
Mycobiome Diversity of the Cave Church of Sts. Peter and Paul in Serbia—Risk Assessment Implication for the Conservation of Rare Cavern Habitat Housing a Peculiar Fresco Painting


 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling Points
2.2. Indoor Microclimate and Moisture Content Measurements
2.3. Determination of Surface Contamination
2.4. In Situ Microscopy
2.5. Optical Microscopy
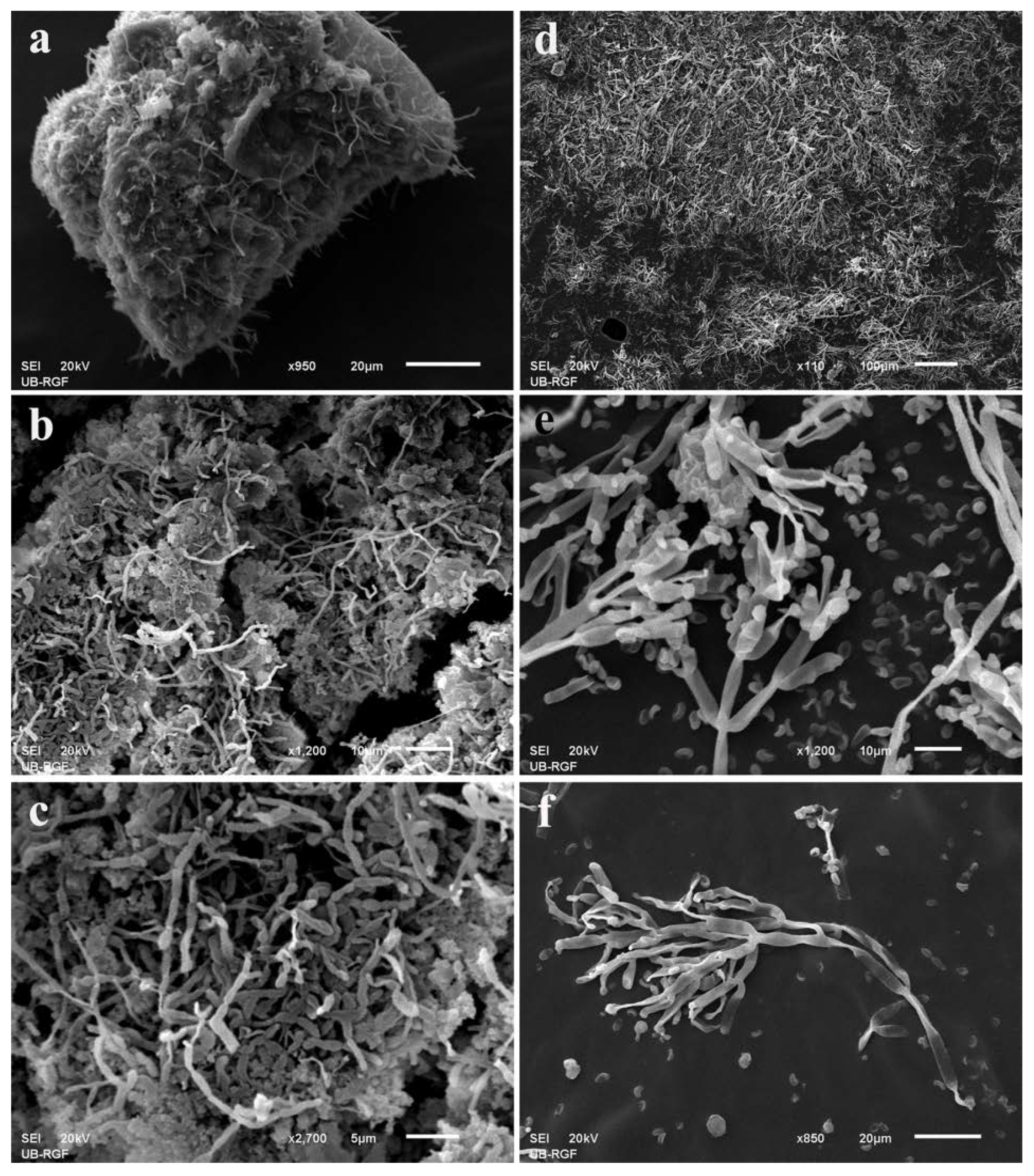
2.6. Scanning Electron Microscopy (SEM)
2.7. Isolation and Identification of Culturable Fungi
2.8. Biodegradation Plate Assays
2.9. Amplicon Sequencing
2.9.1. DNA Extraction, Library Preparation, and NGS Sequencing
2.9.2. Sequence Data Process and Taxonomy Annotation
2.9.3. Statistical Analysis
3. Results
3.1. State of the Investigated Cave Church
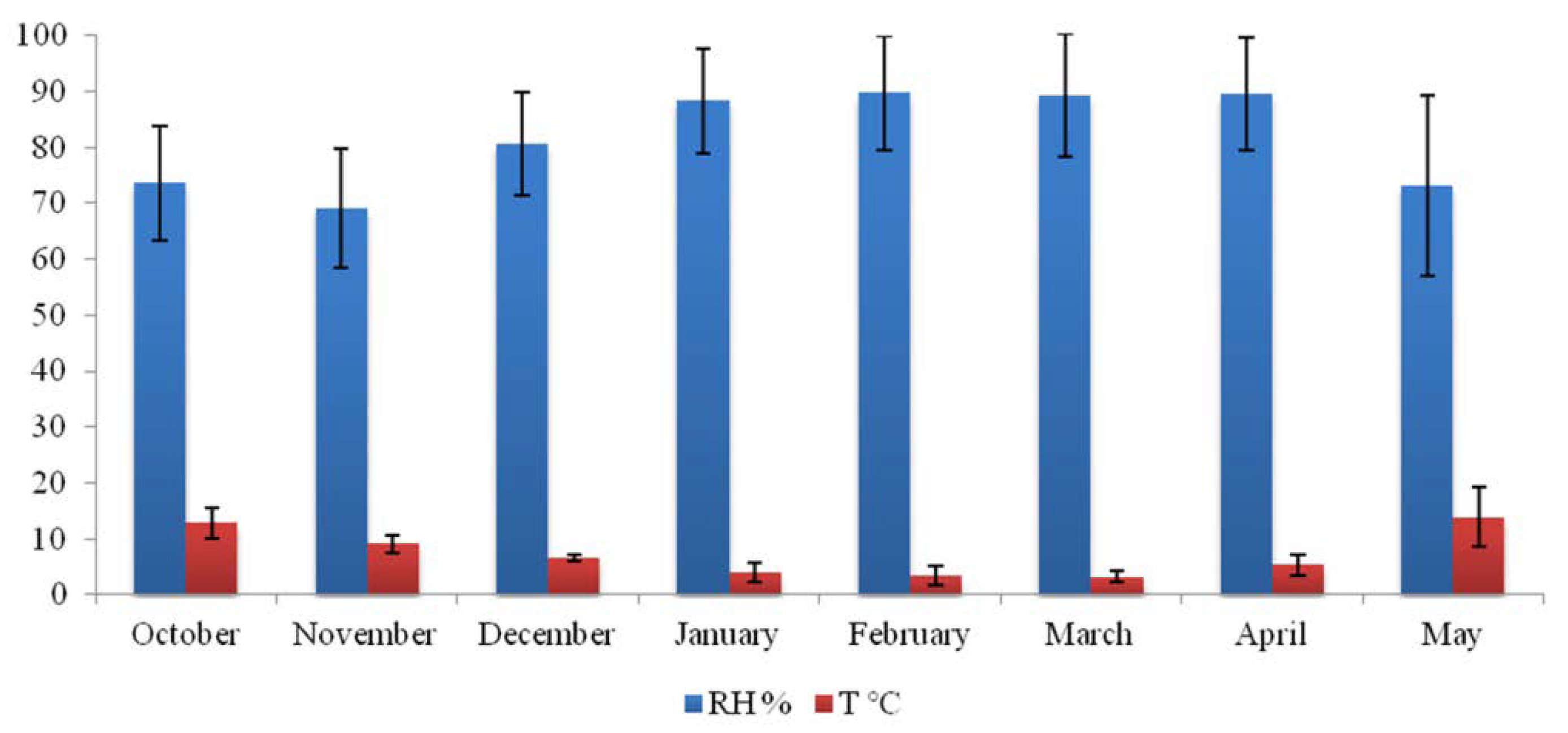
3.2. Indoor Church Microclimate and Material Surface Moisture Content
3.3. Surface Microbial Contamination
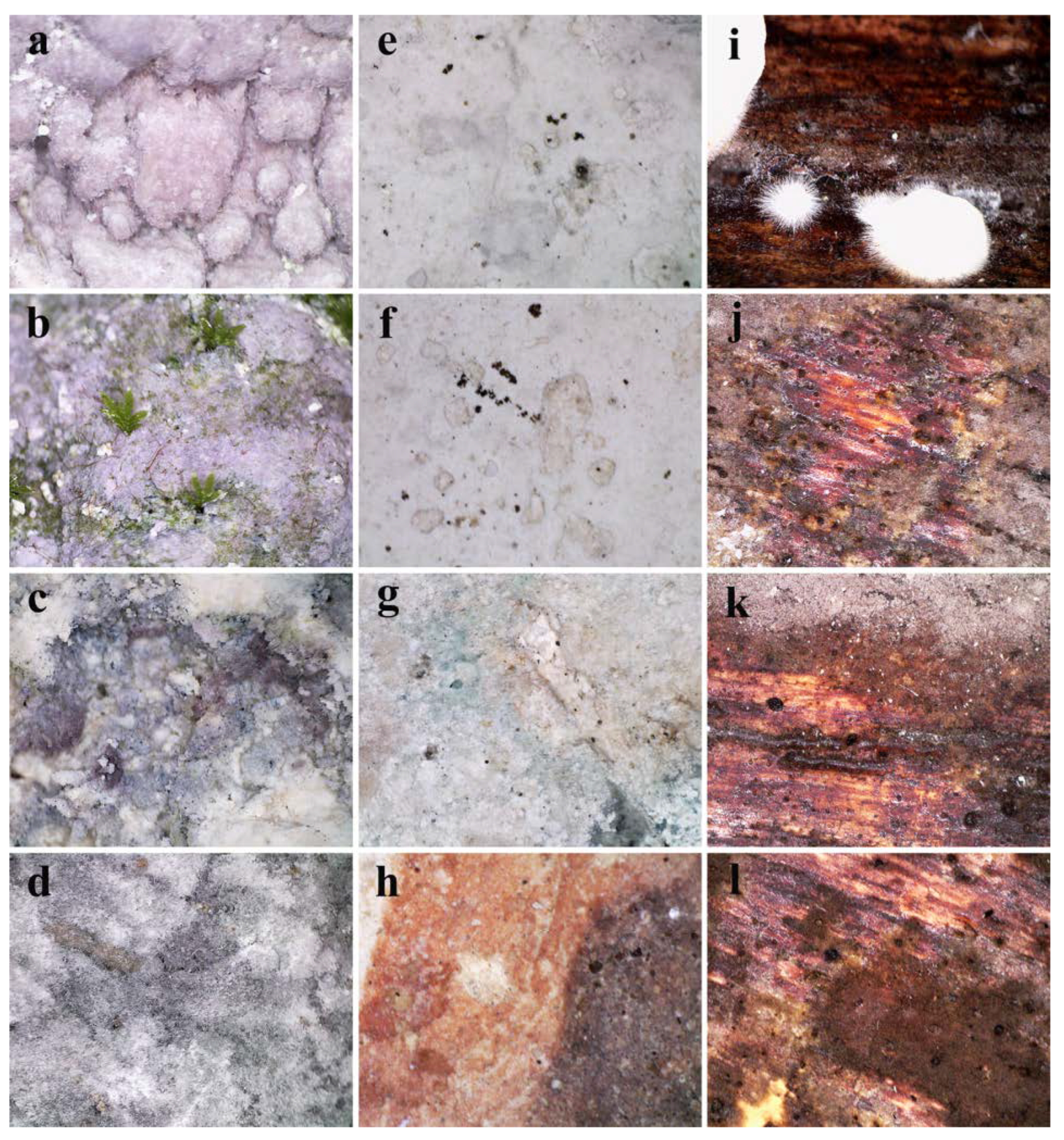
3.4. In Situ Assessment of Micro-Impairments and Surface Microbial Growth
3.5. Fungal Proliferation on Fresco and Iconostasis Surfaces
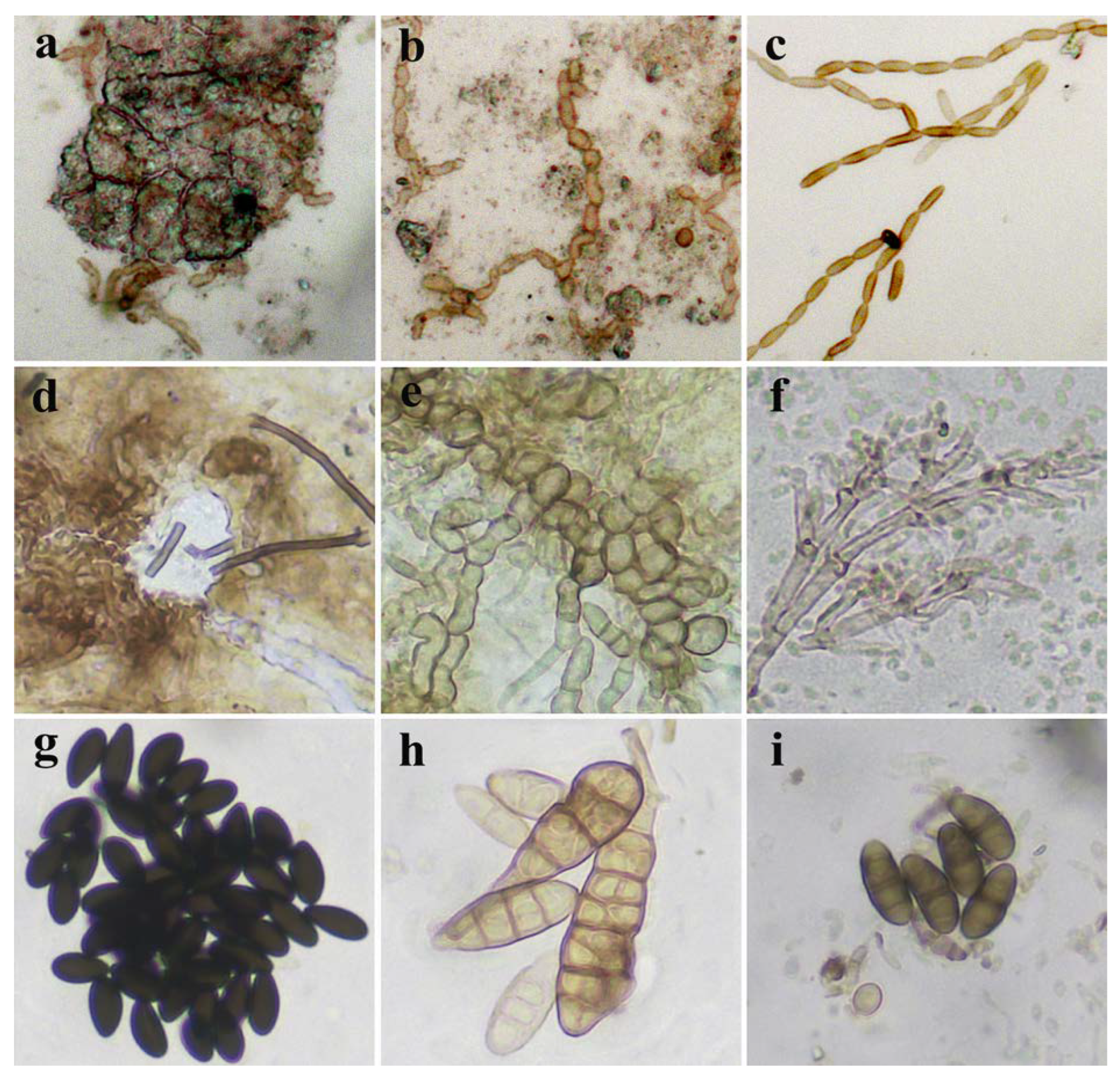
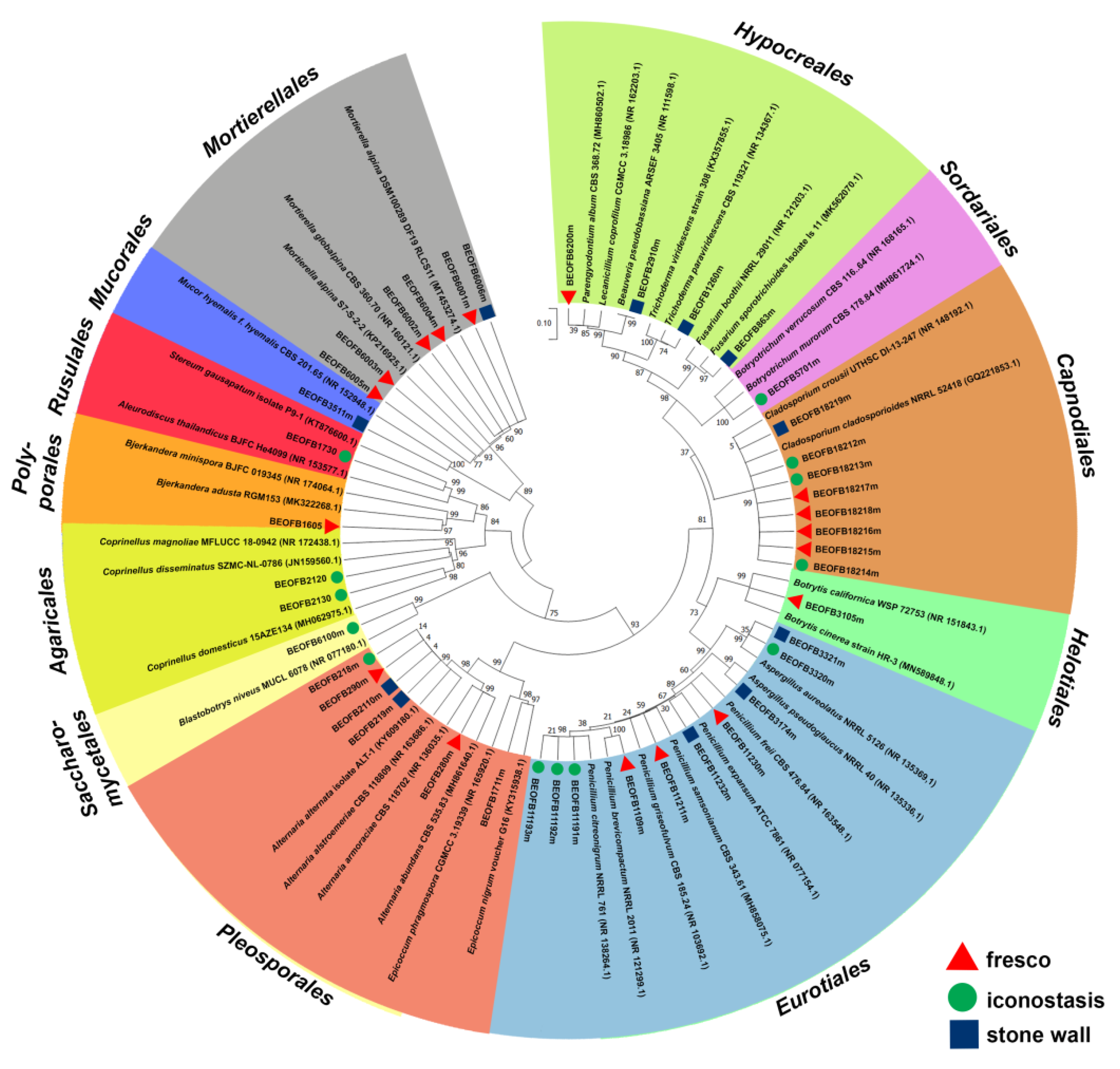
3.6. Culturable Mycobiome
3.7. Biodegradative Profile of Isolated Fungi
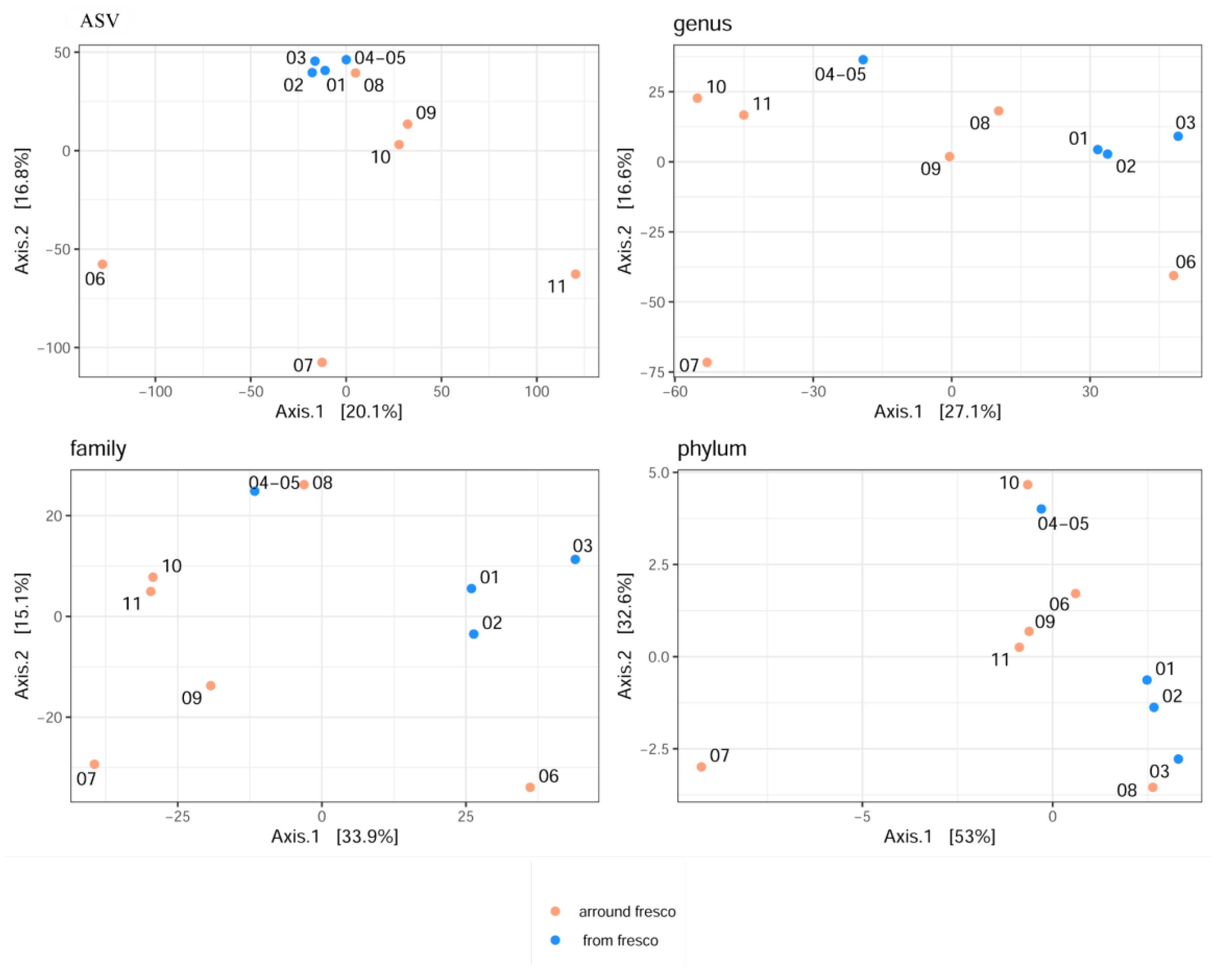
3.8. Diversity of Microbial Communities
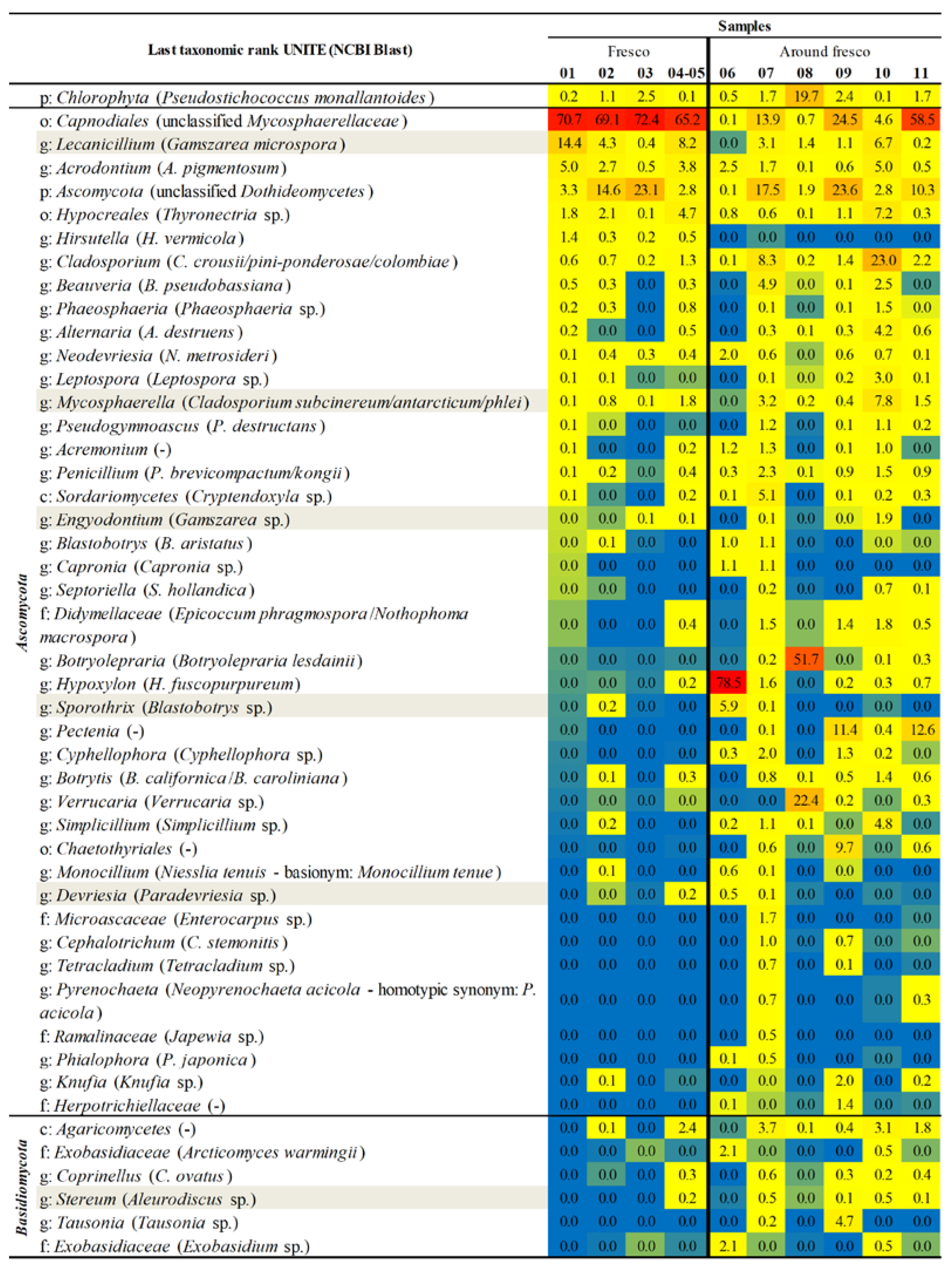
3.9. Total Microbial Composition
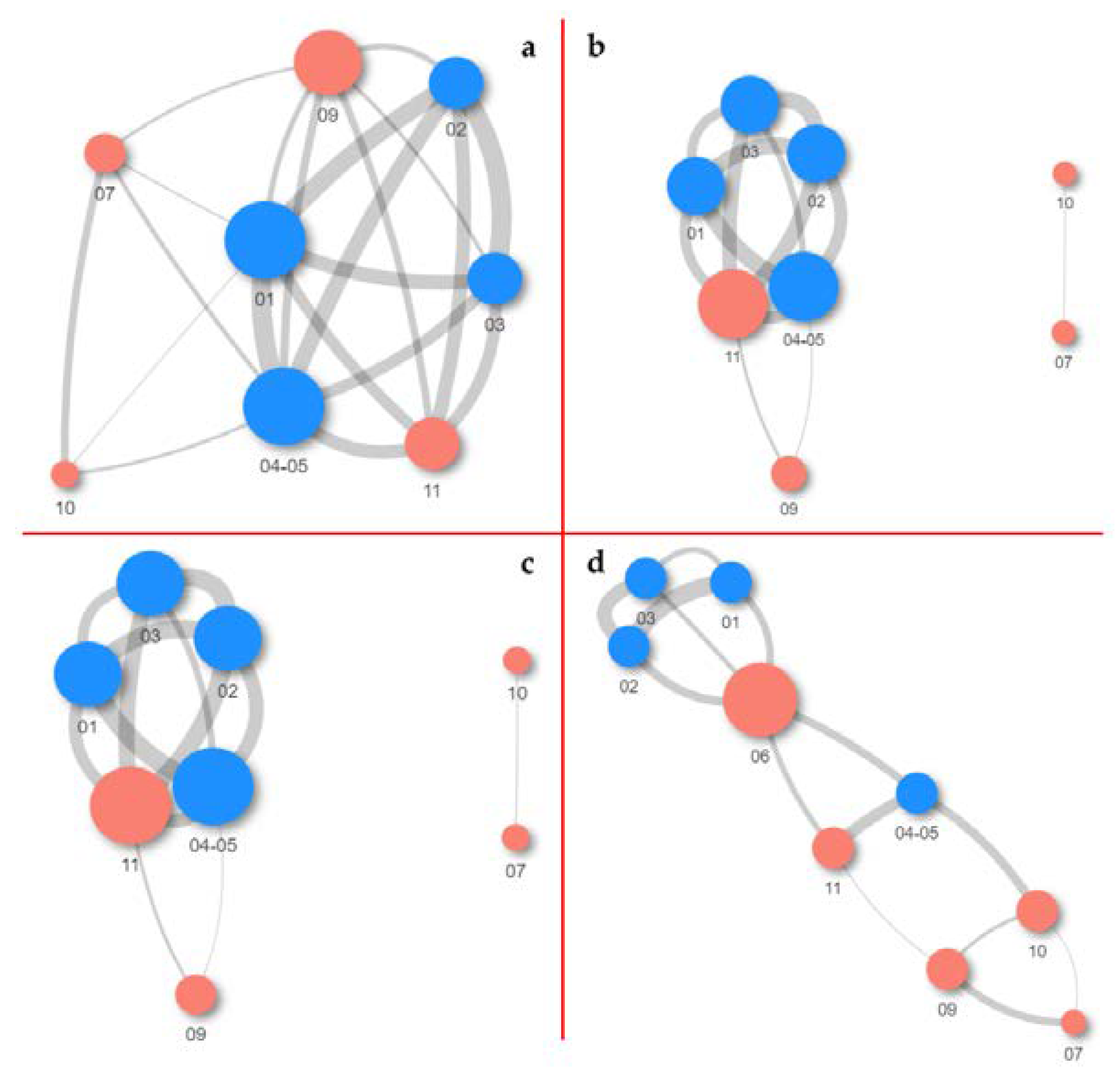
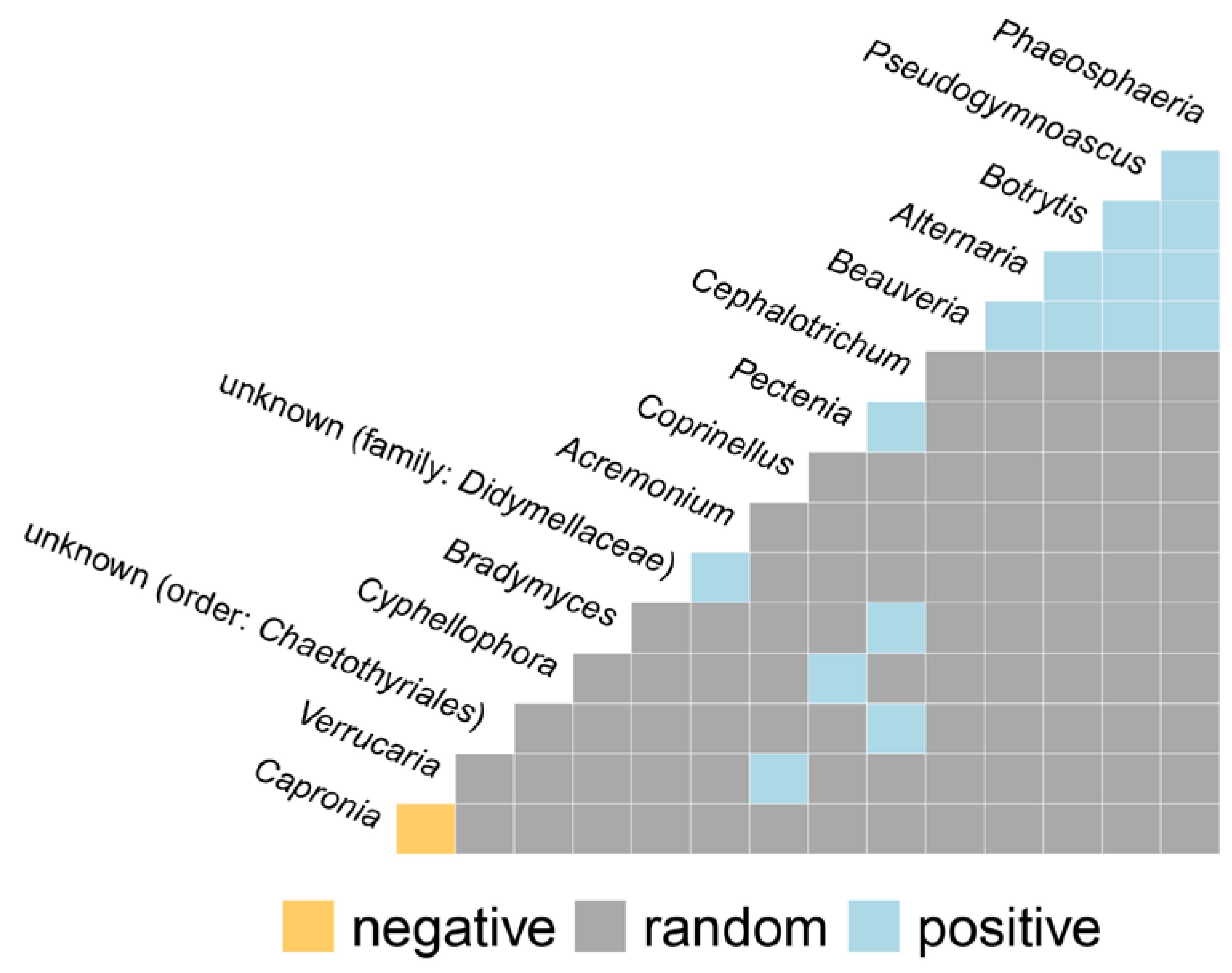
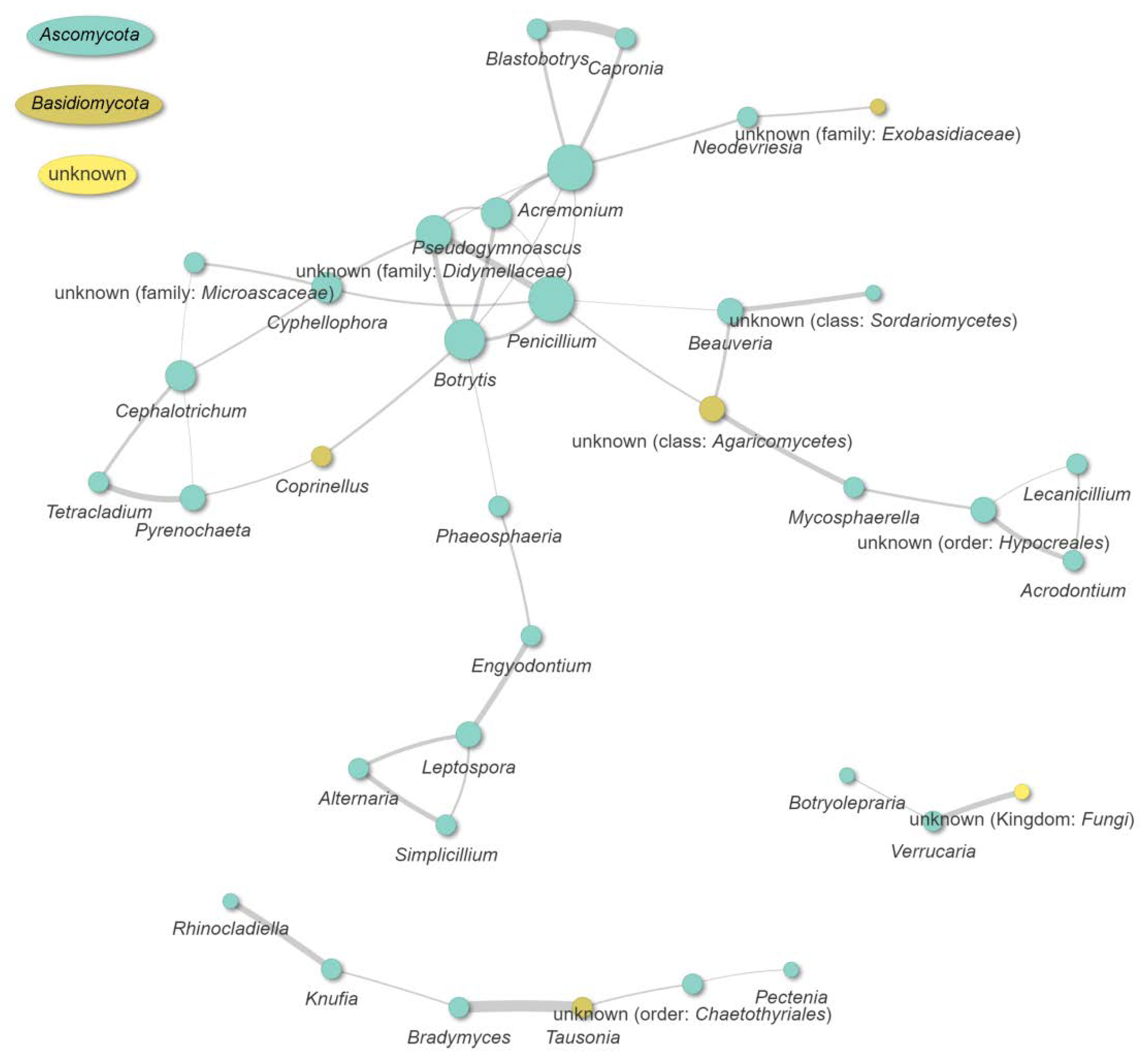
3.10. Co-Occurrence and Relative Abundance Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, E.L.S.; Resende-Stoianoff, M.A.A.; Lopes Ferreira, R. Mycological study for a management plan of a neotropical show cave (Brazil). Int. J. Speleol. 2013, 42, 267–277. [Google Scholar] [CrossRef]
- Gomoiu, I.; Mohanu, D.; Mohanu, I. Cercetări interdisciplinare privind conservarea picturilor murale din biserica rupestră de la Corbii de Piatră. Caiete ARA 2010, 1, 87–102. [Google Scholar] [CrossRef]
- Municchia, A.C.; Bartoli, F.; Taniguchi, Y.; Giordani, P.; Caneva, G. Evaluation of the biodeterioration activity of lichens in the Cave Church of Üzümlü (Cappadocia, Turkey). Int. Biodeterior. Biodegrad. 2018, 127, 160–169. [Google Scholar] [CrossRef]
- Caneva, G.; Bartoli, F.; Fontani, M.; Mazzeschi, D.; Visca, P. Changes in biodeterioration patterns of mural paintings: Multi-temporal mapping for a preventive conservation strategy in the Crypt of the Original Sin (Matera, Italy). J. Cult. Herit. 2019, 40, 59–68. [Google Scholar] [CrossRef]
- Garavelli, A.; Andriani, G.F.; Fioretti, G.; Iurilli, V.; Marsico, A.; Pinto, D. The “Sant’Angelo in Criptis” cave church in Santeramo in Colle (Apulia, South Italy): A multidisciplinary study for the evaluation of conservation state and stability assessment. Geosciences 2021, 11, 382. [Google Scholar] [CrossRef]
- Albertano, P. Cyanobacterial biofilms in monuments and caves. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Amsterda, The Netherlands, 2012; pp. 317–343. [Google Scholar]
