
In Vitro Assessment of Organic and Residual Fractions of Nematicidal Culture Filtrates from Thirteen Tropical Trichoderma Strains and Metabolic Profiles of Most-Active

 , ,
, ,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
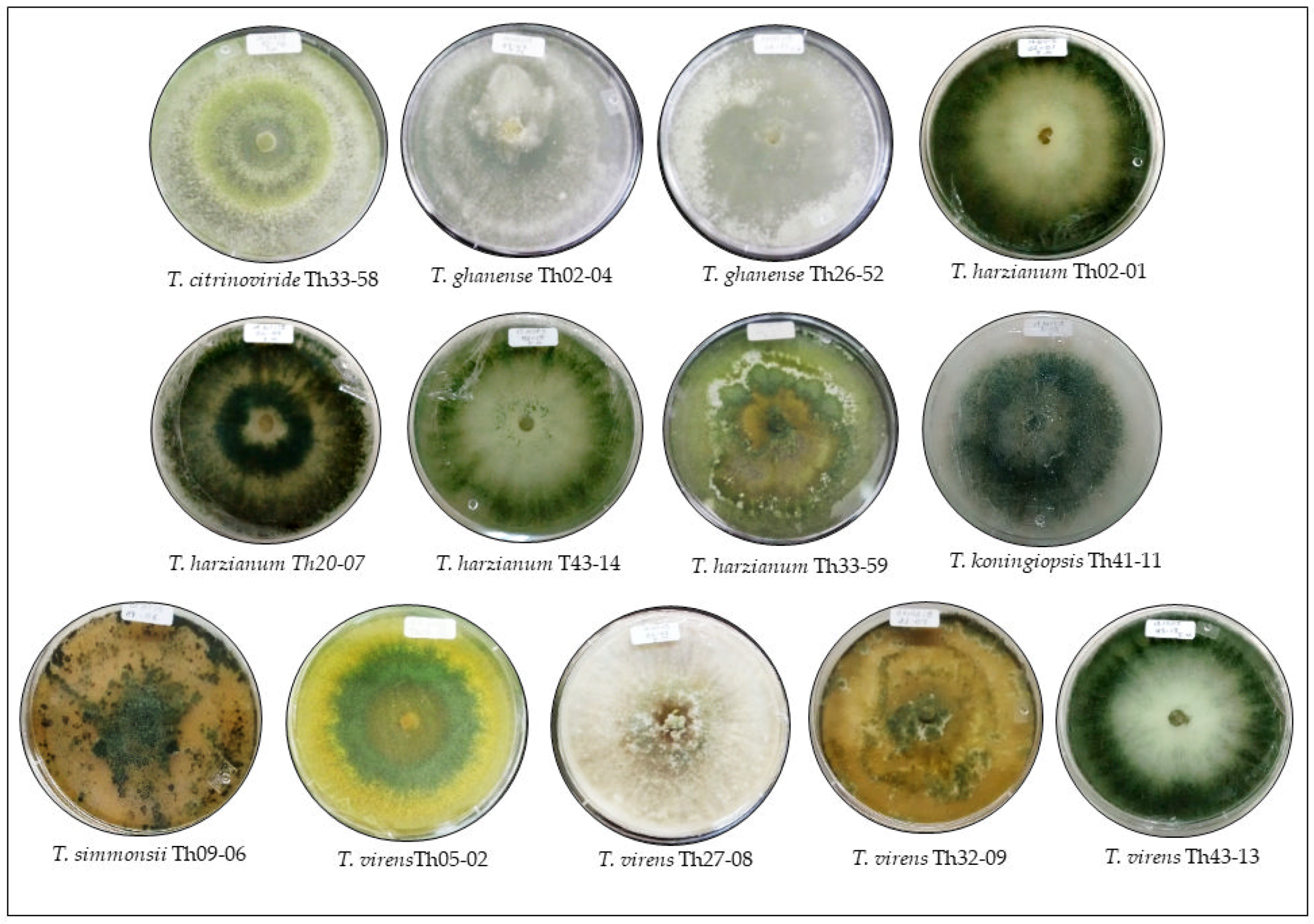
2.1. Trichoderma Strains
Liquid Culture of Trichoderma Strains
2.2. Preparation of Fractions from Culture Filtrates
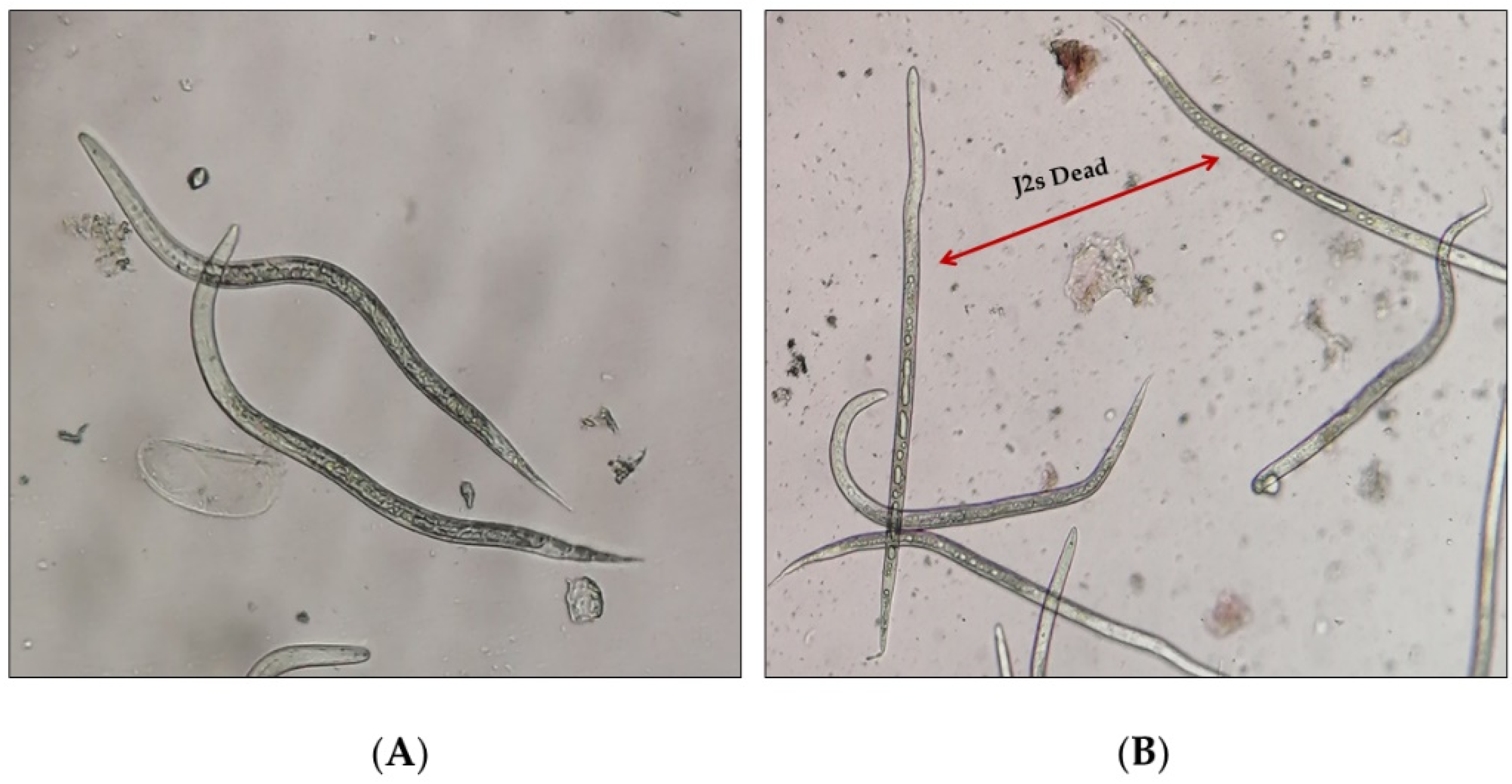
2.3. Nematicidal Bioassay
2.3.1. Nematode Inoculum
2.3.2. Sample Preparation and Nematicidal Bioassay
2.4. Liquid Chromatography-Diode Array Detector-Electrospray-High Resolution Mass Spectrometry
3. Results
3.1. Nematicidal Activity
3.2. Identification of Components in Active Fractions from Trichoderma Strains by LC-DAD-ESI-HRMS
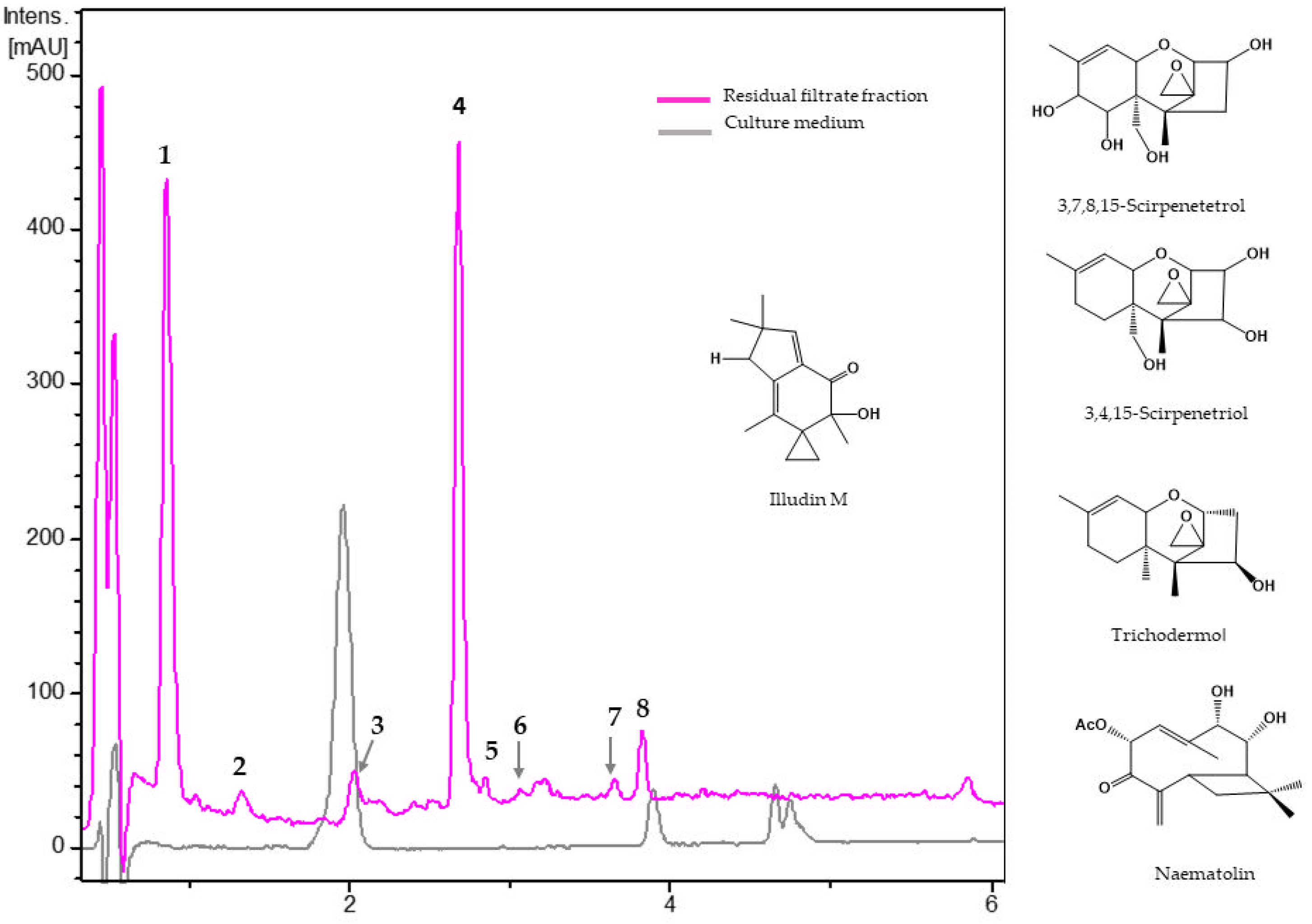
3.2.1. Metabolites from Residual Filtrate Fraction of Trichoderma harzianum Th43-14

