
Effect of Shock Waves on the Growth of Aspergillus niger Conidia: Evaluation of Germination and Preliminary Study on Gene Expression

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
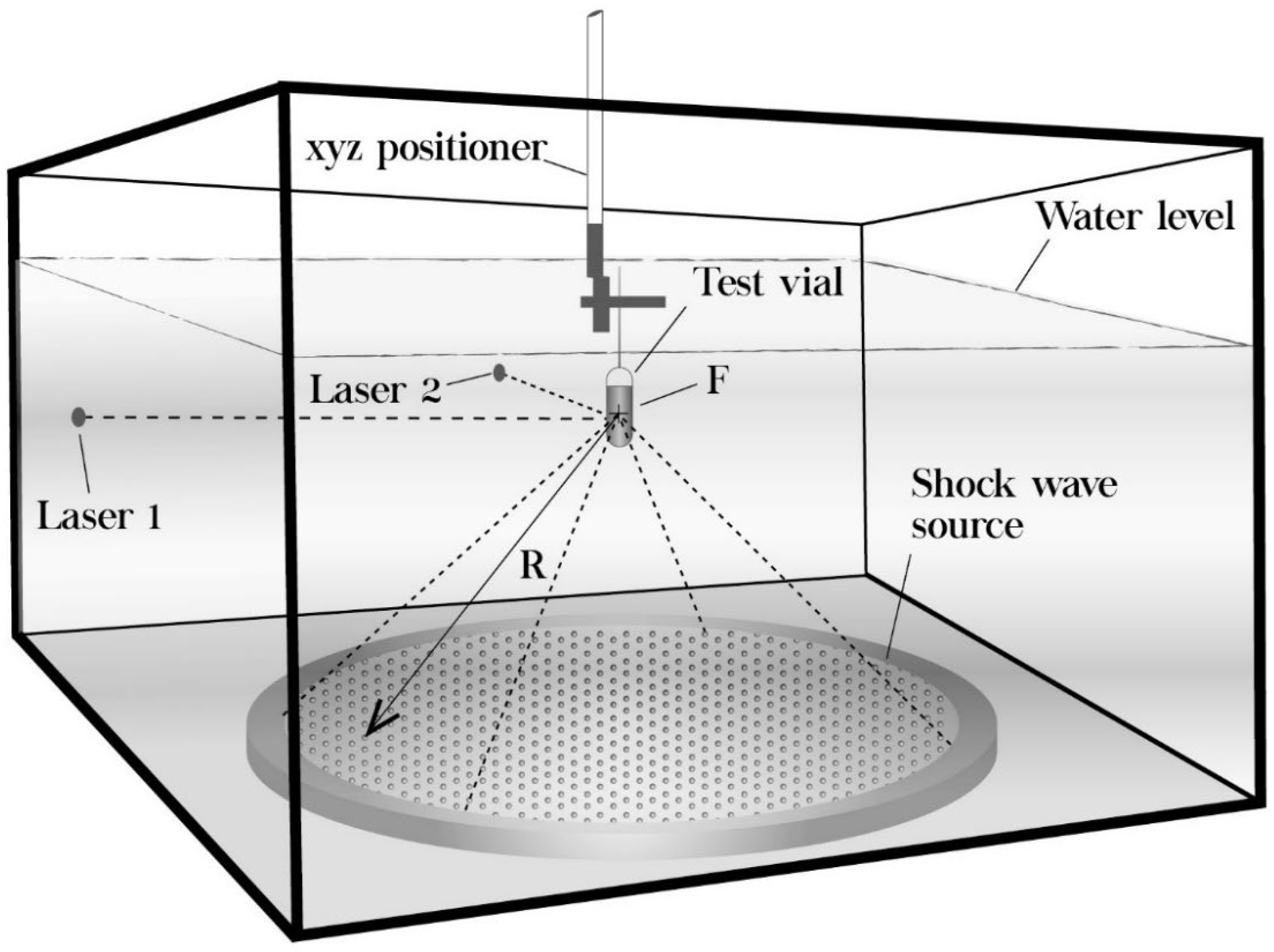
2.1. Experimental Setup
2.2. Pressure Measurements and High-Speed Imaging
2.3. Fungal Cell Culture and Sample Preparation
2.4. Shock Wave Application
2.5. Measurement of Cell Viability
2.6. Morphological Analysis and Its Quantification
2.7. Statistical Design and Data Analyses
2.8. Scanning Electron Microscopy
2.9. Cell Permeabilization Test
2.10. RNA Extraction
2.11. RT-PCR
3. Results
3.1. Pressure Measurements and High-Speed Imaging
3.2. Cell Viability
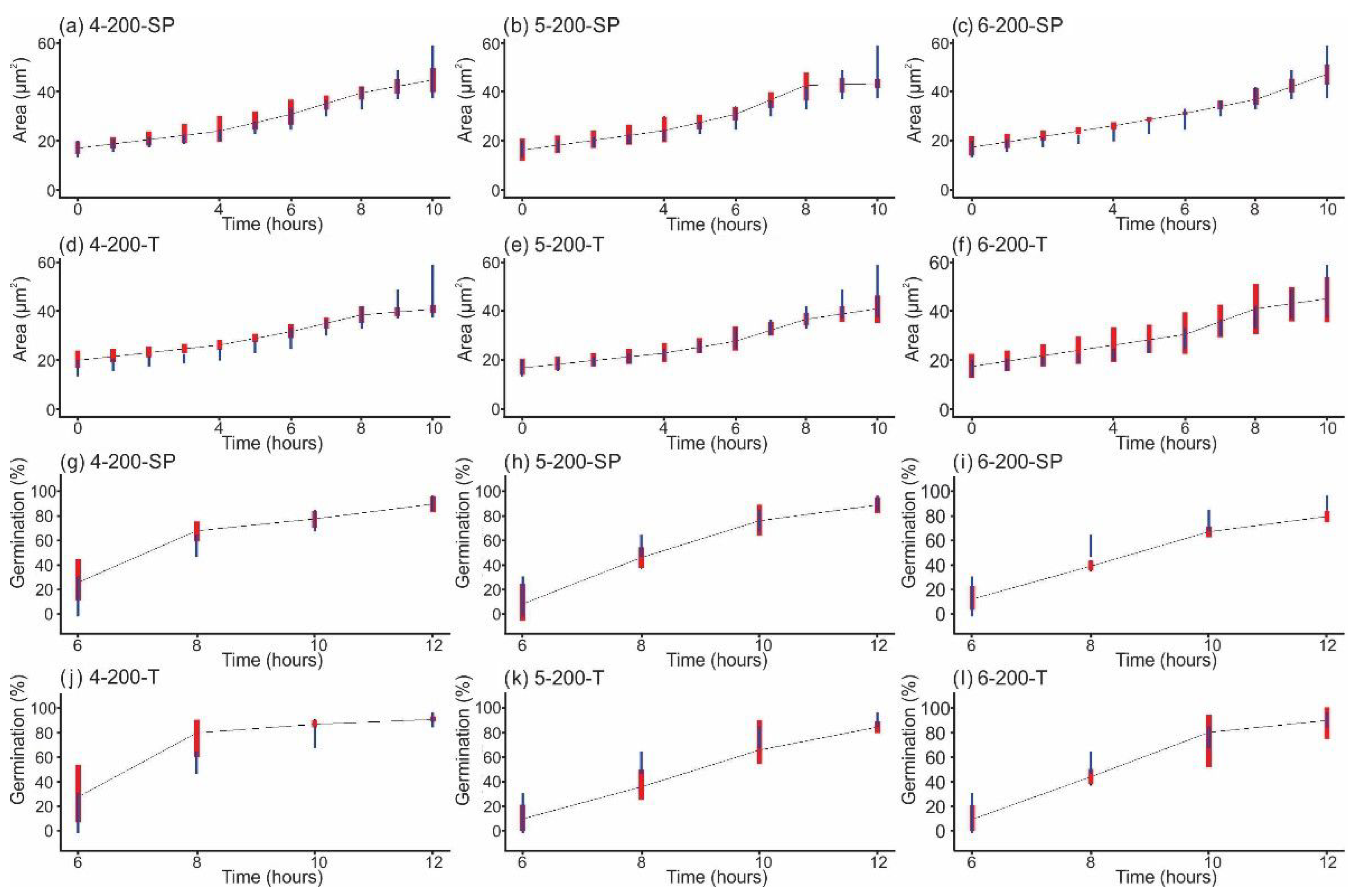
3.3. Growth and Germination of Conidia after Shock Wave Exposure
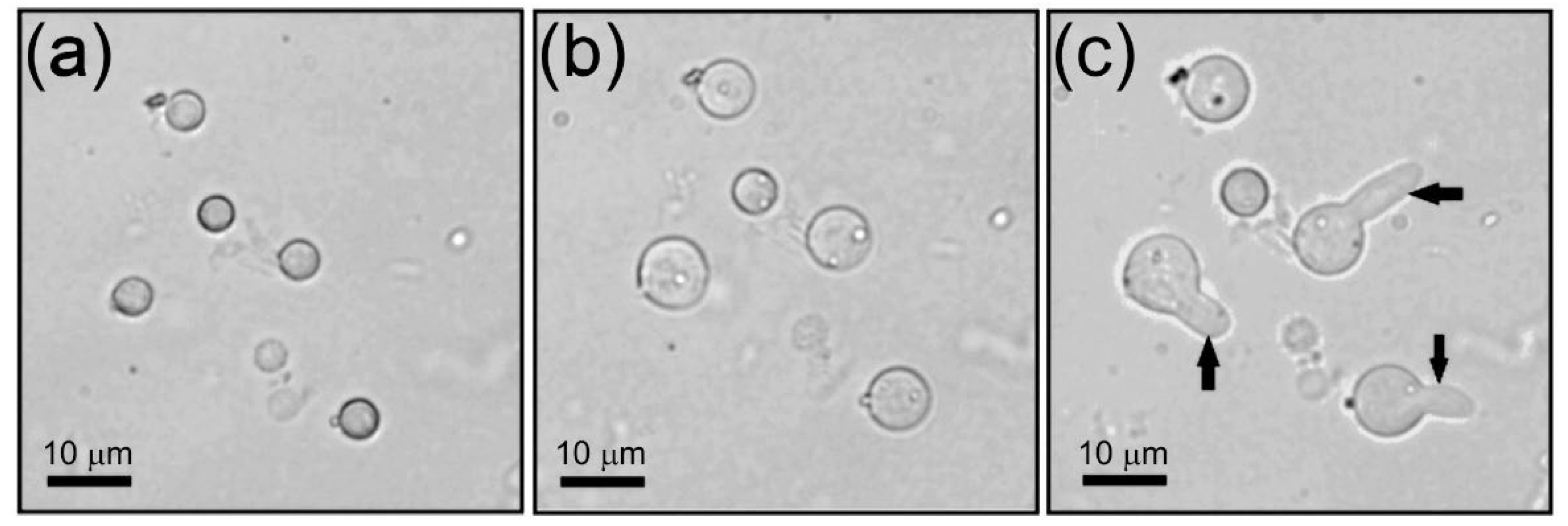
3.4. Surface Morphology of Aspergillus niger Conidia
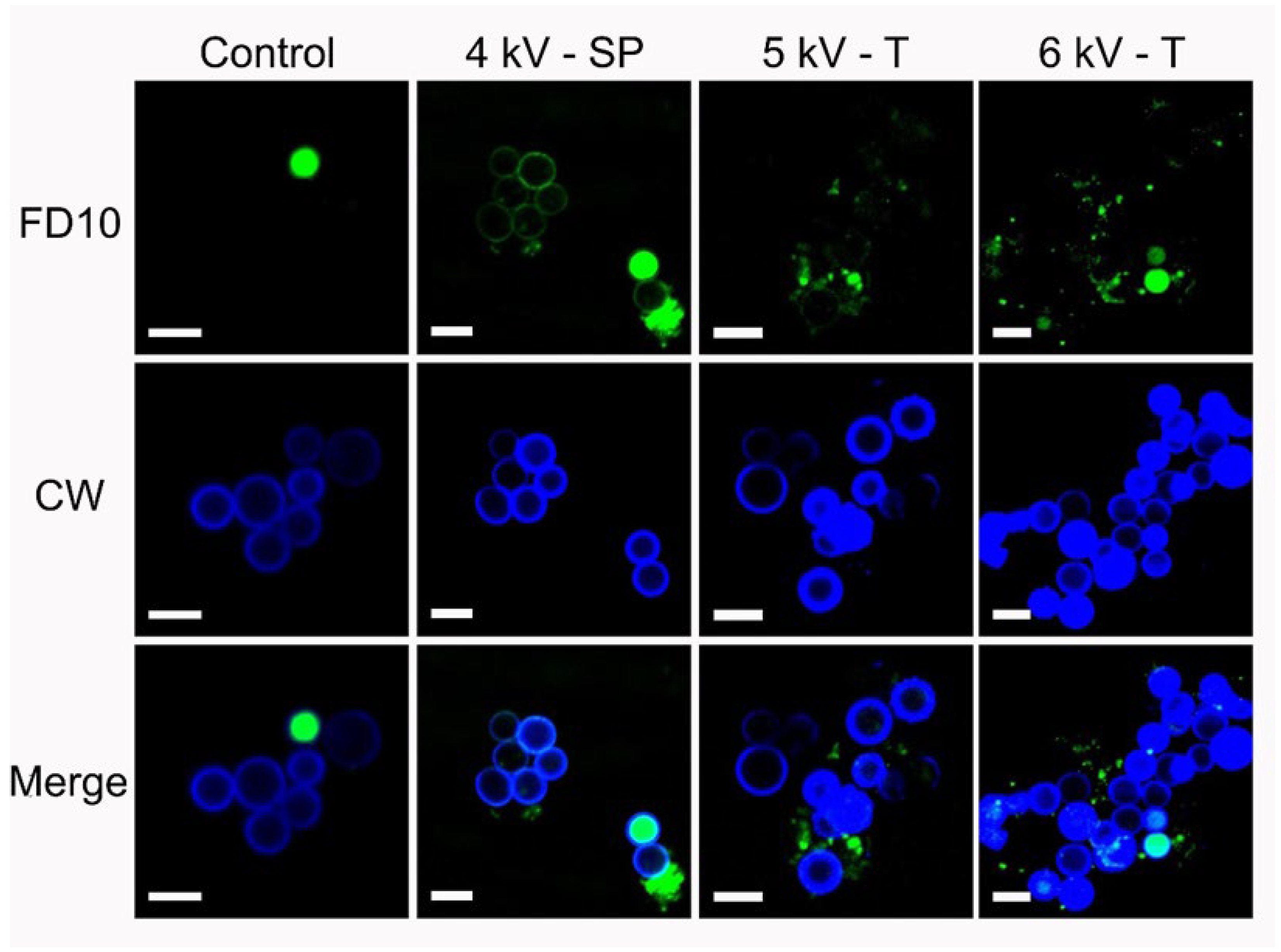
3.5. Cell Permeabilization
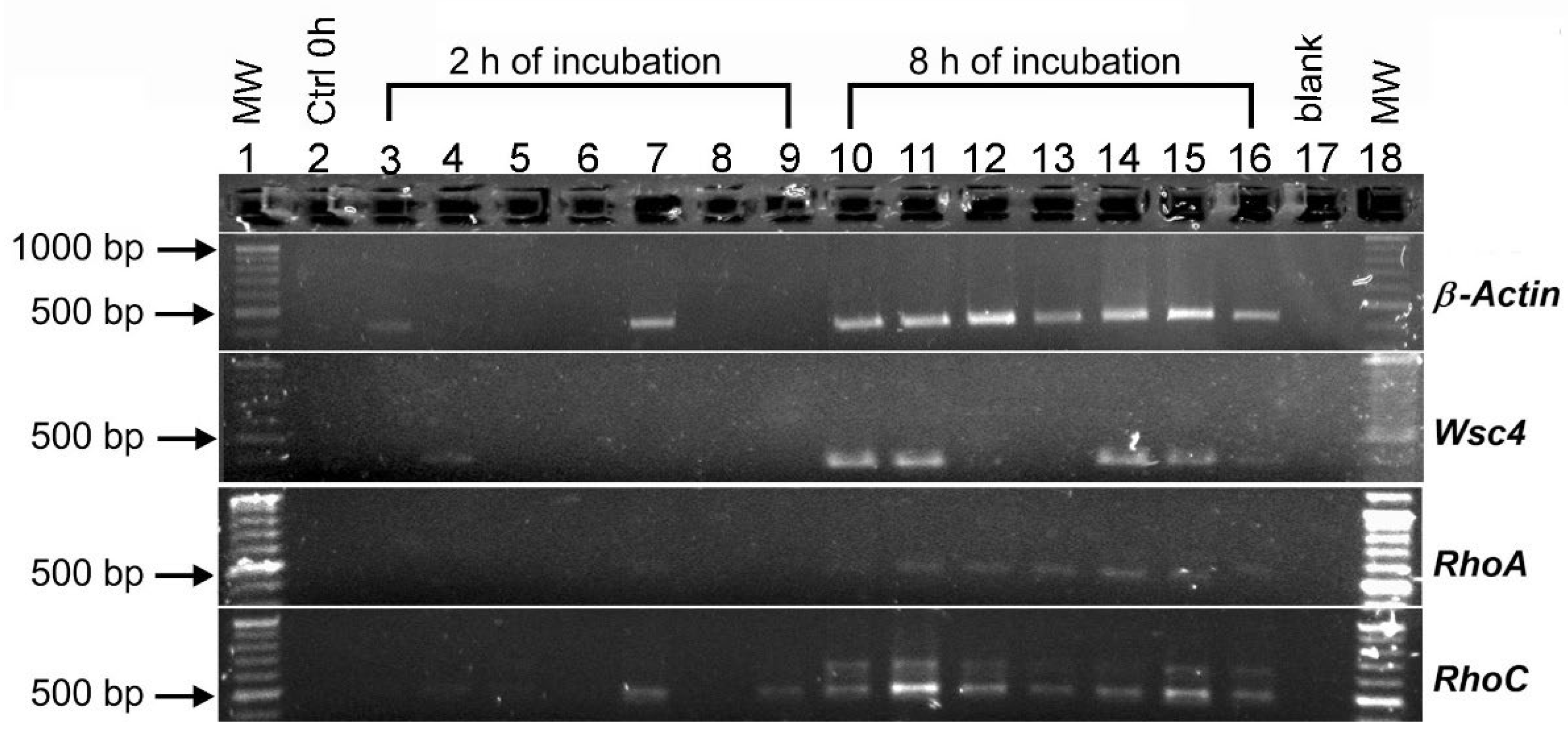
3.6. The Effect of Shock Wave Treatments and Incubation Time on the Expression of β-Actin, Wsc4, RhoA and RhoC Genes of Aspergillus niger
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CFU | Colony-forming units |
| CWI | Cell wall integrity |
| ER | Endoplasmic reticulum |
| Fps | Frames per second |
| FWHM | Full width at half maximum |
| MAPK | Mitogen-activated protein kinase |
| SP | Single pulse |
| T | Tandem |
References
- Yu, L.Y.; Li, L.X.; Yao, L.L.; Zheng, J.W.; Wang, B.; Pan, L. A special phenotype of aconidial Aspergillus niger SH2 and its mechanism of formation via CRISPRi. J. Fungi 2022, 8, 679. [Google Scholar] [CrossRef] [PubMed]
- Lubertozzi, D.; Keasling, J.D. Developing Aspergillus as a host for heterologous expression. Biotechnol. Adv. 2009, 27, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Fleissner, A.; Dersch, P. Expression and export: Recombinant protein production systems for Aspergillus. Appl. Microbiol. Biotechnol. 2010, 87, 1255–1270. [Google Scholar] [CrossRef] [PubMed]
- Ward, O.P. Production of recombinant proteins by filamentous fungi. Biotechnol. Adv. 2012, 30, 1119–1139. [Google Scholar] [CrossRef] [PubMed]
- Soccol, C.R.; Vandenberghe, L.P.S.; Rodrigues, C.; Pandey, A. New perspectives for citric acid production and application. Food Technol. Biotechnol. 2006, 44, 141–149. [Google Scholar]