- Sterflinger, K.; Piñar, G. Microbial deterioration of cultural heritage and works of art—Tilting at windmills? Appl. Microbiol. Biotechnol. 2013, 97, 9637–9646. [Google Scholar] [CrossRef] [PubMed]
- Dimkić, I.; Fira, D.; Janakiev, T.; Kabić, J.; Stupar, M.; Nenadić, M.; Unković, N.; Ljaljević Grbić, M. The microbiome of bat guano: For what is this knowledge important? Appl. Microbiol. Biotechnol. 2021, 5, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Paiva, D.S.; Fernandes, L.; Trovão, J.; Mesquita, N.; Tiago, I.; Portugal, A. Uncovering the Fungal Diversity Colonizing Limestone Walls of a Forgotten Monument in the Central Region of Portugal by High-Throughput Sequencing and Culture-Based Methods. Appl. Sci. 2022, 12, 10650. [Google Scholar] [CrossRef]
- Nováková, A.; Hubka, V.; Valinová, Š.; Kolařík, M.; Hillebrand-Voiculescu, A.M. Cultivable microscopic fungi from an underground chemosynthesis-based ecosystem: A preliminary study. Folia Microbiol. 2018, 63, 43–55. [Google Scholar] [CrossRef]
- Trovão, J.; Gil, F.; Catarino, L.; Soares, F.; Tiago, I.; Portugal, A. Analysis of fungal deterioration phenomena in the first Portuguese King tomb using a multi-analytical approach. Int. Biodeter. Biodegr 2020, 149, 104933. [Google Scholar] [CrossRef]
- Wang, W.; Ma, X.; Ma, Y.; Mao, L.; Wu, F.; Ma, X.; An, L.; Feng, H. Molecular characterization of airborne fungi in caves of the Mogao Grottoes, Dunhuang, China. Int. Biodeterior. Biodegrad. 2011, 65, 726–731. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi: Their role in deterioration of cultural heritage. Fungal Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Rosado, T.; Mirão, J.; Candeias, A.; Caldeira, A.T. Microbial communities analysis assessed by pyrosequencing—A new approach applied to conservation state studies of mural paintings. Anal. Bioanal. Chem. 2014, 406, 887–895. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, H.; Du, Y.; Tian, T.; Xiang, T.; Liu, X.; Wu, F.; An, L.; Wang, W.; Gu, J.; et al. The community distribution of bacteria and fungi on ancient wall paintings of the Mogao Grottoes. Sci. Rep. 2015, 5, 7752. [Google Scholar] [CrossRef] [PubMed]
- Unković, N.; Dimkić, I.; Stupar, M.; Stanković, S.; Vukojević, J.; Ljaljević Grbić, M. Biodegradative potential of fungal isolates from sacral ambient: In vitro study as risk assessment implication for the conservation of wall paintings. PLoS ONE 2018, 13, e0190922. [Google Scholar] [CrossRef]
- Savković, Ž.; Stupar, M.; Unković, N.; Knežević, A.; Vukojević, J.; Ljaljević Grbić, M. Fungal Deterioration of Cultural Heritage Objects. In Biodegradation Technology of Organic and Inorganic Pollutants; Mendes, K.F., de Sousa, R.N., Mielke, K.C., Eds.; IntechOpen: London, UK, 2021; pp. 267–288. [Google Scholar]
- Suphaphimol, N.; Suwannarach, N.; Purahong, W.; Jaikang, C.; Pengpat, K.; Semakul, N.; Yimklan, S.; Jongjitngam, S.; Jindasu, S.; Thiangtham, S.; et al. Identification of microorganisms dwelling on the 19th Century Lanna mural paintings from Northern Thailand using culture-dependent and -independent approaches. Biology 2022, 11, 228. [Google Scholar] [CrossRef] [PubMed]
