
Airborne Alt a 1 Dynamic and Its Relationship with the Airborne Dynamics of Alternaria Conidia and Pleosporales Spores



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling of Airborne Fungal Spores
2.3. Sampling of Airborne Allergens
2.4. Meteorological Data
2.5. Statistical Analysis
3. Results
3.1. Summary of Aerobiological and Meteorological Results
3.2. Airborne Fungal Aeroallergens vs. Spores and vs. Meteorology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horner, W.E.; Helbling, A.; Salvaggio, J.E.; Lehrer, S.B. Fungal allergens. Clin. Microbiol. Rev. 1995, 8, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Twaroch, T.E.; Curin, M.; Valenta, R.; Swoboda, I. Mold allergens in Respiratory Allergy: From Structure to Therapy. Allergy Asthma Immunol. Res. 2015, 7, 205–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez-Pereira, A.M.; De Linares, C.; Canela, M.; Belmonte, J. Spatial distribution of fungi from the analysis of aerobiological data with a gamma function. Aerobiologia 2021, 37, 461–477. [Google Scholar] [CrossRef]
- Kołodziejczyk, K.; Bożek, A.; Jarząb, J.; Gawlik, R. The clinical differences of asthma in patients with molds allergy. Adv. Respir. Med. 2016, 84, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutsen, A.P.; Bush, R.K.; Demain, J.G.; Denning, D.W.; Dixit, A.; Fairs, A.; Greenberger, P.A.; Kariuki, B.; Kita, H.; Kurup, V.P.; et al. Fungi and allergic lower respiratory tract diseases. J. Allergy Clin. Immunol. 2012, 129, 280–291. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [Green Version]
- Simon-Nobbe, B.; Denk, U.; Pöll, V.; Rid, R.; Breitenbach, M. The spectrum of fungal allergy. Int. Arch. Allergy Immunol. 2008, 145, 58–86. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, J.G.; Benham, R.W.; Kesten, B.M. Asthma due to a fungus-Alternaria. JAMA 1930, 94, 6. [Google Scholar] [CrossRef]
- D’Amato, G.; Chatzigeorgiou, G.; Corsico, R.; Gioulekas, D.; Jäger, L.; Jäger, S.; Kontou-Fili, K.; Kouridakis, S.; Liccardi, G.; Meriggi, A.; et al. Evaluation of the prevalence of skin prick test positivity to Alternaria and Cladosporium in patients with suspected respiratory allergy. Allergy 1997, 52, 711–716. [Google Scholar] [CrossRef]
- Pereira, C.; Valero, A.; Loureiro, C.; Davila, I.; Martinez-Cocera, C.; Murio, C.; Rico, P.; Palomino, R. Iberian study of aeroallergens sensitisation in allergic rhinitis. Eur. Ann. Allergy Clin. Immunol. 2006, 38, 186. [Google Scholar]
- Bartra, J.; Belmonte, J.; Torres-Rodriguez, J.M.; Cistero-Bahima, A. Sensitization to Alternaria in patients with respiratory allergy. Front. Biosci. 2009, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crameri, R.; Garbani, M.; Rhyner, C.; Huitema, C. Fungi: The neglected allergenic sources. Allergy 2014, 69, 176–185. [Google Scholar] [CrossRef]
- Ozdemir, O. Molds and respiratory allergy—Part 1. MOJ Immunol. 2015, 2, 00045. [Google Scholar] [CrossRef]
- López Couso, V.P.; Tortajada-Girbés, M.; Rodriguez Gil, D.; Martínez Quesada, J.; Palacios Pelaez, R. Fungi Sensitization in Spain: Importance of the Alternaria alternata Species and Its Major Allergen Alt a 1 in the Allergenicity. J. Fungi 2021, 7, 631. [Google Scholar] [CrossRef]
- Singh, S.M.; Naidu, J.; Pouranik, M. Ungual and cutaneous phaeohyphomycosis caused by Alternaria alternata and Alternaria chlamydospora. J. Med. Vet. Mycol. 1990, 28, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef]
