
Polyphasic Characterization of Four Aspergillus Species as Potential Biocontrol Agents for White Mold Disease of Bean

 ,
,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Plant Materials
2.2. Morphological Characterization of Aspergillus Isolates
2.3. Scanning Electron Microscopy (SEM)
2.4. Mycotoxin Analysis
2.5. Sequence Analysis
2.6. Biocontrol Assays
2.7. Data Analysis
3. Results
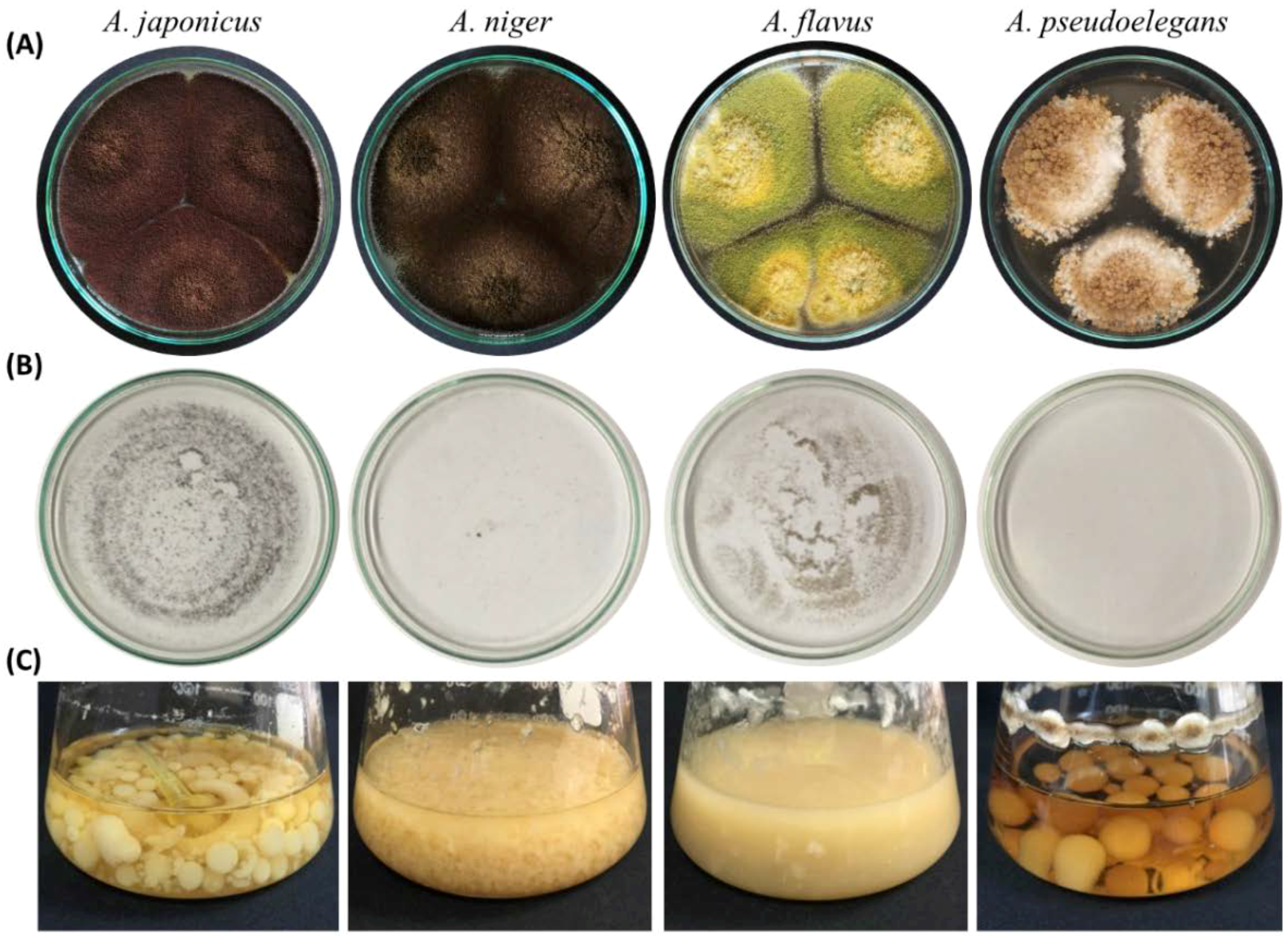
3.1. Polyphasic Characterization of Aspergillus Species
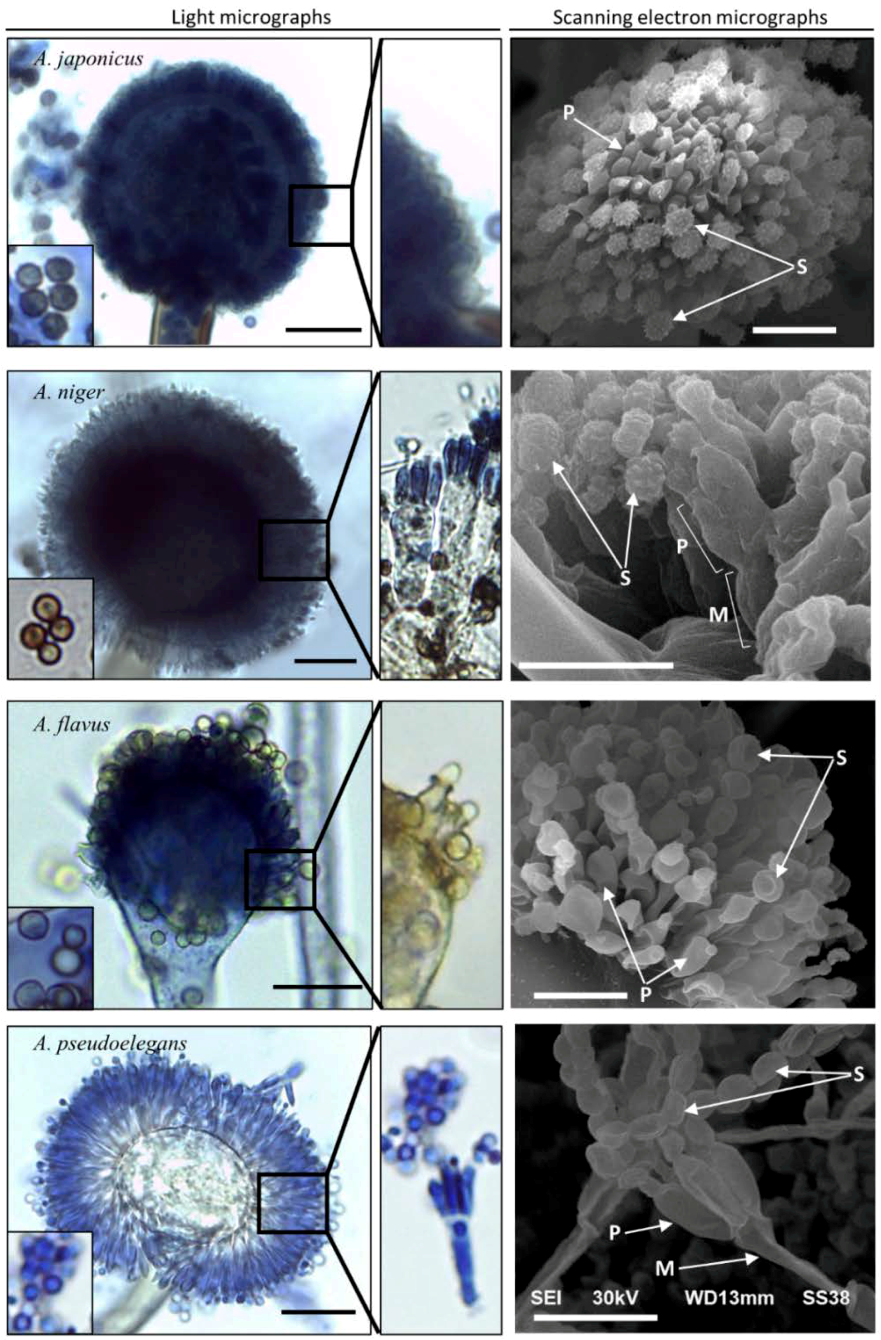
3.1.1. Microscopic Features of Aspergillus spp.
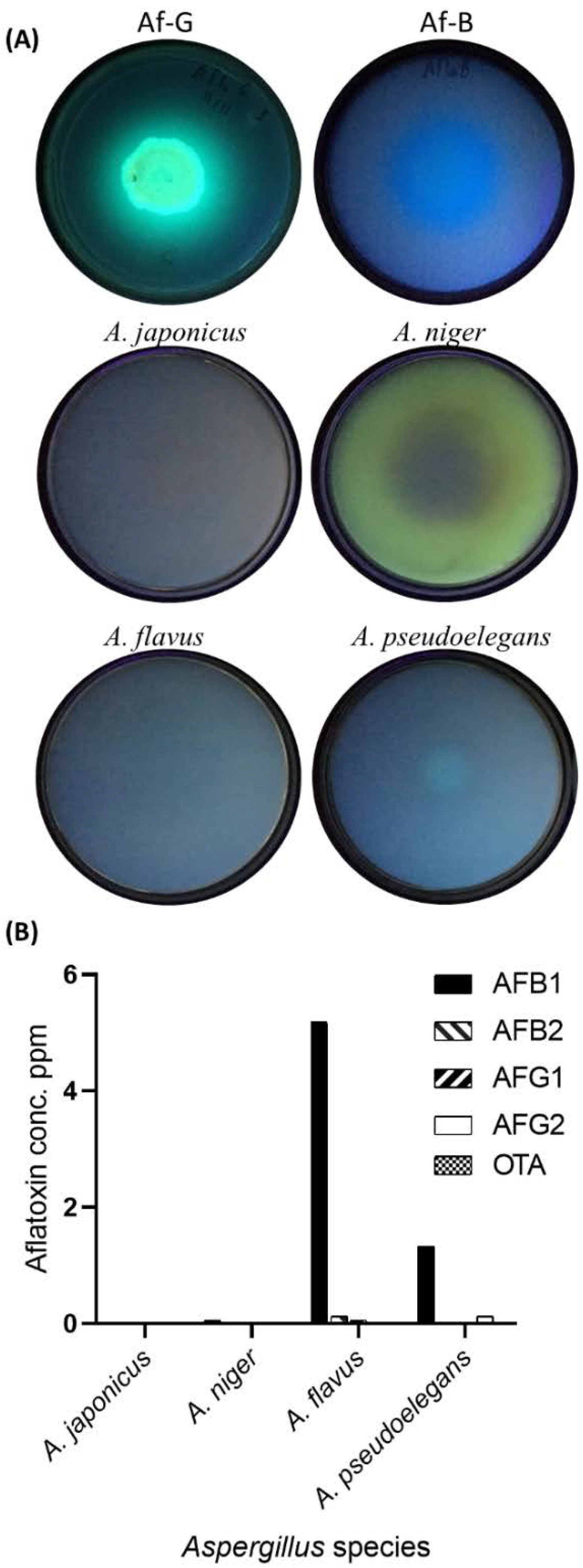
3.1.2. Mycotoxin Secretions by Aspergillus spp.
3.1.3. Genetic Variations within β-tubulin of Aspergillus spp.
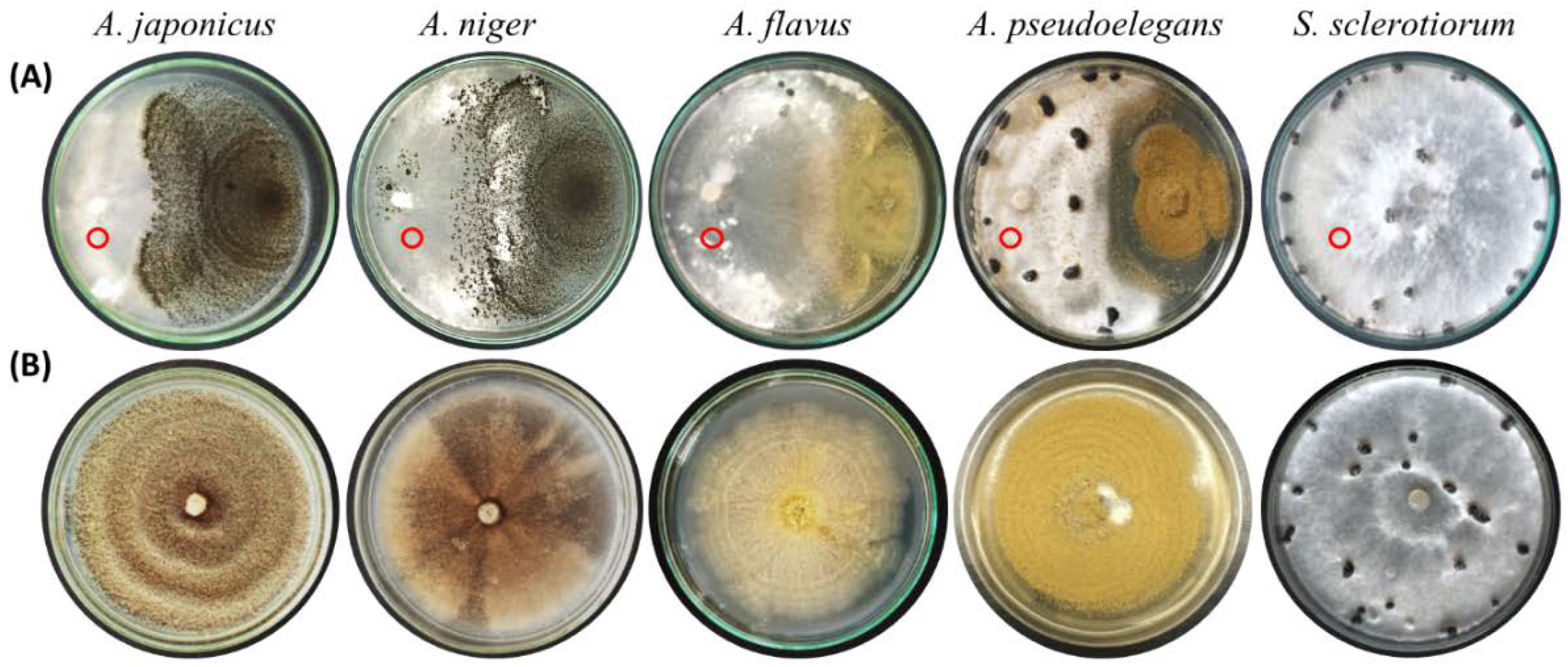
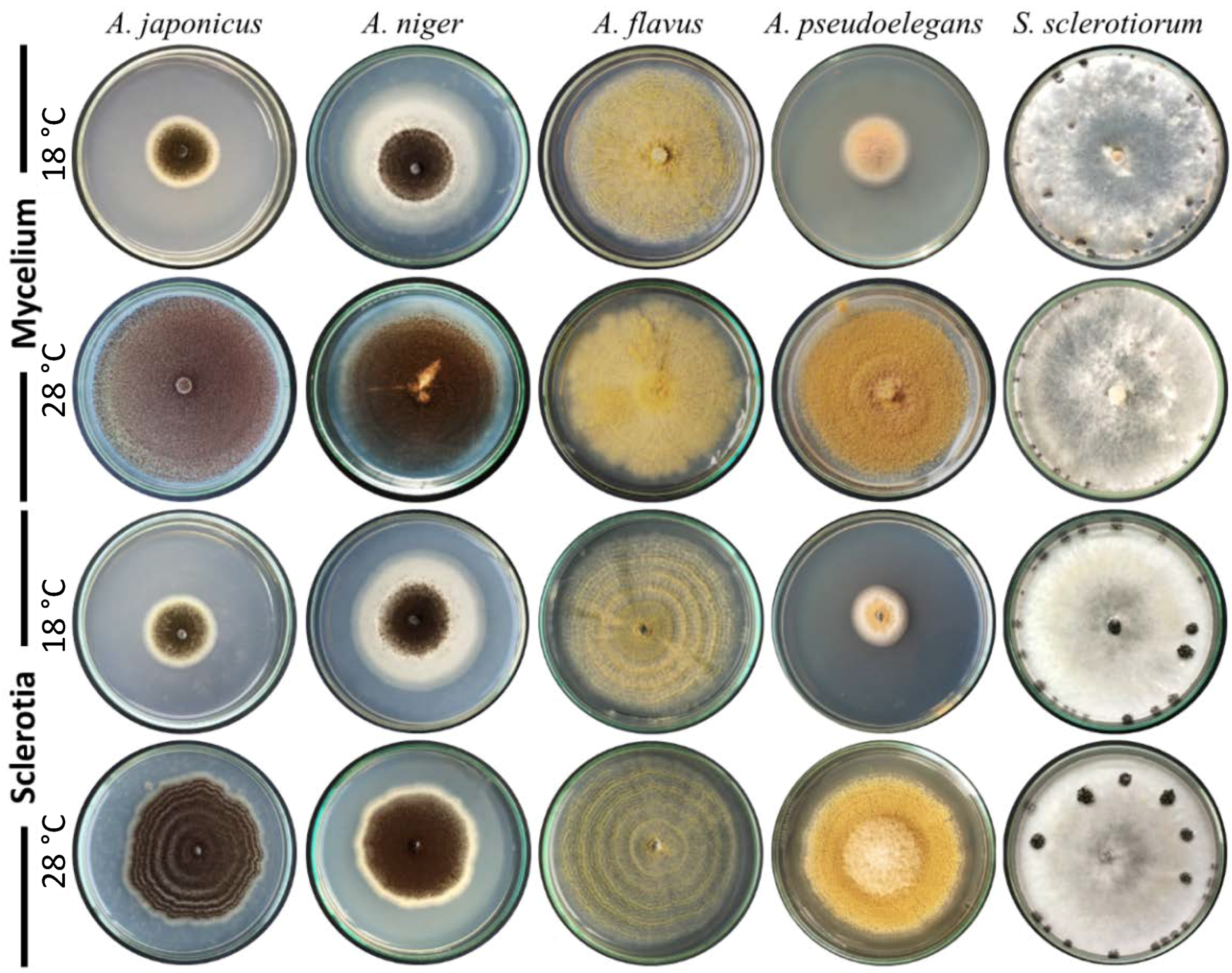
3.2. Aspergillus Deteriorates the Mycelium of S. sclerotiorum and Colonizes Its Sclerotia
3.3. Aspergillus spp. Retain Their Biocontrol Potential at a Wide Range of Temperature
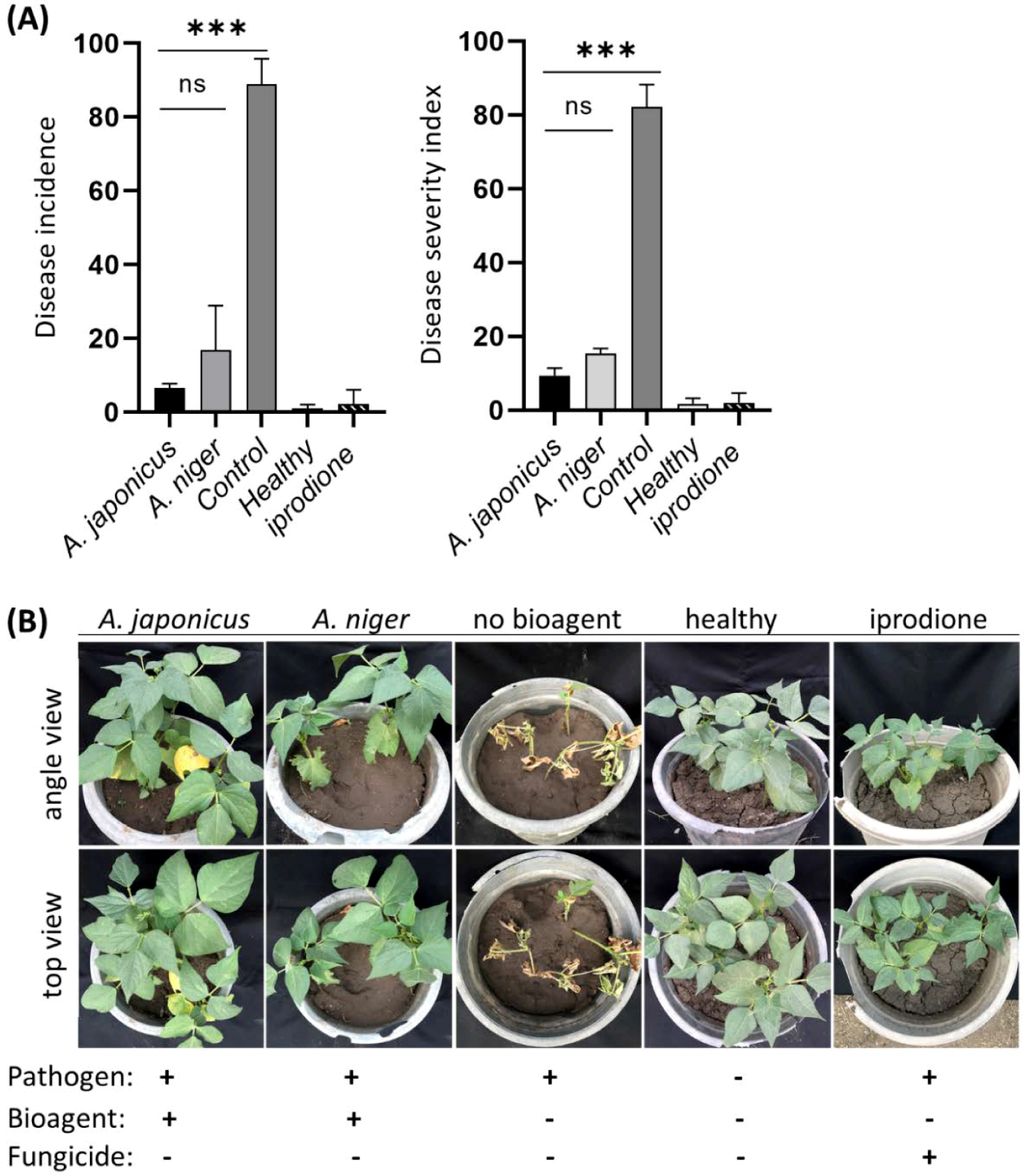
3.4. Aspergillus Suppresses White Mold Disease on Legumes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peltier, A.J.; Bradley, C.A.; Chilvers, M.I.; Malvick, D.K.; Mueller, D.S.; Wise, K.A.; Esker, P.D. Biology, Yield Loss and Control of Sclerotinia Stem Rot of Soybean. J. Integr. Pest Manag. 2012, 3, B1–B7. [Google Scholar] [CrossRef]
- Boland, G.J.; Hall, R. Index of Plant Hosts of Sclerotinia Sclerotiorum. Can. J. Plant Pathol. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Purdy, L.H. Sclerotinia Sclerotiorum: History, Diseases and Symptomatology, Host Range, Geographic Distribution, and Impact. Phytopathology 1979, 69, 875. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia Sclerotiorum (Lib.) de Bary: Biology and Molecular Traits of a Cosmopolitan Pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Willbur, J.; McCaghey, M.; Kabbage, M.; Smith, D.L. An Overview of the Sclerotinia Sclerotiorum Pathosystem in Soybean: Impact, Fungal Biology, and Current Management Strategies. Trop. Plant Pathol. 2019, 44, 3–11. [Google Scholar] [CrossRef]
- Liang, X.; Rollins, J.A. Mechanisms of Broad Host Range Necrotrophic Pathogenesis in Sclerotinia Sclerotiorum. Phytopathology 2018, 108, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.J.; Gent, D.H.; Pethybridge, S.J.; Hay, F.S. Site-Specific Risk Factors of White Mould Epidemics in Bean (Phaseolus Vulgaris) in Tasmania, Australia. N. Zeal. J. Crop Hortic. Sci. 2012, 40, 147–159. [Google Scholar] [CrossRef]
- Koenning, S.R.; Wrather, J.A. Suppression of Soybean Yield Potential in the Continental United States by Plant Diseases from 2006 to 2009. Plant Health Prog. 2010, 11, 5. [Google Scholar] [CrossRef]
