
SterylAcetyl Hydrolase 1 (BbSay1) Links Lipid Homeostasis to Conidiogenesis and Virulence in the Entomopathogenic Fungus Beauveria bassiana




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Cultivation
2.2. Bioinformatic and Transcriptional Analyses of BbSay1 in B. bassiana
2.3. Gene Disruption and Complementation
2.4. Biochemical and Phenotypic Assays
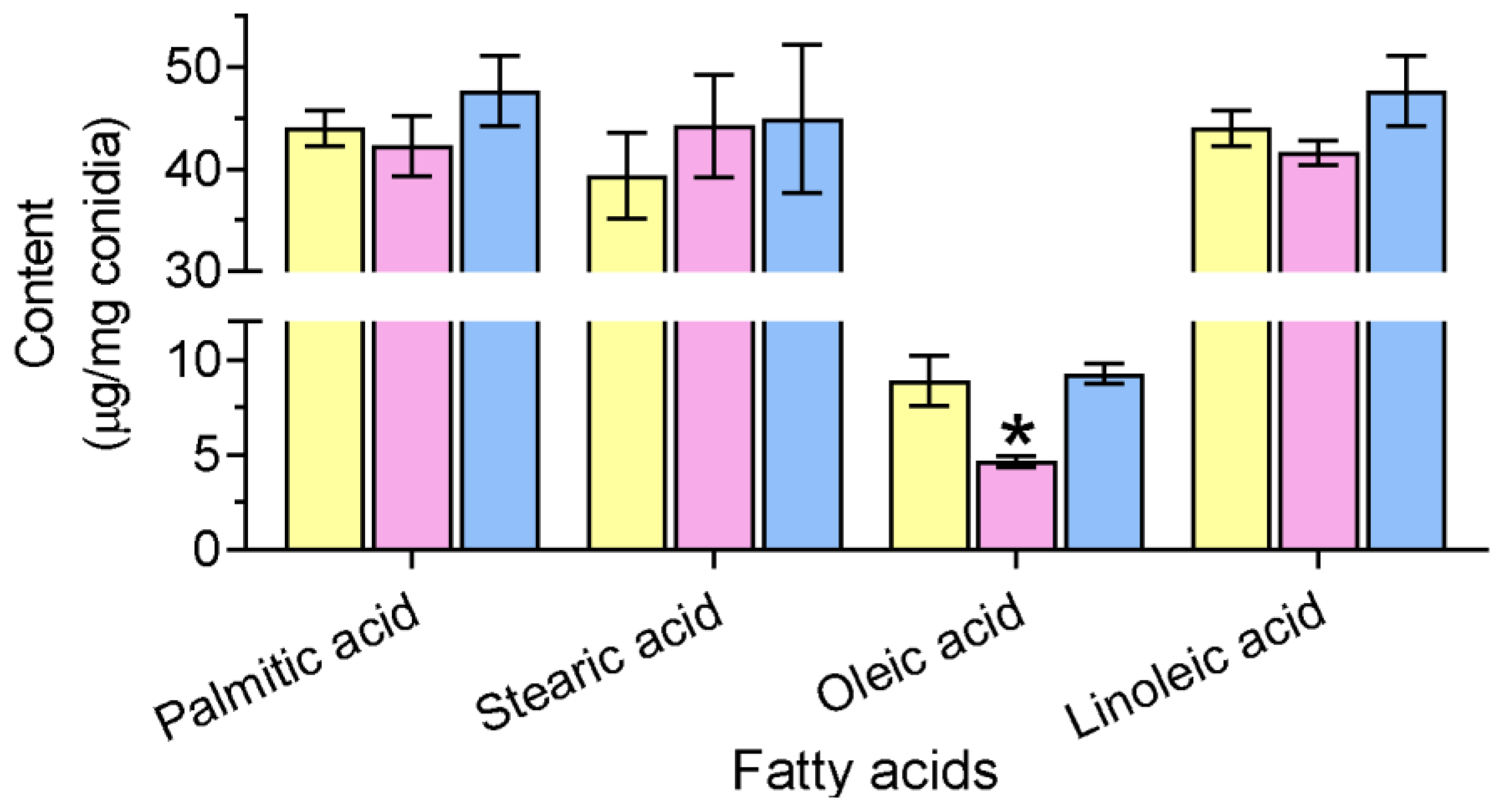
2.4.1. Quantification of Conidial FAs
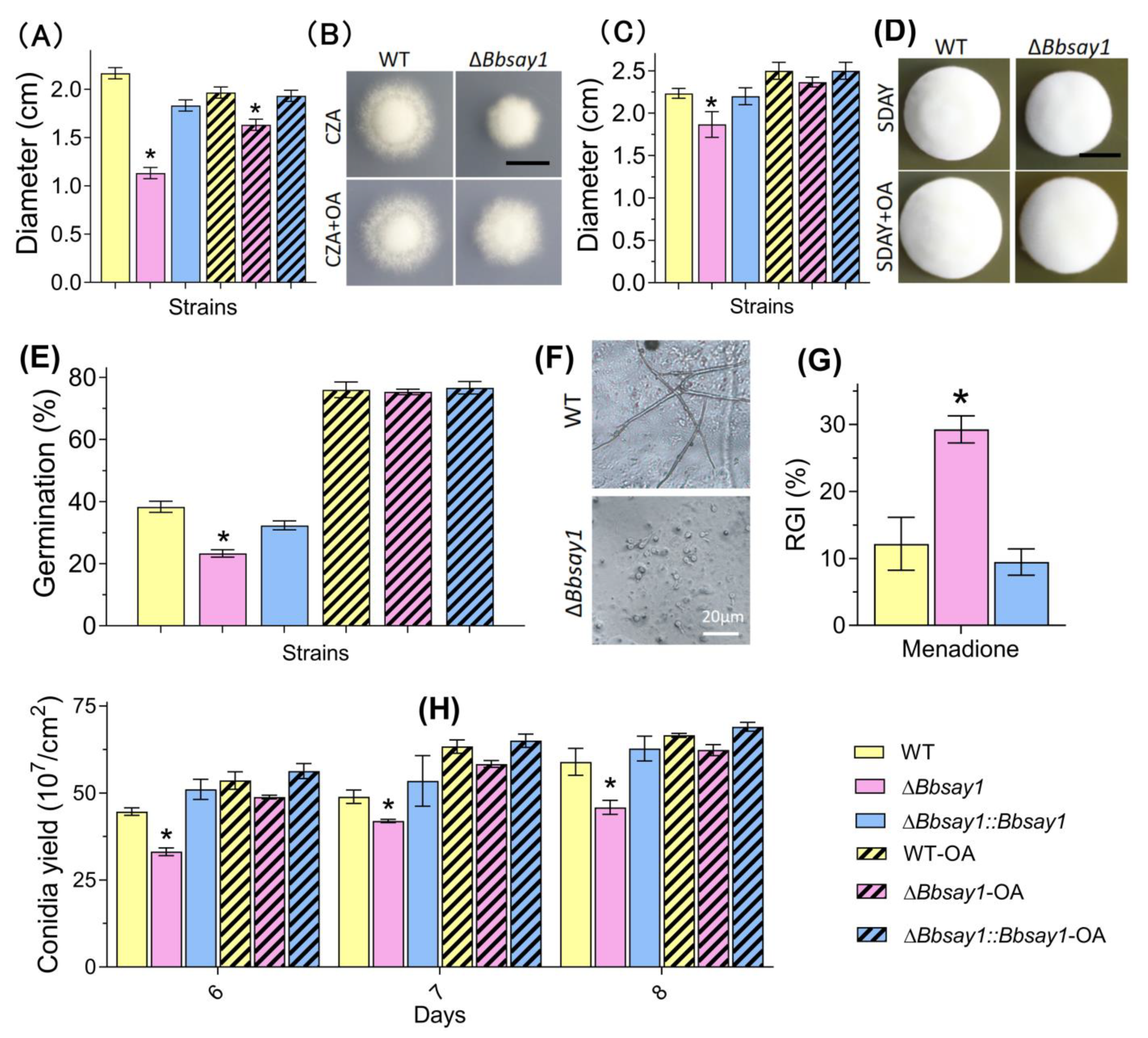
2.4.2. Vegetative Growth
2.4.3. Conidial Production
2.4.4. Conidia Germination
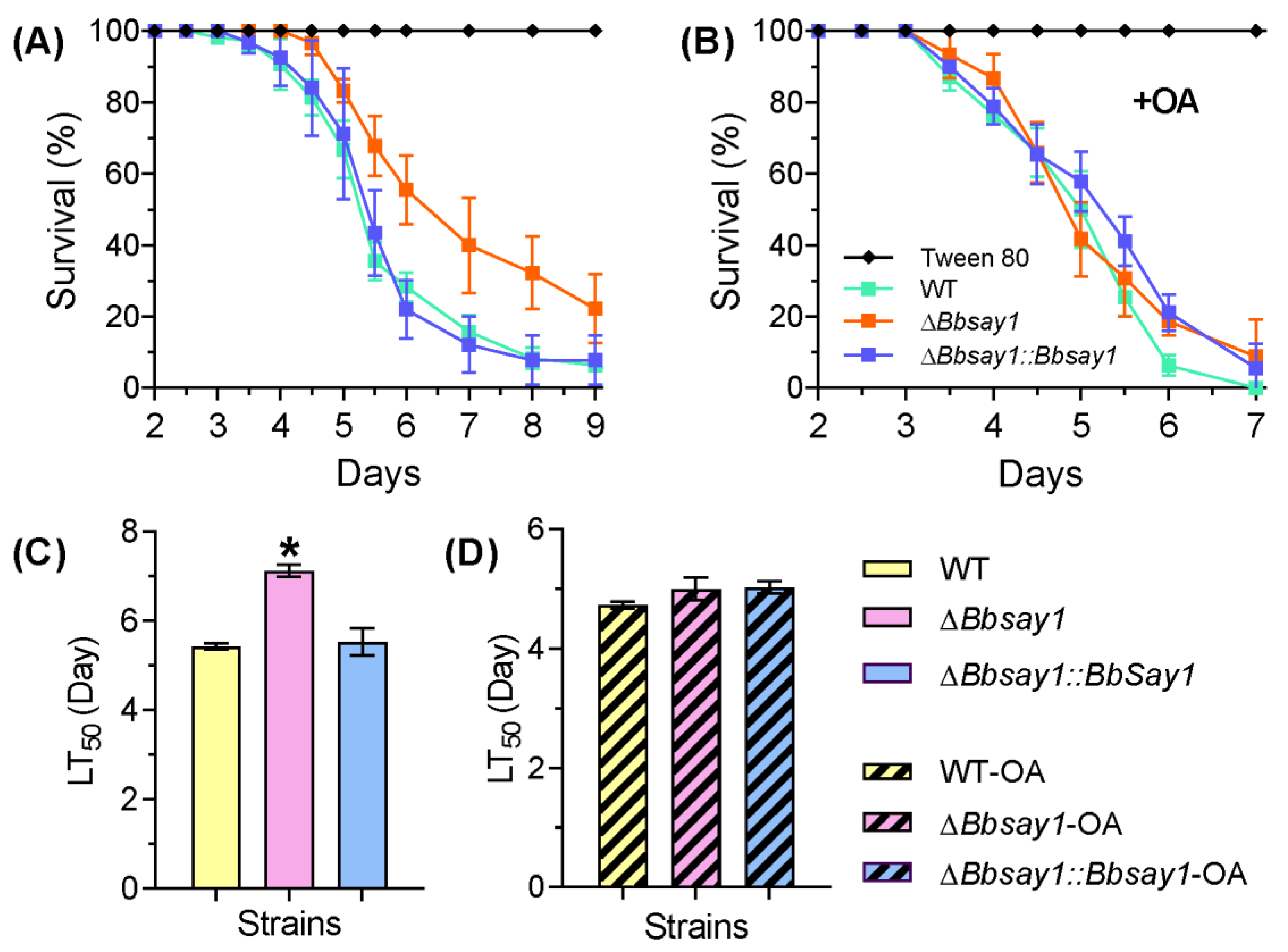
2.4.5. Insect Bioassay
2.5. Lipidomic Analyses
2.6. Assay for Membrane Integrity
2.7. Data Analyses
3. Results
3.1. Bioinformatic Analysis and Molecular Manipulation of BbSay1
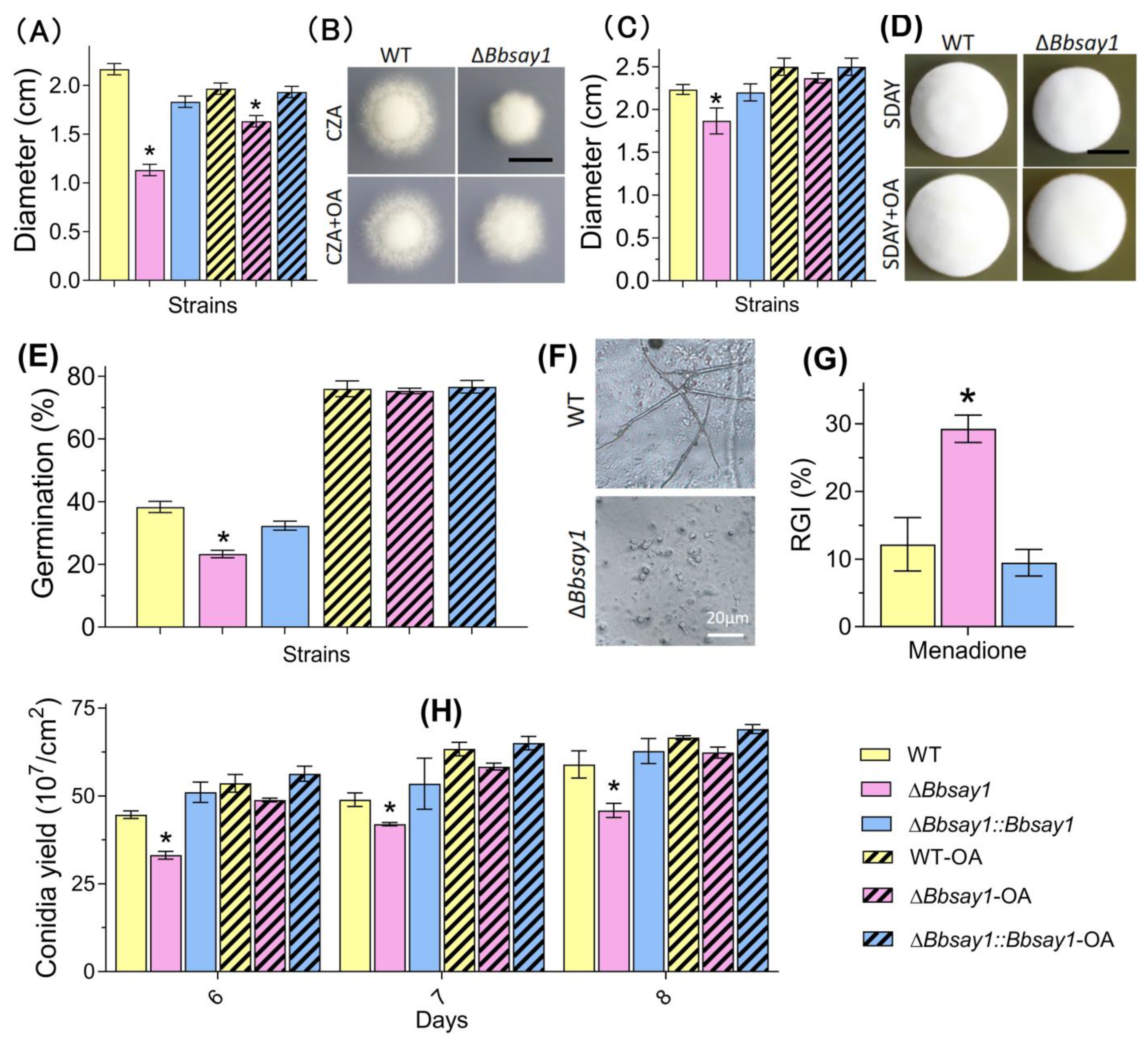
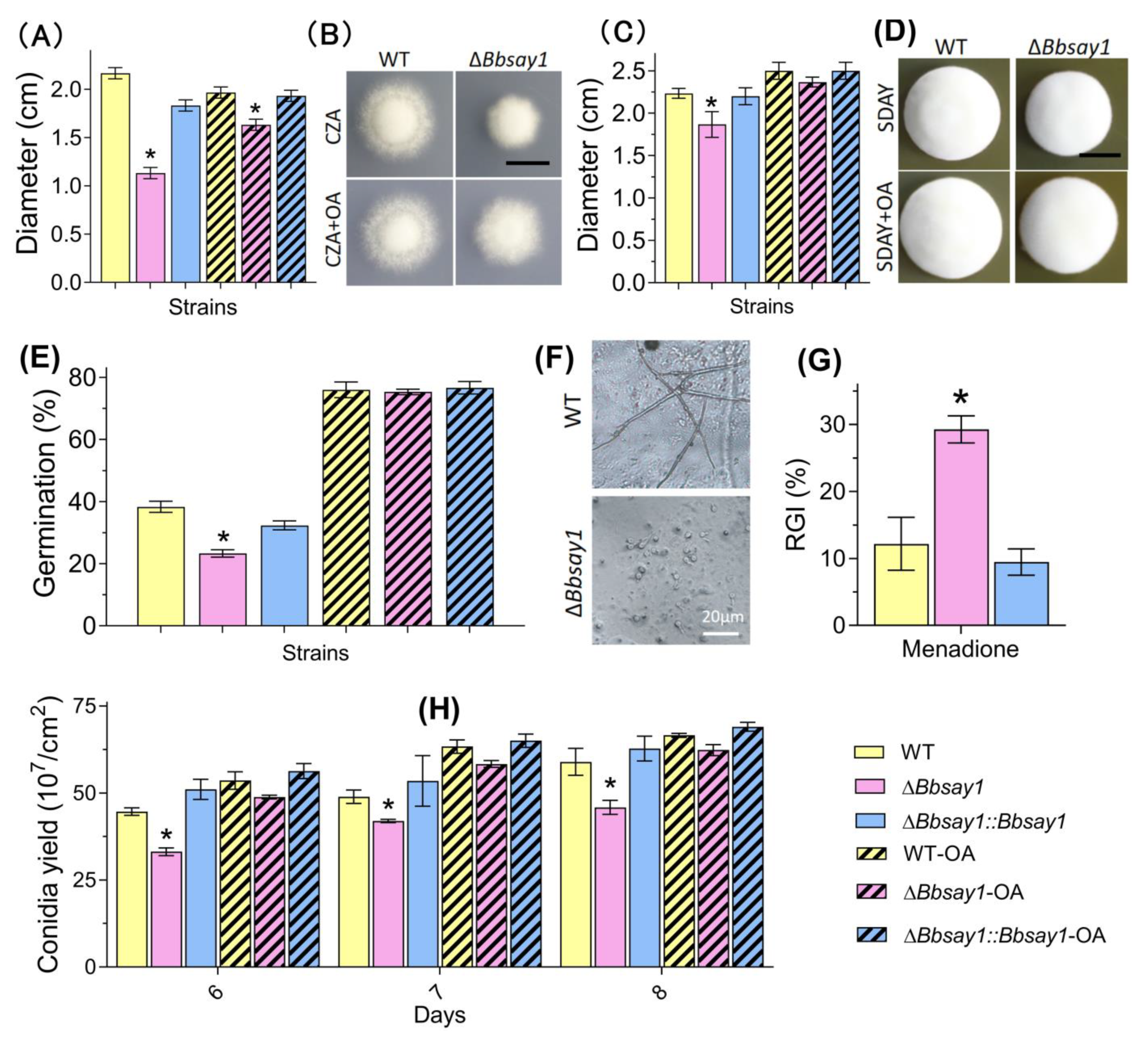
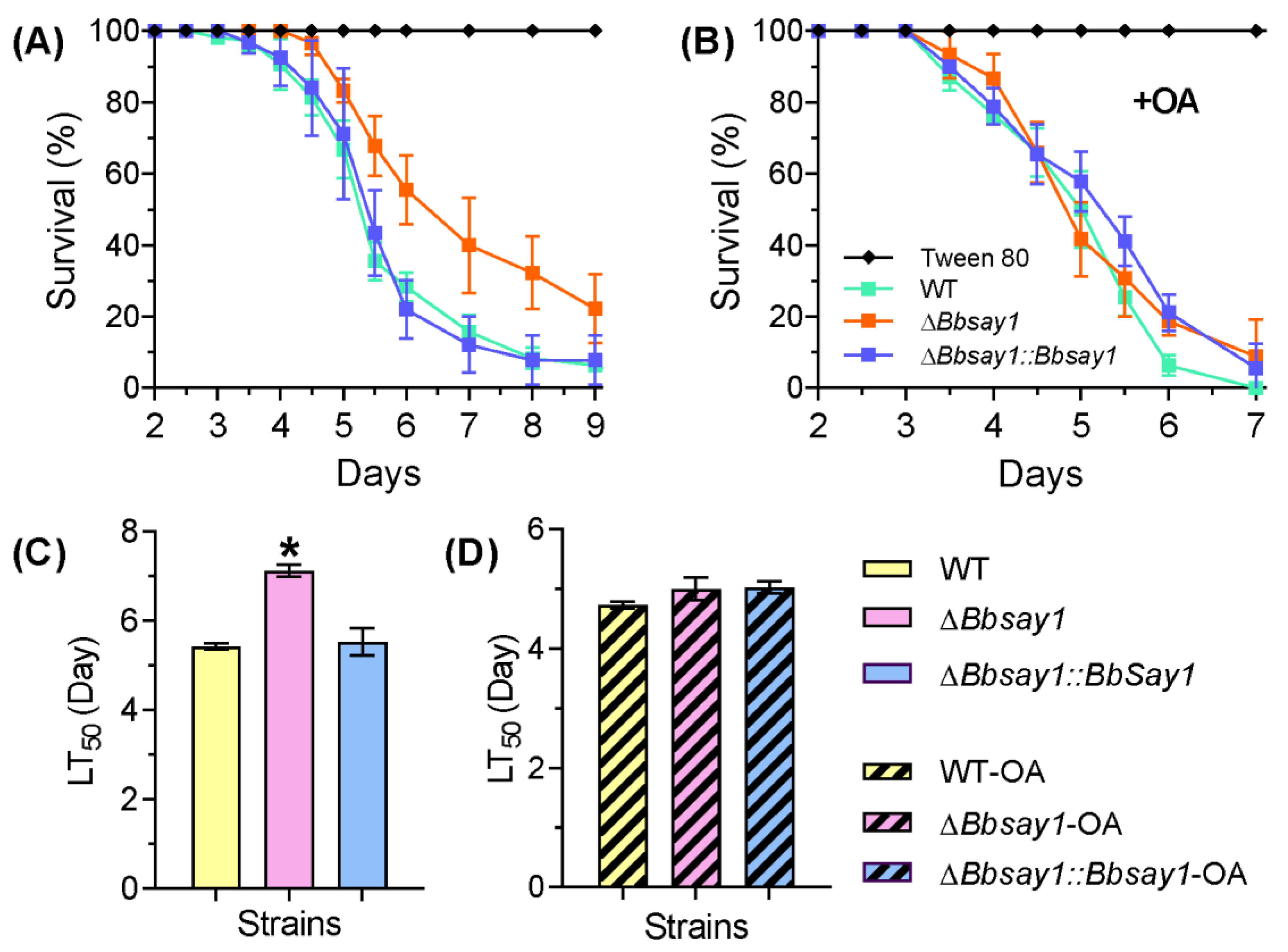
3.2. Effects of Gene Loss on Fungal Phenotypes
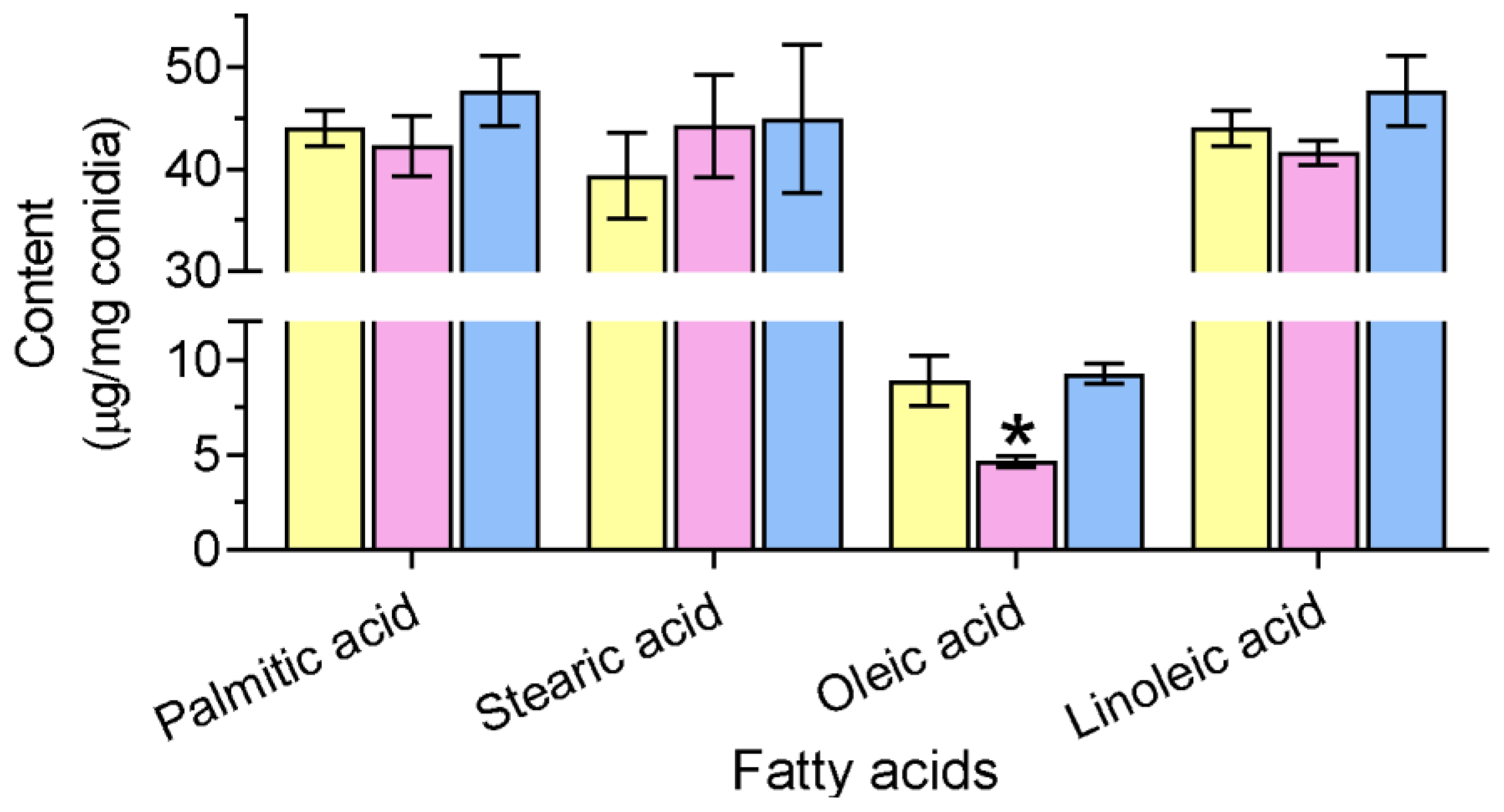
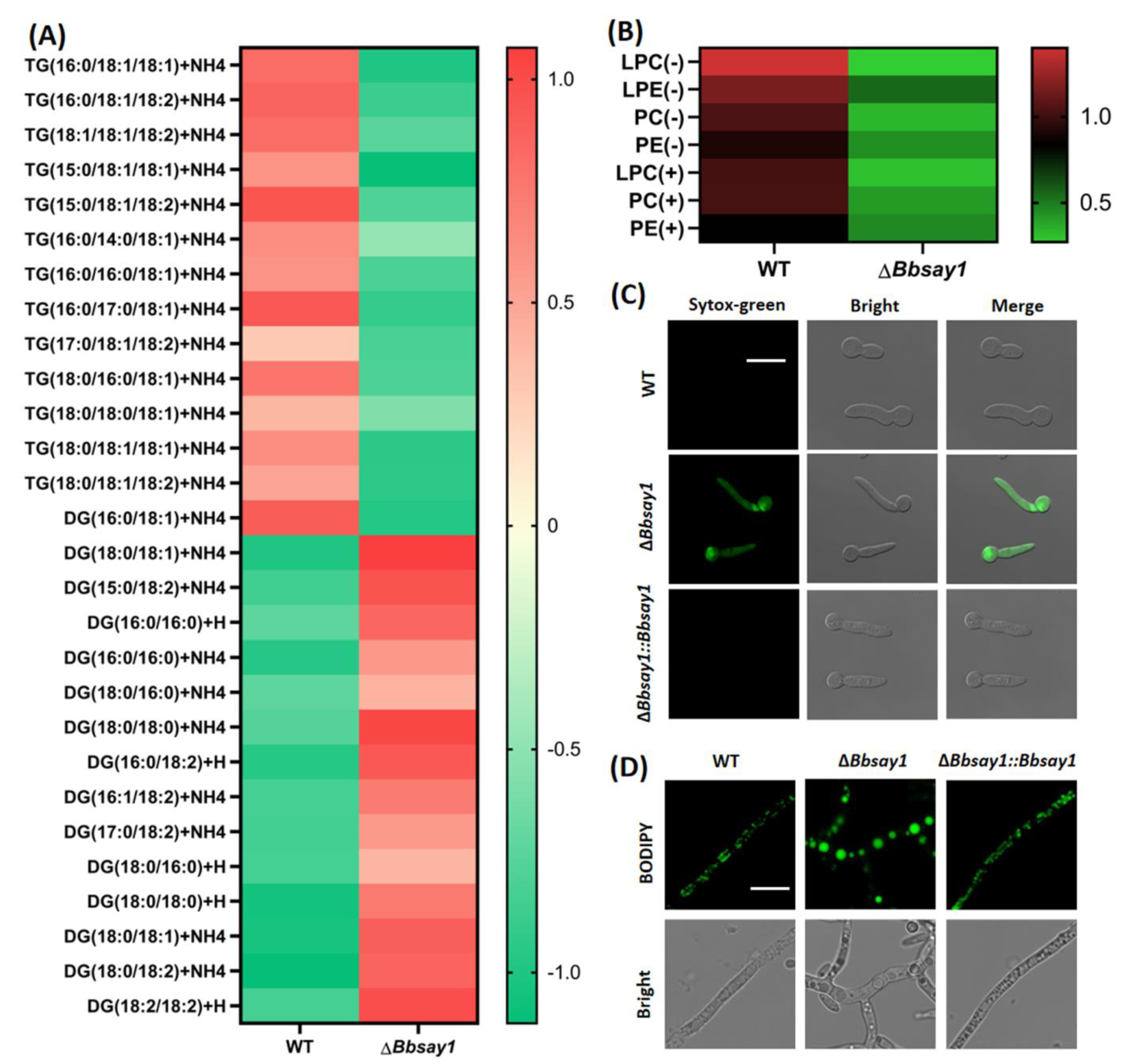
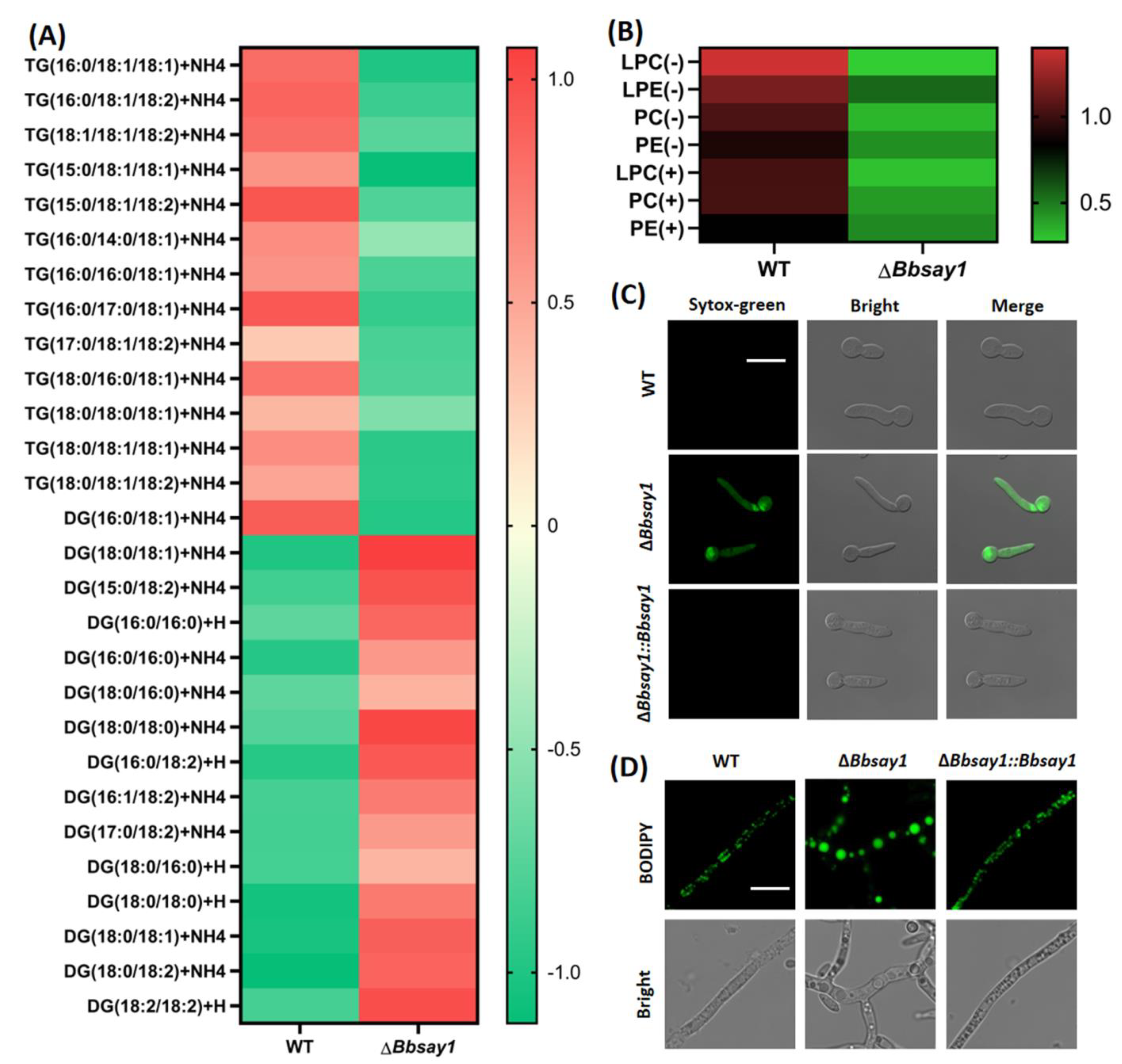
3.3. Gene Loss Affects Cellular Lipidomics and Membrane Integrity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De la Cruz, Q.R.; Roussos, S.; Hernandez, D.; Rodriguez, R.; Castillo, F.; Aguilar, C.N. Challenges and opportunities of the bio-pesticides production by solid-state fermentation: Filamentous Fungi as a Model. Crit. Rev. Biotechnol. 