
The Genus Xanthagaricus: An Updated Global Species Distribution and Phylogeny with the Description of Two New Species from Oman

 ,
, 
Abstract
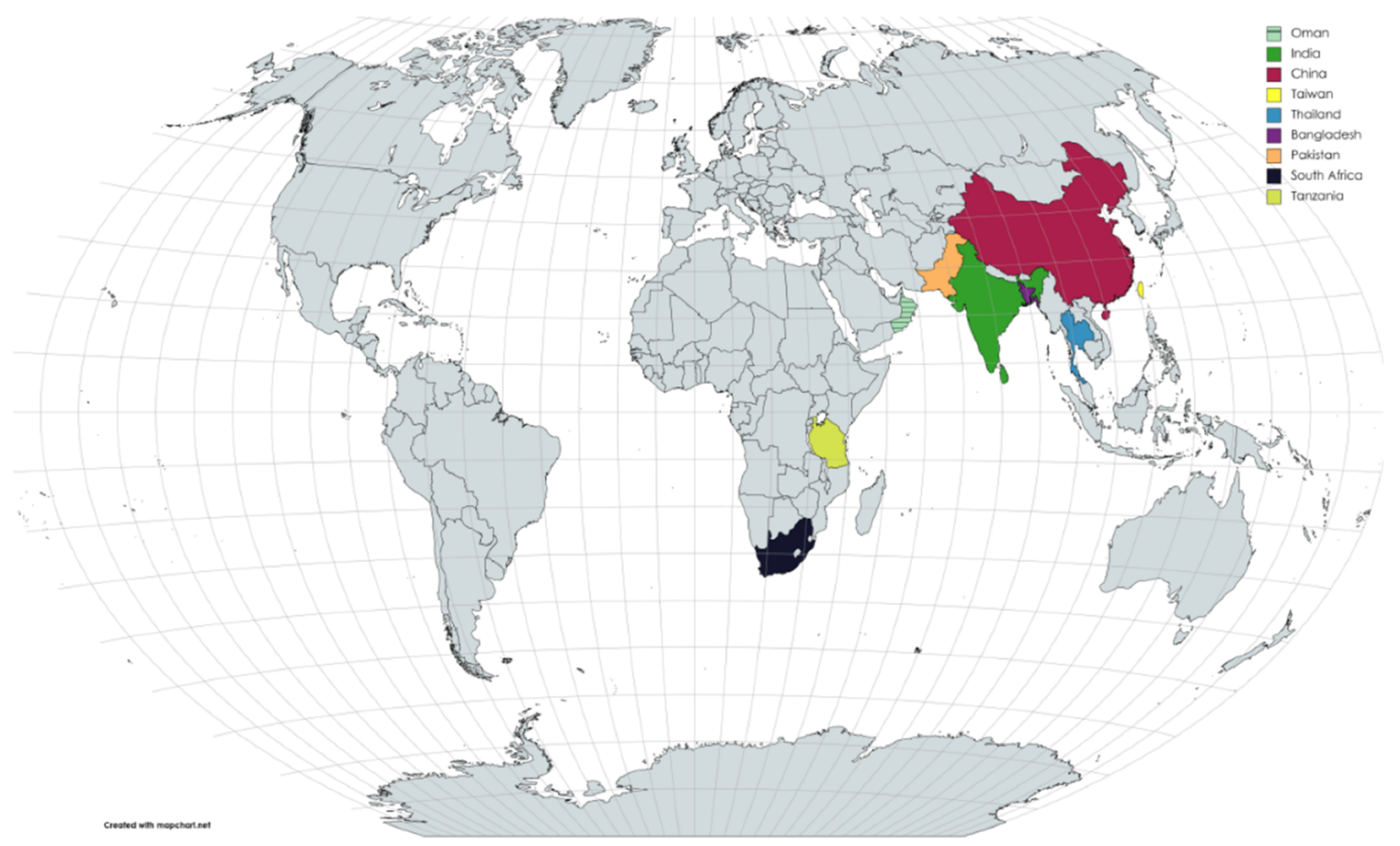
:1. Introduction
2. Materials and Methods
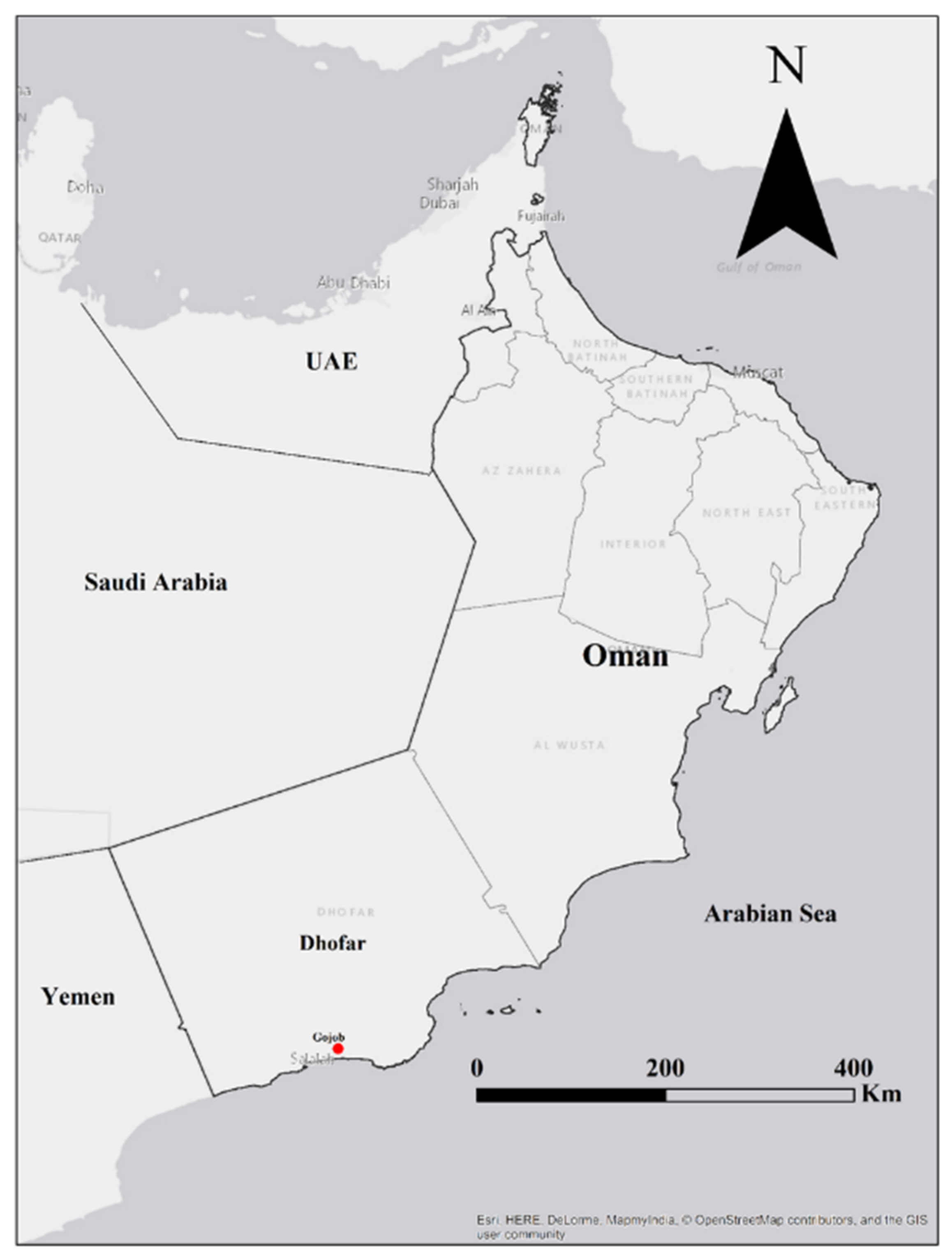
2.1. Sampling and Morphological Observations
2.2. Molecular Identification
2.3. Phylogenetic Analyses
3. Results
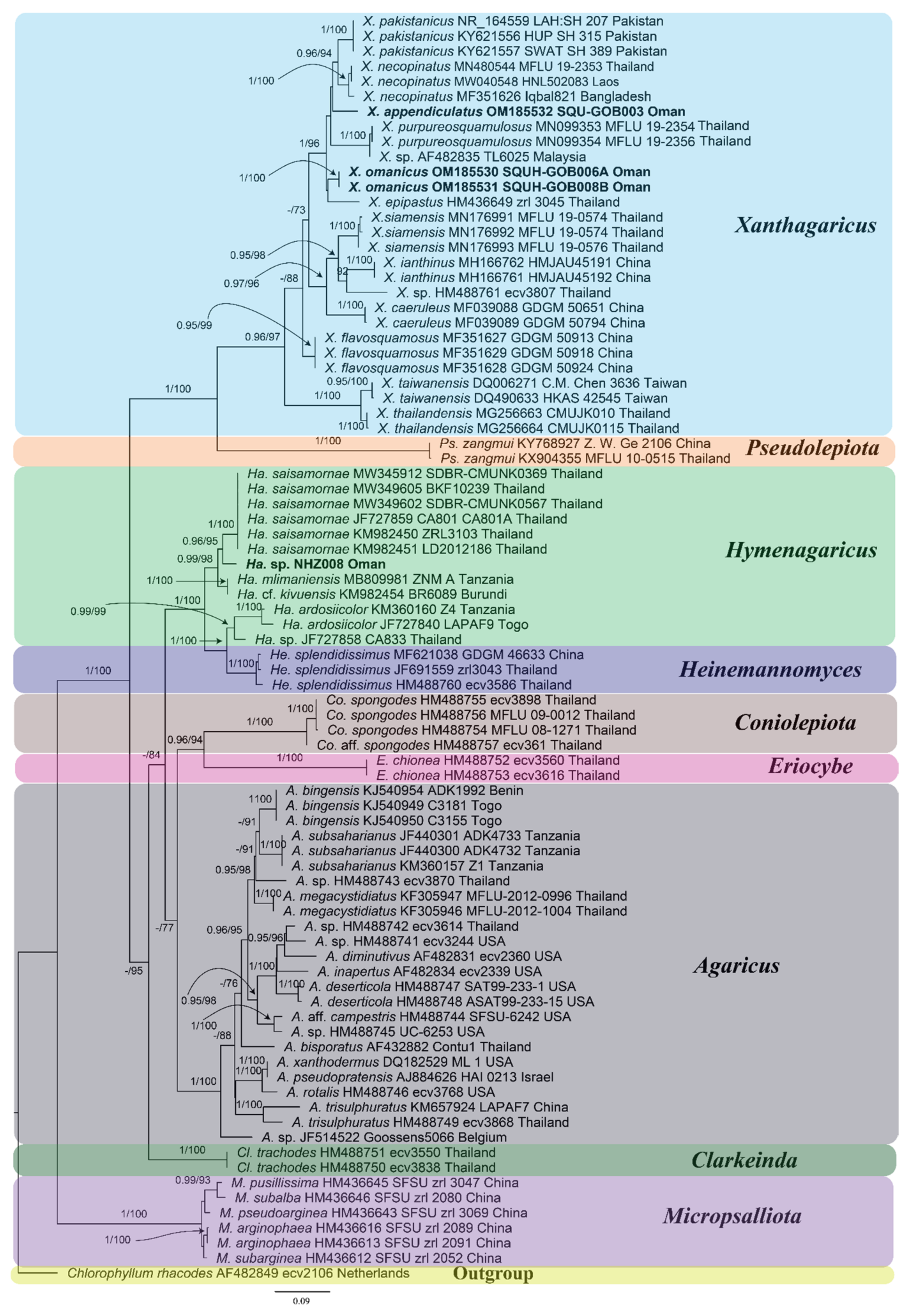
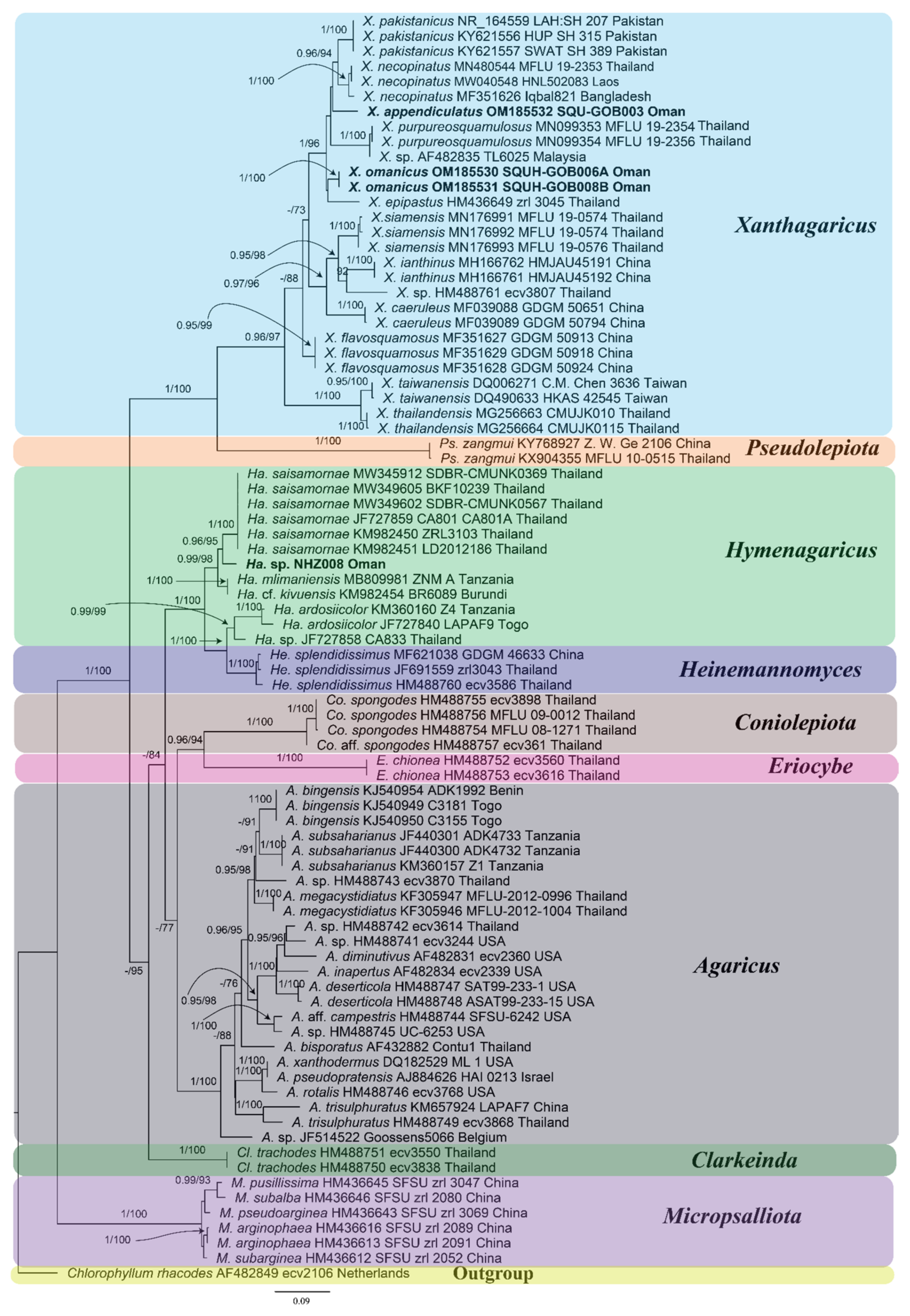
3.1. Molecular Phylogenetic Analyses
3.2. Taxonomy
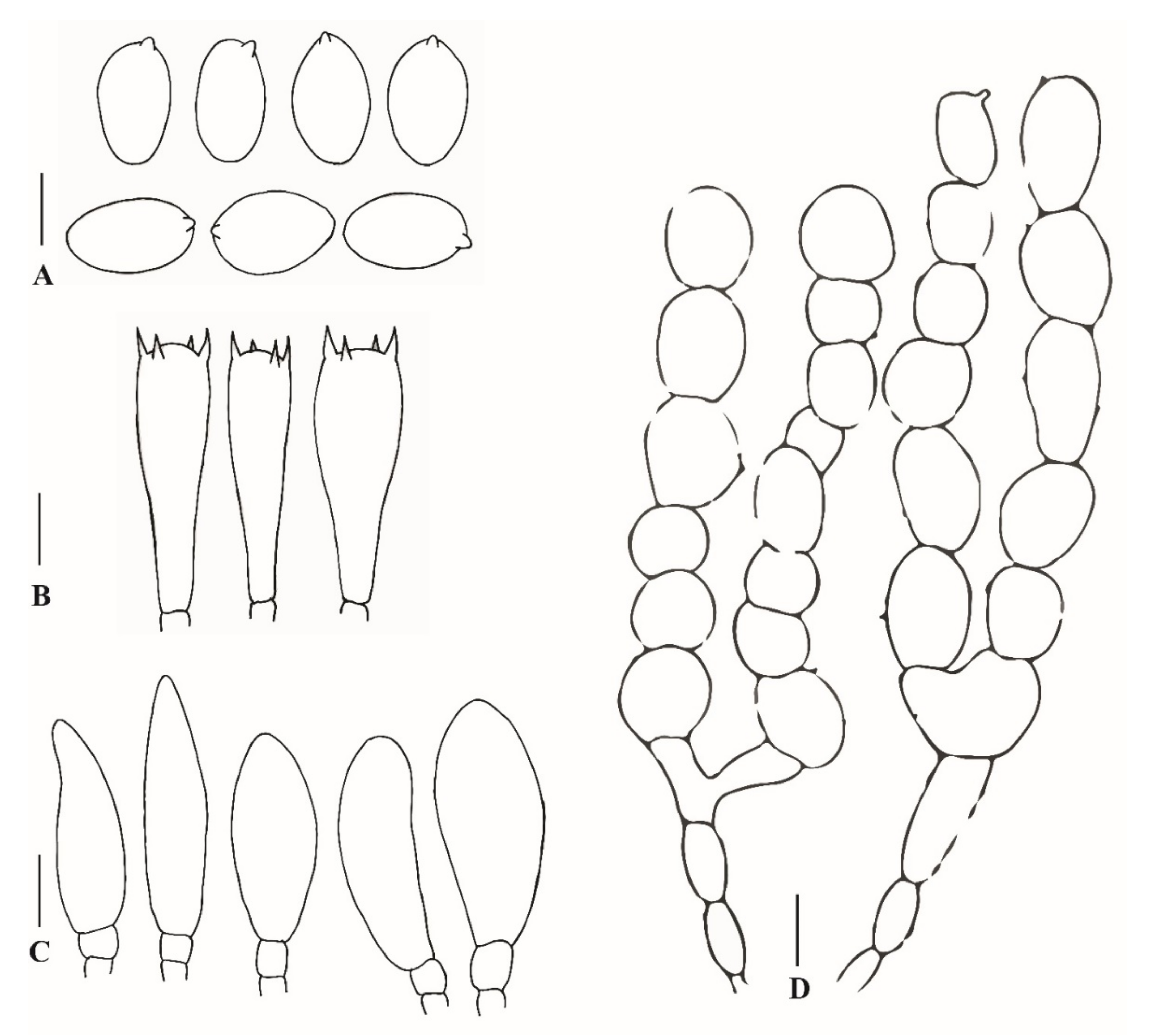
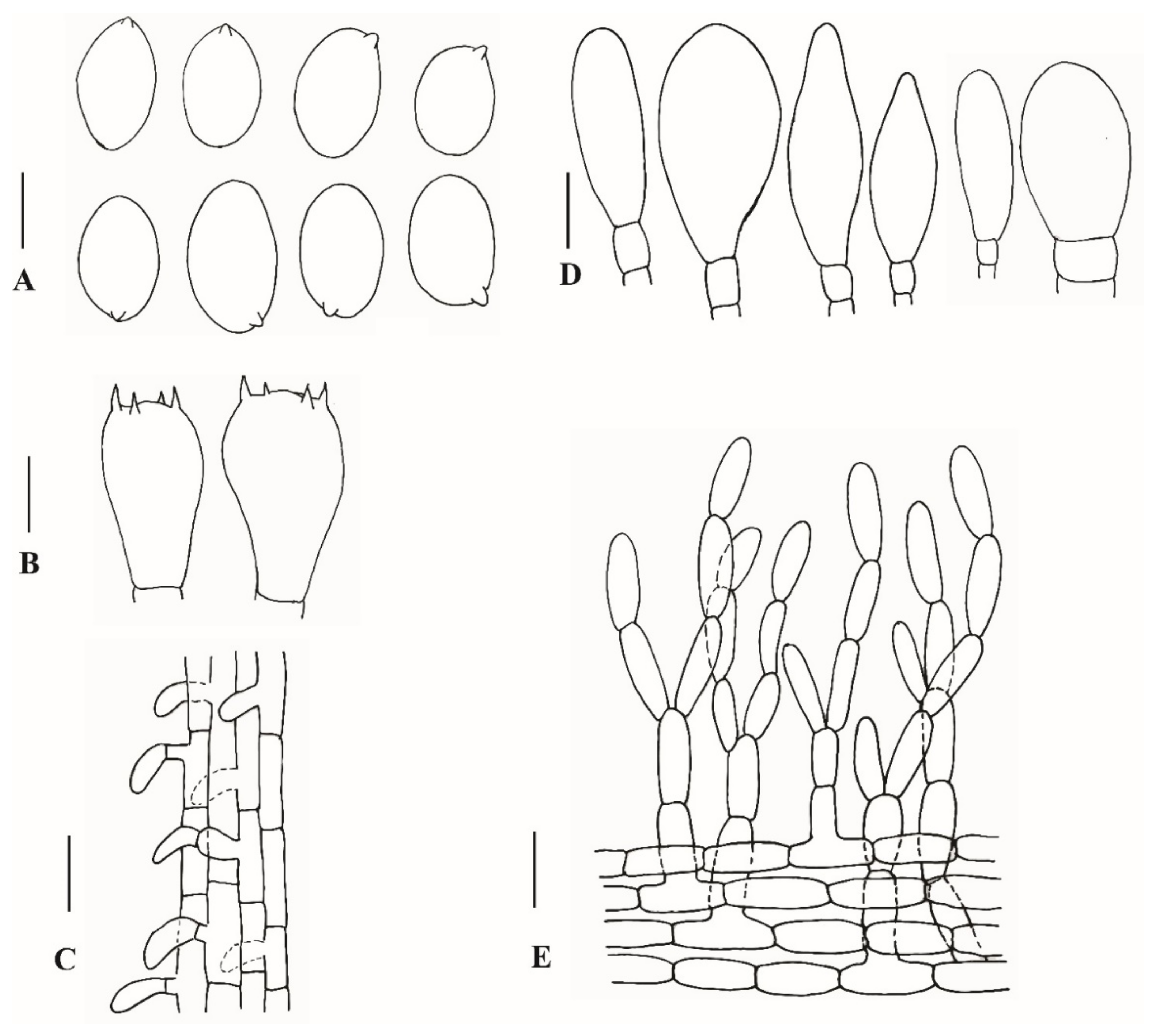
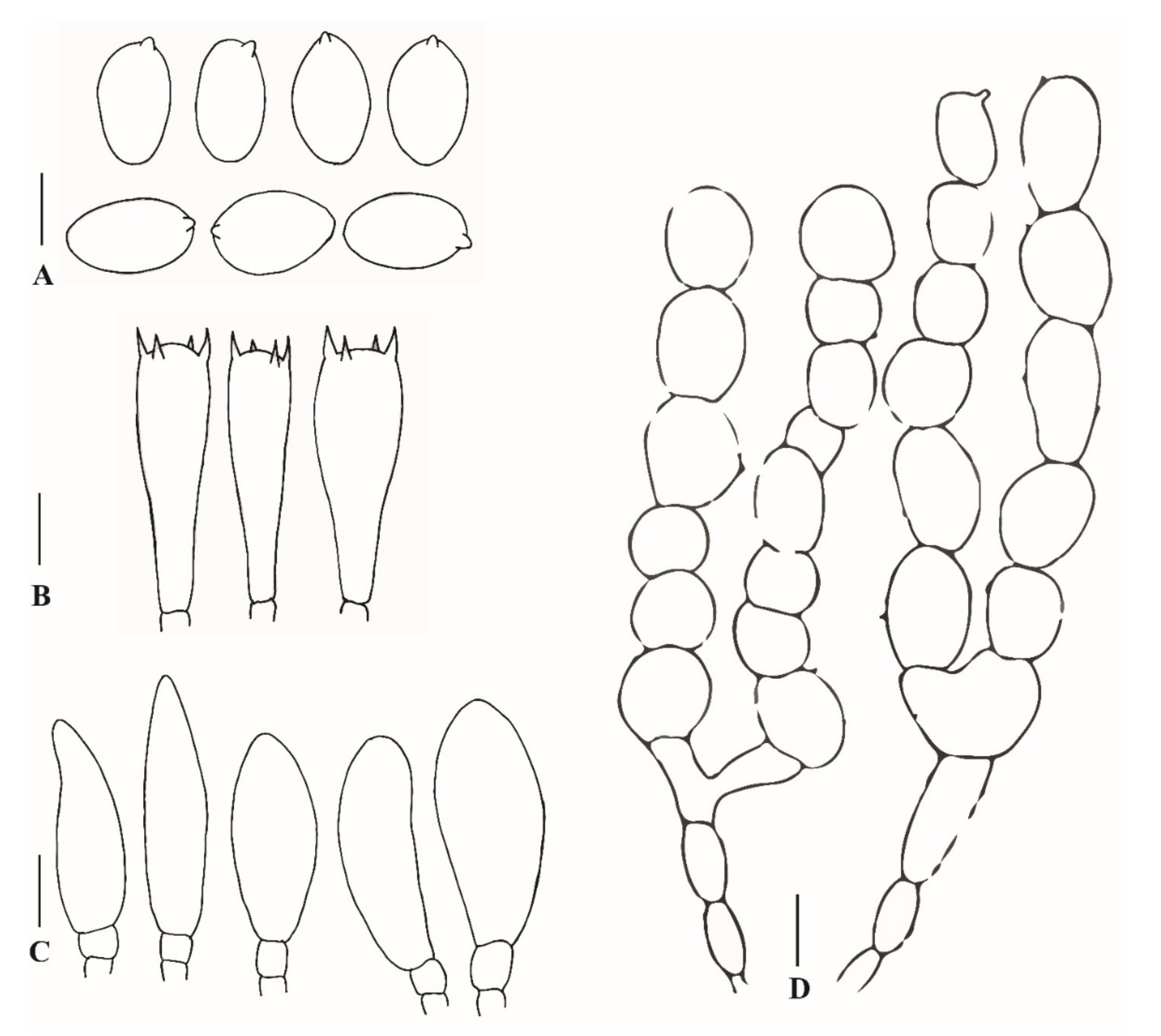
3.2.1. Xanthagaricus appendiculatus Al-Sadi and S. Hussain, sp. nov.
3.2.2. Xanthagaricus omanicus Al-Kharousi, Al-Sadi and S. Hussain, sp. nov.
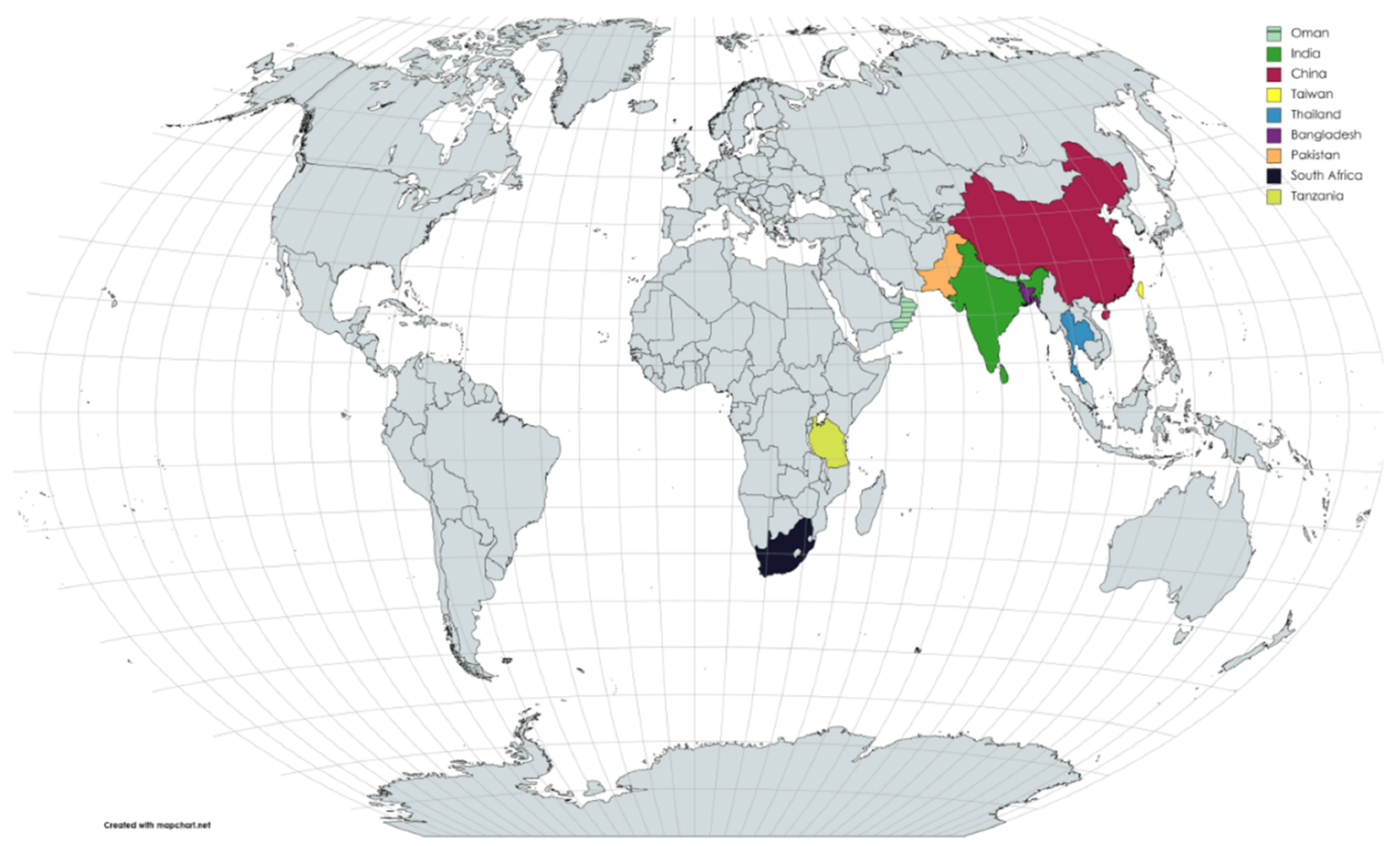
4. Discussion
| 1 | Basidiomata small-sized with pileus (3–15 mm diam) | 2 |
| - | Basidiomata small-sized with pileus >15 mm diam | 13 |
| 2 | Basidiospores size up to 5 µm in length | 3 |
| - | Basidiospores size >5 µm in length | 9 |
| 3 | Pileus covered with yellowish or brownish squamules | 4 |
| - | Pileus 3–9 mm diam, covered with violet squamules, basidiospores 4.2–4.8 × 2.9–3.4 µm | X. ianthinus |
| 4 | Pileus convex to plano-convex | 5 |
| - | Pileus 5–15 mm diam, conico-campanulate, basidiospores 3.8–5 × 2.9–3.6 µm | X.viridulus |
| 5 | Pileus conico-convex or plano-convex | 6 |
| - | Pileus 9–12 mm diam, campanulate, basidiospores 4–5 × 3–3.2 µm | X. ochraceoluteus. |
| 6 | Pileus conico-convex, smokey brown or yellowish | 7 |
| - | Pileus 4–6 mm diam, ochre-brown, plano-convex, basidiospores 3.7–4.4 × 2.6–3 µm | X. myriostictus. |
| 7 | Pileus brownish, conico-convex, covered with brownish-squamules | 8 |
| - | Pileus 10–15 mm diam, brownish, plano-convex, basidiospores 4–5 × 2.7–3.2 µm | X. necopinatus. |