- Vieira, R.F.; Paula Júnior, T.J.; Teixeira, H.; Carneiro, J.E.D.S. White Mold Management in Common Bean by Increasing within-Row Distance between Plants. Plant Dis. 2010, 94, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, J.P.; Staveley, J.; Phelps, K.; Young, C.S.; Whipps, J.M. Ascospore Release and Survival in Sclerotinia Sclerotiorum. Mycol. Res. 2003, 107, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Valencia, C.; Ferrera-Cerrato, R.; Quintanar-Zúñiga, R.E.; Flores-Ortiz, C.M.; Guzmán, G.J.M.; Alarcón, A.; Larsen, J.; García-Barradas, O. Morphological Development of Sclerotia by Sclerotinia Sclerotiorum: A View from Light and Scanning Electron Microscopy. Ann. Microbiol. 2015, 65, 765–770. [Google Scholar] [CrossRef]
- Butler, M.J.; Day, A.W. Fungal Melanins: A Review. Can. J. Microbiol. 1998, 44, 1115–1136. [Google Scholar] [CrossRef]
- Butler, M.J.; Gardiner, R.B.; Day, A.W. Degradation of Melanin or Inhibition of Its Synthesis: Are These a Significant Approach as a Biological Control of Phytopathogenic Fungi? Biol. Control 2005, 32, 326–336. [Google Scholar] [CrossRef]
- Butler, M.J.; Gardiner, R.B.; Day, A.W. Melanin Synthesis by Sclerotinia Sclerotiorum. Mycologia 2009, 101, 296–304. [Google Scholar] [CrossRef]
- Chen, J.; Ullah, C.; Reichelt, M.; Beran, F.; Yang, Z.; Gershenzon, J.; Hammerbacher, A.; Vassão, D.G. The Phytopathogenic Fungus Sclerotinia Sclerotiorum Detoxifies Plant Glucosinolate Hydrolysis Products via an Isothiocyanate Hydrolase. Nat. Commun. 2020, 11, 3090. [Google Scholar] [CrossRef] [PubMed]
- Adam, P.B.; Ayers, W.A. Ecology of Sclerotinia Species. Phytopathology 1979, 69, 896–899. [Google Scholar] [CrossRef]
- Adams, P.B. The Potential of Mycoparasites for Biological Control of Plant Diseases. Annu. Rev. Phytopathol. 1990, 28, 59–72. [Google Scholar] [CrossRef]
- Steadman, J.R. Control of Plant Disease Caused by Sclerotinia Species. Phytopathology 1979, 69, 904–907. [Google Scholar] [CrossRef]
- Jones, E.E.; Rabeendran, N.; Stewart, A. Biocontrol of Sclerotinia Sclerotiorum Infection of Cabbage by Coniothyrium Minitans and Trichoderma Spp. Biocontrol. Sci. Technol. 2014, 24, 1363–1382. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Brandan, C.P.; Petroselli, G.; Erra-Balsells, R.; Audisio, M.C. Biocontrol of Sclerotinia Sclerotiorum (Lib.) de Bary on Common Bean by Native Lipopeptide-Producer Bacillus Strains. Microbiol. Res. 2018, 211, 21–30. [Google Scholar] [CrossRef]
- Zhang, F.F.; Ge, H.; Zhang, F.F.; Guo, N.; Wang, Y.; Chen, L.; Ji, X.; Li, C. Biocontrol Potential of Trichoderma Harzianum Isolate T-Aloe against Sclerotinia Sclerotiorum in Soybean. Plant Physiol. Biochem. 2016, 100, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Roberts, D.P.; Xie, L.; Yu, C.; Li, Y.; Qin, L.; Hu, L.; Zhang, Y.; Liao, X. Biological Control of Sclerotinia Disease by Aspergillus Sp. on Oilseed Rape in the Field. Biocontrol. Sci. Technol. 2016, 26, 1526–1537. [Google Scholar] [CrossRef]
- Patibanda, A.K.; Sen, B. Mechanism of Aspergillus Niger van Teigh Antagonism towards Fusarium Oxysporum Schlect f Sp Melonis Sny & Hans, Muskmelon Wilt Pathogen. J. Biol. Control 2005, 19, 115–120. [Google Scholar]
- Israel, S.; Lodha, S. Biological Control of Fusarium Oxysporum f. Sp. Cumini with Aspergillus Versicolor. Phytopathol. Mediterr. 2005, 44, 3–11. [Google Scholar]