2015, 35, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A.; Luo, Z.; Keyhani, N.O. Improving mycoinsecticides for insect biological control. Appl. Microbiol. Biotechnol. 2015, 99, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic Fungi Versus the Insect Cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Wanchoo, A.; Lewis, M.W.; Keyhani, N.O. Lectin mapping reveals stage-specific display of surface carbohydrates in in vitro and haemolymph-derived cells of the entomopathogenic fungus Beauveria Bassiana. Microbiology 2009, 155, 3121–3133. [Google Scholar] [CrossRef] [Green Version]
- Shah, P.A.; Pell, J.K. Entomopathogenic fungi as biological control agents. Appl. Microbiol. Biotechnol. 2003, 61, 413–423. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Lopez-Jimenez, J.A.; Perez-Berna, A.J.; Huang, I.-C.; Jansson, H.; Salinas, B.J.; Villalain, J.; Read, N.D.; Lopez-Llorca, L.V. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Mol. Microbiol. 2010, 75, 1021–1032. [Google Scholar] [CrossRef]
- Peng, Y.-J.; Wang, J.-J.; Lin, H.-Y.; Ding, J.-L.; Feng, M.-G.; Ying, S.-H. HapX, an indispensable bZIP transcription factor for iron acquisition, regulates infection initiation by orchestrating conidial oleic acid homeostasis and cytomembrane functionality in mycopathogen Beauveria bassiana. Msystems 2020, 5, e00695. [Google Scholar] [CrossRef]
- Calvo, A.M.; Gardner, H.W.; Keller, N.P. Genetic connection between fatty acid metabolism and sporulation in Aspergillus nidulans. J. Biol. Chem. 2001, 276, 25766–25774. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, G.M.; Trono, D. Recombinant lipases and phospholipases and their use as biocatalysts for industrial applications. Int. J. Mol. Sci. 2015, 16, 20774–20840. [Google Scholar] [CrossRef] [Green Version]
- Ballweg, S.; Ernst, R. Control of membrane fluidity: The OLE Pathway in Focus. Biol. Chem. 2017, 398, 215–228. [Google Scholar] [CrossRef]
- Gsaller, F.; Hortschansky, P.; Beattie, S.R.; Klammer, V.; Tuppatsch, K.; Lechner, B.E.; Rietzschel, N.; Werner, E.R.; Vogan, A.A.; Chung, D.; et al. The Janus transcription factor HapX controls fungal adaptation to both iron starvation and iron excess. EMBO J. 2014, 33, 2261–2276. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.; Koffel, R.; Schneiter, R. An acetylation/deacetylation cycle controls the export of sterols and steroids from S. cerevisiae. EMBO J. 2007, 26, 5109–5119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.; Ying, S.