| 8 | Pileus covered with pale brownish squamules, basidiopsores, subglobse, 4–6 × 4.1–3.8 µm | X. globisporus. |
| - | Pileus with smokey squamules, basidiopsores, ellipsoid, 4.1–4.9 × 2.9–3.4 µm | X. rubescens. |
| 9 | Pileus plano-convex, covered with yellowish to brownish squamules | 10 |
| - | Pileus 10–15 mm diam, with grayish-violet squamules, subhemispherical, basidiospores 5–6 × 3–3.5 µm | X. caeruleus |
| 10 | Pileus with yellowish-brown or violet-brown squamules | 11 |
| - | Pileus 10–15 mm diam, greenish squamules, basidiospores 5–5.5 × 3.3–4 µm | X. subepipastus. |
| 11 | Pileus with yellowish or yellowish-brown squamules | 12 |
| - | Pileus 4.5–10 mm diam, violet-brown squamules, basidiospores 4.6–6 × 3.5–4 µm | X. purpureosquamulosus. |
| 12 | Pileus 5–10 mm diam, convex with yellowish squamules, basidiospores 5–6.3 × 3.5–4.2 µm | X. gracilis |
| - | Pileus 8–13 mm diam, with yellowish-brown squamules, basidiospores 5–5.5 × 3–3.5 µm | X. flavosquamusus. |
| 13 | Pileus 15–20 mm diam | 14 |
| - | Basidiomata medium-sized, pileus >20 mm diam | 18 |
| 14 | Basidiospores <5.5 µm in length | 15 |
| - | Basidiospores >5.5 µm in length | 17 |
| 15 | Pileus convex to campanulate | 16 |
| - | Pileus chestnut-brown, conico-convex, basidiospores ellipsoid, 4.3–5.1 × 2.7–3.1 µm | X.erinaceus. |
| 16 | Pileus dark brown, convex, basidiopsores ellipsoid, yellowish, 3.7–4.7 × 2.8–3.4 µm | X. epipastus. |
| - | Pileus campanulate, basidiospores globose to subglobose, 4.2–5 × 2.9–3.5 µm | X. rufomarginatus. |
| 17 | Pileus conico-convex, covered with yellowish-brown squamules, basidiospores globose to broadly ellipsoid, 7–7.5 × 6–7 µm | X. pakistanicus. |
| - | Pileus plano-convex, covered with purplish squamules, basidiospores ellipsoid to amygdaliform, 6.5–7.5 × 4–5 µm | X. appendiculatus. |
