
Neopestalotiopsis siciliana sp. nov. and N. rosae Causing Stem Lesion and Dieback on Avocado Plants in Italy

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Characterization
2.2. DNA Extraction, PCR, and Phylogenetic Analysis
2.3. Pathogenicity Test
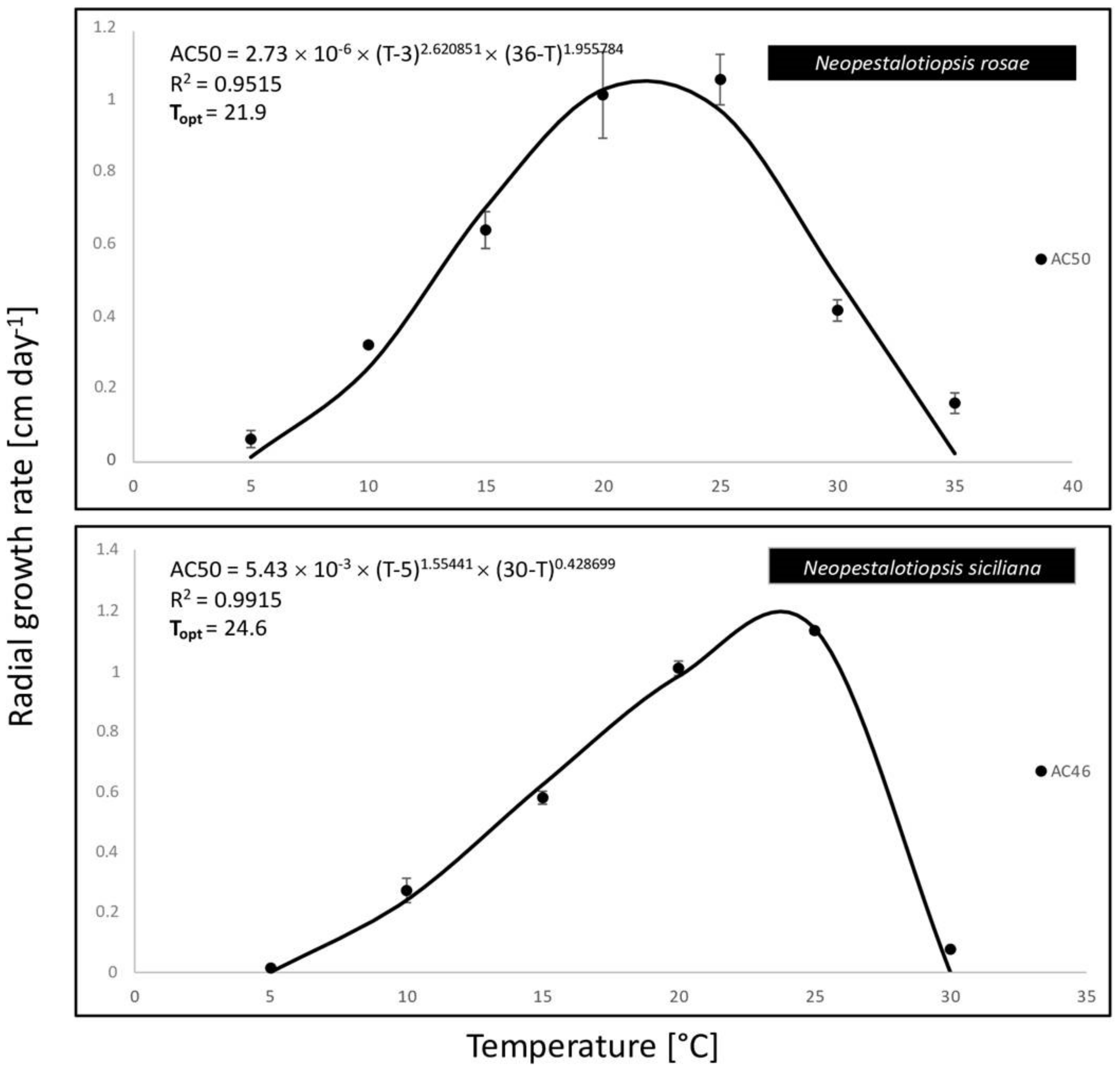
2.4. Data Analysis
3. Results
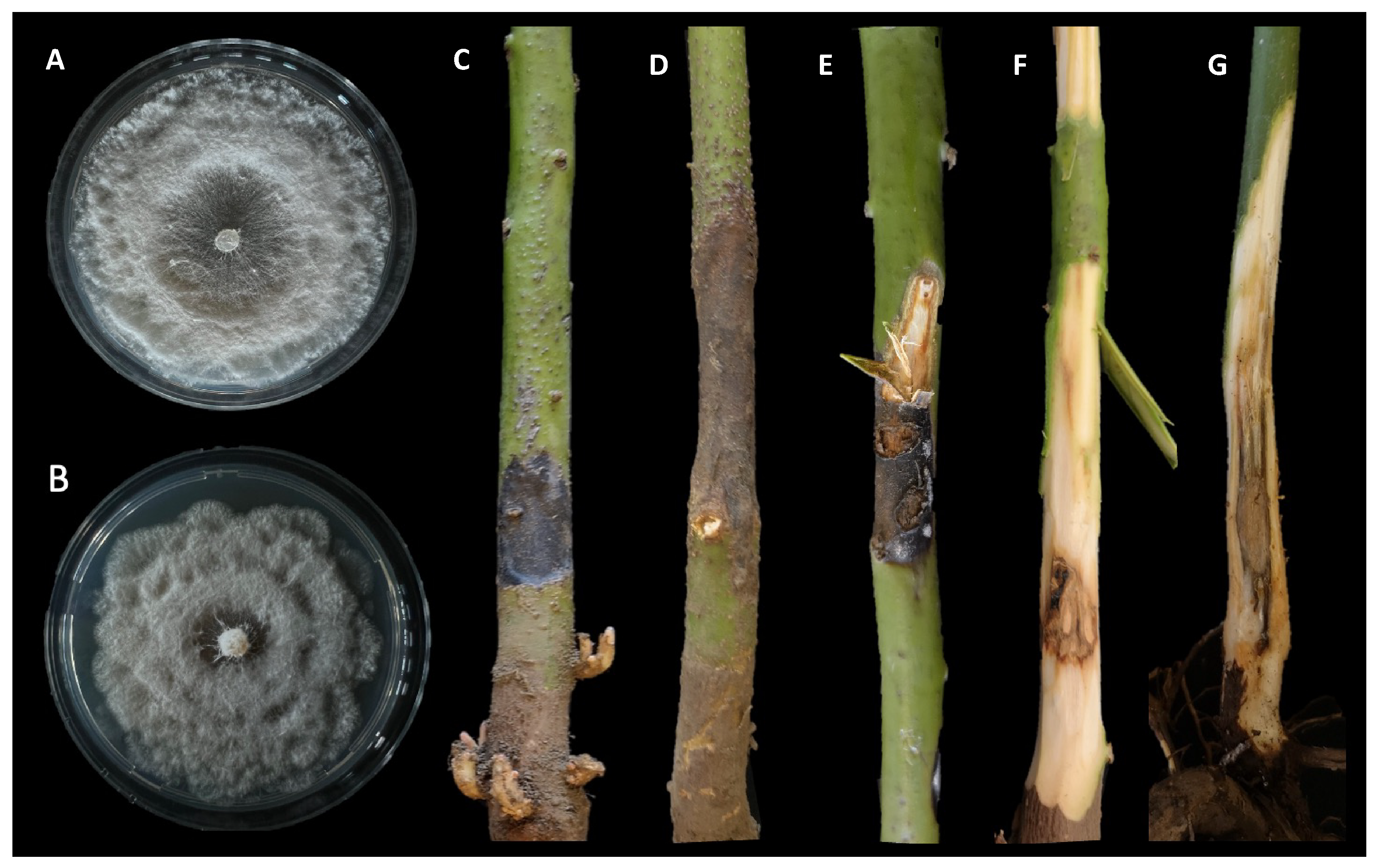
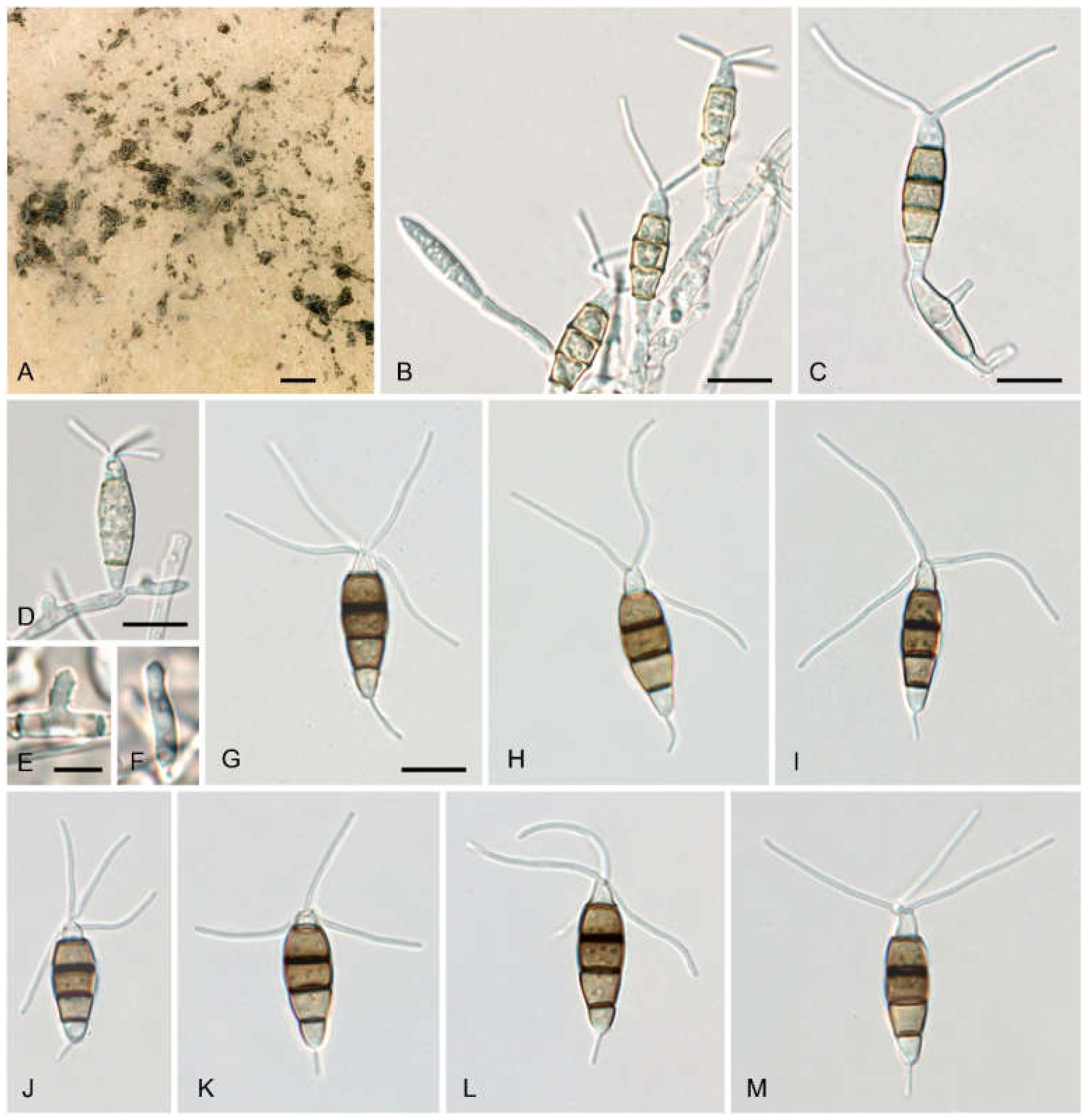
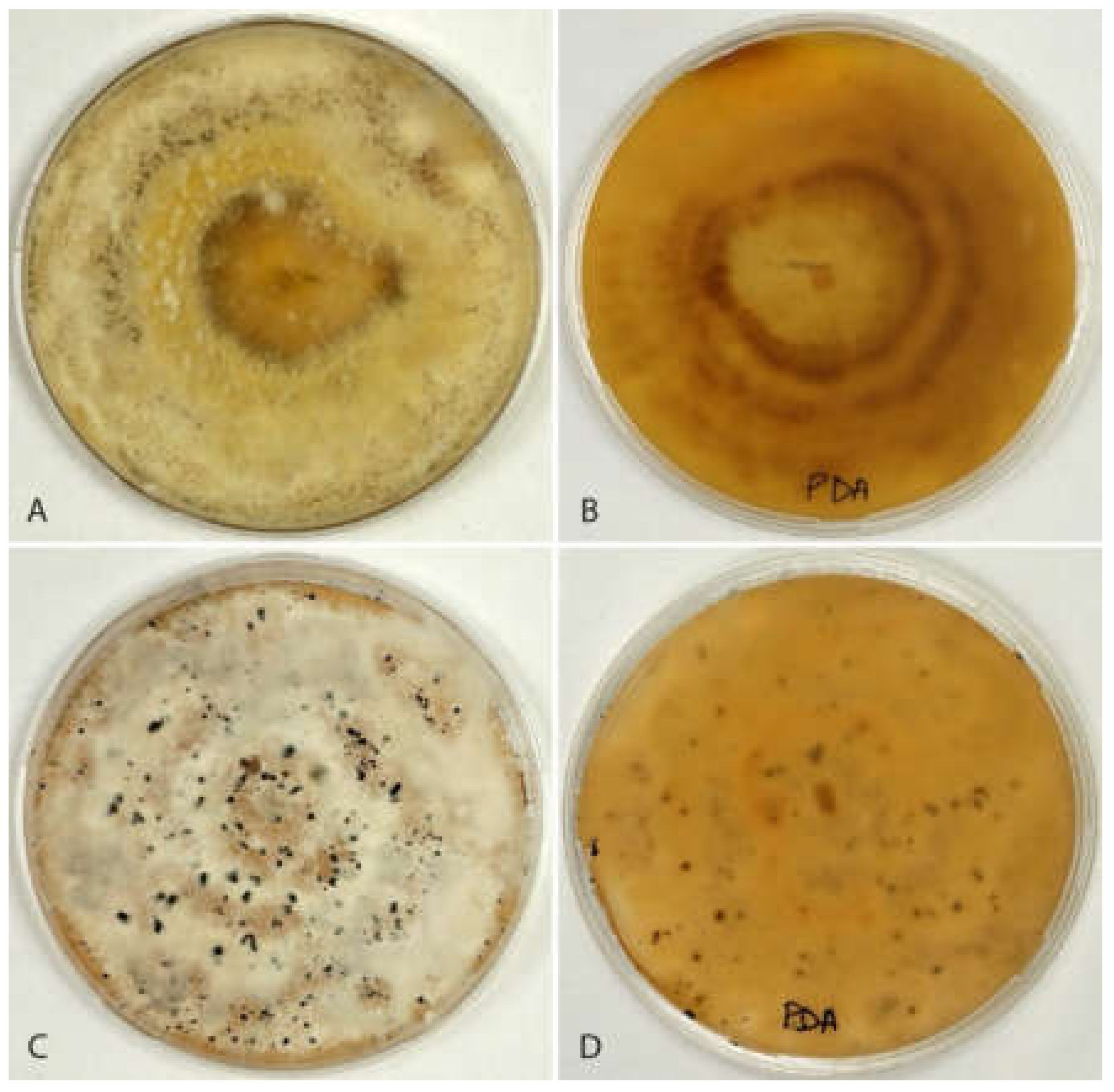