-H.; Zheng, P.; Wang, Z.-L.; Zhang, S.; Xie, X.-Q.; Shang, Y.; St Leger, R.J.; Zhao, G.-P.; Wang, C. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lechner, M.; Findeis, S.; Steiner, L.; Marz, M.; Stadler, P.F.; Prohaska, S.J. Proteinortho: Detection of (Co-)orthologs in Large-Scale Analysis. BMC Bioinform. 2011, 12, 124. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-X.; He, P.-H.; Feng, M.-G.; Ying, S.-H. BbSNF1 contributes to cell differentiation, extracellular acidification, and virulence in Beauveria bassiana, a filamentous entomopathogenic fungus. Appl. Microbiol. Biotechnol. 2014, 98, 8657–8673. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, J.-J.; Peng, Y.-J.; Feng, M.-G.; Ying, S.-H. Functional analysis of the mitochondrial gene mitofilin in the filamentous entomopathogenic fungus Beauveria bassiana. Fungal Genet. Biol. 2019, 132, 103250. [Google Scholar] [CrossRef]
- He, P.-H.; Dong, W.-X.; Chu, X.-L.; Feng, M.-G.; Ying, S.-H. The cellular proteome is affected by a gelsolin (BbGEL1) during morphological transitions in aerobic surface versus liquid growth in the entomopathogenic fungus Beauveria bassiana. Environ. Microbiol. 2016, 18, 4153–4169. [Google Scholar] [CrossRef]