| 18 | Pileus 20–50 mm diam | 19 |
| - | Pileus 70–90 mm diam, applanate, covered with yellowish squamules, basidiospores ellipsoid to broadly ellipsoid, 7–8.5 × 5–5.5 µm | X. omanicus. |
| 19 | Pileus convex to plano-convex, with brownish squamules | 20 |
| - | Pileus conico-campanulate, squamules pale orange, basidiospores ellipsoid to elongate, yellowish-brown, 6–7.5 × 4–4.5 µm | X. thailandensis. |
| 20 | Basidiospores ellipsoid, yellowish to pale-brownish | 21 |
| - | Basidiospores ovoid, brownish-yellow | 24 |
| 21 | Basidiospores ellipsoid with truncate base or oblong | 22 |
| - | Basidiospores ellipsoid with regular base, yellowish or brownish | 23 |
| 22 | Basidiospores ellipsoid to oblong, yellowish-brown, 4–5.5 × 2.5–3 µm | X. siamensis. |
| - | Basidiospores ellipsoid with truncate base, brownish-yellow, 5–6 × 3.4–4.2 µm | X. luteolosporus. |
| 23 | Basidiospores golden-yellow, 6–7.4 × 4–4.8 µm | X. chrysosporus. |
| - | Basidiospores brownish-yellow, 5–5.5 × 3–4 µm | X. taiwanensis. |
| 24 | Basidiospores ovoid with truncate base, yellowish-brown, 4.2–5 × 2.9–3.5 µm | X. subaeruginosus. |
| - | Basidiospores without trancate base, brownish-yellow, 4.9–5.8 × 3.7–4.3 µm | X. calicutensis. |
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heinemann, P.; Little Flower, S. Hymenagaricus (Agaricaceae) de Kerala (Inde) et de Sri Lanka. Bull. Jard. Bot. Natl. Belg. 1984, 54, 151–182. [Google Scholar] [CrossRef]
- Little Flower, S.R.; Hosagoudar, V.B.; Abraham, T.K. Xanthagaricus, a new generic name in the family Agaricaceae. New Bot. 1997, 24, 93–100. [Google Scholar]
- Vellinga, E.C.; Sysouphanthong, P.; Hyde, K.D. The family Agaricaceae: Phylogenies and two new white-spored genera. Mycologia 2011, 103, 494–509. [Google Scholar] [CrossRef] [Green Version]