- Adebola, M.; Amadi, J. Screening Three Aspergillus Species for Antagonistic Activities against the Cocoa Black Pod Organism (Phytophthora Palmivora). Agric. Biol. J. N. Am. 2010, 1, 362–365. [Google Scholar] [CrossRef]
- Jang, J.Y.; Choi, Y.H.; Shin, T.S.; Kim, T.H.; Shin, K.S.; Park, H.W.; Kim, Y.H.; Kim, H.; Choi, G.J.; Jang, K.S.; et al. Biological Control of Meloidogyne Incognita by Aspergillus Niger F22 Producing Oxalic Acid. PLoS ONE 2016, 11, e0156230. [Google Scholar] [CrossRef] [PubMed]
- El-Debaiky, S.A. Antagonistic Studies and Hyphal Interactions of the New Antagonist Aspergillus Piperis against Some Phytopathogenic Fungi in Vitro in Comparison with Trichoderma Harzianum. Microb. Pathog. 2017, 113, 135–143. [Google Scholar] [CrossRef]
- Alshannaq, A.F.; Gibbons, J.G.; Lee, M.K.; Han, K.H.; Hong, S.B.; Yu, J.H. Controlling Aflatoxin Contamination and Propagation of Aspergillus Flavus by a Soy-Fermenting Aspergillus Oryzae Strain. Sci. Rep. 2018, 8, 16871. [Google Scholar] [CrossRef]
- Boughalleb-M’Hamdi, N.; Salem, I.B.; M’Hamdi, M. Evaluation of the Efficiency of Trichoderma, Penicillium, and Aspergillus Species as Biological Control Agents against Four Soil-Borne Fungi of Melon and Watermelon. Egypt. J. Biol. Pest Control 2018, 28, 25. [Google Scholar] [CrossRef]
- Jin, N.; Liu, S.M.; Peng, H.; Huang, W.K.; Kong, L.A.; Wu, Y.H.; Chen, Y.P.; Ge, F.Y.; Jian, H.; Peng, D.L. Isolation and Characterization of Aspergillus Niger NBC001 Underlying Suppression against Heterodera Glycines. Sci. Rep. 2019, 9, 591. [Google Scholar] [CrossRef]
- Melo, I.S.; Faull, J.L.; Nascimento, R.S. Antagonism of Aspergillus Terreus to Sclerotinia Sclerotiorum. Braz. J. Microbiol. 2006, 37, 417–419. [Google Scholar] [CrossRef]
- Hu, X.; Webster, G.; Xie, L.; Yu, C.; Li, Y.; Liao, X. A New Mycoparasite, Aspergillus sp. ASP-4, Parasitizes the Sclerotia of Sclerotinia Sclerotiorum. Crop. Prot. 2013, 54, 15–22. [Google Scholar] [CrossRef]
- Hu, X.; Qin, L.; Roberts, D.P.; Lakshman, D.K.; Gong, Y.; Maul, J.E.; Xie, L.; Yu, C.; Li, Y.; Hu, L.; et al. Characterization of Mechanisms Underlying Degradation of Sclerotia of Sclerotinia Sclerotiorum by Aspergillus Aculeatus Asp-4 Using a Combined QRT-PCR and Proteomic Approach. BMC Genom. 2017, 18, 674. [Google Scholar] [CrossRef] [PubMed]
- Idan, A.A.; Sijam, K.; Kadir, J.; Rashid, T.S.; Awla, H.K.; Alsultan, W. Biological Control of Pyricularia Oryzae Using Antifungal Compounds Produced by Aspergillus Niger. Am. J. Plant Sci. 2017, 08, 2445–2460. [Google Scholar] [CrossRef]
- Gao, T.; Hao, F.; Yang, D.; Bie, Z.; Li, G. Oxalic Acid Produced by Aspergillus Niger Y-1 Is Effective for Suppression of Bacterial Fruit Blotch of Watermelon Seedlings. Biol. Control 2017, 112, 28–33. [Google Scholar] [CrossRef]
- Xing, Y.M.; Yin, W.Q.; Liu, M.M.; Wang, C.L.; Guo, S.X. Oxalic Acid and Sclerotial Differentiation of Polyporus Umbellatus. Sci. Rep. 2015, 5, srep10759. [Google Scholar] [CrossRef]
- Atallah, O.; Yassin, S. Aspergillus Spp. Eliminate Sclerotinia Sclerotiorum by Imbalancing the Ambient Oxalic Acid Concentration and Parasitizing Its Sclerotia. Environ. Microbiol. 2020, 22, 5265–5279. [Google Scholar] [CrossRef]