- Miller, R.I. Nodular granulomatous fungal skin diseases of cats in the United Kingdom: A retrospective review. Vet. Dermatol. 2010, 21, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Genovese, L.M.; Whitbread, T.J.; Campbell, C.K. Cutaneous nodular phaeohyphomycosis in five horses associated with Alternaria alternata infection. Vet. Rec. 2001, 148, 55. [Google Scholar] [CrossRef]
- Gruber-Schley, S.; Thalmann, A. Zum Vorkommen von Alternaria spp. und deren Toxine in Getreide und mögliche Zusammenhänge mit Leistungsminderungen landwirtschaftlicher Nutztiere. Landwirtsch. Forsch 1988, 41, 11–29. [Google Scholar]
- Simmons, E.G. Alternaria: An identification manual. CBS Biodivers. Ser. 2007, 6, 644. [Google Scholar]
- Logrieco, A.; Bottalico, A.; Mulé, G.; Moretti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some Mediterranean crops. In Epidemiology of Mycotoxin Producing Fungi; Springer: Berlin/Heidelberg, Germany, 2003; pp. 645–667. [Google Scholar]
- Van der Waals, J.E.; Korsten, L.; Aveling, T.A.S.; Denner, F.D.N. Influence of environmental factors on field concentrations of Alternaria solani conidia above a South African potato crop. Phytoparasitica 2003, 31, 353–364. [Google Scholar] [CrossRef]
- Maya-Manzano, J.M.; Fernández-Rodríguez, S.; Hernández-Trejo, F.; Díaz-Pérez, G.; Gonzalo-Garijo, Á.; Silva-Palacios, I.; Muñoz-Rodríguez, A.F.; Tormo-Molina, R. Seasonal Mediterranean pattern for airborne spores of Alternaria. Aerobiologia 2012, 28, 515–525. [Google Scholar] [CrossRef]
- Grinn-Gofroń, A.; Nowosad, J.; Bosiacka, B.; Camacho, I.; Pashley, C.; Belmonte, J.; De Linares, C.; Ianovici, N.; Maya Manzano, J.M.; Sadyś, M.; et al. Airborne Alternaria and Cladosporium fungal spores in Europe: Forecasting possibilities and relationships with meteorological parameters. Sci. Total Environ. 2019, 653, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Vélez-Pereira, A.M.; De Linares, C.; Canela, M.A.; Belmonte, J. Logistic regression models for predicting daily airborne Alternaria and Cladosporium concentration levels in Catalonia (NE Spain). Int. J. Biometeorol. 2019, 63, 1541–1553. [Google Scholar] [CrossRef] [PubMed]
- Kustrzeba-Wójcicka, I.; Siwak, E.; Terlecki, G.; Wolańczyk-Mędrala, A.; Mędrala, W. Alternaria alternata and its allergens: A comprehensive review. Clin. Rev. Allergy Immunol. 2014, 47, 354–365. [Google Scholar] [CrossRef]
- De Vouge, M.W.; Thaker, A.J.; Curran, I.H.; Zhang, L.; Muradia, G.; Rode, H.; Vijay, H.M. Isolation and expression of a cDNA clone encoding an Alternaria alternata Alt a 1 subunit. Int. Arch. Allergy Immunol. 1996, 111, 385–395. [Google Scholar] [CrossRef]
- Wagner, G.E.; Gutfreund, S.; Fauland, K.; Keller, W.; Valenta, R.; Zangger, K. Backbone resonance assignment of Alt a 1, a unique β-barrel protein and the major allergen of Alternaria alternata. Biomol. NMR Assign. 2014, 8, 229–231. [Google Scholar] [CrossRef]
- Mitakakis, T.Z.; Barnes, C.; Tovey, E.R. Spore germination increases allergen release from Alternaria. J. Allergy Clin. Immunol. 2001, 107, 388–390. [Google Scholar] [CrossRef]
- Green, B.J.; Mitakakis, T.Z.; Tovey, E.R. Allergen detection from 11 fungal species before and after germination. J. Allergy Clin. Immunol. 2003, 111, 285–289. [Google Scholar] [CrossRef]
- Green, B.J.; Sercombe, J.K.; Tovey, E.R. Fungal fragments and undocumented conidia function as new aeroallergen sources. J. Allergy Clin. Immunol. 2005, 115, 1043–1048. [Google Scholar] [CrossRef]