- de Nobel, H.; Ruiz, C.; Martin, H.; Morris, W.; Brul, S.; Molina, M.; Klis, F.M. Cell wall perturbation in yeast results in dual phosphorylation of the Slt2/Mpk1 MAP kinase and in an Slt2-mediated increase in FKS2–lacZ expression, glucanase resistance and thermotolerance. Microbiology 2000, 146, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Mol, P.; Hellingwerf, K.; Brul, S. Dynamics of cell wall structure in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2002, 26, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Pel, H.J.; de Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; de Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.J.; Arentshorst, M.; Roos, E.D.; van den Hondel, C.A.M.J.J.; Meyer, V.; Ram, A.F.J. Functional Characterization of Rho GTPases in Aspergillus Niger Uncovers Conserved and Diverged Roles of Rho Proteins within Filamentous Fungi. Mol. Microbiol. 2011, 79, 1151–1167. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, M.R.; Krijgsheld, P.; Bleichrodt, R.; Menke, H.; Stam, H.; Stark, J.; Wösten, H.A.B.; Dijksterhuis, J. Germination of conidia of Aspergillus niger is accompanied by major changes in RNA profiles. Stud. Mycol. 2013, 74, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, M.R.; Krijgsheld, P.; Wyatt, T.T.; Golovina, E.A.; Menke, H.; Dekker, A.; Stark, J.; Stam, H.; Bleichrodt, R.; Wösten, H.A.B.; et al. The effect of natamycin on the transcriptome of conidia of Aspergillus niger. Stud. Mycol. 2013, 74, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Osherov, N.; May, G.S. The molecular mechanisms of conidial germination. FEMS Microbiol. Lett. 2001, 199, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Maubon, D.; Park, S.; Tanguy, M.; Huerre, M.; Schmitt, C.; Prévost, M.C.; Perlin, D.S.; Latgé, J.P.; Beauvais, A. AGS3, an α (1–3) glucan synthase gene family member of Aspergillus fumigatus, modulates mycelium growth in the lung of experimentally infected mice. Fungal Genet. Biol. 2006, 43, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Taheri-Talesh, N.; Horio, T.; Araujo-Bazán, L.; Dou, X.; Espeso, E.A.; Peñalva, M.A.; Osmani, S.A.; Oakley, B.R. The tip growth apparatus of Aspergillus nidulans. Mol. Biol. Cell 2008, 19, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.; Wu, B.; Ram, A.F.J. Aspergillus as a multi-purpose cell factory: Current status and perspectives. Biotechnol. Lett. 2011, 33, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Lanchun, S.; Bochu, W.; Zhiming, L.; Chuanren, D.; Chuanyun, D.; Sakanishi, A. The research into the influence of low-intensity ultrasonic on the growth of S. cerevisiaes. Colloid Surf. B 2003, 30, 43–49. [Google Scholar] [CrossRef]