- Cray, J.A.; Bell, A.N.W.; Bhaganna, P.; Mswaka, A.Y.; Timson, D.J.; Hallsworth, J.E. The Biology of Habitat Dominance; Can Microbes Behave as Weeds? Microb. Biotechnol. 2013, 6, 453–492. [Google Scholar] [CrossRef]
- Šimonovičová, A.; Vojtková, H.; Nosalj, S.; Piecková, E.; Švehláková, H.; Kraková, L.; Drahovská, H.; Stalmachová, B.; Kučová, K.; Pangallo, D. Aspergillus Niger Environmental Isolates and Their Specific Diversity Through Metabolite Profiling. Front. Microbiol. 2021, 12, 658010. [Google Scholar] [CrossRef] [PubMed]
- Nadumane, V.K.; Venkatachalam, P.; Gajaraj, B. Chapter 19-Aspergillus Applications in Cancer Research. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 243–255. ISBN 978-0-444-63505-1. [Google Scholar]
- Sun, B.T.; Akutse, K.S.; Xia, X.F.; Chen, J.H.; Ai, X.; Tang, Y.; Wang, Q.; Feng, B.W.; Goettel, M.S.; You, M.S. Endophytic Effects of Aspergillus Oryzae on Radish (Raphanus Sativus) and Its Herbivore, Plutella Xylostella. Planta 2018, 248, 705–714. [Google Scholar] [CrossRef]
- Backman, P.A.; Sikora, R.A. Endophytes: An Emerging Tool for Biological Control. Biol. Control 2008, 46, 1–3. [Google Scholar] [CrossRef]
- Lubna; Asaf, S.; Hamayun, M.; Gul, H.; Lee, I.J.; Hussain, A. Aspergillus Niger CSR3 Regulates Plant Endogenous Hormones and Secondary Metabolites by Producing Gibberellins and Indoleacetic. Acid. J. Plant Interact. 2018, 13, 100–111. [Google Scholar] [CrossRef]
- Hamayun, M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I. Endophytic Fungus Aspergillus Japonicus Mediates Host Plant Growth under Normal and Heat Stress Conditions. Biomed Res. Int. 2018, 3, 7696831. [Google Scholar]
- Ojewumi, M.E.; Anenih, E.V.; Taiwo, O.S.; Adekeye, B.T.; Awolu, O.O.; Ojewumi, E.O. A Bioremediation Study of Raw and Treated Crude Petroleum Oil Polluted Soil with Aspergillus Niger and Pseudomonas Aeruginosa. J. Ecol. Eng. 2018, 19, 226–235. [Google Scholar] [CrossRef]
- Ren, W.X.; Li, P.J.; Geng, Y.; Li, X.J. Biological Leaching of Heavy Metals from a Contaminated Soil by Aspergillus Niger. J. Hazard. Mater. 2009, 167, 164–169. [Google Scholar] [CrossRef]
- Gulzar, T.; Huma, T.; Jalal, F.; Iqbal, S.; Abrar, S.; Kiran, S.; Nosheen, S.; Hussain, W.; Rafique, M.A. Bioremediation of Synthetic and Industrial Effluents by Aspergillus Niger Isolated from Contaminated Soil Following a Sequential Strategy. Molecules 2017, 22, 2244. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped Potential: Exploiting Fungi in Bioremediation of Hazardous Chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Yang, L.; Lübeck, M.; Lübeck, P.S. Aspergillus as a Versatile Cell Factory for Organic Acid Production. Fungal. Biol. Rev. 2017, 31, 33–49. [Google Scholar] [CrossRef]
- Wakai, S.; Arazoe, T.; Ogino, C.; Kondo, A. Future Insights in Fungal Metabolic Engineering. Bioresour. Technol. 2017, 245, 1314–1326. [Google Scholar] [CrossRef]