- Kirk, P.M.; Canon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008. [Google Scholar]
- Hosen, M.I.; Song, Z.P.; Gates, G.; Karunarathna, S.C.; Chowdhury, M.S.M.; Li, T.H. Two new species of Xanthagaricus and some notes on Heinemannomyces from Asia. MycoKeys 2017, 28, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Afshan, N.S.; Ahmad, H.; Sher, H.; Khalid, A.N. Xanthagaricus pakistanicus sp. nov. (Agaricaceae), the first report of the genus from Pakistan. Turk. J. Bot. 2018, 42, 123–133. [Google Scholar] [CrossRef]
- Ge, Z.W.; Chen, C.M.; Yang, Z.L. A new species of the genus Hymenagaricus (Basidiomycota) from Taiwan and its phylogenetic position inferred from ITS and nLSU sequences. Cryptogam. Mycol. 2008, 29, 259–265. [Google Scholar]
- Pegler, D.N. Agaric Flora of Sri Lanka. Kew Bull. Addit. Ser. 1986, 12, 1–519. [Google Scholar]
- Kumla, J.; Suwannarach, N.; Sringernyuang, K.; Lumyong, S. Xanthagaricus thailandensis sp. nov. (Agaricales, Basidiomycota), from northern Thailand. Phytotaxa 2018, 348, 109–117. [Google Scholar] [CrossRef]
- Liu, S.Y.; Liu, J.K.; Syouphanthong, P.; Hyde, K.D.; Lumyong, S. Xanthagaricus siamensis sp. nov. (Agaricaceae), a new species with dull green lamellae from northern Thailand. Phytotaxa 2020, 437, 14–22. [Google Scholar] [CrossRef]
- Sysouphanthong, P.; Thongklang, N.; Liu, J.-K.; Vellinga, E.C. Description of Lepiotaceous Fungal Species of the Genera Chlorophyllum, Clarkeinda, Macrolepiota, Pseudolepiota, and Xanthagaricus, from Laos and Thailand. Diversity 2021, 13, 666. [Google Scholar] [CrossRef]
- Reid, D.A.; Eicker, A. The genus Hymenagaricus Heinem. in South Africa. S. Afr. J. Bot. 1995, 61, 293–297. [Google Scholar] [CrossRef] [Green Version]
- Reid, D.A.; Eicker, A. South African Fungi 8. Three new species of Hymenagaricus from South Africa, with a revised key to South African taxa. S. Afr. J. Bot. 1998, 64, 356–360. [Google Scholar] [CrossRef] [Green Version]