- Zhao, L.; Cao, G.L.; Wang, A.J.; Ren, H.Y.; Dong, D.; Liu, Z.N.; Guan, X.Y.; Xu, C.J.; Ren, N.Q. Fungal pretreatment of cornstalk with Phanerochaete chrysosporium for enhancing enzymatic saccharification and hydrogen production. Bioresour. Technol. 2012, 114, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Khanal, S.N.; Anand, S.; Muthukumarappan, K. Evaluation of high-intensity ultrasonication for the inactivation of endospores of 3 bacillus species in nonfat milk. J. Dairy Sci. 2014, 97, 5952–5963. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Chen, S.; Dai, C.; Sun, L.; Sun, W.; Tang, Y.; Xiong, F.; He, R.; Ma, H. Effects of ultrasound on microbial growth and enzyme activity. Ultrason. Sonochem. 2017, 37, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.; Kiel, J.A.K.W.; Driessen, A.J.M.; Bovenberg, R.A.L.; Nygard, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synth. Biol. 2016, 5, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Lichius, A.; Ruiz, D.M.; Zeilinger, S. Genetic Transformation of Filamentous Fungi: Achievements and challenges. In Grand Challenges in Fungal Biotechnology. Grand Challenges in Biology and Biotechnology; Nevalainen, H., Ed.; Springer: Cham, Switzerland; Zug, Switzerland, 2020; pp. 123–164. [Google Scholar]
- Torres-Mendoza, D.; Ortega, H.E.; Cubilla-Rios, L. Patents on endophytic fungi related to secondary metabolites and biotransformation applications. J. Fungi 2020, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Magaña-Ortíz, D.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. Extracellular expression in Aspergillus niger of an antibody fused to Leishmania sp. antigens. Curr. Microbiol. 2018, 75, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Coconi-Linares, N.; Magaña-Ortíz, D.; Guzmán-Ortiz, D.A.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. High-yield production of manganese peroxidase, lignin peroxidase, and versatile peroxidase in Phanerochaete chrysosporium. Appl. Microbiol. Biotechnol. 2014, 98, 9283–9294. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.L.; Magaña-Ortíz, D.; Gómez-Lim, M.; Fernández, F.; Loske, A.M. Physical methods for genetic transformation of fungi and yeast. Phys. Life Rev. 2014, 11, 184–203. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Tovar, L.; Magaña-Ortíz, D.; Fernández, F.; Guzmán-Quesada, M.; Sandoval-Fernández, J.A.; Ortíz-Vázquez, E.; Loske, A.M.; Gómez-Lim, M.A. Efficient transformation of Mycosphaerella fijiensis by underwater shock waves. J. Microbiol. Methods 2015, 119, 98–105. [Google Scholar] [CrossRef]