- Xie, X.-Q.; Li, F.; Ying, S.-H.; Feng, M.-G. Additive contributions of two manganese-cored superoxide dismutases (MnSODs) to antioxidation, UV tolerance and virulence of Beauveria bassiana. PLoS ONE 2012, 7, e30298. [Google Scholar]
- Ding, J.-L.; Hou, J.; Li, X.-H.; Feng, M.-G.; Ying, S.-H. Transcription activator Swi6 interacts with Mbp1 in MluI cell cycle box-binding complex and regulates hyphal differentiation and virulence in Beauveria bassiana. J. Fungi 2021, 7, 411. [Google Scholar] [CrossRef]
- Ding, J.-L.; Hou, J.; Feng, M.-G.; Ying, S.-H. Transcriptomic analyses reveal comprehensive responses of insect hemocytes to mycopathogen Beauveria bassiana, and fungal virulence-related cell wall protein assists pathogen to evade host cellular defense. Virulence 2020, 11, 1352–1365. [Google Scholar] [CrossRef]
- Keyhani, N.O. Lipid biology in fungal stress and virulence: Entomopathogenic Fungi. Fungal Biol. 2018, 122, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Currie, E.; Guo, X.; Christiano, R.; Chitraju, C.; Kory, N.; Harrison, K.; Haas, J.; Walther, T.C.; Farese, R.J. High confidence proteomic analysis of yeast LDs identifies additional droplet proteins and reveals connections to dolichol synthesis and sterol acetylation. J. Lipid Res. 2014, 55, 