- Popović, S.; Subakov-Simić, G.; Stupar, M.; Unković, N.; Krunić, O.; Savić, N.; Ljaljević Grbić, M. Cave biofilms: Characterization of phototrophic cyanobacteria and algae and chemotrophic fungi from three caves in Serbia. J. Cave Karst Stud. 2017, 79, 10–23. [Google Scholar] [CrossRef]
- Rakocija, M. Manastiri i Crkve Južne i Istočne Srbije; Zavod za Zaštitu Spomenika Kulture Niš: Niš, Srbija, 2013. [Google Scholar]
- Unković, N.; Ljaljević Grbić, M.; Stupar, M.; Vukojević, J.; Subakov-Simić, G.; Jelikić, A.; Stanojević, D. ATP bioluminescence method: Tool for rapid screening of organic and microbial contaminants on deteriorated mural paintings. Nat. Prod. Res. 2019, 33, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Urzì, C.; De Leo, F. Sampling with adhesive tape strips: An easy and rapid method to monitor microbial colonization on monument surfaces. J. Microbiol. Methods 2001, 44, 1–11. [Google Scholar] [CrossRef]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Sampson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; Westerdijk Fungal Biodiversity Institute: Utrecht, Netherlands, 2019. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Savković, Ž.; Stupar, M.; Unković, N.; Ivanović, Ž.; Blagojević, J.; Popović, S.; Vukojević, J.; Ljaljević Grbić, M. Diversity and seasonal dynamics of culturable airborne fungi in a cultural heritage conservation facility. Int Biodeterior Biodegrad 2021, 157, 105163. [Google Scholar] [CrossRef]
- Albertano, P.; Urzì, C. Structural interactions among epilithic cyanobacteria and heterotrophic microorganisms in Roman hypogea. Microb. Ecol. 1999, 38, 244–252. [Google Scholar] [CrossRef]
- Savković, Ž.; Stupar, M.; Unković, N.; Ivanović, Ž.; Blagojević, J.; Vukojević, J.; Ljaljević Grbić, M. In vitro biodegradation potential of airborne Aspergilli and Penicillia. Sci. Nat. 2019, 106, 8. [Google Scholar] [CrossRef]
- Borrego, S.; Guiamet, P.; de Saravia, S.G.; Batistini, P.; Garcia, M.; Lavin, P.; Perdomo, I. The quality of air at archives and the biodeterioration of photographs. Int. Biodeterior. Biodegrad. 2010, 64, 139–145. [Google Scholar] [CrossRef]
- Saran, S.; Isar, J.; Saxena, R.K. A modified method for the detection of microbial proteases on agar plates using tannic acid. J. Biochem. Biophys. Methods 2007, 70, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Pangallo, D.; Kraková, L.; Chovanová, K.; Šimonovičová, A.; De Leo, F.; Urzì, C. Analysis and comparison of the microflora isolated from fresco surface and from surrounding air environment through molecular and biodegradative assays. World J. Microbiol. Biotechnol. 2012, 28, 2015–2027. [Google Scholar] [CrossRef]
- Teather, R.M.; Wood, P.J. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ. Microbiol. 1982, 43, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.L.; Zhang, Z.; Wu, M.; Wu, Y.; Feng, J.X. Isolation, screening, and identification of cellulolytic bacteria from natural reserves in the subtropical region of China and optimization of cellulase production by Paenibacillus terrae ME27-1. BioMed Res. Int. 2014, 2014, 512497. [Google Scholar] [CrossRef]