- Rojas-Sánchez, U.; López-Calleja, A.C.; Millán-Chiu, B.E.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. Enhancing the yield of human erythropoietin in Aspergillus niger by introns and CRISPR-Cas9. Protein Exp. Purif. 2020, 168, 105570. [Google Scholar] [CrossRef]
- Gómez-Lim, M.A.; Ortíz, D.M.; Fernández, F.; Loske, A.M. Transformation of Fungi using Shock Waves. In Genetic Transformation Systems in Fungi; van den Berg, M.A., Maruthachalam, K., Eds.; Springer: New York, NY, USA, 2015; Volume 1, pp. 209–219. [Google Scholar]
- Loske, A.M.; Fernández, F.; Magaña-Ortíz, D.; Coconi-Linares, N.; Ortíz-Vázquez, E.; Gómez-Lim, M.A. Tandem shock waves to enhance genetic transformation of Aspergillus niger. Ultrasonics 2014, 54, 1656–1662. [Google Scholar] [CrossRef]
- Magaña-Ortíz, D.; Coconi-Linares, N.; Ortiz-Vazquez, E.; Fernández, F.; Loske, A.M.; Gómez-Lim, M.A. A novel and highly efficient method for genetic transformation of fungi employing shock waves. Fungal Genet. Biol. 2013, 56, 9–16. [Google Scholar] [CrossRef]
- Ohl, C.D.; Ikink, R. Shock-wave-induced jetting of micron-size bubble. Phys. Rev. Lett. 2003, 90, 214502. [Google Scholar] [CrossRef]
- Klaseboer, E.; Fong, S.W.; Turangan, C.K.; Khoo, B.C.; Szeri, A.J.; Calvisi, M.L.; Sankin, G.N.; Zhong, P. Interaction of lithotripter shockwaves with single inertial cavitation bubbles. J. Fluid Mech. 2007, 593, 33–56. [Google Scholar] [CrossRef]
- Brujan, E.A.; Ikeda, T.; Matsumoto, Y. On the pressure of cavitation bubbles. Exp. Therm. Fluid Sci. 2008, 32, 1188–1191. [Google Scholar] [CrossRef]
- Johnsen, E.; Colonius, T. Shock-induced collapse of a gas bubble in shock wave lithotripsy. J. Acoust. Soc. Am. 2008, 124, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Philip, B.; Levin, D.E. Wsc1 and Mid2 are cell surface sensors for cell wall integrity signaling that act through Rom2, a guanine nucleotide exchange factor for Rho1. Mol. Cell. Biol. 2001, 21, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Ohl, C.D.; Wolfrum, B. Detachment and sonoporation of adherent HeLa-cells by shock wave-induced cavitation. Biochim. Biophys. Acta 2003, 1624, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.R. Control of acoustic cavitation with application to lithotripsy. J. Acoust. Soc. Am. 1997, 102, 1250. [Google Scholar] [CrossRef]