1465–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Siloto, R.M.; Lehner, R.; Stone, S.J.; Weselake, R.J. Acyl-CoA:diacylglycerol acyltransferase: Molecular Biology, Biochemistry and Biotechnology. Prog. Lipid Res. 2012, 51, 350–377. [Google Scholar] [CrossRef]
- Walther, T.C.; Farese, R.J. Lipid droplets and cellular lipid metabolism. Annu. Rev. Biochem. 2012, 81, 687–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, S.-H.; Liu, J.; Chu, X.-L.; Xie, X.-Q.; Feng, M.-G. The autophagy-related genes BbATG1 and BbATG8 have different functions in differentiation, stress resistance and virulence of mycopathogen Beauveria bassiana. Sci. Rep. 2016, 6, e26376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.-Y.; Pang, M.-Y.; Feng, M.-G.; Ying, S.-H. A peroxisomal sterol carrier protein 2 (Scp2) contributes to lipid trafficking in differentiation and virulence of the insect pathogenic fungus Beauveria bassiana. Fungal Genet. Biol. 2022, 158, e103651. [Google Scholar] [CrossRef]
- Li, X.-H.; Peng, Y.-J.; Ding, J.-L.; Feng, M.-G.; Ying, S.-H. A homologue of yeast acyl-CoA synthetase Faa1 contributes to cytomembrane functionality involved in development and virulence in the insect pathogenic fungus Beauveria bassiana. Microb. Pathog. 2022, 164, e105419. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.