- Kuwahara, M.; Glenn, J.K.; Morgan, M.A.; Gold, M.H. Separation and characterization of two extracellular H2O2-dependent oxidases from ligninolytic cultures of Phanerochaete chrysosporium. FEBS Lett. 1984, 169, 247–250. [Google Scholar] [CrossRef]
- Casciello, C.; Tonin, F.; Berini, F.; Fasoli, E.; Marinelli, F.; Pollegioni, L.; Rosini, E. A valuable peroxidase activity from the novel species Nonomuraea gerenzanensis growing on alkali lignin. Biotechnol. Rep. 2017, 13, 9–57. [Google Scholar] [CrossRef]
- Senthivelan, T.; Kanagaraj, J.; Panda, R.C.; Narayani, T. Screening and production of a potential extracellular fungal laccase from Penicillium chrysogenum: Media optimization by response surface methodology (RSM) and central composite rotatable design (CCRD). Biotechnol. Rep. 2019, 23, e00344. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge–accurate paired shotgun read merging via overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Pavoine, S.; Dufour, A.B.; Chessel, D. From dissimilarities among species to dissimilarities among communities: A double principal coordinate analysis. J. Theor. Biol. 2004, 228, 523–537. [Google Scholar] [CrossRef]
- Wright, E.S. DECIPHER: Harnessing local sequence context to improve protein multiple sequence alignment. BMC Bioinform. 2015, 16, 322. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic species co-occurrence analysis in R. J. Stat. Softw. 2016, 69, 1–17. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Foster, Z.S.; Sharpton, T.J.; Grünwald, N.J. Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-tailed prior distributions for sequence count data: Removing the noise and preserving large differences. Bioinformatics 2019, 35, 2084–2092. [Google Scholar] [CrossRef]
- Zarzo, M.; Fernández-Navajas, A.; García-Diego, F.J. Long-term monitoring of fresco paintings in the Cathedral of Valencia (Spain) through humidity and temperature sensors in various locations for preventive conservation. Sensors 2011, 11, 8685–8710. [Google Scholar] [CrossRef]
- Florian, M.L.E. Fungal Facts: Solving Fungal Problems in Heritage Collections; Archetype Publications: London, UK, 2002. [Google Scholar]
- Chlebicki, A.; Zielenkiewicz, U.; Wilczek, A.M. Fungi are not involved in biofilm formation on rock wall in subterranean arsenic mine in Poland. Nova Hedwig. 2014, 99, 255–269. [Google Scholar] [CrossRef]
- Lupan, I.; Ianc, M.B.; Kelemen, B.S.; Carpa, R.; Rosca-Casian, O.; Chiriac, M.T.; Popescu, O. New and old microbial communities colonizing a seventeenth-century wooden church. Folia Microbiol. 2014, 59, 45–51. [Google Scholar] [CrossRef]
- Popović, S.; Subakov Simić, G.; Stupar, M.; Unković, N.; Predojević, D.; Jovanović, J.; Ljaljević Grbić, M. Cyanobacteria, algae and microfungi present in biofilm from Božana Cave (Serbia). Int. J. Speleol. 2015, 44, 141–149. [Google Scholar] [CrossRef][Green Version]
- Unković, N.; Erić, S.; Šarić, K.; Stupar, M.; Savković, Ž.; Stanković, S.; Stanojević, O.; Dimkić, I.; Vukojević, J.; Ljaljević Grbić, M. Biogenesis of secondary mycogenic minerals related to wall paintings deterioration process. Micron 2017, 100, 1–9. [Google Scholar] [CrossRef]