- Rantio–Lehtimäki, A.; Viander, M.; Koivikko, A. Airborne birch pollen antigens in different particle sizes. Clin. Exp. Allergy 1994, 24, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Schäppi, G.F.; Taylor, P.E.; Pain, M.C.; Cameron, P.A.; Dent, A.W.; Staff, I.A.; Suphioglu, C. Concentrations of major grass group 5 allergens in pollen grains and atmospheric particles: Implications for hay fever and allergic asthma sufferers sensitized to grass pollen allergens. Clin. Exp. Allergy 1999, 29, 633–641. [Google Scholar] [CrossRef] [PubMed]
- De Linares, C.; Nieto-Lugilde, D.; Alba, F.; Díaz de la Guardia, C.; Galán, C.; Trigo, M.M. Detection of airborne allergen (Ole e 1) in relation to Olea europaea pollen in S Spain. Clin. Exp. Allergy 2007, 37, 125–132. [Google Scholar] [CrossRef] [PubMed]
- De Linares, C.; Diaz de la Guardia, C.; Lugilde, D.; Alba, F. Airborne study of grass allergen (Lol p 1) in different-sized particles. Int. Arch. Allergy Immunol. 2010, 152, 49–57. [Google Scholar] [CrossRef]
- Buters, J.T.; Weichenmeier, I.; Ochs, S.; Pusch, G.; Kreyling, W.; Boere, A.J.F.; Schober, W.; Behrendt, H. The allergen Bet v 1 in fractions of ambient air deviates from birch pollen counts. Allergy 2010, 65, 850–858. [Google Scholar] [CrossRef]
- Jato, V.; Rodríguez-Rajo, F.J.; González-Parrado, Z.; Elvira-Rendueles, B.; Moreno-Grau, S.; Vega-Maray, A.; Fernández-González, D.; Asturias, J.A.; Suárez-Cervera, M. Detection of airborne Par j 1 and Par j 2 allergens in relation to Urticaceae pollen counts in different bioclimatic areas. Ann. Allergy Asthma Immunol. 2010, 105, 50–56. [Google Scholar] [CrossRef]
- Hong, S.; Cramer, R.; Lawrence, C.; Pryora, B. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef]
- Sáenz-de-Santamaría, M.; Postigo, I.; Gutierrez-Rodríguez, A.; Cardona, G.; Guisantes, J.A.; Asturias, J.; Martinez, J. The major allergen of Alternaria alternata (Alt a 1) is expressed in other members of the Pleosporaceae family. Mycoses 2006, 49, 91–95. [Google Scholar] [CrossRef]
- Schmechel, D.; Green, B.; Blachere, F.; Janotka, E.; Beezhold, D. The Importance of antibody crossreactivity for the development of environmental immunoassays for Alternaria Alternata. J. Allergy Clin. Immunol. 2008, 121, S178. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.; Postigo, I.; Guisantes, J.; Suñén, E.; Martínez, J. Identification of allergens homologous to Alt a 1 from Stemphylium botryosum and Ulocladium botrytis. Med. Mycol. 2011, 49, 892–896. [Google Scholar]
- Postigo, I.; Gutiérrez-Rodríguez, A.; Fernández, J.; Guisantes, J.; Suñén, E.; Martínez, J. Diagnostic value of Alt a 1, fungal enolase and manganese-dependent superoxide dismutase in the component-resolved diagnosis of allergy to Pleosporaceae. Clin. Exp. Allergy 2011, 41, 443–451. [Google Scholar] [CrossRef]
- Chruszcz, M.; Chapman, M.; Osinski, T.; Solberg, R.; Demas, M.; Porebski, P.J.; Majorek, K.A.; Pomés, A.; Minor, W. Alternaria alternata allergen Alt a 1: A unique b-barrel protein dimer found exclusively in fungi. J. Allergy Clin. Immunol. 2012, 130, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teifoori, F.; Shams-Ghahfarokhia, M.; Razzaghi-Abyanehb, M.; Martinez, J. Gene profiling and expression of major allergen Alt a 1 in Alternaria alternata and related members of the Pleosporaceae family. Rev. Iberoam. Micol. 2019, 36, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Rúa-Guiraldo, A. Aerobiología de las Esporas de Pleosporales en Ambientes Intra y Extradomiciliarios de Barcelona. Aplicación a la Clínica en Población Alérgica. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, July 2013. Available online: https://www.tdx.cat/handle/10803/120550 (accessed on 10 November 2021).