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | UV (nm) | [M + H]+ m/z | Molecular Formula | Putative Metabolite | |
|---|---|---|---|---|---|---|
| Experimental | Theoretical | |||||
| 1 | 0.88 | 200, 215 | 317.1599 | 317.1595 | C15H24O7 | unknown |
| 2 | 1.35 | 205, 210 | 299.1492 | 299.1489 | C15H22O6 | 3,7,8,15-Scirpenetetrol |
| 3 | 2.05 | 200, 220 | 249.1488 | 249.1485 | C15H20O3 | Illudin M |
| 4 | 2.69 | 200, 220 | 255.1591 | 255.1591 | C14H22O4 | unknown |
| 5 | 2.86 | 200, 220 | 283.1541 | 283.1540 | C15H22O5 | 3,4,15-Scirpenetriol |
| 6 | 3.09 | 200, 218, 270 | 309.1692 | 309.1697 | C17H24O5 | Naematolin |
| 7 | 3.66 | 200, 220 | 553.3000 | 553.3007 | C29H44O10 | unknown |
| 8 | 3.84 | 200, 220 | 251.1644 | 251.1642 | C15H22O3 | Trichodermol |
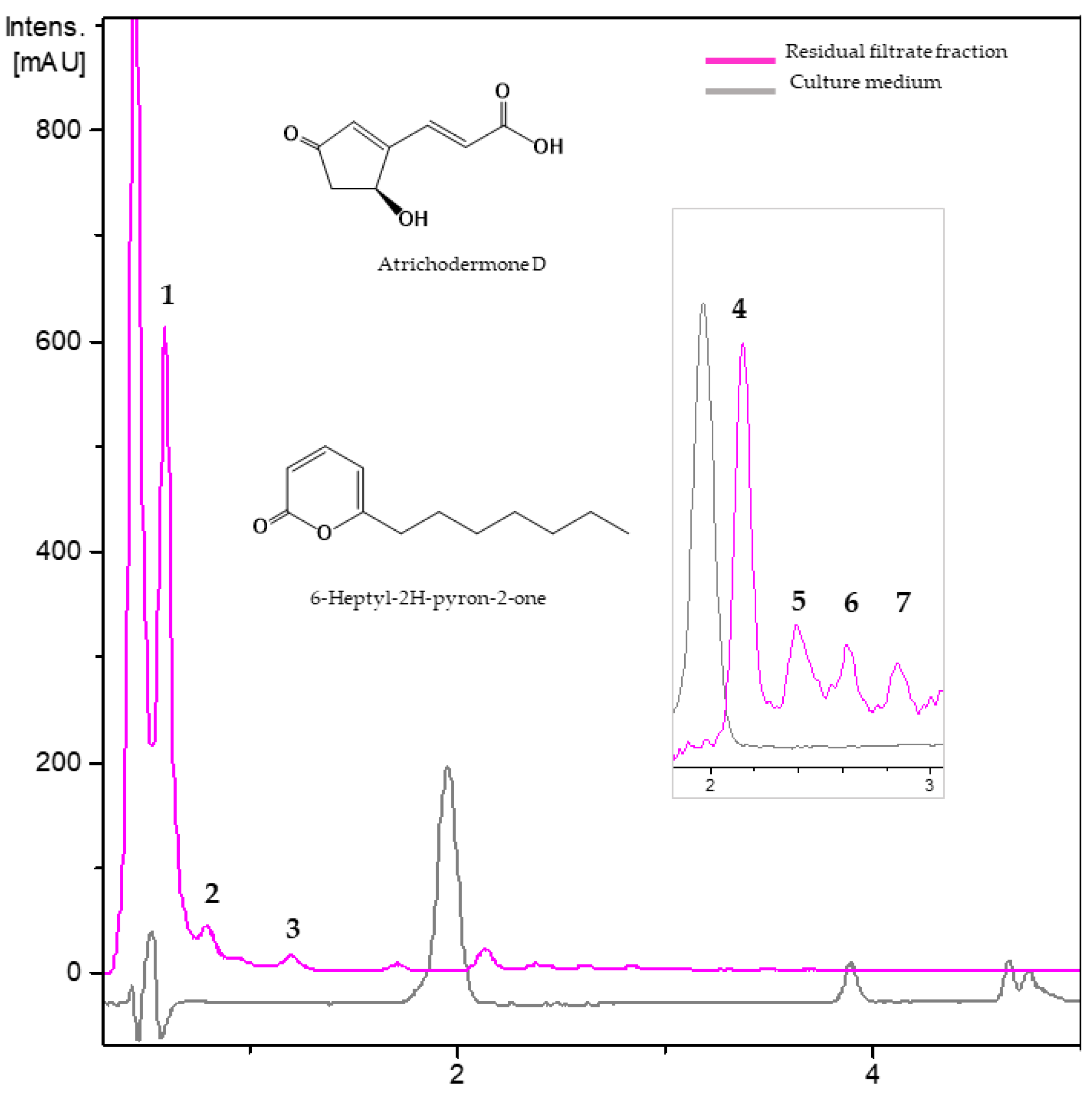
3.2.2. Metabolites from Residual Filtrate Fraction of Trichoderma ghanense Th26-52