3.1. Isolation and Morphological Characterization
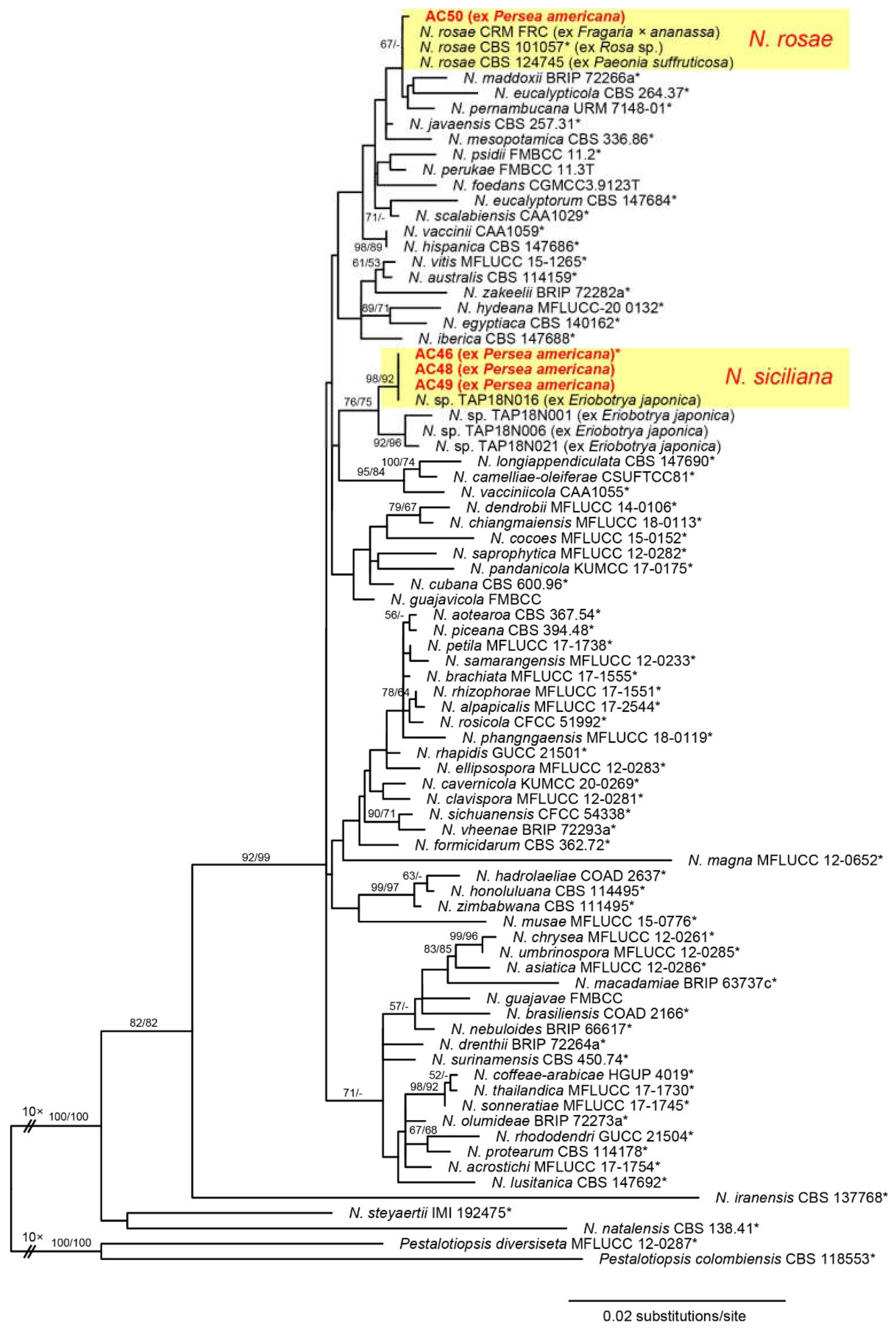
3.2. Phylogenetic Analysis
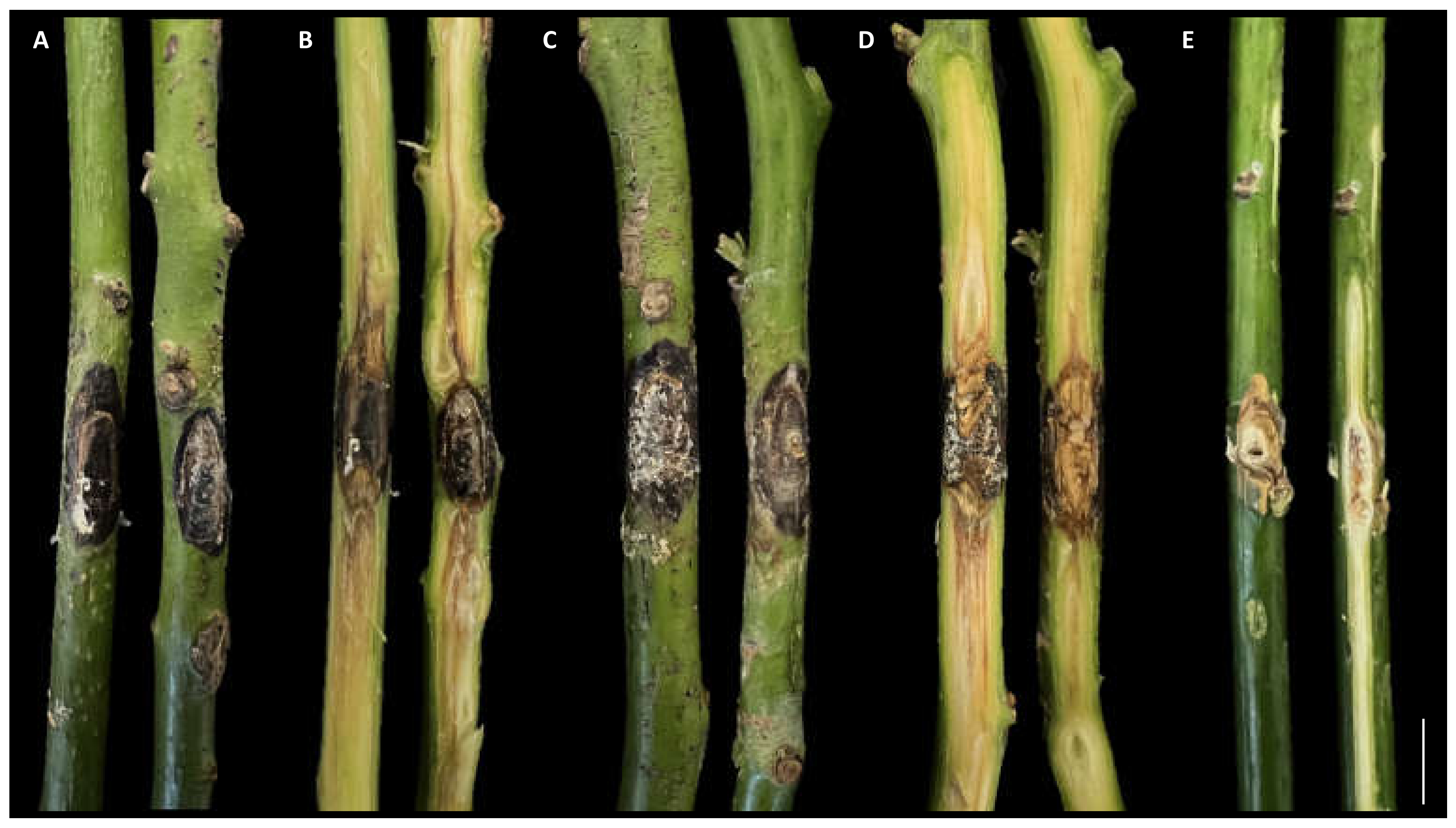
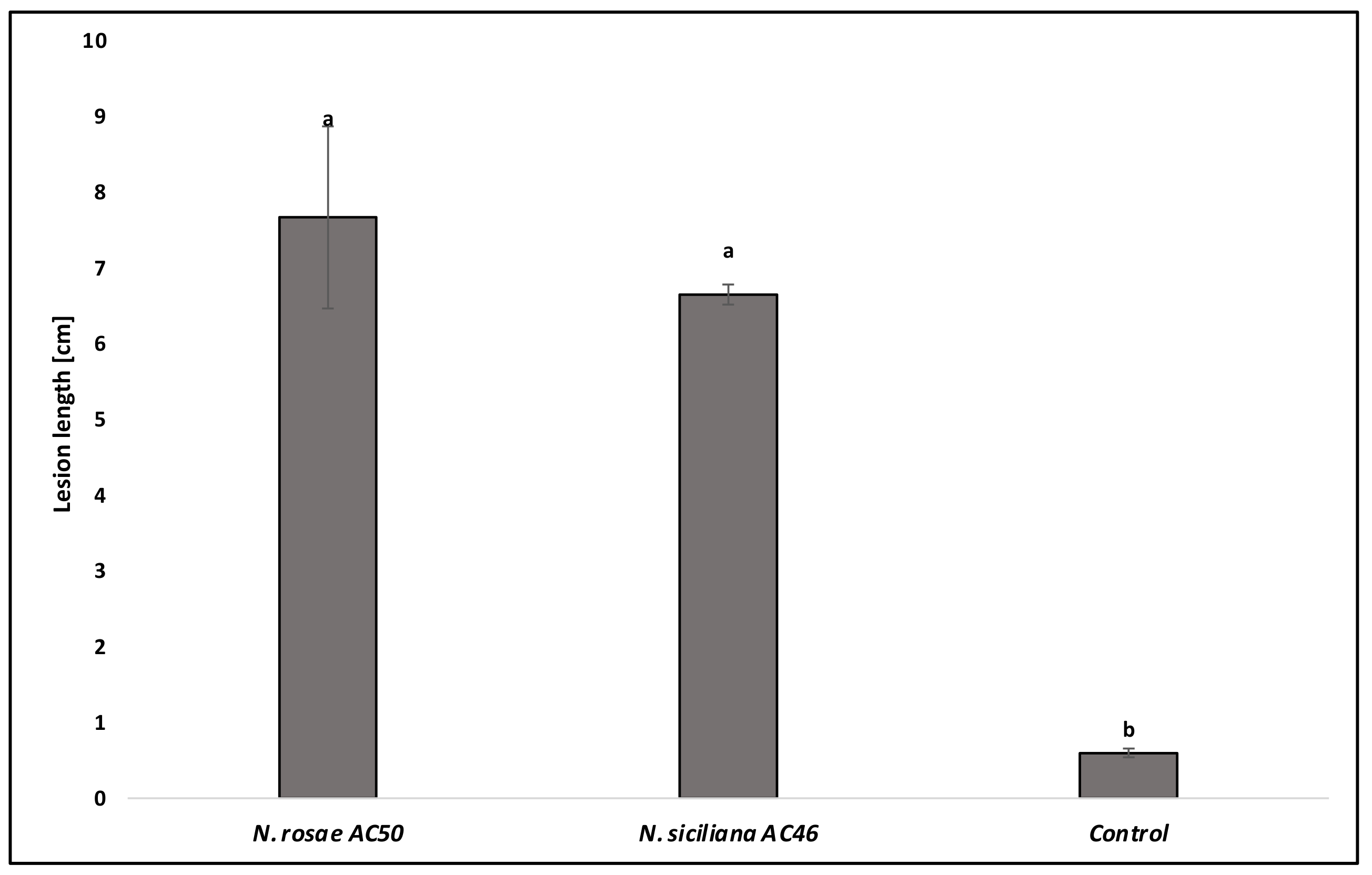
3.3. Pathogenicity Test
3.4. Morphological Description of Neopestalotiopsis rosae Isolates from Avocado
4. Taxonomy
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database; FAO: Rome, Italy, 2022; Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 11 April 2022).