- Bisht, V.; Arora, N.; Singh, B.P.; Pasha, S.; Gaur, S.N.; Sridhara, S. Epi p 1, an allergenic glycoprotein of Epicoccum purpurascens is a serine protease. FEMS Immunol. Med. Microbiol. 2004, 42, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allue Andrade, J.L. Phytoclimatic atlas of Spain. Taxonomies; Instituto Nacional de Investigaciones Agrarias, Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1990. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köpen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Chazarra, A.; Flórez, E.; Peraza, B.; Tohá, T.; Lorenzo, B.; Criado, E.; Moreno, J.V.; Romero, R.; Botey, R. Mapas Climáticos de España (1981–2010) y ETo (1996–2016); Ministerio para la Transición Ecológica, Agencia Estatal de Meteorología: Madrid, Spain, 2018. [Google Scholar] [CrossRef]
- Hirst, J.M. An automatic volumetric spore-trap. Ann. Appl. Biol. 1952, 39, 257–265. [Google Scholar] [CrossRef]
- Galán, C.; Cariñanos, P.; Alcázar, P.; Domínguez, E. Manual de Calidad y Gestión de la Red Española de Aerobiología; [Quality Manual and Management Spanish Aerobiology Network]; Servicio de Publicaciones de la Universidad de Córdoba: Cordoba, Spain, 2007. [Google Scholar]
- Galán, C.; Smith, M.; Thibaudon, M.; Frenguelli, G.; Oteros, J.; Gehrig, R.; Berger, U.; Clot, B.; Brandao, R.; the EAS QC Working Group. Pollen monitoring: Minimum requirements and reproducibility of analysis. Aerobiologia 2017, 30, 385–395. [Google Scholar] [CrossRef]
- Galán, C.; Ariatti, A.; Bonini, M.; Clot, B.; Crouzy, B.; Dahl, A.; Fernández-González, D.; Frenguelli, G.; Gehrig, R.; Isard, S.; et al. Recommended terminology for aerobiological studies. Aerobiología 2017, 33, 293–295. [Google Scholar] [CrossRef]
- De Linares, C.; Belmonte, J.; Delgado, R. New methodology for the measurement of airborne allergens. In Abstract Book of the Seventh European Symposium on Aerobiology: Bioaerosols and Environmental Impacts; European Aerobiology Society: Córdoba, Spain, 2020; p. 119. [Google Scholar]
- Indoor Biotechnologies. Available online: https://inbio.com/elisa-2.0/elisa-2.0-kits-mold (accessed on 10 November 2021).