| No. | Rt (min) | UV (nm) | [M + H]+ m/z | Molecular Formula | Putative Metabolite | |
|---|---|---|---|---|---|---|
| Experimental | Theoretical | |||||
| 1 | 0.63 | 200, 218 | 261.0966 | 261.0969 | C11H16O7 | unknown |
| 2 | 0.86 | 218, 230, 300 | 168.0656 | 168.0655 | C8H9NO3 | unknown |
| 3 | 1.22 | 205, 290 | 155.0700 | 155.0703 | C8H10O3 | Unknown |
| 4 | 2.16 | 200, 220, 310 | 169.0493 | 169.0495 | C8H8O4 | atrichodermone D |
| 5 | 2.40 | 200, 210, 240 | 225.0756 | 225.0758 | C11H12O5 | Unknown |
| 6 | 2.64 | 200, 230, 270 | 235.1328 | 235.1329 | C14H18O3 | Unknown |
| 7 | 2.89 | 200, 220, 280 | 195.1379 | 195.1380 | C12H18O2 | 6-heptyl-2H-pyron-2-one |
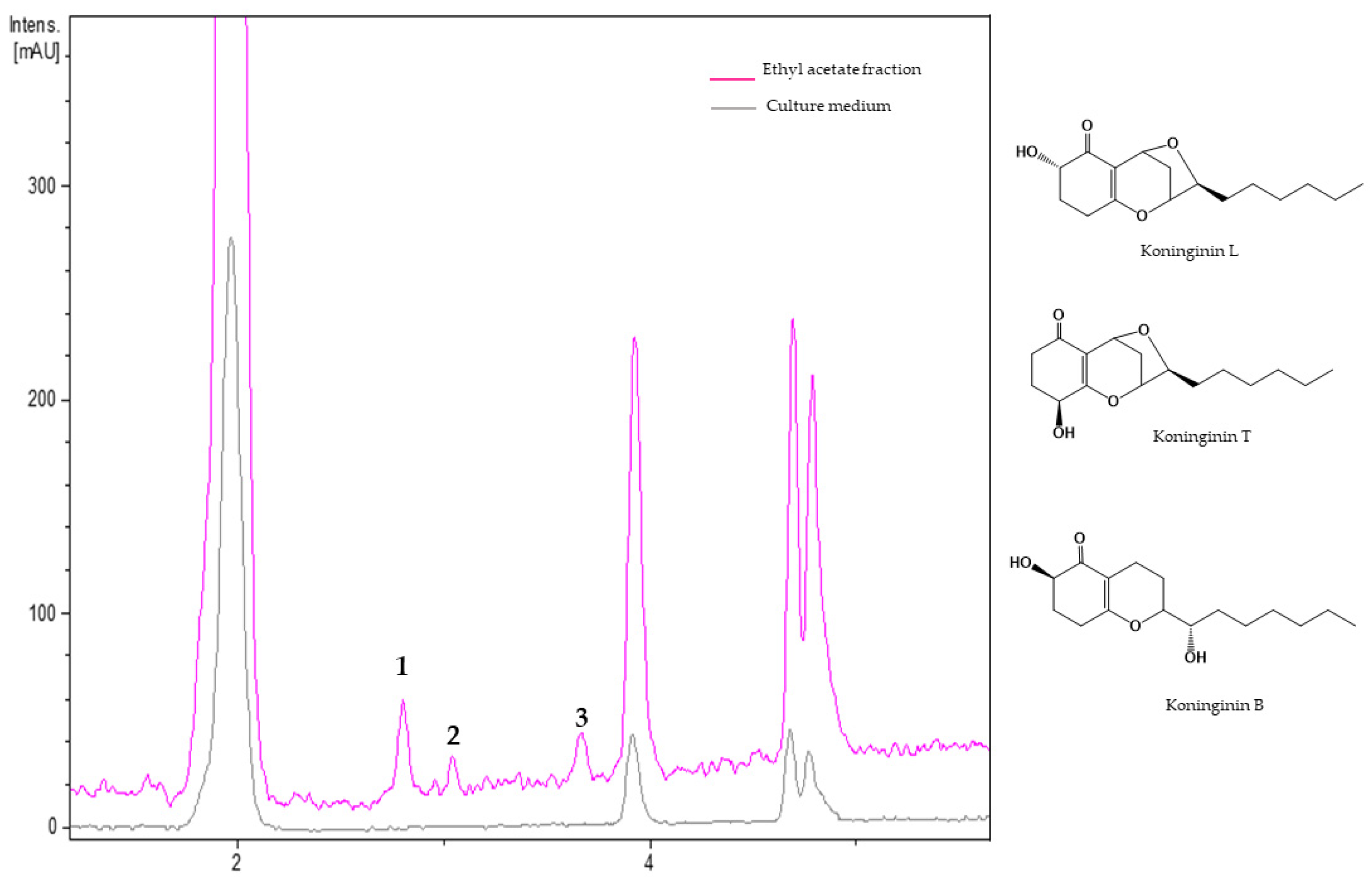
3.2.3. Metabolites from Ethyl Acetate Fraction of Trichoderma koningiopsis Th41-11
| No. | Rt (min) | UV (nm) | [M + H]+ m/z | Molecular Formula | Putative Metabolite | |
|---|---|---|---|---|---|---|