- Madariaga, J.M.; Costantini, I.; Castro, K. Analysis of the Degradation of Medieval Mural Paintings in the Open Air Abandoned Church of Ribera, North of Spain. In Raman Spectroscopy in Archeology and Art History; Vandenabeele, P., Edwards, H., Eds.; The Royal Society of Chemistry: London, UK, 2018; Volume 2, pp. 213–233. [Google Scholar]
- Ogórek, R. Fungal Communities on Rock Surfaces in Demänovská Ice Cave and Demänovská Cave of Liberty (Slovakia). Geomicrobiol. J. 2018, 35, 266–276. [Google Scholar] [CrossRef]
- Jurado, V.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain. Appl. Sci. 2021, 11, 8858. [Google Scholar] [CrossRef]
- Zucconi, L.; Canini, F.; Isola, D.; Caneva, G. Fungi affecting wall paintings of historical value: A worldwide meta-analysis of their detected diversity. Appl. Sci. 2022, 12, 2988. [Google Scholar] [CrossRef]
- Irbe, I.; Karadelev, M.; Andersone, I.; Andersons, B. Biodeterioration of external wooden structures of the Latvian cultural heritage. J. Cult. Herit. 2012, 13, S79–S84. [Google Scholar] [CrossRef]
- Kovachovska, L.; Rusevska, K.; Karadelev, M. Diversity of wood-inhabiting fungi (Basidiomycota) in Macedonian cultural heritage. Maced. J. Ecol. Environ. 2020, 22, 5–16. [Google Scholar]
- Sequeira, S.O.; De Carvalho, H.P.; Mesquita, N.; Portugal, A.; Macedo, M.F. Fungal stains on paper: Is what you see what you get? Conserv. Património 2019, 32, 18–27. [Google Scholar] [CrossRef]
- Trovão, J.; Tiago, I.; Catarino, L.; Gil, F.; Portugal, A. In vitro analyses of fungi and dolomitic limestone interactions: Bioreceptivity and biodeterioration assessment. Int. Biodeterior. Biodegrad. 2020, 155, 105107. [Google Scholar] [CrossRef]
- Leplat, J.; François, A.; Bousta, F. Parengyodontium album, a frequently reported fungal species in the cultural heritage environment. Fungal Biol. Rev. 2020, 34, 126–135. [Google Scholar] [CrossRef]
- Dakal, T.C.; Cameotra, S.S. Microbially induced deterioration of architectural heritages: Routes and mechanisms involved. Environ. Sci. Eur. 2012, 24, 36. [Google Scholar] [CrossRef]
- Pangallo, D.; Chovanova, K.; Šimonovičová, A.; Ferianc, P. Investigation of microbial community isolated from indoor artworks and air environment: Identification, biodegradative abilities, and DNA typing. Can. J. Microbiol. 2009, 55, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Macêdo, D.P.C.; Neves, R.P.; Souza-Motta, C.M.D.; Magalhães, O.M.C. Engyodontium album fungaemia: The first reported case. Braz. J. Microbiol. 2007, 38, 110–112. [Google Scholar] [CrossRef]
- Borrego, S.; Molina, A.; Santana, A. Fungi in archive repositories environments and the deterioration of the graphics documents. EC Microbiol. 2017, 11, 205–226. [Google Scholar]
- Ávila-Hernández, J.G.; Carrillo-Inungaray, M.L.; De, R.; La Cruz-Quiroz, J.E.; Paz, D.B.; Parra, R.; Aguilar, C.M.; del Campo, R. Beauveria bassiana secondary metabolites: A review inside their production systems, biosynthesis, and bioactivities. Mex. J. Biotechnol. 2020, 5, 1–33. [Google Scholar] [CrossRef]
- Boddy, L.; Rayner, A.D.M. Population structure, inter-mycelial interactions and infection biology of Stereum gausapatum. Trans. Br. Mycol. Soc. 1982, 78, 337–351. [Google Scholar] [CrossRef]