- Cecchi, L. From pollen count to pollen potency: The molecular era of aerobiology. Eur. Respir. J. 2013, 42, 898–900. [Google Scholar] [CrossRef]
- Moreno-Grau, S.; Elvira-Rendueles, B.; Moreno, J.; García-Sánchez, A.; Vergara, N.; Asturias, J.A.; Arilla, M.C.; Ibarrola, I.; Seoane-Camba, J.A.; Suárez-Cervera, M. Correlation between Olea europaea and Parietaria judaica pollen counts and quantification of their major allergens Ole e 1 and Par j 1-Par j 2. Ann. Allergy Asthma Immunol. 2006, 96, 858–864. [Google Scholar] [CrossRef]
- Rodríguez-Rajo, F.J.; Jato, V.; González-Parrado, Z.; Elvira-Rendueles, B.; Moreno-Grau, S.; Vega-Maray, A.; Fernández-González, D.; Asturias, J.A.; Suárez-Cervera, M. The combination of airborne pollen and allergen quantification to re-liably assess the real pollinosis risk in different bioclimatic areas. Aerobiologia 2011, 27, 1–12. [Google Scholar] [CrossRef]
- Buters, J.T.; Thibaudon, M.; Smith, M.; Kennedy, R.; Rantio-Lehtimäki, A.; Albertini, R.; Reese, G.; Weber, B.; Galán, C.; Brandao, R.; et al. The HIALINE working group. Release of Bet v 1 from birch pollen from 5 European countries. Results from the HIALINE study. Atmos. Environ. 2012, 55, 496–505. [Google Scholar] [CrossRef] [Green Version]
- De Linares, C.; Plaza, M.P.; Valle, A.M.; Alcázar, P.; Díaz de la Guardia, C.; Galán, C. Airborne Cupressaceae Pollen and Its Major Allergen, Cup a 1, in Urban Green Areas of Southern Iberian Peninsula. Forests 2021, 12, 254. [Google Scholar] [CrossRef]
- O’Hollaren, M.T.; Yunginger, J.W.; Offord, K.P.; Somers, M.J.; O’Connell, E.J.; Ballard, D.J.; Sachs, M.I. Exposure to an aeroallergen as a possible precipitating factor in respiratory arrest in young patients with asthma. N. Engl. J. Med. 1991, 324, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Brito, F.F.; Alonso, A.M.; Carnés, J.; Martín-Martín, R.; Fernández-Caldas, E.; Galindo, P.A.; Alfaya, T.; Amo-Salas, M. Correlation between Alt a 1 levels and clinical symptoms in Alternaria alternata-monosensitized patients. J. Investig. Allergol. Clin. Immunol. 2012, 22, 154–159. [Google Scholar]
- Grewling, Ł.; Nowak, M.; Szymańska, A.; Kostecki, Ł.; Bogawski, P. Temporal variability in the allergenicity of airborne Alternaria spores. Med. Mycol. 2019, 57, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Plaza, M.P.; Alcázar, P.; Hernández-Ceballos, M.A.; Galán, C. Mismatch in aeroallergens and airborne grass pollen concentrations. Atmos. Environ. 2016, 144, 361–369. [Google Scholar] [CrossRef]
- Plaza, M.P.; Alcázar, P.; Galán, C. Correlation between airborne Olea europaea pollen concentrations and levels of the major allergen Ole e 1 in Córdoba, Spain, 2012-2014. Int. J. Biometeorol. 2016, 60, 1841–1847. [Google Scholar] [CrossRef]
- Hirhish, U. Study on the Biological air Quality in Bellaterra (Barcelona) and Vitoria-Gasteiz: Pollen and Spores and Their Allergens. Ph.D. Thesis, Universitat Autònoma de Barcelona, Barcelona, Spain, May 2019. Available online: https://www.tdx.cat/handle/10803/666755#page=1 (accessed on 10 November 2021).