- Lukes, P.; Fernández, F.; Gutiérrez-Aceves, J.; Fernández, E.; Alvarez, U.M.; Sunka, P.; Loske, A.M. Tandem shock waves in medicine and biology: A review of potential applications and successes. Shock Waves 2016, 26, 1–23. [Google Scholar] [CrossRef]
- López-Marín, L.M.; Millán-Chiu, B.E.; Castaño-González, K.; Aceves, C.; Fernández, F.; Varela-Echavarría, A.; Loske, A.M. Shock wave-induced damage and poration in eukaryotic cell membranes. J. Membr. Biol. 2017, 250, 41–52. [Google Scholar] [CrossRef]
- Kodama, T.; Takayama, K. Dynamic behavior of bubbles during extracorporeal shock-wave lithotripsy. Ultrasound Med. Biol. 1998, 24, 723–738. [Google Scholar] [CrossRef]
- Tzanakis, I.; Eskin, D.G.; Georgoulas, A.; Fytanidis, D.K. Incubation pit analysis and calculation of the hydrodynamic impact pressure from the implosion of an acoustic cavitation bubble. Ultrason. Sonochem. 2014, 21, 866–878. [Google Scholar] [CrossRef]
- Ramachandran, S.; Fontanille, P.; Pandey, A.; Larroche, C. Permeabilization and inhibition of the germination of spores of Aspergillus niger for gluconic acid production from glucose. Bioresour. Technol. 2008, 99, 4559–4565. [Google Scholar] [CrossRef]
- Gortikov, M.; Yakubovich, E.; Wang, Z.; López-Giráldez, F.; Tu, Y.; Townsend, J.P.; Yarden, O. Differential expression of cell wall remodeling genes is part of the dynamic phase-specific transcriptional program of conidial germination of Trichoderma asperelloides. J. Fungi 2022, 8, 854. [Google Scholar] [CrossRef] [PubMed]
- Baltussen, T.J.H.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular mechanisms of conidial germination in Aspergillus spp. Microbiol. Mol. Biol. Rev. 2020, 84, e00049-00019. [Google Scholar] [CrossRef] [PubMed]
- Verburg, K.; van Neer, J.; Duca, M.; de Cock, H. Novel treatment approach for Aspergilloses by targeting germination. J. Fungi 2022, 8, 758. [Google Scholar] [CrossRef] [PubMed]
- Schachtschabel, D.; Arentshorst, M.; Lagendijk, E.L.; Ram, A.F.J. Vacuolar H+-ATPase plays a key role in cell wall biosynthesis of Aspergillus niger. Fungal Genet. Biol. 2012, 49, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Rasmussen, C.; Roca, M.G.; Read, N.D. Hyphal homing, fusion and mycelial interconnectedness. Trends Microbiol. 2004, 12, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, T.; Simenel, C.; Dubreucq, G.; Adam, O.; Delepierre, M.; Lemoine, J.; Vorgias, C.E.; Diaquin, M.; Latgé, J.P. Molecular organization of the alkali-insoluble fraction of Aspergillus fumigatus cell wall. J. Biol. Chem. 2000, 275, 27594–27607. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 1–25. [Google Scholar] [CrossRef]