- Caldeira, A.T.; Schiavon, N.; Mauran, G.; Salvador, C.; Rosado, T.; Mirão, J.; Candeias, A. On the biodiversity and biodeteriogenic activity of microbial communities present in the hypogenic Environment of the Escoural Cave, Alentejo, Portugal. Coatings 2021, 11, 209. [Google Scholar] [CrossRef]
- Trovão, J.; Mesquita, N.; Paiva, D.S.; Paiva de Carvalho, H.; Avelar, L.; Portugal, A. Can arthropods act as vectors of fungal dispersion in heritage collections? A case study on the archive of the University of Coimbra, Portugal. Int. Biodeterior. Biodegrad. 2013, 79, 49–55. [Google Scholar] [CrossRef]
- Franco-Castillo, I.; Misra, A.; Laratte, S.; Gommeaux, M.; Perarnau, R.; Vaillant-Gaveau, N.; Pierlot, C.; Mitchell, S.G.; Streb, C.; Eyssautier-Chuine, S. New protective coatings against lampenflora growing in the Pommery Champagne cellar. Int. Biodeterior. Biodegrad. 2022, 173, 105459. [Google Scholar] [CrossRef]
- McIlroy de La Rosa, J.P.; Warke, P.A.; Smith, B.J. Microscale biopitting by the endolithic lichen Verrucaria baldensis and its proposed role in mesoscale solution basin development on limestone. Earth Surf. Process. Landf. 2012, 37, 374–384. [Google Scholar] [CrossRef]
- Santo, A.P.; Cuzman, O.A.; Petrocchi, D.; Pinna, D.; Salvatici, T.; Perito, B. Black on white: Microbial growth darkens the external marble of Florence cathedral. Appl. Sci. 2021, 11, 6163. [Google Scholar] [CrossRef]
- Tang, A.M.; Jeewon, R.; Hyde, K.D. A re-evaluation of the evolutionary relationships within the Xylariaceae based on ribosomal and protein-coding gene sequences. Fungal Divers. 2009, 34, 127–155. [Google Scholar]
- Caneva, G.; Nugari, M.P.; Nugari, M.P.; Salvadori, O. Plant Biology for Cultural Heritage: Biodeterioration and Conservation; Getty Publications: Los Angeles, CA, USA, 2008. [Google Scholar]
- Wang, M.M.; Shenoy, B.D.; Li, W.; Cai, L. Molecular phylogeny of Neodevriesia, with two new species and several new combinations. Mycologia 2017, 109, 965–974. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Point | Zone of Cleanliness | Material Moisture Content (%) |
|---|---|---|
| Sample 01—depiction of Jesus | 5.4—Danger zone | 2.2 |
| Sample 02—entire fresco except depiction of Jesus | 5.6—Danger zone | 3.0 |
| Sample 03—gray discoloration on fresco | 5.4—Danger zone | 1.6 |
| Samples 04–05—fresco surface damage | 5.3 – Danger zone | 1.7 |
| Sample 06—pinkish gray powdery surface of iconostasis | 6.4—Danger zone | 17.1 |
| Sample 07—blue-green deposits on stone wall under the fresco | 6.1—Danger zone | 20.1 |
| Sample 08—green patina on stone wall | 5.7—Danger zone | 23.6 |
| Sample 09—black deposits on stone wall | 5.5—Danger zone | 23.8 |
| Sample 10—white deposits on stone wall | 5.3—Danger zone | 22.4 |
| Sample 11—pink deposits on stone wall | 5.5—Danger zone | 22.0 |
| No. | Strain | Isolate | Origin | Biodegradation Activity |
|---|---|---|---|---|
| 1 | BEOFB218m | Alternaria alternata | Iconostasis | CA, HA, LA (Lac, LiP, MnP) |
| 2 | BEOFB3320m | Aspergillus aureolatus | LA (Lac, MnP), PP (#9b9400) | |
| 3 | BEOFB6100m | Blastobotrys niveus | - | |
| 4 | BEOFB5701m | Botryotrichum murorum | CA, HA, LA (Lac, LiP, MnP) | |