- Migliore, G.; Farina, V.; Guccione, G.D.; Schifani, G. Quality determinants of avocado fruit consumption in Italy. Implications for small farms. Calitatea 2018, 19, 148–153. [Google Scholar]
- Zentmyer, G.A. Avocado. In Compendium of Tropical Fruit Diseases; Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G., Ohr, H.D., Eds.; APS Press: St. Paul, MA, USA, 1994; pp. 71–72. [Google Scholar]
- Zentmyer, G.A. Phytophthora Cinnamomi and the Disease it Causes; APS Press: St Paul, MA, USA, 1980. [Google Scholar]
- López-Herrera, C.J.; Melero-Vara, J.M. Diseases of avocado caused by soil fungi in the southern Mediterranean coast of Spain. In Proceedings of the Second World Avocado Congress California Avocado Society: The Shape of Things to Come, Orange, CA, USA, 21–26 April 1991; Lovatt, C.J., Holthe, P.A., Arpaia, M.L., Eds.; California Avocado Society: Riverside, CA, USA, 1992; Volume 1, pp. 119–121. [Google Scholar]
- Fiorenza, A.; Aiello, D.; Leonardi, G.R.; Continella, A.; Polizzi, G. First report of Rosellinia necatrix causing white root rot on avocado in Italy. Plant Dis. 2021, 105, 3294. [Google Scholar] [CrossRef]
- Parkinson, L.E.; Shivas, R.G.; Dann, E.K. Pathogenicity of nectriaceous fungi on avocado in Australia. Phytopathology 2017, 107, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Aiello, D.; Gusella, G.; Vitale, A.; Guarnaccia, V.; Polizzi, G. Cylindrocladiella peruviana and Pleiocarpon algeriense causing stem and crown rot on avocado (Persea americana). Eur. J. Plant Pathol. 2020, 158, 419–430. [Google Scholar] [CrossRef]
- Vitale, A.; Aiello, D.; Guarnaccia, V.; Perrone, G.; Stea, G.; Polizzi, G. First report of root rot caused by Ilyonectria (= Neonectria) macrodidyma on avocado (Persea americana) in Italy. J. Phytopathol. 2012, 160, 156–159. [Google Scholar] [CrossRef]
- McDonald, V.; Eskalen, A. Botryosphaeriaceae species associated with avocado branch cankers in California. Plant Dis. 2011, 95, 1465–1473. [Google Scholar] [CrossRef] [Green Version]
- Menge, J.A.; Ploetz, R.C. Diseases of avocado. In Diseases of Tropical Fruit Crops; Ploetz, R.C., Ed.; CABI Publishing: Wallingford, UK, 2003; pp. 35–71. [Google Scholar]
- Hartill, W.F.T.; Everett, K.R. Inoculum sources and infection pathways of pathogens causing stem-end rots of ‘Hass’ avocado (Persea americana). N. Zeal. J. Crop Hortic. Sci. 2002, 30, 249–260. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Vitale, A.; Cirvilleri, G.; Aiello, D.; Susca, A.; Epifani, F.; Perrone, G.; Polizzi, G. Characterisation and pathogenicity of fungal species associated with branch cankers and stem-end rot of avocado in Italy. Eur. J. Plant Pathol. 2016, 146, 963–976. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Sandoval-Denis, M.; Aiello, D.; Polizzi, G.; Crous, P.W. Neocosmospora perseae sp. nov., causing trunk cankers on avocado in Italy. Fungal Syst. Evol. 2018, 1, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Guarnaccia, V.; Aiello, D.; Papadantonakis, N.; Polizzi, G.; Gullino, M.L. First report of branch cankers on avocado (Persea americana) caused by Neocosmospora (syn. Fusarium) perseae in Crete (Greece). J. Plant Pathol. 2021, 104, 419–420. [Google Scholar] [CrossRef]
- Freeman, S.; Katan, T.; Shabi, E. Characterization of Colletotrichum species responsible for anthracnose diseases of various fruits. Plant Dis. 1998, 82, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Silva-Rojas, H.V.; Ávila-Quezada, G.D. Phylogenetic and morphological identification of Colletotrichum boninense: A novel causal agent of anthracnose in avocado. Plant Pathol. 2011, 60, 899–908. [Google Scholar] [CrossRef]
- Sharma, G.; Maymon, M.; Freeman, S. Epidemiology, pathology and identification of Colletotrichum including a novel species associated with avocado (Persea americana) anthracnose in Israel. Sci. Rep. 2017, 7, 15839. [Google Scholar] [CrossRef]
- Kimaru, S.K.; Monda, E.; Cheruiyot, R.C.; Mbaka, J.; Alakonya, A. Morphological and molecular identification of the causal agent of anthracnose disease of avocado in Kenya. Int. J. Microbiol. 2018, 2018, 4568520. [Google Scholar] [CrossRef] [Green Version]
- Valencia, A.L.; Torres, R.; Latorre, B.A. First report of Pestalotiopsis clavispora and Pestalotiopsis spp. causing postharvest stem end rot of avocado in Chile. Plant Dis. 2011, 95, 492. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Vitale, A.; Polizzi, G. Occurrence of Pestalotiopsis uvicola causing leaf spots and stem blight on bay laurel (Laurus nobilis) in Sicily. Plant Dis. 2005, 89, 1362. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innes, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–555. [Google Scholar] [CrossRef]
- Voglmayr, H.; Friebes, G.; Gardiennet, A.; Jaklitsch, W.M. Barrmaelia and Entosordaria in Barrmaeliaceae (fam. nov., Xylariales) and critical notes on Anthostomella-like genera based on multigene phylogenies. Mycol. Prog. 2018, 17, 155–177. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Voglmayr, H.; Gardiennet, A.; Jaklitsch, W.M. Asterodiscus and Stigmatodiscus, two new apothecial dothideomycete genera and the new order Stigmatodiscales. Fungal Divers. 2016, 80, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Werle, E.; Schneider, C.; Renner, M.; Völker, M.; Fiehn, W. Convenient single-step, one tube purification of PCR products for direct sequencing. Nucleic Acids Res. 1994, 22, 4354. [Google Scholar] [CrossRef]
- Voglmayr, H.; Jaklitsch, W.M. Prosthecium species with Stegonsporium anamorphs on Acer. Mycol. Res. 2008, 112, 885–905. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Jayawardena, R.S.; Chen, Y.; Wen, T.C.; Meepol, W.; Hyde, K.D. Morphological and phylogenetic characterization of novel pestalotioid species associated with mangroves in Thailand. Mycosphere 2019, 10, 531–578. [Google Scholar] [CrossRef]
- Kumar, V.; Cheewangkoon, R.; Gentekaki, E.; Maharachchikumbura, S.S.N.; Brahmange, R.S.; Hyde, K.D. Neopestalotiopsis alpapicalis sp. nov., a new endophyte from tropical mangrove trees in Krabi Province (Thailand). Phytotaxa 2019, 393, 251–262. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Cai, L.; Chukeatirote, E.; Wu, W.P.; Sun, X.; Hyde, K.D. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Divers. 2012, 56, 95–129. [Google Scholar] [CrossRef] [Green Version]