- Sánchez-Arreguin, J.A.; Ortiz-Castellanos, M.L.; Robledo-Briones, A.M.; León-Ramírez, C.G.; Martínez-Soto, D.; Ruiz-Herrera, J. Chitosan is necessary for the structure of the cell wall, and full virulence of Ustilago maydis. J. Fungi 2022, 8, 813. [Google Scholar] [CrossRef]
- Subroto, E.; van Neer, J.; Valdes, I.; de Cock, H. Growth of Aspergillus fumigatus in biofilms in comparison to Candida albicans. J. Fungi 2022, 8, 48. [Google Scholar] [CrossRef]
- Pogorelko, G.; Lionetti, V.; Bellincampi, D.; Zabotina, O. Cell wall integrity: Targeted post-synthetic modifications to reveal its role in plant growth and defense against pathogens. Plant Signal. Behav. 2013, 8, e25435. [Google Scholar] [CrossRef]
- Gomez-Gomez, A.; Brito-de la Fuente, E.; Gallegos, C.; Garcia-Perez, J.V.; Benedito, J. Combination of supercritical CO2 and high-power ultrasound for the inactivation of fungal and bacterial spores in lipid emulsions. Ultrason. Sonochem. 2012, 76, 105636. [Google Scholar] [CrossRef] [PubMed]
- Damveld, R.A.; Arentshorst, M.; Franken, A.; VanKuyk, P.A.; Klis, F.M.; Van Den Hondel, C.A.; Ram, A.F. The Aspergillus niger MADS-box transcription factor RlmA is required for cell wall reinforcement in response to cell wall stress. Mol. Microbiol. 2005, 58, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Millet, N.; Latgé, J.P.; Mouyna, I. Members of glycosyl-hydrolase family 17 of A. fumigatus differentially affect morphogenesis. J. Fungi 2018, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.; Delgado-Gonzalez, E.; López-Marín, L.M.; Millán-Chiu, B.E.; Fernández, F.; Rodriguez-Castelan, J.; Muñoz-Torres, C.; Carrasco, G.; Anguiano, B.; Loske, A.M.; et al. Shock wave application increases the antineoplastic effect of molecular iodine supplement in breast cancer xenografts. Ultrasound Med. Biol. 2020, 46, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Kaminskyj, S.G. Fundamentals of growth, storage, genetics and microscopy of Aspergillus nidulans. Fungal Genet. Rep. 2001, 48, 25–31. [Google Scholar] [CrossRef]
- Efron, B.; Hastie, T. Computer Age Statistical Inference: Algorithms, Evidence and Data Science; Cambridge University Press: Cambridge, UK, 2016; Available online: https://web.stanford.edu/~hastie/CASI_files/PDF/casi.pdf (accessed on 14 June 2022).