- De Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative Genomics Reveals High Biological Diversity and Specific Adaptations in the Industrially and Medically Important Fungal Genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef]
- McClenny, N. Laboratory Detection and Identification of Aspergillus Species by Microscopic Observation and Culture: The Traditional Approach. Med. Mycol. 2005, 43, 125–128. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, Identification and Nomenclature of the Genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus Section Flavi and Their Production of Aflatoxins, Ochratoxins and Other Mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Visagie, C.M.; Varga, J.; Houbraken, J.; Meijer, M.; Kocsubé, S.; Yilmaz, N.; Fotedar, R.; Seifert, K.A.; Frisvad, J.C.; Samson, R.A. Ochratoxin Production and Taxonomy of the Yellow Aspergilli (Aspergillus Section Circumdati). Stud. Mycol. 2014, 78, 1–61. [Google Scholar] [CrossRef]
- Atallah, O.O.; Osman, A.; Ali, M.A.S.; Sitohy, M. Soybean β-Conglycinin and Catfish Cutaneous Mucous P22 Glycoproteins Deteriorate Sporangial Cell Walls of Pseudoperonospora Cubensis and Suppress Cucumber Downy Mildew. Pest Manag. Sci. 2021, 77, 3313–3324. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Bozzola, J.J.; Russell, L.D. Electron Microscopy: Principles and Techniques for Biologists, 2nd ed.; Jones and Bartlett: Boston, MA, USA, 1999; ISBN 978-0763701925. [Google Scholar]
- Kuo, J. Electron Microscopy: Methods and Protocols. In Methods in Molecular Biology, 2nd ed.; Kuo, J., Ed.; Humana Press: Totowa, NJ, USA, 2007; Volume 369, p. 624. ISBN 978-1588295736. [Google Scholar]
- Lemke, P.A.; Davis, D.; Iyer, S.K.; Creech, G.W. Fluorometric Analysis of Iodinated Aflatoxin in Minicultures of Aspergillus Flavus and Aspergillus Parasiticus. J. Ind. Microbiol. 1988, 3, 119–125. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Yassin, S.M.; Aly, A.Z.; Abdel-Kader, D.A.; Morsy, K.M.; Atallah, O.O. Antagonistic Potential of Rhizospheric Biocontrol Agents against Soybean Root Rot- Wilt Disease Complex Syndrome. Zagazig J. Agric. Res. 2019, 46, 1395–1418. [Google Scholar] [CrossRef]
- Abdelrhim, A.S.; Mazrou, Y.S.A.; Nehela, Y.; Atallah, O.O.; El-ashmony, R.M.; Dawood, M.F.A. Silicon Dioxide Nanoparticles Induce Innate Immune Responses and Activate Antioxidant Machinery in Wheat against Rhizoctonia Solani. Plants 2021, 10, 2758. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, X.; Chen, Y.; Qin, H.; Huang, L.; Han, Q. Inhibitory Efficacy of Endophytic Bacillus Subtilis EDR4 against Sclerotinia Sclerotiorum on Rapeseed. Biol. Control 2014, 78, 67–76. [Google Scholar] [CrossRef]
- Silva, D.M.; Batista, L.R.; Rezende, E.F.; Fungaro MH, P.; Sartori, D.; Alves, E. Identification of Fungi of the Genus Aspergillus Section Nigri Using Polyphasic Taxonomy. Braz. J. Microbiol. 2011, 42, 761–773. [Google Scholar] [CrossRef]
- Nyongesa, B.W.; Okoth, S.; Ayugi, V. Identification Key for Aspergillus Species Isolated from Maize and Soil of Nandi County, Kenya. Adv. Microbiol. 2015, 5, 205–229. [Google Scholar] [CrossRef]