| Experimental | Theoretical | |||||
| 1 | 2.81 | 200, 255 | 281.1750 | 281.1747 | C16H24O4 | Koninginin L |
| 2 | 3.04 | 200, 255 | 281.1749 | 281.1747 | C16H24O4 | Koninginin T |
| 3 | 3.68 | 200, 260 | 283.1906 | 283.1904 | C16H26O4 | Koninginin B |
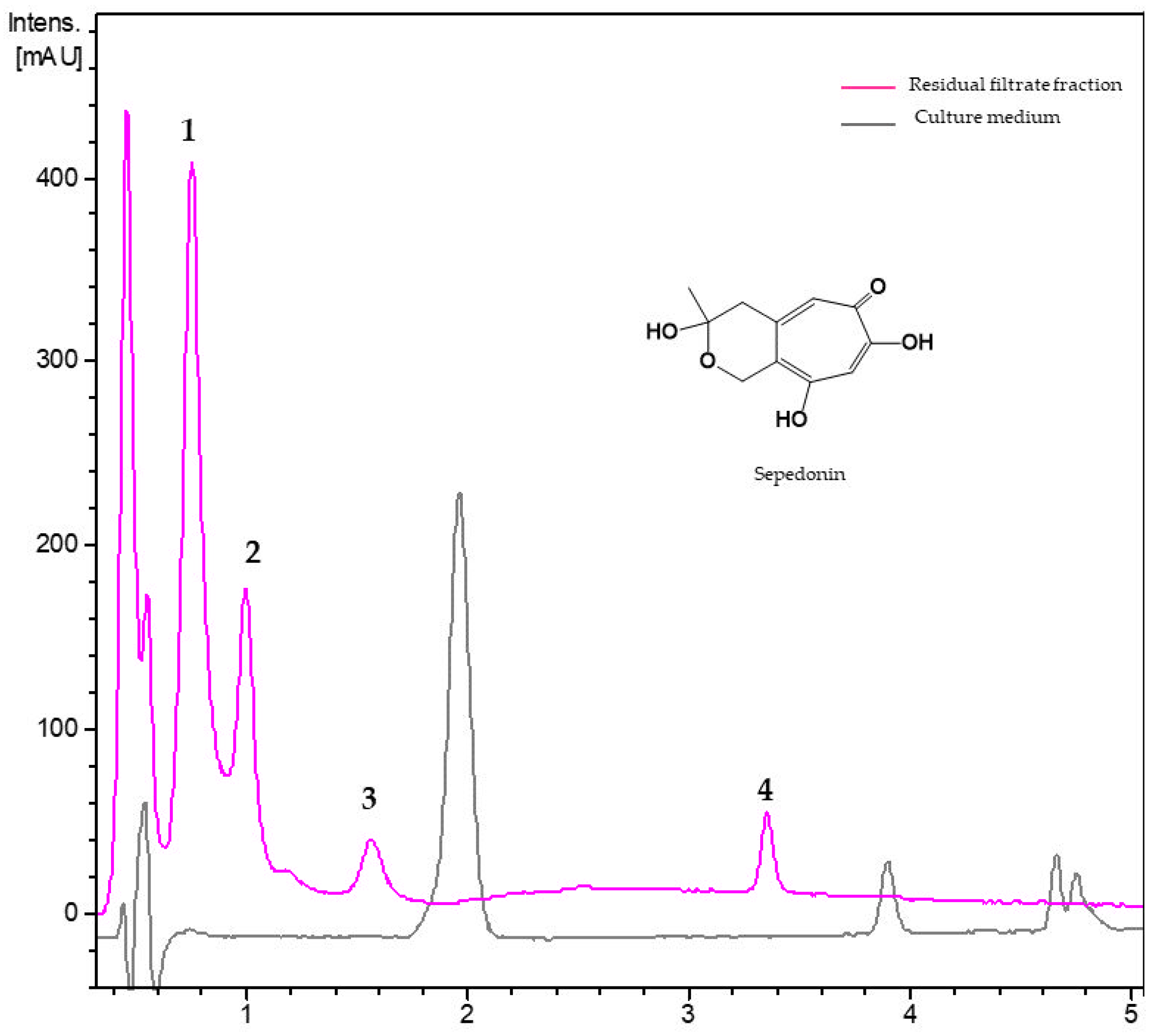
3.2.4. Metabolites from Residual Filtrate Fraction of Trichoderma virens Th27-08
| No. | Rt (min) | UV (nm) | [M + H]+ m/z | Molecular Formula | Putative Metabolite | |
|---|---|---|---|---|---|---|
| Experimental | Theoretical | |||||
| 1 | 0.78 | 205, 240, 290 | 207.0653 | 207.0657 | C11H10O4 | unknown |
| 2 | 1.00 | 210, 225, 295, | 225.0757 | 225.0762 | C11H12O5 | sepedonin |
| 3 | 1.57 | 205, 240, 290 | 207.0652 | 207.0657 | C11H10O4 | unknown |
| 4 | 3.35 | 215, 255, 300 | 277.1435 | 277.1438 | C16H20O4 | unknown |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 20 November 2021).