- Querol, X.; Alastuey, A.; Rodriguez, S.; Plana, F.; Mantilla, E.; Ruiz, C.R. Monitoring of PM10 and PM2.5 around primary particulate anthropogenic emission sources. Atmos. Environ. 2001, 35, 845–858. [Google Scholar] [CrossRef]
- Agarwal, M.K.; Swanson, M.C.; Reed, C.E.; Yunginger, J.W. Immunochemical quantitation of airborne short ragweed, Alternaria, antigen E, and Alt-I allergens: A two-year prospective study. J. Allergy Clin. Immunol. 1983, 72, 40–45. [Google Scholar] [CrossRef]
- Martínez, J.; Gutiérrez, A.; Postigo, I.; Cardona, G.; Guisantes, J. Variability of Alt a 1 expression by different strains of Alternaria alternata. J. Investig. Allergol. Clin. Immunol. 2006, 16, 279–282. [Google Scholar] [PubMed]
- Grinn-Gofroń, A.; Bosiacka, B. Effects of meteorological factors on the composition of selected fungal spores in the air. Aerobiologia 2015, 31, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damialis, A.; Mohammad, A.B.; Halley, J.M.; Gange, A.C. Fungi in a changing world: Growth rates will be elevated, but spore production may decrease in future climates. Int. J. Biometeorol. 2015, 59, 1157–1167. [Google Scholar] [CrossRef]
- Sousa, L.; Camacho, I.C.; Grinn-Gofroń, A.; Camacho, R. Monitoring of anamorphic fungal spores in Madeira region (Portugal), 2003–2008. Aerobiologia 2016, 32, 303–315. [Google Scholar] [CrossRef]
- Grewling, Ł.; Bogawski, P.; Szymańska, A.; Nowak, M.; Kostecki, Ł.; Smith, M. Particle size distribution of the major Alternaria alternata allergen, Alt a 1, derived from airborne spores and subspore fragments. Fungal Biol. 2020, 124, 219–227. [Google Scholar] [CrossRef]


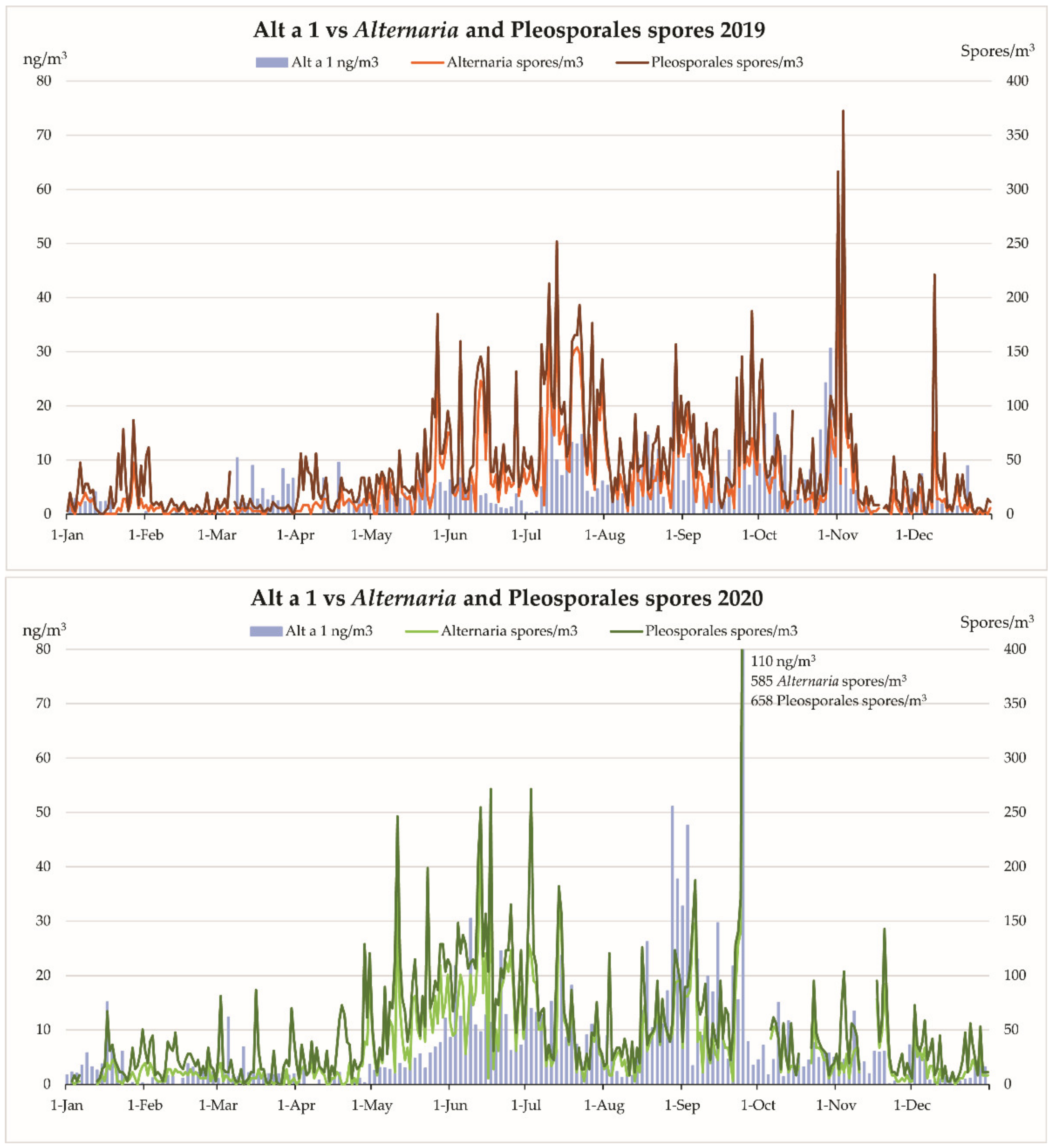{kind=link}
| 2017 | 2019 | 2020 | ||
|---|---|---|---|---|
| Multi/vial Cyclone sampler | Alt a 1 AAIn (ng·day/m3) | ND | __ | __ |
| Peak (ng/m3) | ND | __ | __ | |
| Peak day | ND | __ | __ | |
| Nr of days with Alt a 1 | 0 | __ | __ | |
| Nr of analyzed days | 359 | __ | __ | |