| 5 | BEOFB18212m | Cladosporium cladosporioides | - | |
| 6 | BEOFB2120 | Coprinellus disseminatus | AP (6.73), HA, LA (Lac, LiP, MnP) | |
| 7 | BEOFB11191m | Penicillium citreonigrum | LA (Lac, MnP) | |
| 8 | BEOFB1730 | Stereum gausapatum | CA, HA, LA (Lac, MnP), PP (#eaa221) | |
| 9 | BEOFB280m | Alternaria abundans | Fresco | - |
| 10 | BEOFB290m | Alternaria brassicae | - | |
| 11 | BEOFB1605 | Bjerkandera adusta | AP (6.65), PA, PP (#beb72e) | |
| 12 | BEOFB3105m | Botrytis cinerea | AP (4.80) | |
| 13 | BEOFB18216m | Cladosporium cladosporioides | AP (6.72), CD, PA | |
| 14 | BEOFB6001m | Mortierella alpina | - | |
| 15 | BEOFB6200m | Parengyodontium album | PA | |
| 16 | BEOFB1109m | Penicillium brevicompactum | - | |
| 17 | BEOFB11230m | Penicillium frei | AP (4.27), PA, PP (#8db600) | |
| 18 | BEOFB11211m | Penicillium samsonianum | AP (4.75), CD, PA | |
| 19 | BEOFB219m | Alternaria alternata | Stone | - |
| 20 | BEOFB3321m | Aspergillus aureolatus | PP (#9b9400) | |
| 21 | BEOFB3174m | Aspergillus pseudoglaucus | - | |
| 22 | BEOFB2910m | Beauveria pseudobassiana | PP (#e9e450) | |
| 23 | BEOFB18219m | Cladosporium cladosporioides | - | |
| 24 | BEOFB2130 | Coprinellus domesticus | - | |
| 25 | BEOFB1711m | Epicoccum nigrum | - | |
| 26 | BEOFB863m | Fusarium sporotrichioides | - | |
| 27 | BEOFB6006m | Mortierella alpina | AP (4.90) | |
| 28 | BEOFB3511m | Mucor hiemalis | - | |
| 29 | BEOFB11132m | Penicillium expansum | AP (6.72) | |
| 30 | BEOFB1260m | Trichoderma viridescens | - |
| Indices | Description | Mean | St. Dev. | Min. | Max. | Kruskal–Wallis Sign. (p < 0.05) |
|---|---|---|---|---|---|---|
| Phylum | ||||||
| OBS | Fresco | 3.00 | 0.00 | 3.00 | 3.00 | 0.224 |
| Around Fresco | 3.67 | 1.21 | 3.00 | 6.00 | ||
| Chao1 | Fresco | 3.00 | 0.00 | 3.00 | 3.00 | 0.224 |
| Around Fresco | 3.67 | 1.21 | 3.00 | 6.00 | ||
| ACE | Fresco | 3.00 | 0.00 | 3.00 | 3.00 | 0.224 |
| Around Fresco | 3.67 | 1.21 | 3.00 | 6.00 | ||
| Shannon | Fresco | 0.11 | 0.09 | 0.02 | 0.22 | 0.019 |
| Around Fresco | 0.34 | 0.13 | 0.16 | 0.52 | ||
| Gini–Simpson | Fresco | 0.05 | 0.04 | 0.01 | 0.11 | 0.019 |
| Around Fresco | 0.17 | 0.09 | 0.06 | 0.32 | ||
| Family | ||||||
| OBS | Fresco | 56.75 | 29.88 | 22.00 | 95.00 | 0.136 |
| Around Fresco | 109.50 | 43.16 | 45.00 | 153.00 | ||
| Chao1 | Fresco | 57.65 | 30.14 | 22.00 | 95.75 | 0.136 |
| Around Fresco | 110.77 | 44.09 | 45.20 | 157.09 | ||
| ACE | Fresco | 57.88 | 30.13 | 22.23 | 95.96 | 0.136 |
| Around Fresco | 111.45 | 43.77 | 46.08 | 157.69 | ||
| Shannon | Fresco | 1.17 | 0.33 | 0.78 | 1.58 | 0.088 |
| Around Fresco | 2.11 | 0.86 | 1.08 | 3.24 | ||
| Gini–Simpson | Fresco | 0.49 | 0.06 | 0.42 | 0.56 | 0.088 |
| Around Fresco | 0.72 | 0.21 | 0.38 | 0.92 | ||
| Genus | ||||||
| OBS | Fresco | 90.00 | 61.07 | 26.00 | 173.00 | 0.136 |
| Around Fresco | 181.17 | 91.37 | 60.00 | 276.00 | ||
| Chao1 | Fresco | 93.43 | 61.44 | 26.00 | 175.10 | 0.136 |
| Around Fresco | 184.92 | 91.94 | 62.50 | 285.05 | ||
| ACE | Fresco | 92.86 | 61.53 | 26.19 | 175.25 | 0.136 |
| Around Fresco | 185.07 | 90.79 | 64.38 | 283.09 | ||
| Shannon | Fresco | 1.22 | 0.37 | 0.78 | 1.69 | 0.088 |