- Li, M.F.; Li, G.H.; Zhang, K.Q. Non-volatile metabolites from Trichoderma spp. Metabolites 2019, 9, 58. [Google Scholar] [CrossRef]
- Li, G.; Zhang, K.; Xu, J.; Dong, J.; Liu, Y. Nematicidal substances from fungi. Recent Pat. Biotechnol. 2007, 1, 212–233. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, G.; Khan, A.; Khan, A.A.; Ali, A.; Mohhamad, H.I. Biological control: A novel strategy for the control of the plant parasitic nematodes. Ant. Leeuw. 2021, 114, 885–912. [Google Scholar] [CrossRef]
- Kaushik, N.; Díaz, C.E.; Chhipa, H.; Julio, L.F.; Andrés, M.F.; González-Coloma, A. Chemical composition of an aphid antifeedant extract from an endophytic fungus Trichoderma sp. EFI671. Microorganisms 2020, 8, 420. [Google Scholar] [CrossRef]
- Javaid, A.; Ali, S. Herbicidal activity of culture filtrates of Trichoderma spp. against two problematic weeds of wheat. Nat. Prod. Res. 2011, 25, 730–740. [Google Scholar] [CrossRef]
- Baazeem, A.; Almanea, A.; Manikandan, P.; Alorabi, M.; Vijayaraghavan, P.; Abdel-Hadi, A. In vitro antibacterial, antifungal, nematocidal and growth promoting activities of Trichoderma hamatum FB10 and its secondary metabolites. J. Fungi 2021, 7, 331. [Google Scholar] [CrossRef] [PubMed]
- TariqJaveed, M.; Farooq, T.; Al-Hazmi, A.S.; Hussain, M.D.; Rehman, A.U. Role of Trichoderma as a biocontrol agent (BCA) of phytoparasitic nematodes and plant growth inducer. J. Invertebr. Pathol. 2021, 183, 107626107626. [Google Scholar] [CrossRef]
- Braithwaite, M.; Clouston, A.; Minchin, R.; Yardley, J.; Nieto-Jacobo, M.F.; Mendoza-Mendoza, A.; Steyaert, J.; Hill, R.; Marshall, J.; Stewart, A. The density-dependent effect of initial nematode population levels on the efficacy of Trichoderma as a bio-nematicide against Meloidogyne hapla on tomato. Australas. Plant Pathol. 2016, 45, 473–479. [Google Scholar] [CrossRef]
- Cardona-Piedrahita, L.F.; Castaño-Zapata, J.; Ceballos-Aguirre, N. Respuesta de quince introducciones de tomate cherry (Solanum lycopersicum L.) al nemátodo nodulador (Meloidogyne spp. Goeldi) e identificación de las especies. Rev. Acad. Colomb. Cienc. Exactas. Fis. Nat. 2016, 40, 450–460. [Google Scholar] [CrossRef][Green Version]
- Abd-Elgawad, M.M.M.; Askary, T.H. Fungal and bacterial nematicides in integrated nematode management strategies. Egypt. J. Biol. Pest. Control 2018, 28, 74. [Google Scholar] [CrossRef]
- Hassan, M.A.; Pham, T.H.; Shi, H.; Zheng, J. Nematodes threats to global food security. Acta Agric. Scand, Sect. B-Soil Plant Sci. 2013, 63, 420–425. [Google Scholar] [CrossRef]
- Collange, B.; Navarrete, M.; Peyre, G.; Mateille, T.; Tchamitchian, M. Root-knot nematode (Meloidogyne) management in vegetable crop production: The challenge of an agronomic system analysis. Crop Protect. 2011, 30, 1251–1262. [Google Scholar] [CrossRef]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: Metabolites from nematophagous ascomycetes. App. Microbiol. Biotechnol. 2016, 100, 3799–3812. [Google Scholar] [CrossRef] [PubMed]
- Baldoni, D.B.; Antoniolli, Z.I.; Mazutti, M.A.; Jacques, R.J.S.; Dotto, A.C.; de Oliveira Silveira, A.; Ferraz, R.C.; Soares, V.B.; de Souza, A.R.C. Chitinase production by Trichoderma koningiopsis UFSMQ40 using solid state fermentation. Braz. J. Microbiol. 2020, 51, 1897–1908. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Yao, M.; Wang, H.; Zhao, D.; Zhu, X.; Wang, Y.; Liu, X.; Duan, Y.; Chen, L. Isolation and effect of Trichoderma citrinoviride Snef1910 for the biological control of root-knot nematode, Meloidogyne incognita. BMC Microbiol. 2020, 20, 1–11. [Google Scholar] [CrossRef]
- Khan, R.A.A.; Najeeb, S.; Mao, Z.; Ling, J.; Yang, Y.; Li, Y.; Xie, B. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic bacteria and root-knot nematode. Microorganisms 2020, 8, 401. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.A.; Ruqqia, A.; Sultana, V.; Ara, J.; Ehteshamul-Haque, S. Nematicidal potential of culture filtrates of soil fungi associated with rhizosphere and rhizoplane of cultivated and wild plants. Pak. J. Bot. 2012, 44, 1041–1046. [Google Scholar]
- Elgorban, A.M.; Abdel-Wahab, M.A.; Bahkali, A.H.; Al-Sum, B.A. Biocontrol of Meloidogyne javanica on tomato plants by Hypocrea lixii (the Teleomorph of Trichoderma harzianum). Clean-Soil Air Water 2014, 42, 1464–1469. [Google Scholar] [CrossRef]