- Bezerra, J.D.P.; Machado, A.R.; Firmino, A.L.; Rosado, A.W.C.; Souza, C.A.F.; Souza-Motta, C.M.; Freire, K.T.L.S.; Paiva, L.M.; Magalhaes, O.M.C.; Pereira, O.L.; et al. Mycological diversity description I. Acta Bot. Bras. 2018, 32, 656–666. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Yang, Q.; Li, H. Morphology, phylogeny, and pathogenicity of pestalotioid species on Camellia oleifera in China. J. Fungi 2021, 7, 1080. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tibpromma, S.; Zhang, F.; Xu, J.; Chethana, K.W.T.; Karunarathna, S.C.; Mortimer, P.E. Neopestalotiopsis cavernicola sp. nov. from Gem Cave in Yunnan Province, China. Phytotaxa 2021, 512, 1–27. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Song, Y.; Geng, K.; Hyde, K.D.; Zhao, W.; Wei, J.G.; Kang, J.C.; Wang, Y. Two new species of Pestalotiopsis from Southern China. Phytotaxa 2013, 126, 22–30. [Google Scholar] [CrossRef]
- Ma, X.Y.; Maharachchikumbura, S.S.; Chen, B.W.; Hyde, K.D.; Mckenzie, E.H.; Chomnunti, P.; Kang, J.C. Endophytic pestalotiod taxa in Dendrobium orchids. Phytotaxa 2019, 419, 268–286. [Google Scholar] [CrossRef]
- Prasannath, K.; Shivas, R.G.; Galea, V.J.; Akinsanmi, O.A. Neopestalotiopsis species associated with flower diseases of Macadamia integrifolia in Australia. J. Fungi 2021, 7, 771. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvarado, P.; Edwards, J.; Moreno, G.; Sharma, R.; et al. Fungal Planet description sheets: 371–399. Persoonia 2015, 35, 264–327. [Google Scholar] [CrossRef]
- Diogo, E.; Gonçalves, C.I.; Silva, A.C.; Valente, C.; Bragança, H.; Phillips, A.J. Five new species of Neopestalotiopsis associated with diseased Eucalyptus spp. in Portugal. Mycol. Prog. 2021, 20, 1441–1456. [Google Scholar] [CrossRef]
- Ul Haq, I.; Ijaz, S.; Khan, N.A. Genealogical concordance of phylogenetic species recognition-based delimitation of Neopestalotiopsis species associated with leaf spots and fruit canker disease affected guava plants. Pak. J. Agric. Sci. 2021, 58, 1301–1313. [Google Scholar] [CrossRef]
- Freitas, E.F.S.; Da Silva, M.; Barros, M.V.P.; Kasuya, M.C.M. Neopestalotiopsis hadrolaeliae sp. nov., a new endophytic species from the roots of the endangered orchid Hadrolaelia jongheana in Brazil. Phytotaxa 2019, 416, 211–220. [Google Scholar] [CrossRef]
- Huanluek, N.; Jjayawardena, R.S.; Maharachchikumbura, S.S.N.; Harishchandra, D.L. Additions to pestalotioid fungi in Thailand: Neopestalotiopsis hydeana sp. nov. and Pestalotiopsis hydei sp. nov. Phytotaxa 2021, 479, 23–43. [Google Scholar] [CrossRef]
- Ayoubi, N.; Soleimani, M.J. Strawberry fruit rot caused by Neopestalotiopsis iranensis sp. nov., and N. mesopotamica. Curr. Microbiol. 2016, 72, 329–336. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Nisa, S.; Jeff-Ego, O.S.; Shivas, R.G.; Drenth, A. Dry flower disease of macadamia in Australia caused by Neopestalotiopsis macadamiae sp. nov. and Pestalotiopsis macadamiae sp. nov. Plant Dis. 2017, 101, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Maharachchikumbura, S.S.; Guo, L.D.; Chukeatirote, E.; Hyde, K.D. Improving the backbone tree for the genus Pestalotiopsis; addition of P. steyaertii and P. magna sp. nov. Mycol. Prog. 2014, 13, 617–624. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal Planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef] [PubMed]
- Silvério, M.L.; Cavalcanti, M.A.Q.; Silva, G.A.; Oliveira, R.J.V.; Bezerra, J.L. A new epifoliar species of Neopestalotiopsis from Brazil. Agrotropica 2016, 28, 151–158. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.A.; Swart, L.; Denman, S.; Taylor, J.E.; Bezuidenhout, C.M.; Palm, M.E.; Marincowitz, S.; Groenewald, J.Z. Fungal pathogens of Proteaceae. Persoonia 2011, 27, 20–45. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Zeng, X.Y.; Yuan, J.; Zhang, Q.; He, Y.K.; Wang, Y. Two new species of Neopestalotiopsis from southern China. Biodivers. Data J. 2021, 9, e70446. [Google Scholar] [CrossRef]
- Rebollar-Alviter, A.; Silva-Rojas, H.V.; Fuentes-Aragón, D.; Acosta-González, U.; Martínez-Ruiz, M.; Parra-Robles, B.E. An emerging strawberry fungal disease associated with root rot, crown rot and leaf spot caused by Neopestalotiopsis rosae in Mexico. Plant Dis. 2020, 104, 2054–2059. [Google Scholar] [CrossRef]
- Santos, J.; Hilário, S.; Pinto, G.; Alves, A. Diversity and pathogenicity of pestalotioid fungi associated with blueberry plants in Portugal, with description of three novel species of Neopestalotiopsis. Eur. J. Plant Pathol. 2022, 162, 539–555. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, X.; Tian, C. Identification and characterization of leaf-inhabiting fungi from Castanea plantations in China. J. Fungi 2021, 7, 64. [Google Scholar] [CrossRef]
- Nozawa, S.; Uchikawa, K.; Suga, Y.; Watanabe, K. Infection sources of Pestalotiopsis sensu lato related to loquat fruit rot in Nagasaki Prefecture, Japan. J. Gen. Plant. Pathol. 2020, 86, 173–179. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Liu, M.; Maharachchikumbura, S.S.N.; Zang, W.; Xing, Q.K.; Hyde, K.D.; Nilthong, S.; Li, X.; Yan, J. Neopestalotiopsis vitis sp. nov. causing grapevine leaf spot in China. Phytotaxa 2016, 258, 63–74. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP* 4.0b10: Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar] [CrossRef]
- Analytical Software. Statistix 10: User’s Manual; Analytical Software: Tallahassee, FL, USA, 2013. [Google Scholar]
- Moral, J.; Jurado-Bello, J.; Sánchez, M.I.; de Oliveira, R.; Trapero, A. Effect of temperature, wetness duration, and planting density on olive anthracnose caused by Colletotrichum spp. Phytopathology 2012, 102, 974–981. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gálvez, E.; Hilário, S.; Lopes, A.; Alves, A. Diversity and pathogenicity of Lasiodiplodia and Neopestalotiopsis species associated with stem blight and dieback of blueberry plants in Peru. Eur. J. Plant Pathol. 2020, 157, 89–102. [Google Scholar] [CrossRef]