| High-volume sampler | Alt a 1 AAIn (ng·day/m3) | __ | 861.5 | 1352.0 |
| Peak (ng/m3) | __ | 30.6 | 109.7 | |
| Peak day | __ | 29-Oct | 25-Sep | |
| Nr of days with Alt a 1 | __ | 155 | 173 | |
| Nr of analyzed days | __ | 182 | 183 | |
| Hirst sampler | Alternaria spores ASIn (Spore·day/m3) | 10,343 | 10,108 | 11,768 |
| Peak (spores/m3) | 316 | 305 | 585 | |
| Peak day | 22-Oct | 3-Nov | 25-Sep | |
| Nr of days with Alternaria spores | 335 | 326 | 316 | |
| Analyzed days | 361 | 363 | 339 | |
| Pleosporales spores ASIn (Spore·day/m3) | 16,285 | 15,733 | 17,657 | |
| Peak (spores/m3) | 356 | 372 | 658 | |
| Peak day | 22-Oct | 3-Nov | 25-Sep | |
| Nr of days with Pleosporales spores | 357 | 356 | 335 | |
| Nr of analyzed days | 361 | 363 | 339 | |
| Meteorological data | TMax (°C) | 21.7 | 21.6 | 21.5 |
| TMin (°C) | 9.4 | 9.3 | 9.6 | |
| TMean (°C) | 15.6 | 15.5 | 15.5 | |
| P 0–24 (mm) | 468.8 | 553.0 | 847.2 | |
| RH (%) | 68.8 | 68.6 | 73.8 |
| 2017 | Alt a 1 | Alternaria spores | Pleosporales spores | TMax (°C) | TMin (°C) | TMean (°C) | P 0–24 (mm) | RH (%) | |
| Alt a 1 | Spearman Test | ||||||||
| N | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Alternaria spores | Spearman Test | 1.000 | 0.860 ** | 0.639 ** | 0.601 ** | 0.631 ** | −0.194 ** | −0.224 ** | |
| N | 0 | 362 | 362 | 362 | 362 | 362 | 362 | 362 | |
| Pleosporales spores | Spearman Test | 0.860 ** | 1.000 | 0.504 ** | 0.560 ** | 0.537 ** | 0.065 | −0.092 | |
| N | 0 | 362 | 362 | 362 | 362 | 362 | 362 | 362 | |
| 2019 | Alt a | Alternaria spores | Pleosporales spores | TMax (°C) | TMin (°C) | TMean (°C) | P 0–24 (mm) | RH (%) | |
| Alt a 1 | Spearman Test | 1.000 | 0.548 ** | 0.492 ** | 0.538 ** | 0.577 ** | 0.579 ** | −0.043 | −0.056 |
| N | 155 | 153 | 155 | 155 | 155 | 155 | 155 | 155 | |
| Alternaria spores | Spearman Test | 0.548 ** | 1.000 | 0.875 ** | 0.649 ** | 0.649 ** | 0.672 ** | −0.077 | −0.300 ** |