- Hegedus, D.D.; Rimmer, S.R. Sclerotinia Sclerotiorum: When “to Be or Not to Be” a Pathogen? FEMS Microbiol. Lett. 2005, 251, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Gerlagh, M.; Goossen-Van De Geijn, H.M.; Fokkema, N.J.; Vereijken, P.F.G. Long-Term Biosanitation by Application of Coniothyrium Minitans on Sclerotinia Sclerotiorum-Infected Crops. Phytopathology 1999, 89, 141–147. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, D.; Kirk, W.; Hao, J. Use of Coniothyrium Minitans and Other Microorganisms for Reducing Sclerotinia Sclerotiorum. Biol. Control 2012, 60, 225–232. [Google Scholar] [CrossRef]
- Meijer, M.; Houbraken, J.A.M.P.; Dalhuijsen, S.; Samson, R.A.; de Vries, R.P. Growth and Hydrolase Profiles Can Be Used as Characteristics to Distinguish Aspergillus Niger and other Black Aspergilli. Stud. Mycol. 2011, 69, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Pařenicová, L.; Skouboe, P.; Frisvad, J.; Samson, R.A.; Rossen, L.; Ten Hoor-Suykerbuyk, M.; Visser, J. Combined Molecular and Biochemical Approach Identifies Aspergillus Japonicus and Aspergillus Aculeatus as Two Species. Appl. Environ. Microbiol. 2001, 67, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Frisvad, J.C.; Kocsubé, S.; Brankovics, B.; Tóth, B.; Szigeti, G.; Samson, R.A. New and Revisited Species in Aspergillus Section Nigri. Stud. Mycol. 2011, 69, 1–17. [Google Scholar] [CrossRef]
- Dyer, P.S.; O’Gorman, C.M. Sexual Development and Cryptic Sexuality in Fungi: Insights from Aspergillus Species. FEMS Microbiol. Rev. 2012, 36, 165–192. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Petersen, L.M.; Lyhne, E.K.; Larsen, T.O. Formation of Sclerotia and Production of Indoloterpenes by Aspergillus Niger and Other Species in Section. PLoS ONE 2014, 9, e94857. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, T.R.; Nielsen, K.F.; Arentshorst, M.; Park, J.; van den Hondel, C.A.; Frisvad, J.C.; Ram, A.F. Submerged Conidiation and Product Formation by Aspergillus Niger at Low Specific Growth Rates Are Affected in Aerial Developmental Mutants. Appl. Environ. Microbiol. 2011, 77, 5270–5277. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rajendran, C.; Muthappa, B.N. Saitoa, a New Genus of Plectomycetes. Proc. Plant Sci. 1980, 89, 185–191. [Google Scholar] [CrossRef]
- Dutcher, R.J.D. Aspergillic Acid; an Antibiotic Substance Produced by Aspergillus Flavus. J. Biol. Chem. 1958, 232, 785–795. [Google Scholar] [CrossRef]
- Uka, V.; Moore, G.G.; Arroyo-Manzanares, N.; Nebija, D.; De Saeger, S.; Diana Di Mavungu, J. Secondary Metabolite Dereplication and Phylogenetic Analysis Identify Various Emerging Mycotoxins and Reveal the High Intra-Species Diversity in Aspergillus Flavus. Front. Microbiol. 2019, 10, 667. [Google Scholar] [CrossRef]
- Ola, A.R.B.; Metboki, G.; Lay, C.S.; Sugi, Y.; Rozari, P.; De Darmakusuma, D.; Hakim, E.H. Single Production of Kojic Acid by Aspergillus Flavus and the Revision of Flufuran. Molecules 2019, 24, 4200. [Google Scholar] [CrossRef] [PubMed]