- Xavier, K.V.; Yu, X.; Vallad, G.E. First report of Neopestalotiopsis rosae causing foliar and fruit spots on pomegranate in Florida. Plant Dis. 2020, 105, 504. [Google Scholar] [CrossRef]
- Baggio, J.S.; Forcelini, B.B.; Wang, N.Y.; Ruschel, R.G.; Mertely, J.C.; Peres, N.A. Outbreak of leaf spot and fruit rot in Florida strawberry caused by Neopestalotiopsis spp. Plant Dis. 2021, 105, 305–315. [Google Scholar] [CrossRef]
- Sun, Q.; Harishchandra, D.; Jia, J.; Zuo, Q.; Zhang, G.; Wang, Q.; Yan, J.; Zhang, W.; Li, X. Role of Neopestalotiopsis rosae in causing root rot of strawberry in Beijing, China. Crop Prot. 2021, 147, 105710. [Google Scholar] [CrossRef]
- Wu, H.Y.; Tsai, C.Y.; Wu, Y.M.; Ariyawansa, H.A.; Chung, C.L.; Chung, P.C. First report of Neopestalotiopsis rosae causing leaf blight and crown rot on strawberry in Taiwan. Plant Dis. 2021, 105, 487. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Lian, T.T.; Mai, X.M.; Jeewon, R.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Zeng, Y.J.; Tian, S.L.; Xie, N. New geographical records of Neopestalotiopsis and Pestalotiopsis species in Guangdong Province, China. Asian J. Mycol. 2020, 3, 510–530. [Google Scholar] [CrossRef]
- Shetty, K.G.; Rivadeneira, D.V.; Jayachandran, K.; Walker, D.M. Isolation and molecular characterization of the fungal endophytic microbiome from conventionally and organically grown avocado trees in South Florida. Mycol. Prog. 2016, 15, 977–986. [Google Scholar] [CrossRef]
- Espinoza, J.G.; Briceño, E.X.; Keith, L.M.; Latorre, B.A. Canker and twig dieback of blueberry caused by Pestalotiopsis spp. and a Truncatella sp. in Chile. Plant Dis. 2008, 92, 1407–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.H.; Dong, P.P.; Fang, S.Q.; Li, M.F.; Liu, W.B.; Miao, W.G. First report of avocado dieback disease caused by Pestalotiopsis longiseta in China. Plant Dis. 2018, 102, 2660. [Google Scholar] [CrossRef]
- Cardoso, J.E.; Maia, C.B.; Pessoa, M.N. Occurrence of Pestalotiopsis psidii and Lasiodiplodia theobromae causing stem rot of guava plants in the State of Ceará, Brazil. Fitopatol. Bras. 2002, 27, 320. [Google Scholar] [CrossRef]
- Gibson, I.A.S.; Howland, P. Graft failure in young Cupressus lusitanica. East Afr. Agric. For. J. 1969, 35, 52–54. [Google Scholar] [CrossRef]
- Rego, C.; Nascimento, T.; Cabral, A.; Oliveira, H. Fungi associated with young vine decline in Portugal: Results of nine years surveys. IOBC/WPRS Bull. 2006, 29, 123–126. [Google Scholar]
- Ismail, A.M.; Cirvilleri, G.; Polizzi, G. Characterisation and pathogenicity of Pestalotiopsis uvicola and Pestalotiopsis clavispora causing grey leaf spot of mango (Mangifera indica L.) in Italy. Eur. J. Plant Pathol. 2013, 135, 619–625. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain 1 | Host/Substrate | Origin | GenBank Accession Numbers 2 | Reference | ||
|---|---|---|---|---|---|---|---|
| ITS | TEF1 | TUB2 | |||||
| Neopestalotiopsis acrostichi | MFLUCC 17-1754 T | Acrostichum aureum | Thailand | MK764272 | MK764316 | MK764338 | [30] |
| N. alpapicalis | MFLUCC 17-2544 T | Rhizophora mucronata | Thailand | MK357772 | MK463547 | MK463545 | [31] |
| N. aotearoa | CBS 367.54 T | Canvas | New Zealand | KM199369 | KM199526 | KM199454 | [32] |
| N. asiatica | MFLUCC 12-0286 T | Unidentified tree | China | JX398983 | JX399049 | JX399018 | [32] |
| N. australis | CBS 114159 T | Telopea sp. | Australia | KM199348 | KM199537 | KM199348 | [21] |
| N. brachiata | MFLUCC 17-1555 T | Rhizophora apiculata | Thailand | MK764274 | MK764318 | MK764340 | [30] |
| N. brasiliensis | COAD 2166 T | Psidium guajava | Brazil | MG686469 | MG692402 | MG692400 | [33] |
| N. camelliae-oleiferae | CSUFTCC81 T | Camellia oleifera | China | OK493585 | OK507955 | OK562360 | [34] |
| N. cavernicola | KUMCC 20-0269 T | Cave rock surface | China | MW545802 | MW550735 | MW557596 | [35] |
| N. chiangmaiensis | MFLUCC 18-0113 T | Dead leaves | Thailand | N/A | MH388404 | MH412725 | [36] |
| N. chrysea | MFLUCC 12-0261 T | Pandanus sp. | China | JX398985 | JX399051 | JX399020 | [32] |
| N. clavispora | MFLUCC 12-0281 T | Magnolia sp. | China | JX398979 | JX399045 | JX399014 | [32] |
| N. cocoes | MFLUCC 15-0152 T | Cocos nucifera | Thailand | NR_156312 | KX789689 | N/A | [30] |
| N. coffeae-arabicae | HGUP 4019 T | Coffea arabica | China | KF412649 | KF412646 | KF412643 | [37] |
| N. cubana | CBS 600.96 T | Leaf litter | Cuba | KM199347 | KM199521 | KM199438 | [21] |
| N. dendrobii | MFLUCC 14-0106 T | Dendrobium cariniferum | Thailand | MK993571 | MK975829 | MK975835 | [38] |
| N. drenthii | BRIP 72264a T | Macadamia integrifolia | Australia | MZ303787 | MZ344172 | MZ312680 | [39] |
| N. egyptiaca | CBS 140162 T | Mangifera indica | Egypt | KP943747 | KP943748 | KP943746 | [40] |
| N. ellipsospora | MFLUCC 12-0283 T | Dead plant materials | China | JX398980 | JX399047 | JX399016 | [32] |
| N. eucalypticola | CBS 264.37 T | Eucalyptus globulus | N/A | KM199376 | KM199551 | KM199431 | [21] |
| N. eucalyptorum | CBS 147684 T | Eucalyptus globulus | Portugal | MW794108 | MW805397 | MW802841 | [41] |
| N. foedans | CGMCC 3.9123 T | Mangrove plant | China | JX398987 | JX399053 | JX399022 | [32] |
| N. formicidarum | CBS 362.72 T | Dead Formicidae (ant) | Ghana | KM199358 | KM199517 | KM199455 | [21] |
| N. guajavae | FMBCC 11.1 T | Psidium guajava | Pakistan | MF783085 | MH460868 | MH460871 | [42] |
| N. guajavicola | FMBCC 11.4 T | Psidium guajava | Pakistan | MH209245 | MH460870 | MH460873 | [42] |
| N. hadrolaeliae | COAD 2637 T | Hadrolaelia jongheana | Brazil | MK454709 | MK465122 | MK465120 | [43] |
| N. hispanica | CBS 147686 T | Eucalyptus globulus | Portugal | MW794107 | MW805399 | MW802840 | [41] |
| N. honoluluana | CBS 114495 T | Telopea sp. | USA | KM199364 | KM199548 | KM199457 | [21] |
| N. hydeana | MFLUCC 20-0132 T | Artocarpus heterophyllus | Thailand | MW266069 | MW251129 | MW251119 | [44] |
| N. iberica | CBS 147688 T | Eucalyptus globulus | Portugal | MW794111 | MW805402 | MW802844 | [41] |
| N. iranensis | CBS 137768 T | Fragaria × ananassa | Iran | KM074048 | KM074051 | KM074057 | [45] |
| N. javaensis | CBS 257.31 T | Cocos nucifera | Indonesia | KM199357 | KM199548 | KM199457 | [21] |
