaissie, E.J. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar] [CrossRef] [PubMed]
- Schmale, D.G., III; Ross, S.D.; Fetters, T.L.; Tallapragada, P.; Wood-Jones, A.K.; Dingus, B. Isolates of Fusarium graminearum collected 40–320 m above ground level cause Fusarium head blight in wheat and produce trichothecene mycotoxins. Aerobiologia 2012, 28, 1–11. [Google Scholar] [CrossRef]
- Palmero, D.; Rodríguez, J.M.; de Cara, M.; Camacho, F.; Iglesias, C.; Tello, J.C. Fungal microbiota from rain water and pathogenicity of Fusarium species isolated from atmospheric dust and rainfall dust. J. Ind. Microbiol. Biotechnol. 2011, 38, 13–20. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, D.; Murray, B.J.; Ross, J.F.; Whale, T.F.; Price, H.C.; Atkinson, J.D.; Umo, N.S.; Webb, M.E. The relevance of nanoscale biological fragments for ice nucleation in clouds. Sci. Rep. 2015, 5, 8082. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Ishihara, Y.; Tokuyama, T. Characteristics of ice-nucleation activity in Fusarium avenaceum IFO 7158. Biosci. Biotechnol. Biochem. 1994, 58, 2273–2274. [Google Scholar] [CrossRef]
- Kunert, A.T.; Pöhlker, M.L.; Tang, K.; Krevert, C.S.; Wieder, C.; Speth, K.R.; Hanson, L.E.; Morris, C.E.; Schmale, D.G., III; Pöschl, U.; et al. Macromolecular fungal ice nuclei in Fusarium: Effects of physical and chemical processing. Biogeosciences 2019, 16, 4647–4659. [Google Scholar] [CrossRef]
- Richard, C.; Martin, J.G.; Pouleur, S. Ice nucleation activity identified in some phytopathogenic Fusarium species. Phytoprotection 1996, 77, 83–92. [Google Scholar] [CrossRef][Green Version]
- Tsumuki, H.; Konno, H. Ice nuclei produced by Fusarium sp. isolated from the gut of the rice stem borer, Chilo suppressalis Walker (Lepidoptera: Pyralidae). Biosci. Biotechnol. Biochem. 1994, 58, 578–579. [Google Scholar] [CrossRef]
- Humphreys, T.L.; Castrillo, L.A.; Lee, M.R. Sensitivity of partially purified ice nucleation activity of Fusarium acuminatum SRSF 616. Curr. Microbiol. 2001, 42, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.P.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef]
- Cole, R.J.; Cox, R.H. Handbook of Toxic Fungal Metabolites; Acamedic Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Sangare, L.; Zhao, Y.; Folly, Y.M.E.; Chang, J.; Li, J.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Liu, Y. Aflatoxin B₁ degradation by a Pseudomonas strain. Toxins 2014, 6, 3028–3040. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Qian, Y.; Chen, N.; Ling, T.; Wang, J.; Jiang, H.; Wang, X.; Qi, K.; Zhou, Y. Detoxification of aflatoxin B1 by Stenotrophomonas sp. CW117 and characterization the thermophilic degradation process. Environ. Pollut. 2020, 261, 114178. [Google Scholar] [CrossRef]
- Coluzza, I.; Creamean, J.; Rossi, M.J.; Wex, H.; Alpert, P.A.; Bianco, V.; Boose, Y.; Dellago, C.; Felgitsch, L.; Fröhlich-Nowoisky, J.; et al. Perspectives on the future of Ice nucleation research: Research needs and unanswered questions identified from two international workshops. Atmosphere 2017, 8, 138. [Google Scholar] [CrossRef]
- Vali, G. Quantitative evaluation of experimental results on the heterogeneous freezing nucleation of supercooled liquids. J. Atmos. Sci. 1971, 28, 402–409. [Google Scholar] [CrossRef]
- Andrews, S.; Krueger, F.; Segonds-Pichon, A.; Biggins, L.; Krueger, C.; Wingett, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010; Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 6 January 2022).