-J.; Ding, J.-L.; Lin, H.-Y.; Feng, M.-G.; Ying, S.-H. A virulence-related lectin traffics into eisosome and contributes to functionality of cytomembrane and cell-wall in the insect-pathogenic fungus Beauveria bassiana. Fungal Biol. 2021, 125, 914–922. [Google Scholar] [CrossRef] [PubMed]
- He, P.-H.; Wang, X.-X.; Chu, X.-L.; Feng, M.-G.; Ying, S.-H. RNA sequencing analysis identifies the metabolic and developmental genes regulated by BbSNF1 during conidiation of the entomopathogenic fungus Beauveria bassiana. Curr. Genet. 2015, 61, 143–152. [Google Scholar] [CrossRef]
- Wilson, R.A.; Chang, P.K.; Dobrzyn, A.; Ntambi, J.M.; Zarnowski, R.; Keller, N.P. Two Δ9-stearic acid desaturases are required for Aspergillus nidulans growth and development. Fungal Genet. Biol. 2004, 41, 501–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.-S.; Menardo, F.; Xue, M.-F.; Zhang, X.-J.; Gong, S.-J.; Yang, L.-J.; Shi, W.-Q.; Yu, D.-Z. Transcriptome analyses shed new insights into primary metabolism and regulation of Blumeria graminis f. sp. tritici during conidiation. Front. Plant Sci. 2017, 8, e1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhu, J.; Hu, J.; Meng, X.; Zhang, Q.; Zhu, K.; Chen, X.; Chen, X.; Li, G.; Wang, Z.; et al. Functional characterization