- Hosen, M.I.; Song, Z.; Gates, G.; Li, T.H. Xanthagaricus caeruleus, a new species with ink-blue lamellae from southeast China. Mycoscience 2017, 59, 190–194. [Google Scholar] [CrossRef]
- Wang, F.; Qi, L.L.; Zhou, X.; Li, Y. A new species and a new record of Xanthagaricus (Agaricaceae, Agaricales) from China. Phytotaxa 2018, 371, 241–250. [Google Scholar] [CrossRef]
- Henderson, J.C. The development of tourist destinations in the Gulf: Oman and Qatar compared. Tour. Plan. Dev. 2015, 12, 350–361. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Al-Sadi, A.M.; Al-Kharousi, M.; Al-Saady, N.; Hyde, K.D. A checklist of fungi in Oman. Phytotaxa 2016, 273, 219–261. [Google Scholar] [CrossRef]
- Fouda, M.M.; Hermosa, G.V., Jr.; Al-Harthi, S.M. Status of fish biodiversity in the Sultanate of Oman. Convention on Biological Diversity, First National Report. Dir. Nat. Reserve Sultanate Oman 1997, 19, 21. [Google Scholar]
- Carranza, S.; Xipell, M.; Tarroso, P.; Gardner, A.; Arnold, E.N.; Robinson, M.D.; Simó-Riudalbas, M.; Vasconcelos, R.; de Pous, P.; Amat, F.; et al. Diversity, distribution and conservation of the terrestrial reptiles of Oman (Sauropsida, Squamata). PLoS ONE 2018, 13, e0190389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sharhan, A.S.; Rizk, Z.A.; Nairn, A.E.M.; Bakhit, D.W.; Alhajari, S.A. (Eds.) Hydrogeology of an Arid Region: The Arabian Gulf and Adjoining Areas; Elsevier: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Burns, S.J.; Fleitmann, D.; Mudelsee, M.; Neff, U.; Matter, A.; Mangini, A. A 780-year annually resolved record of Indian Ocean monsoon precipitation from a speleothem from south Oman. J. Geophys. Res. Atmos. 2002, 107, ACL-9-1–ACL-9-9. [Google Scholar] [CrossRef] [Green Version]