| N. longiappendiculata | CBS 147690 T | Eucalyptus globulus | Portugal | MW794112 | MW805404 | MW802845 | [41] |
| N. lusitanica | CBS 147692 T | Eucalyptus globulus | Portugal | MW794110 | MW805406 | MW802843 | [41] |
| N. macadamiae | BRIP 63737c T | Macadamia integrifolia | Australia | KX186604 | KX186627 | KX186654 | [46] |
| N. maddoxii | BRIP 72266a T | Macadamia integrifolia | Australia | MZ303782 | MZ344167 | MZ312675 | [39] |
| N. magna | MFLUCC 12-0652 T | Pteridium sp. | France | KF582795 | KF582791 | KF582793 | [47] |
| N. mesopotamica | CBS 336.86 T | Pinus brutia | Turkey | KM199362 | KM199555 | KM199441 | [21] |
| N. musae | MFLUCC 15-0776 T | Musa sp. | Thailand | NR_156311 | KX789685 | KX789686 | [30] |
| N. natalensis | CBS 138.41 T | Acacia mollissima | South Africa | NR_156288 | KM199552 | KM199466 | [47] |
| N. nebuloides | BRIP 66617 T | Sporobolus jacquemontii | Australia | MK966338 | MK977633 | MK977632 | [48] |
| N. olumideae | BRIP 72273a T | Macadamia integrifolia | Australia | MZ303790 | MZ344175 | MZ312683 | [39] |
| N. pandanicola | KUMCC 17-0175 T | Pandanus sp. | China | N/A | MH388389 | MH412720 | [36] |
| N. pernambucana | URM 7148-01 T | Vismia guianensis | Brazil | KJ792466 | KU306739 | N/A | [49] |
| N. perukae | FMBCC 11.3 T | Psidium guajava | Pakistan | MH209077 | MH523647 | MH460876 | [42] |
| N. petila | MFLUCC 17-1738 T | Rhizophora apiculata | Thailand | MK764276 | MK764320 | MK764342 | [30] |
| N. phangngaensis | MFLUCC 18-0119 T | Pandanus sp. | Thailand | MH388354 | MH388390 | MH412721 | [36] |
| N. piceana | CBS 394.48 T | Picea sp. | UK | KM199368 | KM199527 | KM199453 | [21] |
| N. protearum | CBS 114178 T | Leucospermum cuneiforme | Zimbabwe | JN712498 | KM199542 | KM199463 | [50] |
| N. psidii | FMBCC 11.2 T | Psidium guajava | Pakistan | MF783082 | MH460874 | MH477870 | [42] |
| N. rhapidis | GUCC 21501 T | Rhododendron simsii | China | MW931620 | MW980442 | MW980441 | [51] |
| N. rhizophorae | MFLUCC 17-1551 T | Rhizophora mucronata | Thailand | MK764277 | MK764321 | MK764343 | [30] |
| N. rhododendri | GUCC 21504 T | Rhododendron simsii | China | MW979577 | MW980444 | MW980443 | [51] |
| N. rosae | CBS 101057 T | Rosa sp. | New Zealand | KM199359 | KM199523 | KM199429 | [21] |
| N. rosae | CBS 124745 | Paeonia suffruticosa | USA | KM199360 | KM199524 | KM199430 | [21] |
| N. rosae | CRM-FRC | Fragaria × ananassa | Mexico | MN385718 | MN268532 | MN268529 | [52] |
| N. rosae | AC50 | Persea americana | Italy | ON117810 | ON107276 | ON209165 | this study |
| N. rosicola | CFCC 51992 T | Rosa chinensis | China | KY885239 | KY885243 | KY885245 | [30] |
| N. samarangensis | MFLUCC 12-0233 T | Syzygium samarangense | Thailand | JQ968609 | JQ968611 | JQ968610 | [30] |
| N. saprophytica | MFLUCC 12-0282 T | Magnolia sp. | China | JX398982 | JX399048 | JX399017 | [21] |
| N. scalabiensis | CAA1029 T | Vaccinium corymbosum | Portugal | MW969748 | MW959100 | MW934611 | [53] |
| N. sichuanensis | CFCC 54338 T | Castanea mollissima | China | MW166231 | MW199750 | MW218524 | [54] |
| N. siciliana | AC46 | Persea americana | Italy | ON117813 | ON107273 | ON209162 | this study |
| N. siciliana | AC48 | Persea americana | Italy | ON117812 | ON107274 | ON209163 | this study |
| N. siciliana | AC49 | Persea americana | Italy | ON117811 | ON107275 | ON209164 | this study |
| N. sonneratiae | MFLUCC 17-1745 T | Sonneronata alba | Thailand | MK764280 | MK764324 | MK764346 | [30] |
| N. sp. | TAP18N001 | Eriobotrya japonica | Japan | LC427126 | LC427128 | LC427127 | [55] |
| N. sp. | TAP18N006 | Eriobotrya japonica | Japan | LC427141 | LC427143 | LC427142 | [55] |
| N. sp. | TAP18N016 | Eriobotrya japonica | Japan | LC427168 | LC427170 | LC427169 | [55] |
| N. sp. | TAP18N021 | Eriobotrya japonica | Japan | LC427183 | LC427185 | LC427184 | [55] |
| N. steyaertii | IMI 192475 T | Eucalytpus viminalis | Australia | KF582796 | KF582792 | KF582794 | [47] |
| N. surinamensis | CBS 450.74 T | Soil under Elaeis guineensis | Suriname | KM199351 | KM199518 | KM199465 | [21] |
| N. thailandica | MFLUCC 17-1730 T | Rhizophora mucronata | Thailand | MK764281 | MK764325 | MK764347 | [30] |
| N. umbrinospora | MFLUCC 12-0285 T | unidentified plant | China | JX398984 | JX399050 | JX399019 | [32] |
| N. vaccinii | CAA1059 T | Vaccinium corymbosum | Portugal | MW969747 | MW959099 | MW934610 | [53] |
| N. vacciniicola | CAA1055 T | Vaccinium corymbosum | Portugal | MW969751 | MW959103 | MW934614 | [53] |
| N. vheenae | BRIP 72293a T | Macadamia integrifolia | Australia | MZ303792 | MZ344177 | MZ312685 | [39] |
| N. vitis | MFLUCC 15-1265 T | Vitis vinifera | China | KU140694 | KU140676 | KU140685 | [56] |
| N. zakeelii | BRIP 72282a T | Macadamia integrifolia | Australia | MZ303789 | MZ344174 | MZ312682 | [39] |
| N. zimbabwana | CBS 111495 T | Leucospermum cuneiforme | Zimbabwe | MH554855 | KM199545 | KM199456 | [21] |
| Pestalotiopsis colombiensis | CBS 118553 T | Eucalyptus grandis × urophylla | Colombia | KM199307 | KM199488 | KM199421 | [21] |
| Pestalotiopsis diversiseta | MFLUCC 12-0287 T | Dead plant material | China | NR_120187 | JX399073 | JX399040 | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiorenza, A.; Gusella, G.; Aiello, D.; Polizzi, G.; Voglmayr, H. Neopestalotiopsis siciliana sp. nov. and N. rosae Causing Stem Lesion and Dieback on Avocado Plants in Italy. J. Fungi 2022, 8, 562. https://doi.org/10.3390/jof8060562
Fiorenza A, Gusella G, Aiello D, Polizzi G, Voglmayr H. Neopestalotiopsis siciliana sp. nov. and N. rosae Causing Stem Lesion and Dieback on Avocado Plants in Italy. Journal of Fungi. 2022; 8(6):562. https://doi.org/10.3390/jof8060562
Chicago/Turabian StyleFiorenza, Alberto, Giorgio Gusella, Dalia Aiello, Giancarlo Polizzi, and Hermann Voglmayr. 2022. "Neopestalotiopsis siciliana sp. nov. and N. rosae Causing Stem Lesion and Dieback on Avocado Plants in Italy" Journal of Fungi 8, no. 6: 562. https://doi.org/10.3390/jof8060562
APA StyleFiorenza, A., Gusella, G., Aiello, D., Polizzi, G., & Voglmayr, H. (2022). Neopestalotiopsis siciliana sp. nov. and N. rosae Causing Stem Lesion and Dieback on Avocado Plants in Italy. Journal of Fungi, 8(6), 562. https://doi.org/10.3390/jof8060562