| N | 153 | 362 | 362 | 362 | 362 | 362 | 362 | 362 | |
| Pleosporales spores | Spearman Test | 0.492 ** | 0.875 ** | 1.000 | 0.541 ** | 0.619 ** | 0.599 ** | 0.163 * | −0.129 * |
| N | 155 | 362 | 362 | 362 | 362 | 362 | 362 | 362 | |
| 2020 | Alt a 1 | Alternaria spores | Pleosporales spores | TMax (°C) | TMin (°C) | TMean (°C) | P 0–24 (mm) | RH (%) | |
| Alt a 1 | Spearman Test | 1.000 | 0.631 ** | 0.556 ** | 0.566 ** | 0.550 ** | 0.571 ** | −0.016 | −0.229 * |
| N | 173 | 160 | 160 | 173 | 173 | 173 | 173 | 173 | |
| Alternaria spores | Spearman Test | 0.631 ** | 1.000 | 0.856 ** | 0.614 ** | 0.556 ** | 0.602 ** | −0.145 ** | −0.326 ** |
| N | 160 | 339 | 338 | 339 | 339 | 339 | 339 | 339 | |
| Pleosporales spores | Spearman Test | 0.556 ** | 0.856 ** | 1.000 | 0.476 ** | 0.535 ** | 0.518 ** | 0.138 * | −0.119 * |
| N | 160 | 338 | 338 | 338 | 338 | 338 | 338 | 338 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Linares, C.; Navarro, D.; Puigdemunt, R.; Belmonte, J. Airborne Alt a 1 Dynamic and Its Relationship with the Airborne Dynamics of Alternaria Conidia and Pleosporales Spores. J. Fungi 2022, 8, 125. https://doi.org/10.3390/jof8020125
De Linares C, Navarro D, Puigdemunt R, Belmonte J. Airborne Alt a 1 Dynamic and Its Relationship with the Airborne Dynamics of Alternaria Conidia and Pleosporales Spores. Journal of Fungi. 2022; 8(2):125. https://doi.org/10.3390/jof8020125
Chicago/Turabian StyleDe Linares, Concepción, David Navarro, Rut Puigdemunt, and Jordina Belmonte. 2022. "Airborne Alt a 1 Dynamic and Its Relationship with the Airborne Dynamics of Alternaria Conidia and Pleosporales Spores" Journal of Fungi 8, no. 2: 125. https://doi.org/10.3390/jof8020125
APA StyleDe Linares, C., Navarro, D., Puigdemunt, R., & Belmonte, J. (2022). Airborne Alt a 1 Dynamic and Its Relationship with the Airborne Dynamics of Alternaria Conidia and Pleosporales Spores. Journal of Fungi, 8(2), 125. https://doi.org/10.3390/jof8020125