- Jindapunnapat, K.; Chinnasri, B.; Kwankuae, S. Biological control of root-knot nematodes (Meloidogyne enterolobii) in guava by the fungus Trichoderma harzianum. J. Dev. Sustain. Agric. 2013, 8, 110–118. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Barbetti, M.J.; Li, H.; Woo, S.L.; Lorito, M. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant. Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Yan, Y.; Mao, Q.; Wang, Y.; Zhao, J.; Fu, Y.; Yang, Z.; Pen, X.; Zhang, M.; Bai, B.; Liu, A.; et al. Trichoderma harzianum induces resistance to root-knot nematodes by increasing secondary metabolite synthesis and defense-related enzyme activity in Solanum lycopersicum L. Biol. Control 2021, 158, 104609. [Google Scholar] [CrossRef]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Bansal, R.; Pachauri, S.; Gururajaiah, D.; Sherkhane, P.D.; Khan, Z.; Gupta, S.; Banerjee, K.; Kumar, A.; Mukherjee, P.K. Dual role of a dedicated GAPDH in the biosynthesis of volatile and non-volatile metabolites-novel insights into the regulation of secondary metabolism in Trichoderma virens. Microbiol. Res. 2021, 253, 126862. [Google Scholar] [CrossRef]
- Candelero, D.J.; Cristóbal, A.J.; Reyes, R.A.; Gamboa, A.M.M.; Ruíz, S.E.; Tun, S.J.M. Trichoderma spp. promotoras del crecimiento en plántulas de Capsicum chinense Jacq. y antagónicas contra Meloidogyne incognita. Phyton. Int. J. Exp. Bot. 2015, 84, 113–119. [Google Scholar] [CrossRef]
- Cristóbal-Alejo, J.; Cetz-Chi, J.I.; Tún-Suárez, J.M.; Moo-Koh, F.A.; Peraza-Luna, F.A.; Candelero-De la Cruz, J. Filtrados fúngicos de Trichoderma con actividad nematicida contra Meloidogyne incognita (Kofoid & White) Chitwood. Rev. Prot. Veg. 2018, 33, 1–8. [Google Scholar]
- Moo-Koh, F.A.; Cristóbal-Alejo, J.; Reyes-Ramírez, A.; Tun Suárez, J.M.; Gamboa-Angulo, M. Identificación molecular de aislados de Trichoderma spp. y su actividad promotora en Solanum lycopersicum L. Investig. Cienc. 2017, 75, 5–11. [Google Scholar] [CrossRef]
- Moo Koh, F.A.; Cristóbal Alejo, J.; Reyes Ramírez, A.; Tun Suárez, J.M.; Gamboa Angulo, M.; Islas-Flores, I.R. Incompatibilidad interespecífica de especies de Trichoderma contra Meloidogyne incognita en Solanum lycopersicum. Sci. Fungorum 2018, 47, 37–45. [Google Scholar] [CrossRef][Green Version]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Cristóbal-Alejo, J.; Tun-Suárez, J.M.; Moguel-Catzin, S.; Marbán-Mendoza, N.; Medina-Baizabal, L.; Simá-Polanco, P.; Peraza-Sanchez, S.R.; Gamboa-Angulo, M.M. In vitro sensitivity of Meloidogyne incognita to extracts from native yucatecan plants. Nematropica 2006, 36, 89–97. [Google Scholar]
- Julio, L.F.; González-Coloma, A.; Burillo, J.; Diaz, C.E.; Andrés, M.F. Nematicidal activity of the hydrolate byproduct from the semi industrial vapor pressure extraction of domesticated Artemisia absinthium against Meloidogyne javanica. Crop Prot. 2017, 94, 33–37. [Google Scholar] [CrossRef]
- Schneider-Orelli, O. Entomologisches Praktikum: Einführung in die land-und. Forstwirtschaftliche Insektenkunde; Aarau Sauerländer: Aarau, Switzerland, 1947; pp. 1–237. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión; Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba: Cordobá, Argentina, 2020; Available online: http://www.infostat.com.ar (accessed on 12 July 2021).
- Martín, J.; Crespo, G.; González-Menéndez, V.; Pérez-Moreno, G.; Sánchez-Carrasco, P.; Pérez-Victoria, I.; Ruiz-Pérez, L.M.; González-Packanowska, D.; Vicente, F.; Genilloud, O.; et al. MDN-0104, an antiplasmodial betaine lipid from Heterospora chenopodii. J. Nat. Prod. 2014, 77, 2118–2123. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S. Trichoderma harzianum enhances the production of nematicidal compounds in vitro and improves biocontrol of Meloidogyne javanica by Pseudomonas fluorescens in tomato. Lett. Appl. Microbiol. 2004, 38, 169–175. [Google Scholar] [CrossRef]
- Migunova, V.; Sasanelli, N.; Kurakov, A. Effect of microscopic fungi on larval mortality of the root-knot nematodes Meloidogyne incognita and Meloidogyne javanica. Biol. Integ. Control Plant Pathog. IOBC-WPRS Bull. 2018, 133, 27–31. [Google Scholar]
- Ntalli, N.; Menkissoglu-Spiroudi, U.; Doitsinis, K.; Kalomoiris, M.; Papadakis, E.N.; Boutsis, G.; Dimou, M.; Monokrousos, N. Mode of action and ecotoxicity of hexanoic and acetic acids on Meloidogyne javanica. J. Pest Sci. 2020, 93, 867–877. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Vargiu, S.; Menkissoglu-Spiroudi, U.; Caboni, P. Nematicidal carboxylic acids and aldehydes from Melia azedarach fruits. J. Agric. Food Chem. 2010, 58, 11390–11394. [Google Scholar] [CrossRef]
- Nouranim, S.L.; Mohammadi-Goltapeh, E.; Safaie, N.; Javaran, M.J.; Pourjam, E.; Shams-Bakhsh, M.; Afshar, F.J. The effects of Arthrobotrys oligospora and Arthrobotrys conoides culture filtrates on second stage juvenile mortality and egg hatching of Meloidogyne incognita and Meloidogyne javanica. J. Crop Prot. 2015, 4, 667–674. [Google Scholar]
- Babu, B.V.; Kamra, A.; Paul, S.; Devi, T.P. Antibiosis and egg parasitization in root-knot nematode, Meloidogyne incognita by indigenous isolates of Trichoderma harzianum rifai, 1969 in relation to chitinase and protease levels. Indian J. Nematol. 2019, 49, 187–192. [Google Scholar]