- Edgell, H.S. Arabian Deserts: Nature, Origin and Evolution; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- El-Sheikh, M.A. Population structure of woody plants in the arid cloud forests of Dhofar, southern Oman. Acta Bot. Croat. 2013, 72, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Sale, J.B. The environment of the mountain region of Dhofar. J. Oman Stud. Spec. Rep. 1980, 2, 17–54. [Google Scholar]
- Nizamuddin, M.; Campbell, A.C. Glossophorella, a new genus of the family Dictyotaceae (Dictyotales-Phaeophyta) and its ecology from the coast of the Sultanate of Oman. Pak. J. Bot. 1995, 27, 257–262. [Google Scholar]
- Patzelt, A.; Harrison, T.; Knees, S.G.; Al Harthy, L. Studies in the flora of Arabia: XXXI. New records from the Sultanate of Oman. Edinb. J. Bot. 2014, 71, 161–180. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Waller, J.M.; Bridge, J. Plant diseases and nematodes in the Sultanate of Oman. Proc. Natl. Acad. Sci. USA 1978, 24, 313–326. [Google Scholar] [CrossRef]
- Van Wyk, M.; Al-Adawi, A.O.; Khan, I.A.; Deadman, M.L.; Al-Jahwari, A.A.; Wingfield, B.D.; Ploetz, R.; Wingfield, M.J. Ceratocystis manginecans sp. nov., causal agent of a destructive mango wilt disease in Oman and Pakistan. Fungal Divers. 2007, 27, 213–230. [Google Scholar]
- Symanczik, S.; Błaszkowski, J.; Chwat, G.; Boller, T.; Wiemken, A.; Al-Yahya’ei, M.N. Three new species of arbuscular mycorrhizal fungi discovered at one location in a desert of Oman: Diversispora omaniana, Septoglomus nakheelum and Rhizophagus arabicus. Mycologia 2014, 106, 243–259. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Belda, M.; Holtanová, E.; Halenka, T.; Kalvová, J. Climate classification revisited: From Köppen to Trewartha. Climate Res. 2014, 59, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bookhagen, B.; Thiede, R.C.; Strecker, M.R. Abnormal monsoon years and their control on erosion and sediment flux in the high, arid northwest Himalaya. Earth Planet. Sci. Lett. 2005, 231, 131–146. [Google Scholar] [CrossRef]