of electron-transferring flavoprotein and its dehydrogenase required for fungal development and plant infection by the rice blast fungus. Sci. Rep. 2016, 6, e24911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Y.-J.; Zhang, H.; Feng, M.-G.; Ying, S.-H. SterylAcetyl Hydrolase 1 (BbSay1) Links Lipid Homeostasis to Conidiogenesis and Virulence in the Entomopathogenic Fungus Beauveria bassiana. J. Fungi 2022, 8, 292. https://doi.org/10.3390/jof8030292
Peng Y-J, Zhang H, Feng M-G, Ying S-H. SterylAcetyl Hydrolase 1 (BbSay1) Links Lipid Homeostasis to Conidiogenesis and Virulence in the Entomopathogenic Fungus Beauveria bassiana. Journal of Fungi. 2022; 8(3):292. https://doi.org/10.3390/jof8030292
Chicago/Turabian StylePeng, Yue-Jin, Hao Zhang, Ming-Guang Feng, and Sheng-Hua Ying. 2022. "SterylAcetyl Hydrolase 1 (BbSay1) Links Lipid Homeostasis to Conidiogenesis and Virulence in the Entomopathogenic Fungus Beauveria bassiana" Journal of Fungi 8, no. 3: 292. https://doi.org/10.3390/jof8030292
APA StylePeng, Y.-J., Zhang, H., Feng, M.-G., & Ying, S.-H. (2022). SterylAcetyl Hydrolase 1 (BbSay1) Links Lipid Homeostasis to Conidiogenesis and Virulence in the Entomopathogenic Fungus Beauveria bassiana. Journal of Fungi, 8(3), 292. https://doi.org/10.3390/jof8030292