- Kohl, M. Introduction to Statistical Data Analysis with R, 1st ed.; Bookboon: London, UK, 2015; ISBN 978-87-403-1123-5. [Google Scholar]
- Yun, J.; Lee, H.; Ko, H.J.; Woo, E.R.; Lee, D.G. Fungicidal effect of isoquercitrin via inducing membrane disturbance. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 695–701. [Google Scholar] [CrossRef]
- Arora, M.; Ohl, C.D.; Mørch, K.A. Cavitation inception on microparticles: A self-propelled particle accelerator. Phys. Rev. Lett. 2004, 92, 174501. [Google Scholar] [CrossRef]
- Cleveland, R.O.; McAteer, J.A. The Physics of Shock Wave Lithotripsy. In Smith’s Textbook on Endourology, 3rd ed.; Arthur, D., Smith, M.D., Gopal, H., Badlani, M.D., Glenn, M., Preminger, M.D., Louis, R., Kavoussi, M.D., Eds.; Wiley Online Library: Washington, DC, USA, 2012; Volume 1, pp. 529–558. [Google Scholar]
- Loske, A.M. Medical and Biomedical Applications of Shock Waves, 1st ed.; Springer: Cham, Switzerland, 2017; pp. 19–291. [Google Scholar]
- Kodama, T.; Hamblin, M.R.; Doukas, A.G. Cytoplasmic molecular delivery with shock waves: Importance of impulse. Biophys. J. 2000, 79, 1821–1832. [Google Scholar] [CrossRef]
- Peng, Q.; Li, Y.; Deng, L.; Fang, J.; Yu, X. High hydrostatic pressure shapes the development and production of secondary metabolites of Mariana Trench sediment fungi. Sci. Rep. 2021, 11, 11436. [Google Scholar] [CrossRef]
- Damare, S.R.; Nagarajan, M.; Raghukumar, C. Spore germination of fungi belonging to Aspergillus species under deep-sea conditions. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 670–678. [Google Scholar] [CrossRef]
- Zhao, L.; Schaefer, D.; Marten, M.R. Assessment of elasticity and topography of Aspergillus nidulans spores via atomic force microscopy. Appl. Environ. Microbiol. 2005, 71, 955–960. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhao, L.; Schaefer, D.; Xu, H.; Modi, S.J.; LaCourse, W.R.; Marten, M.R. Elastic properties of the cell wall of Aspergillus nidulans studied with atomic force microscopy. Biotechnol. Progr. 2005, 21, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Millán-Chiu, B.; Camacho, G.; Varela-Echavarría, A.; Tamariz, E.; Fernández, F.; López-Marín, L.M.; Loske, A.M. Shock waves and DNA-cationic lipid assemblies: A synergistic approach to express exogenous genes in human cells. Ultrasound Med. Biol. 2014, 40, 1599–1608. [Google Scholar] [CrossRef] [PubMed]
- Valsecchi, I.; Dupres, V.; Michel, J.P.; Duchateau, M.; Matondo, M.; Chamilos, G.; Saveanu, C.; Guijarro, J.I.; Aimanianda, V.; Lafont, F.; et al. The puzzling construction of the conidial outer layer of Aspergillus fumigatus. Cell Microbiol. 2019, 21, e12994. [Google Scholar] [CrossRef] [PubMed]
- Reese, S.; Chelius, C.; Riekhof, W.; Marten, M.R.; Harris, S.D. Micafungin-induced cell wall damage stimulates morphological changes consistent with microcycle conidiation in Aspergillus nidulans. J. Fungi 2021, 7, 525. [Google Scholar] [CrossRef] [PubMed]
- Staschke, K.A.; Dey, S.; Zaborske, J.M.; Palam, L.R.; McClintick, J.N.; Pan, T.; Edenberg, H.J.; Wek, R.C. Integration of general amino acid control and target of rapamycin (TOR) regulatory pathways in nitrogen assimilation in yeast. J. Biol. Chem. 2010, 285, 16893–16911. [Google Scholar] [CrossRef] [PubMed]
- Peeters, K.; Thevelein, J.M. Glucose Sensing and Signal Transduction in Saccharomyces cerevisiae. In Molecular Mechanisms in Yeast Carbon Metabolism; Piškur, J., Compagno, C., Eds.; Springer: Berlin, Germany, 2014; pp. 21–56. [Google Scholar]
- Mamoun, C.B.; Beckerich, J.M.; Gaillardin, C.; Kepes, F. Disruption of YHC8, a member of the TSR1 gene family, reveals its direct involvement in yeast protein translocation. J. Biol. Chem. 1999, 274, 11296–11302. [Google Scholar] [CrossRef] [PubMed]
- Zu, T.; Verna, J.; Ballester, R. Mutations in WSC genes for putative stress receptors result in sensitivity to multiple stress conditions and impairment of Rlm1-dependent gene expression in Saccharomyces cerevisiae. Mol. Genet. Genom. 2001, 266, 142–155. [Google Scholar] [CrossRef]
- Futagami, T.; Nakao, S.; Kido, Y.; Oka, T.; Kajiwara, Y.; Takashita, H.; Omori, T.; Furukawa, K.; Goto, M. Putative stress sensors WscA and WscB are involved in hypo-osmotic and acidic pH stress tolerance in Aspergillus nidulans. Eukaryot. Cell 2011, 10, 1504–1515. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.M.; Chen, Y.; Zhu, J.; Ying, S.H.; Feng, M.G. Subcellular localization of five singular WSC domain-containing proteins and their roles in Beauveria bassiana responses to stress cues and metal ions. Environ. Microbiol. Rep. 2016, 8, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rocha, A.L.; Roncero, M.I.G.; López-Ramirez, A.; Mariné, M.; Guarro, J.; Martínez-Cadena, G.; Di Pietro, A. Rho1 has distinct functions in morphogenesis, cell wall biosynthesis and virulence of Fusarium oxysporum. Cell Microbiol. 2008, 10, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Delley, P.A.; Hall, M.N. Cell wall stress depolarizes cell growth via hyperactivation of RHO1. J. Cell Biol. 1999, 147, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.E. Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 262–291. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.C.; Kim, J.H.; Han, K.H. The Conserved MAP kinase MpkB regulates development and sporulation without affecting aflatoxin biosynthesis in Aspergillus flavus. J. Fungi 2020, 6, 289. [Google Scholar] [CrossRef]