- Yang, S.; Coleman, J.; Vinatzer, B. Genome Resource: Draft genome of Fusarium avenaceum, strain F156N33, isolated from the atmosphere above Virginia and annotated based on RNA sequencing data. Plant Dis. 2022, 106, 720–722. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Campbell, M.S.; Holt, C.; Moore, B.; Yandell, M. Genome annotation and curation using MAKER and MAKER-P. Curr. Protoc. Bioinform. 2014, 48, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Korf, I. Gene finding in novel genomes. BMC Bioinform. 2004, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Diekhans, M.; Baertsch, R.; Haussler, D. Using native and syntenically mapped cDNA alignments to improve de novo gene finding. Bioinformatics 2008, 24, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.U. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2020, 49, D480–D489. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema Marnix, H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a versatile software package for scalable and robust microbial pangenome analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Stoeckert, C.J.; Roos, D.S., Jr. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Vasimuddin, M.; Misra, S.; Li, H.; Aluru, S. Efficient architecture-aware acceleration of BWA-MEM for multicore systems. In Proceedings of the 2019 IEEE International Parallel and Distributed Processing Symposium, Rio de Janeiro, Brazil, 20–24 May 2019; pp. 314–324. [Google Scholar]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Gurian-Sherman, D.; Lindow, S.E. Differential effects of growth temperature on ice nuclei active at different temperatures that are produced by cells of Pseudomonas syringae. Cryobiology 1995, 32, 129–138. [Google Scholar] [CrossRef]
- Mueller, G.M.; Wolber, P.K.; Warren, G.J. Clustering of ice nucleation protein correlates with ice nucleation activity. Cryobiology 1990, 27, 416–422. [Google Scholar] [CrossRef]
- Ohtaki, S.; Maeda, H.; Takahashi, T.; Yamagata, Y.; Hasegawa, F.; Gomi, K.; Nakajima, T.; Abe, K. Novel hydrophobic surface binding protein, HsbA, produced by Aspergillus oryzae. Appl. Environ. Microbiol. 2006, 72, 2407–2413. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Proctor, R.H. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef]
- Lin, C.; Corem, G.; Godsi, O.; Alexandrowicz, G.; Darling, G.R.; Hodgson, A. Ice nucleation on a corrugated surface. J. Am. Chem. Soc. 2018, 140, 15804–15811. [Google Scholar] [CrossRef] [PubMed]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Lagzian, M.; Latifi, A.M.; Bassami, M.R.; Mirzaei, M. An ice nucleation protein from Fusarium acuminatum: Cloning, expression, biochemical characterization and computational modeling. Biotechnol. Lett. 2014, 36, 2043–2051. [Google Scholar] [CrossRef] [PubMed]
- Anastassopoulos, E. Contribution to the study of eukaryotic ice nucleation: Development of a method for selecting freeze-tolerant tobacco (Nicotiana tabacum L.) plants and isolation of an ice nucleation gene from the fungus Fusarium acumitatum. Ph.D. Thesis, University of Crete, Heraklion, Crete, Greece, 2001. [Google Scholar]
- Kwan, A.H.Y.; Winefield, R.D.; Sunde, M.; Matthews, J.M.; Haverkamp, R.G.; Templeton, M.D.; Mackay, J.P. Structural basis for rodlet assembly in fungal hydrophobins. Proc. Natl. Acad. Sci. USA 2006, 103, 3621. [Google Scholar] [CrossRef]
- Macindoe, I.; Kwan, A.H.; Ren, Q.; Morris, V.K.; Yang, W.; Mackay, J.P.; Sunde, M. Self-assembly of functional, amphipathic amyloid monolayers by the fungal hydrophobin EAS. Proc. Natl. Acad. Sci. USA 2012, 109, E804–E811. [Google Scholar] [CrossRef] [PubMed]
- De Vocht, M.L.; Reviakine, I.; Ulrich, W.-P.; Bergsma-Schutter, W.; Wösten, H.A.B.; Vogel, H.; Brisson, A.; Wessels, J.G.H.; Robillard, G.T. Self-assembly of the hydrophobin SC3 proceeds via two structural intermediates. Protein Sci. 2002, 11, 1199–1205. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Accession | Substrate | Sampling Location | Accession(s) in Other Collections |