- Dutcher, J.D. Aspergillic Acid: An Antibiotic Substance Produced by Aspergillus Flavus: I. General Properties; Formation of Desoxyaspergillic Acid; Structural Conclusions. J. Biol. Chem. 1947, 171, 321–339. [Google Scholar] [CrossRef]
- Vijayalakshmi, P.; Durgadevi, K.B. A Study on Anti-Fungal Activity of Kojic Acid Synthesized by Aspergillus Flavus FJ537130 Strain Isolated from Peanut Soil. Asian J. Microbiol. Biotechnol. Environ. Sci. 2017, 19, 160–164. [Google Scholar]
- Etxebeste, O.; Garzia, A.; Espeso, E.A.; Ugalde, U. Aspergillus Nidulans Asexual Development: Making the Most of Cellular Modules. Trends. Microbiol. 2010, 18, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.P. Effect of Light on Growth and Sporulation of Aspergillus Ornatus. J. Gen. Microbiol. 1976, 95, 39–44. [Google Scholar] [CrossRef]
- Mooney, J.L.; Yager, L.N. Light Is Required for Conidiation in Aspergillus Nidulans. Genes Dev. 1990, 4, 1473–1482. [Google Scholar] [CrossRef]
- Murthy, P.S.; Suzuki, S.; Kusumoto, K.-I. Effect of Light on the Growth and Acid Protease Production of Aspergillus Oryzae. Food Sci. Technol. Res. 2015, 21, 631–635. [Google Scholar] [CrossRef]
- Hedtke, M.; Rauscher, S.; Röhrig, J.; Rodríguez-Romero, J.; Yu, Z.; Fischer, R. Light-Dependent Gene Activation in Aspergillus Nidulans Is Strictly Dependent on Phytochrome and Involves the Interplay of Phytochrome and White Collar-Regulated Histone H3 Acetylation. Mol. Microbiol. 2015, 97, 733–745. [Google Scholar] [CrossRef]
- Sun, W.; Yu, Y.; Chen, J.; Yu, B.; Chen, T.; Ying, H.; Zhou, S.; Ouyang, P.; Liu, D.; Chen, Y. Light Signaling Regulates Aspergillus Niger Bio Fi Lm Formation By. Mol. Biol. Physiol. 2021, 12, 1–13. [Google Scholar]
- Stinnett, S.M.; Espeso, E.A.; Cobeño, L.; Araújo-Bazán, L.; Calvo, A.M. Aspergillus Nidulans VeA Subcellular Localization Is Dependent on the Importin Alpha Carrier and on Light. Mol. Microbiol. 2007, 63, 242–255. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Yang, L.; Zhang, L.; Jiang, D.; Chen, W.; Li, G. Diversity and Biocontrol Potential of Endophytic Fungi in Brassica Napus. Biol. Control 2014, 72, 98–108. [Google Scholar] [CrossRef]
- Verma, A.; Johri, B.N.; Prakash, A. Antagonistic Evaluation of Bioactive Metabolite from Endophytic Fungus, Aspergillus Flavipes KF671231. J. Mycol. 2014, 2014, 371218. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, Y.; Liu, R.; Zhou, F. Endophytic Fungus Aspergillus Tamarii from Ficus Carica L., a New Source of Indolyl Diketopiperazines OH. Biochem. Syst. Ecol. 2012, 45, 31–33. [Google Scholar] [CrossRef]
- Qin, J.; Lyu, A.; Zhang, Q.H.; Yang, L.; Zhang, J.; Wu, M.D.; Li, G.Q. Strain Identification and Metabolites Isolation of Aspergillus Capensis CanS-34A from Brassica Napus. Mol. Biol. Rep. 2019, 46, 3451–3460. [Google Scholar] [CrossRef]
- Li, X.J.; Zhang, Q.; Zhang, A.L.; Gao, J.M. Metabolites from Aspergillus Fumigatus, an Endophytic Fungus Associated with Melia Azedarach, and Their Antifungal, Antifeedant, and Toxic Activities. J. Agric. Food Chem. 2012, 60, 3424–3431. [Google Scholar] [CrossRef] [PubMed]