- Miller, A.G. CPD Site SWA 1. Dhofar Fog Oasis. Oman and Yemen. In Centres of Plant Diversity; Davis, S.D., Heywood, V.H., Hamilton, A.G., Eds.; IUCN Publication Unit: Cambridge, UK, 1994; Volume 1, pp. 143–155. [Google Scholar]
- Munsell, A.H. Munsell Soil Color Charts; Macbeth: Newburgh, NY, USA, 1992. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes, application to the identification of mycorrhizae and rusts. Molec. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Al-Sadi, A.M.; Al-Said, F.A.; Al-Jabri, A.H.; Al-Mahmooli, I.H.; Al-Hinai, A.H.; De Cock, A.W. Occurrence and characterization of fungi and oomycetes transmitted via potting mixtures and organic manures. Crop. Prot. 2011, 30, 38–44. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit 5.0.9: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Molec. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Stadler, T. On incomplete sampling under birth-death models and connections to the sampling-based coalescent. J. Theor. Biol. 2009, 261, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the cipres science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New York, NY, USA; pp. 1–8. [Google Scholar]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. TRACER v 1.6. Computer Program and Documentation Distributed by the Authors. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 3 January 2022).
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, K.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131, Institute of Evolutionary 623 Biology, University of Edinburgh. Available online: http://treebioedacuk/software/figtree/ (accessed on 3 January 2022).