|---|---|---|---|---|
| USDA ARS Culture Collection (NRRL) | 13316 | Turf soil | Pennsylvania, USA | NA |
| 13826 | Carnation | California, USA | NA | |
| 36457 | Barley kernel | USA | CBS 409.86/FRC R-8509/IMI 309353 | |
| 54396 | Soil | Easter Lilly Research Borrkings, Oregon, USA | F49 | |
| 54754 | Corn | Pennsylvania, USA | A-28077 | |
| 66272 | Wheat | Washington, USA | A-28073 | |
| 66944 | Seedling of spruce | Pennsylvania, USA | A-28042 | |
| 66946 | Plant roots, Douglas fir tree | Oregon, USA | A-28020 | |
| 66947 | Seedling of Douglas fir | Pennsylvania, USA | A-28035 | |
| 66948 | Sugar pine tree seedling | Oregon, USA | A-28040 | |
| 66949 | Seedling of spruce | Pennsylvania, USA | A-28041 | |
| 66950 | Seedling of spruce | Pennsylvania, USA | A-28043 | |
| Kansas State University | 11440 | NA | NA | |
| Virginia Tech | F156N33 | Atmosphere | Virginia, USA |
| Strain | Coverage (×) | Assembly Size (bp) | Number of Contigs | Maximum Contig Length (bp) | Minimum Contig Length (bp) | Average Contig Length (bp) | Median Contig Length (bp) | N50 Contig Length (bp) | GC Content (%) | a Assembly BUSCO Coverage (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| F156N33 | 51 | 41,175,306 | 214 | 3,233,628 | 210 | 192,487 | 1075 | 1,472,944 | 48.44 | C:97.8; F:0.5; M:1.7 |
| 11440 | 52 | 42,933,485 | 897 | 1,984,632 | 200 | 48,011 | 538 | 1,024,532 | 48.33 | C:97.7; F:0.5; M:1.8 |
| NRRL 13316 | 61 | 41,704,585 | 964 | 2,164,424 | 200 | 43,399 | 579 | 843,661 | 48.26 | C:97.7; F:0.5; M:1.8 |
| NRRL 13826 | 51 | 44,694,304 | 1465 | 1,780,814 | 200 | 30,660 | 493 | 779,627 | 48.35 | C:97.6; F:0.5; M:1.9 |
| NRRL 36457 | 54 | 38,761,238 | 308 | 2,047,741 | 234 | 125,928 | 1068 | 918,031 | 48.38 | C:97.7; F:0.6; M:1.7 |
| NRRL 54396 | 51 | 49,692,405 | 626 | 1,709,761 | 200 | 79,506 | 659 | 691,401 | 50.72 | C:97.8; F:0.5; M:1.7 |
| NRRL 54754 | 50 | 38,889,550 | 234 | 4,244,225 | 229 | 166,303 | 875 | 1,329,635 | 48.48 | C:97.8; F:0.5; M:1.7 |
| NRRL 66272 | 49 | 39,140,173 | 251 | 2,646,538 | 226 | 156,028 | 1000 | 1,190,042 | 48.47 | C:97.6; F:0.5; M:1.9 |
| NRRL 66944 | 59 | 36,826,384 | 216 | 3,227,342 | 231 | 170,590 | 845 | 1,246,826 | 48.48 | C:97.6; F:0.4; M:2.0 |
| NRRL 66946 | 54 | 40,603,425 | 397 | 2,016,945 | 200 | 102,387 | 945 | 972,920 | 48.42 | C:97.7; F:0.5; M:1.8 |
| NRRL 66947 | 61 | 40,212,181 | 320 | 2,482,218 | 228 | 125,771 | 996 | 1,127,118 | 48.34 | C:97.9; F:0.5; M:1.6 |
| NRRL 66948 | 57 | 38,139,861 | 290 | 2,379,904 | 202 | 131,584 | 1103 | 1,046,193 | 48.31 | C:97.9; F:0.5; M:1.6 |
| NRRL 66949 | 52 | 37,127,996 | 260 | 3,604,089 | 234 | 142,881 | 1021 | 1,215,522 | 48.41 | C:97.7; F:0.4; M:1.9 |
| NRRL 66950 | 51 | 40,854,987 | 317 | 4,247,833 | 214 | 128,969 | 1117 | 1,141,783 | 48.38 | C:97.8; F:0.5; M:1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Rojas, M.; Coleman, J.J.; Vinatzer, B.A. Identification of Candidate Ice Nucleation Activity (INA) Genes in Fusarium avenaceum by Combining Phenotypic Characterization with Comparative Genomics and Transcriptomics. J. Fungi 2022, 8, 958. https://doi.org/10.3390/jof8090958
Yang S, Rojas M, Coleman JJ, Vinatzer BA. Identification of Candidate Ice Nucleation Activity (INA) Genes in Fusarium avenaceum by Combining Phenotypic Characterization with Comparative Genomics and Transcriptomics. Journal of Fungi. 2022; 8(9):958. https://doi.org/10.3390/jof8090958
Chicago/Turabian StyleYang, Shu, Mariah Rojas, Jeffrey J. Coleman, and Boris A. Vinatzer. 2022. "Identification of Candidate Ice Nucleation Activity (INA) Genes in Fusarium avenaceum by Combining Phenotypic Characterization with Comparative Genomics and Transcriptomics" Journal of Fungi 8, no. 9: 958. https://doi.org/10.3390/jof8090958
APA StyleYang, S., Rojas, M., Coleman, J. J., & Vinatzer, B. A. (2022). Identification of Candidate Ice Nucleation Activity (INA) Genes in Fusarium avenaceum by Combining Phenotypic Characterization with Comparative Genomics and Transcriptomics. Journal of Fungi, 8(9), 958. https://doi.org/10.3390/jof8090958