- Goutam, J.; Singh, S.; Kharwar, R.N.; Ramarai, V. In Vitro Potential of Endophytic Fungus Aspergillus Terrus (JAS-2) Associated with Achyranthus Aspera and Study on Its Culture Conditions. Biol. Med. 2016, 8, 7. [Google Scholar] [CrossRef]
- Chaves, F.C.; Gianfagna, T.J.; Aneja, M.; Posada, F.; Peterson, S.W.; Vega, F.E. Aspergillus Oryzae NRRL 35191 from Coffee, a Non-Toxigenic Endophyte with the Ability to Synthesize Kojic Acid. Mycol. Prog. 2012, 11, 263–267. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPLC Parameters | AFB1 | AFB2 | AFG1 | AFG2 | OTA |
|---|---|---|---|---|---|
| Retention time (min) | 5.1–7.1 | 12.9–13.2 | 5.2–5.5 | 8.7–9 | 6.7 |
| Excitation Wavelength | 365 | 365 | 365 | 365 | 335 |
| Emission Wavelength | 450 | 450 | 450 | 450 | 465 |
| Colony Color | Colony Diameter at 28 °C/7 dpi | Sporulation at 18 °C/Dark | Seriation Type | Spore Liberation | Sclerotia on PDA | |
|---|---|---|---|---|---|---|
| A. japonicus | Coffee brown | 90 mm | Normal | Uniseriate | Dense | Absent |
| A. niger | Dark brown | 90 mm | Absent | Biseriate | Light | Absent |
| A. flavus | Yellow green | 90 mm at 5 dpi | Normal | Uniseriate | Dense | Absent |
| A. pseudoelegans | Light brown | 50 mm | Absent | Biseriate | Light | Absent |
| Head Diameter (µm) | Vesicle Diameter (µm) | Spore Diameter (µm) | Conidiophore Width (µm) | Phialide Length (µm) | Spore Surface | |
|---|---|---|---|---|---|---|
| A. japonicus | 33.9 | 25.2 | 2.11 | 7.2 | 4.23 | Rough |
| A. niger | 38.8 | 20.6 | 1.606 | 6.7 | 4.5 | Rough |
| A. flavus | 25.8 | 18.1 | 2.64 | 4.8 | 3.9 | Smooth |
| A. pseudoelegans | 40.2 | 17.4 | 1.343 | 5.2 | 5 | Smooth |
| No. of replicates | 50 | 50 | 100 | 50 | 50 | 100 |
| DI | DS Index | Biocontrol Efficiency | |
|---|---|---|---|
| A. japonicus | 6.5 | 9.3 | 88.5 |
| A. niger | 16.8 | 15.3 | 81.4 |
| Control | 88.9 | 82.1 | N/A |
| Healthy | 1.0 | 1.7 | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atallah, O.O.; Mazrou, Y.S.A.; Atia, M.M.; Nehela, Y.; Abdelrhim, A.S.; Nader, M.M. Polyphasic Characterization of Four Aspergillus Species as Potential Biocontrol Agents for White Mold Disease of Bean. J. Fungi 2022, 8, 626. https://doi.org/10.3390/jof8060626
Atallah OO, Mazrou YSA, Atia MM, Nehela Y, Abdelrhim AS, Nader MM. Polyphasic Characterization of Four Aspergillus Species as Potential Biocontrol Agents for White Mold Disease of Bean. Journal of Fungi. 2022; 8(6):626. https://doi.org/10.3390/jof8060626
Chicago/Turabian StyleAtallah, Osama O., Yasser S. A. Mazrou, Mahmoud M. Atia, Yasser Nehela, Abdelrazek S. Abdelrhim, and Maha M. Nader. 2022. "Polyphasic Characterization of Four Aspergillus Species as Potential Biocontrol Agents for White Mold Disease of Bean" Journal of Fungi 8, no. 6: 626. https://doi.org/10.3390/jof8060626
APA StyleAtallah, O. O., Mazrou, Y. S. A., Atia, M. M., Nehela, Y., Abdelrhim, A. S., & Nader, M. M. (2022). Polyphasic Characterization of Four Aspergillus Species as Potential Biocontrol Agents for White Mold Disease of Bean. Journal of Fungi, 8(6), 626. https://doi.org/10.3390/jof8060626