- Vasara, T.; Salusjärvi, L.; Raudaskoski, M.; Keränen, S.; Penttilä, M.; Saloheimo, M. Interactions of the Trichoderma reesei rho3 with the secretory pathway in yeast and T. reesei. Mol. Microbiol. 2001, 42, 1349–1361. [Google Scholar] [CrossRef] [PubMed]
- Adamo, J.E.; Rossi, G.; Brennwald, P. The Rho GTPase Rho3 has a direct role in exocytosis that is distinct from its role in actin polarity. Mol. Biol. Cell 1999, 10, 4121–4133. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Li, B.; Qin, G.; Tian, S. Function of small GTPase Rho3 in regulating growth, conidiation and virulence of Botrytis cinerea. Fungal Genet. Biol. 2015, 75, 46–55. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence 5′–3′ | Product Size (bp) |
|---|---|---|
| β-Actin | Forward: CACCGGTATCGTTCTGGACTCT | 427 |
| Reverse: ACGGACATCAACATCACACTTCAT | ||
| Wsc4 | Forward: GCGTGGCTCTTCTCAGATT | 378 |
| Reverse: GCACTGCATCGTTCGCTATC | ||
| RhoA | Forward: CGTCCCCTCTCATACCCTGA | 506 |
| Reverse: GCACACATAGTGGAACACGC | ||
| RhoC | Forward: GCACGTTTATGCACCCTCAC | 518 |
| Reverse: CCAGAATGAGCGGGGTAGTG |
| Single-Pulse | CFU (1 × 106)/mL | Viability (%) | Tandem | CFU (1 × 106)/mL | Viability (%) |
|---|---|---|---|---|---|
| Control | 1.40 ± 0.17 a | 100.00 ± 11.84 | Control | 1.40 ± 0.17 | 100.0 ± 11.84 |
| 4 b–50 c | 1.33 ± 0.23 | 94.98 ± 8.27 | 4–50 | 1.28 ± 0.20 | 90.9 ± 6.00 |
| 4–100 | 1.39 ± 0.32 | 98.49 ± 14.28 | 4–100 | 1.37 ± 0.37 | 96.5 ± 18.00 |
| 4–200 | 1.37 ± 0.41 | 96.40 ± 20.40 | 4–200 | 1.31 ± 0.18 | 93.3 ± 4.33 |
| 5–50 | 1.20 ± 0.17 | 86.03 ± 4.49 | 5–50 | 1.18 ± 0.20 | 83.7 ± 7.28 |
| 5–100 | 1.16 ± 0.17 | 82.97 ± 4.57 | 5–100 | 1.30 ± 0.26 | 92.2 ± 10.20 |
| 5–200 | 1.00 ± 0.19 | 71.54 ± 5.87 | 5–200 | 1.18 ± 0.17 | 84.2 ± 4.80 |
| 6–50 | 1.31 ± 0.19 | 93.35 ± 4.73 | 6–50 | 1.17 ± 0.13 | 83.7 ± 1.64 |
| 6–100 | 1.25 ± 0.17 | 89.40 ± 5.24 | 6–100 | 1.31 ± 0.12 | 94.0 ± 0.38 |
| 6–200 | 1.24 ± 0.18 | 88.75 ± 4.89 | 6–200 | 1.22 ± 0.24 | 86.5 ± 9.60 |
| Incubation Time: 2 h (ng/4 µL) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Ctrl 0 h | Ctrl | 4 kV-SP a | 5 kV-SP | 6 kV-SP | 4 kV-T b | 5 kV-T | 6 kV-T | |
| β-Actin | - c | 43.5 | - | - | - | 85 | - | - |
| Wsc4 | - | - | 18.5 | - | - | - | - | - |
| RhoA | - | - | - | - | - | 9.2 | - | - |
| RhoC | - | - | 8.5 | 8.3 | - | 15.2 | - | 11 |
| Incubation Time: 8 h (ng/4 µL) | ||||||||
| Ctrl 0 h | Ctrl | 4 kV-SP | 5 kV-SP | 6 kV-SP | 4 kV-T | 5 kV-T | 6 kV-T | |
| β-Actin | - | 103 | 120 | 156 | 82 | 131.6 | 155 | 90 |
| Wsc4 | - | 54.3 | 59.3 | - | - | 57.5 | 43.3 | 27 |
| RhoA | - | 10 | 14 | 12 | 12.3 | 14.5 | 14.1 | 12 |
| RhoC | - | 16.6 | 56.7 | 23 | 12.6 | 15.6 | 26.4 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larrañaga-Ordaz, D.; Martínez-Maldonado, M.A.; Millán-Chiu, B.E.; Fernández, F.; Castaño-Tostado, E.; Gómez-Lim, M.Á.; Loske, A.M. Effect of Shock Waves on the Growth of Aspergillus niger Conidia: Evaluation of Germination and Preliminary Study on Gene Expression. J. Fungi 2022, 8, 1117. https://doi.org/10.3390/jof8111117
Larrañaga-Ordaz D, Martínez-Maldonado MA, Millán-Chiu BE, Fernández F, Castaño-Tostado E, Gómez-Lim MÁ, Loske AM. Effect of Shock Waves on the Growth of Aspergillus niger Conidia: Evaluation of Germination and Preliminary Study on Gene Expression. Journal of Fungi. 2022; 8(11):1117. https://doi.org/10.3390/jof8111117
Chicago/Turabian StyleLarrañaga-Ordaz, Daniel, Miguel A. Martínez-Maldonado, Blanca E. Millán-Chiu, Francisco Fernández, Eduardo Castaño-Tostado, Miguel Ángel Gómez-Lim, and Achim M. Loske. 2022. "Effect of Shock Waves on the Growth of Aspergillus niger Conidia: Evaluation of Germination and Preliminary Study on Gene Expression" Journal of Fungi 8, no. 11: 1117. https://doi.org/10.3390/jof8111117
APA StyleLarrañaga-Ordaz, D., Martínez-Maldonado, M. A., Millán-Chiu, B. E., Fernández, F., Castaño-Tostado, E., Gómez-Lim, M. Á., & Loske, A. M. (2022). Effect of Shock Waves on the Growth of Aspergillus niger Conidia: Evaluation of Germination and Preliminary Study on Gene Expression. Journal of Fungi, 8(11), 1117. https://doi.org/10.3390/jof8111117