- Tiwari, S.; Pandey, S.; Chauhan, P.S.; Pandey, R. Biocontrol agents in co-inoculation manages root knot nematode [Meloidogyne incognita (Kofoid & White) Chitwood] and enhances essential oil content in Ocimum basilicum L. Ind. Crops Prod. 2017, 97, 292–301. [Google Scholar] [CrossRef]
- Sellami, S.; Benttoumi, N.; Berrahia, S.; Boureghda, H. Evaluation of antagonistic activity of Trichoderma spp. against Meloidogyne incognita. Acta Phytopathol. Entomol. Hung. 2017, 52, 177–184. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Gräfenhan, T.; Zafari, D.; Thrane, U. Trichothecene Production by Trichoderma brevicompactum. J. Agric. Food. Chem. 2005, 53, 8190–8196. [Google Scholar] [CrossRef]
- Schollenberger, M.; Drochner, W.; Müller, H.-M. Fusarium toxins of the scirpentriol subgroup: A review. Mycopathologia 2007, 164, 101–118. [Google Scholar] [CrossRef]
- Ayer, W.A.; Miao, S. Secondary metabolites of the aspen fungus Stachybotrys cylindrospora. Can. J. Chem. 1993, 71, 487–493. [Google Scholar] [CrossRef]
- Barúa, J.E.; de la Cruz, M.; de Pedro, N.; Cautain, B.; Hermosa, R.; Cardoza, R.E.; Gutiérrez, S.; Monte, E.; Vicente, F.; Collado, I.G. Synthesis of trichodermin derivatives and their antimicrobial and cytotoxic activities. Molecules 2019, 24, 3811. [Google Scholar] [CrossRef] [PubMed]
- Kononenko, G.P.; Soboleva, N.A.; Leonov, A.N. 3, 7, 8, 15-Tetrahydroxy-12, 13-epoxytrichothec-9-en in a culture of Fusarium graminearum. Chem. Nat. Comp. 1990, 26, 218–220. [Google Scholar] [CrossRef]
- Mayer, A.; Anke, H.; Sterner, O. Omphalotin, A new cyclic peptide with potent nematicidal activity from Omphalotus olearius I. Fermentation and biological activity. Nat. Prod. Lett. 1997, 10, 25–32. [Google Scholar] [CrossRef]
- McMorris, T.C.; Anchel, M. Fungal metabolites. The structures of the novel sesquiterpenoids illudin-S and -M. J. Am. Chem. Soc. 1965, 87, 1594–1600. [Google Scholar] [CrossRef]
- Nord, C.; Menkis, A.; Broberg, A. Cytotoxic illudane sesquiterpenes from the fungus Granulobasidium vellereum (Ellis and Cragin) Jülich. J. Nat. Prod. 2015, 78, 2559–2564. [Google Scholar] [CrossRef]
- Ito, Y.; Kurita, H.; Yamaguchi, T.; Sato, M.; Okuda, T. Naematolin, a new biologically active substance produced by Naematoloma fasciculare (Fr.) Karst. Chem. Pharm. Bull. 1967, 15, 2009–2010. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Abraham, W.R. Bioactive sesquiterpenes produced by fungi: Are they useful for humans as well? Curr. Med. Chem. 2001, 8, 583–606. [Google Scholar] [CrossRef]
- Al-salihi, S.A.A.; Dao, T.T.; Williams, K.; Bailey, A.M.; Foster, G.D. The biogenetic origin of the biologically active naematolin of Hypholoma species involves an unusual sesquiterpene synthase. Mol. Biotechnol. 2019, 61, 754–762. [Google Scholar] [CrossRef]
- Inchausti, A.; Yaluff, G.; Rojas de Arias, A.; Torres, S.; Ferreira, M.E.; Nakayama, H.; Schinini, A.; Lorenzen, K.; Anke, T.; Fournet, A. Leishmanicidal and trypanocidal activity of extracts and secondary metabolites from basidiomycetes. Phytother. Res. 1997, 11, 193–197. [Google Scholar] [CrossRef]
- Singh, A.; Sarma, B.K.; Singh, U.P.; Singh, R.; Singh, H.B.; Singh, K.P. Metabolite profiling can assist variability analysis in Trichoderma species. Arch. Phytopathol. Pflanzenschutz 2011, 44, 1697–1702. [Google Scholar] [CrossRef]
- Zhou, P.; Wu, Z.; Tan, D.; Yang, J.; Zhou, Q.; Zeng, F.; Zhang, M.; Bie, Q.; Chen, C.; Xue, Y.; et al. Atrichodermones A–C, three new secondary metabolites from the solid culture of an endophytic fungal strain, Trichoderma atroviride. Fitoterapia 2017, 123, 18–22. [Google Scholar] [CrossRef]
- Simon, A.; Dunlop, R.W.; Ghisalberti, E.L.; Sivasithamparam, K. Trichoderma koningii produces a pyrone compound with antibiotic properties. Soil Biol. Biochem. 1988, 20, 263–264. [Google Scholar] [CrossRef]
- Wickel, S.M.; Citron, C.A.; Dickschat, J.S. 2H-Pyran-2-ones from Trichoderma viride and Trichoderma asperellum. Eur. J. Org. Chem. 2013, 2013, 2906–2913. [Google Scholar] [CrossRef]
- Shi, X.-S.; Meng, L.-H.; Li, X.; Wang, D.-J.; Zhou, X.-W.; Du, F.-Y.; Wang, B.-G.; Li, X.-M. Polyketides and terpenoids with potent antibacterial activities from the Artemisia argyi-derived fungus Trichoderma koningiopsis QA-3. Chem. Biodivers. 2020, 17, e2000566. [Google Scholar] [CrossRef]
- Mazzei, P.; Vinale, F.; Woo, S.L.; Pascale, A.; Lorito, M.; Piccolo, A. Metabolomics by proton high-resolution magic-angle-spinning nuclear magnetic resonance of tomato plants treated with two secondary metabolites isolated from Trichoderma. J. Agric. Food Chem. 2016, 64, 3538–3545. [Google Scholar] [CrossRef]