| Around Fresco | 2.28 | 1.02 | 1.10 | 3.55 | ||
| Gini–Simpson | Fresco | 0.49 | 0.06 | 0.42 | 0.56 | 0.088 |
| Around Fresco | 0.73 | 0.21 | 0.38 | 0.93 | ||
| ASV | ||||||
| OBS | Fresco | 155.50 | 108.82 | 47.00 | 306.00 | 0.088 |
| Around Fresco | 340.00 | 169.06 | 141.00 | 526.00 | ||
| Chao1 | Fresco | 158.40 | 110.29 | 47.00 | 310.50 | 0.088 |
| Around Fresco | 349.57 | 171.66 | 141.86 | 551.60 | ||
| ACE | Fresco | 157.79 | 109.58 | 47.00 | 308.76 | 0.088 |
| Around Fresco | 348.55 | 171.91 | 142.27 | 551.26 | ||
| Shannon | Fresco | 2.08 | 0.40 | 1.58 | 2.54 | 0.201 |
| Around Fresco | 3.09 | 1.12 | 1.70 | 4.32 | ||
| Gini–Simpson | Fresco | 0.73 | 0.04 | 0.67 | 0.78 | 0.286 |
| Around Fresco | 0.85 | 0.14 | 0.64 | 0.97 | ||
| Taxon Name | baseMean | log2 FoldChange | lfcSE | Stat | p Value |
|---|---|---|---|---|---|
| Hirsutella | 711.943552 | −10.86822846 | 1.941390071 | −4.567978681 | 4.9245 × 10−6 |
| Pyrenochaeta | 37.8113517 | 21.52668696 | 2.644391638 | 7.384188741 | 1.53386 × 10−13 |
| Phialophora | 101.742202 | 22.76808504 | 2.77610541 | 7.481014576 | 7.3751 × 10−14 |
| Rhinocladiella | 166.739135 | 9.386290822 | 2.156576309 | 3.425007867 | 0.000614782 |
| Taxon Name | p adj | log2 FoldChange_apeglm | lfcSE_apeglm | p value_apeglm | p adj_apeglm |
| Hirsutella | 0.000428432 | −10.2402688 | 1.941237778 | 2.16628 × 10−8 | 1.69692 × 10−6 |
| Pyrenochaeta | 2.00168 × 10−11 | 0.46897268 | 1.186974431 | 3.93628 × 10−16 | 4.62513 × 10−14 |
| Phialophora | 1.9249 × 10−11 | 0.4200758 | 1.154011635 | 2.37508 × 10−16 | 4.62513 × 10−14 |
| Rhinocladiella | 0.040114501 | 10.2864502 | 4.084514356 | 1.34653 × 10−5 | 0.000791086 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ljaljević Grbić, M.; Dimkić, I.; Savković, Ž.; Stupar, M.; Knežević, A.; Jelikić, A.; Unković, N. Mycobiome Diversity of the Cave Church of Sts. Peter and Paul in Serbia—Risk Assessment Implication for the Conservation of Rare Cavern Habitat Housing a Peculiar Fresco Painting. J. Fungi 2022, 8, 1263. https://doi.org/10.3390/jof8121263
Ljaljević Grbić M, Dimkić I, Savković Ž, Stupar M, Knežević A, Jelikić A, Unković N. Mycobiome Diversity of the Cave Church of Sts. Peter and Paul in Serbia—Risk Assessment Implication for the Conservation of Rare Cavern Habitat Housing a Peculiar Fresco Painting. Journal of Fungi. 2022; 8(12):1263. https://doi.org/10.3390/jof8121263
Chicago/Turabian StyleLjaljević Grbić, Milica, Ivica Dimkić, Željko Savković, Miloš Stupar, Aleksandar Knežević, Aleksa Jelikić, and Nikola Unković. 2022. "Mycobiome Diversity of the Cave Church of Sts. Peter and Paul in Serbia—Risk Assessment Implication for the Conservation of Rare Cavern Habitat Housing a Peculiar Fresco Painting" Journal of Fungi 8, no. 12: 1263. https://doi.org/10.3390/jof8121263
APA StyleLjaljević Grbić, M., Dimkić, I., Savković, Ž., Stupar, M., Knežević, A., Jelikić, A., & Unković, N. (2022). Mycobiome Diversity of the Cave Church of Sts. Peter and Paul in Serbia—Risk Assessment Implication for the Conservation of Rare Cavern Habitat Housing a Peculiar Fresco Painting. Journal of Fungi, 8(12), 1263. https://doi.org/10.3390/jof8121263