- Ge, Z.W.; Yang, Z.L. Pseudolepiota zangmui gen. et sp. nov. (Agaricaceae, Basidiomycota), a new white-spored mushroom from China. Phytotaxa 2017, 312, 247–255. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Pileus Diameter (mm) | Stipe Size (mm) | Basidiospores Size (µm) | Squamules Cells/Size (µm) | References |
|---|---|---|---|---|---|
| Xanthagaricus appendiculatus sp. nov. | 15–20 | 40–60 × 2–4 | 6.5–7.5 × 4.0–5.0 | Irregular epithelium/13.5–21.5 | Examined in the present study. |
| X. calicutensis | 30–40 | 45 × 4 | 4.9–5.8 × 3.7–4.3 | Sub-hymeniform/10–20 | [1] |
| X. caeruleus | 10–15 | 22–35 × 1.5–2 | 5–6 × 3–3.5 | Pseudoparenchymatous/9–21 × 9–15 | [5] |
| X. chrysosporus | 20–30 | 30 × 2 | 6.0–7.4 × 4.0–4.8 | Sub-hymeniform/11–14 | [1] |
| X. epipastus | 15–20 | 40 × 2 | 3.7–4.7 × 2.8–3.4 | Sub-hymeniform/10–27 | [1] |
| X. erinaceus | 15–20 | 30 × 1 | 4.3–5.1 × 2.7–3.1 | Hymeniform/10–24 × 6–8 | [1] |
| X. flavidorufus | 20–27 | 70 × 1 | 4.4–5.4 × 3.0–3.5 | Vesiculate/10 | [1,2] |
| X. flavosquamosus | 8–13 | 20–30 × 1.5–2 | 5–5.5 × 3–3.5 | Epithelium/6–12 × 6–10 | [14] |
| X. globisporus | 10 | 20 × 1 | 4.6–5.1 × 4.1–4.8 | Hymeniform, pseudoparenchymatous/10–25 × 8–25 | [1] |
| X. gracilis | 5–10 | 30–35 × 1 | 5.0–6.3 × 3.5–4.2 | Subhymeniform/8–22 × 8–14 | [1] |
| X. ianthinus | 3–9 | 12–24 × 0.8–1.5 | 4.2–4.8 × 2.9–3.4 | Epithelium/7.6–20.7 × 5.4–15.8 | [15] |
| X. luteolosporus | 30–40 | 50 × 4 | 5.0–6.0 × 3.4–4.2 | Hymeniform/7–30 | [1] |
| X. myriostictus | 4–6 | 16 × 1 | 3.7–4.4 × 2.6–3.0 | Hymeniform to pseudoparenchymatous/9–20 | [1] |
| X. necopinatus | 10–15 | 18–28 × 1.5–2 | 4–5 × 2.7–3.2 | Epithelium/9–15 × 6–10 | [5] |
| X. ochraceoluteus | 9–12 | 30 × 1 | 4.0–5.2 × 3.0–3.2 | Pseudoparenchymatous/5–15 | [13] |
| X. omanicus sp. nov. | 70–90 | 50–60 × 3–5 | 7.0–8.5 × 5.0–5.5 | Intricate trichoderm/3.5–7.0 | Examined in the present study. |
| X. pakistanicus | 10–20 | 10–15 × 1–2 | 7.0–7.5 × 6.5–7 | Pseudoparenchymatous epithelium/20–47 × 18–44 | [6] |
| X. purpureosquamulosus | 4.5–10 | 15–30 × 1–1.5 | 4.5–6 × 3.5–4 | Irregular epithelium/7.5–25 | [11] |
| X. rubescens | 10–16 | 20 × 1 | 4.1–4.9 × 2.9–3.4 | Pseudoparenchymatous/15–35 × 8–12 | [1] |
| X. rufomarginatus | 20 | 20 × 3 | 4.0–4.2 × 2.5–3.0 | Pseudoparenchymatous epithelium/7–17 × 5–10 | [12] |
| X. siamensis | 31–54 | 35–65 × 2–6 | 4.0–5.5 × 2.5–3.0 | Cutis/3.5–8 | [10] |
| X. subaeruginosus | 45 | 40(90) × 2 | 4.2–5.0 × 2.9–3.5 | Pseudoparenchymatous to hymeniform/22–40 × 14–35 | [1] |
| X. subepipastus | 10–15 | 15–20 × 1 | 4.8–5.3 × 3.3–3.9 | Subhymeniform to pseudoparenchymatous/11–20 | [1] |
| X. taiwanensis | 15–35 | 40–70 × 6–9 | 5.0–5.5 × 3.0–4.0 | Pseudoparenchymatous/10–20 × 8–12 | [7] |
| X. thailandensis | 30−45 | 35−55 × 5−8 | 6−7.5 × 4−4.5 | Pseudoparenchymatous/5−26 × 4.5−8.5 | [9] |
| X. viridulus | 5–15 | 10–25 × 1 | 3.8–5.0 × 2.9–3.6 | Pseudoparenchymatous to hymeniform/22–40 × 14–35 | [1] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Kharousi, M.; Hussain, S.; Al-Muharabi, M.A.; Al-Shabibi, Z.; Al-Maqbali, D.; Al-Balushi, A.H.; Al-Yahya’ei, M.N.; Al-Saady, N.; Velazhahan, R.; Al-Sadi, A.M. The Genus Xanthagaricus: An Updated Global Species Distribution and Phylogeny with the Description of Two New Species from Oman. J. Fungi 2022, 8, 173. https://doi.org/10.3390/jof8020173
Al-Kharousi M, Hussain S, Al-Muharabi MA, Al-Shabibi Z, Al-Maqbali D, Al-Balushi AH, Al-Yahya’ei MN, Al-Saady N, Velazhahan R, Al-Sadi AM. The Genus Xanthagaricus: An Updated Global Species Distribution and Phylogeny with the Description of Two New Species from Oman. Journal of Fungi. 2022; 8(2):173. https://doi.org/10.3390/jof8020173
Chicago/Turabian StyleAl-Kharousi, Moza, Shah Hussain, Marwa A. Al-Muharabi, Zahra Al-Shabibi, Dua’a Al-Maqbali, Abdullah H. Al-Balushi, Mohamed N. Al-Yahya’ei, Nadiya Al-Saady, Rethinasamy Velazhahan, and Abdullah M. Al-Sadi. 2022. "The Genus Xanthagaricus: An Updated Global Species Distribution and Phylogeny with the Description of Two New Species from Oman" Journal of Fungi 8, no. 2: 173. https://doi.org/10.3390/jof8020173
APA StyleAl-Kharousi, M., Hussain, S., Al-Muharabi, M. A., Al-Shabibi, Z., Al-Maqbali, D., Al-Balushi, A. H., Al-Yahya’ei, M. N., Al-Saady, N., Velazhahan, R., & Al-Sadi, A. M. (2022). The Genus Xanthagaricus: An Updated Global Species Distribution and Phylogeny with the Description of Two New Species from Oman. Journal of Fungi, 8(2), 173. https://doi.org/10.3390/jof8020173