- Cutler, H.G.; Himmelsbach, D.S.; Yagen, B.; Arrendale, R.F.; Jacyno, J.M.; Cole, P.D.; Cox, R.H. Koninginin B: A biologically active congener of koninginin A from Trichoderma koningii. J. Agric. Food Chem. 1991, 39, 977–980. [Google Scholar] [CrossRef]
- Zhou, X.X.; Li, J.; Yang, Y.H.; Zeng, Y.; Zhao, P.J. Three new koninginins from Trichoderma neokongii 8722. Phytochem. Lett. 2014, 8, 137–140. [Google Scholar] [CrossRef]
- Chen, L.; Wu, G.-W.; Liu, D.; Zhuang, W.-Y.; Yin, W.-B. Trichodermatides E and F from fungus Trichoderma applanatum. J. Asian Nat. Prod. Res. 2019, 21, 659–665. [Google Scholar] [CrossRef]
- Shi, X.S.; Wang, D.J.; Li, X.M.; Li, H.L.; Meng, L.H.; Li, X.; Yan, P.; Wang, X.Z.; Wang, B.G. Antimicrobial polyketides from Trichoderma koningiopsis QA-3, an endophytic fungus obtained from the medicinal plant Artemisia argyi. RSC Adv. 2017, 7, 51335–51342. [Google Scholar] [CrossRef]
- Shi, X.S.; Li, H.L.; Li, X.M.; Wang, D.J.; Li, X.; Meng, L.H.; Zhou, X.W.; Wang, B.G. Highly oxygenated polyketides produced by Trichoderma koningiopsis QA-3, an endophytic fungus obtained from the fresh roots of the medicinal plant Artemisia argyi. Bioorg. Chem. 2020, 94, 103448. [Google Scholar] [CrossRef]
- Wang, Y.L.; Hu, B.Y.; Qian, M.A.; Wang, Z.H.; Zou, J.M.; Sang, X.Y.; Li, L.; Luo, D.L.; Zhao, L.X. Koninginin W, a new polyketide from the endophytic fungus Trichoderma koningiopsis YIM PH30002. Chem. Biodivers. 2021, 18, e2100460. [Google Scholar] [CrossRef]
- Nagao, K.; Yoshida, N.; Iwai, K.; Sakai, T.; Tanaka, M.; Miyahara, T. Production of sepedonin by Sepedonium chrysospermum NT-1 in submerged culture. Environ. Sci. 2006, 13, 251–256. [Google Scholar] [PubMed]
- Divekar, P.V.; Raistrick, H.; Dobson, T.A.; Vining, L.C. Studies in the biochemistry of microorganisms: Part 117. Sepedonin, a tropolone metabolite of Sepedonium chrysospermum Fries. Can. J. Chem. 1965, 43, 1835–1848. [Google Scholar] [CrossRef]
- Quang, D.N.; Schmidt, J.; Porzel, A.; Wessjohann, L.; Haid, M.; Arnold, N. Ampullosine, a new isoquinoline alkaloid from Sepedonium ampullosporum (Ascomycetes). Nat. Prod. Commun. 2010, 5, 869–872. [Google Scholar] [CrossRef]
- Hu, Z.; Tao, Y.; Tao, X.; Su, Q.; Cai, J.; Qin, C.; Ding, W.; Li, C. Sesquiterpenes with phytopthogenic fungi inhibitory activities from fungus Trichoderma virens from Litchi chinensis Sonn. J. Agric. Food Chem. 2019, 67, 10646–10652. [Google Scholar] [CrossRef]
- Shi, X.S.; Meng, L.H.; Li, X.M.; Li, X.; Wang, D.J.; Li, H.L.; Zhou, X.W.; Wang, B.G. Trichocadinins B–G: Antimicrobial cadinane sesquiterpenes from Trichoderma virens QA-8, an endophytic fungus obtained from the medicinal plant Artemisia argyi. J. Nat. Prod. 2019, 82, 2470–2476. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.S.; Li, G.H.; Zhao, P.J.; Zheng, X.; Luo, S.L.; Li, L.; Niu, X.M.; Zhang, K.Q. Nematicidal activity of Trichoderma spp. and isolation of an active compound. World J. Microbiol. Biotechnol. 2010, 26, 2297–2302. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Ju, G.L.; Xiao, L.; Zhang, X.F.; Du, F.Y. Cyclodepsipeptides and sesquiterpenes from marine-derived fungus Trichothecium roseum and their biological functions. Mar. Drugs. 2018, 16, 519. [Google Scholar] [CrossRef]
- Ntalli, N.; Ratajczak, M.; Oplos, C.; Menkissoglu-Spiroudi, U.; Adamski, Z. Acetic acid, 2-undecanone, and (E)-2-decenal ultrastructural malformations on Meloidogyne incognita. J. Nematol. 2016, 48, 248–260. [Google Scholar] [CrossRef] [PubMed]






| Trichoderma Species | Key | GenBank Number | Activity | Place of Collection |
|---|---|---|---|---|
| T. citrinoviride Bissett 1984 | Th33-58 | MF078653 | A/B | Ticul |
| T. ghanense Yoshim. Doi, Y. Abe & Sugiy 1987 | Th02-04 | MF078652 | A/B | Tizimín |
| Th26-52 | MF078651 | A/B | Tahdziú | |
| T. harzianum Rifai 1969 | Th02-01 | MF952887 | A/B | Tizimín |
| Th20-07 | MF078650 | A | Tzucacab | |
| Th43-14 | MF078649 | A | San Felipe | |
| Th33-59 | MF078648 | A/B | Ticul | |
| T. koningiopsis Samuels, Carm. Suárez & H.C. Evans 2006 | Th41-11 | MF952888 | A/B | Sanahcat |
| T. simmonsii P. Chaverri, F.B. Rocha, Samuels, Degenkolb & Jaklitsch 2015 | Th09-06 | MF078647 | A/B | Dzidzantun |
| T. virens (J.H. Mill., Giddens & A.A. Foster) Arx 1987 | Th05-02 | MF952889 | A/B | Dzilam González |
| Th27-08 | MF078646 | A | Chacsinkin | |
| Th32-09 | MF078645 | A | Oxkutzcab | |
| Th43-13 | MF078644 | A | San Felipe |
| Trichoderma Species | Strains | Mean Percentage Mortality | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Meloidogyne incognita | Meloidogyne javanica | ||||||||||
| CF | EA | RF | CF | EA | RF | ||||||
| 100% | 50% | 1 µg/µL | 100% | 50% | 25% | 100% | 1 µg/µL | 100% | 50% | ||
| T. citrinoviride | Th33-58 | 62 b | 21 c | 30 c | 100 a | 66 c | 13 c | 15 d | 8 b | 100 a | 1 h |
| T. ghanense | Th02-04 | 100 a | 100 a | 20 d | 100 a | 63 c | 24 c | 99 a | 0 c | 100 a | 16 e |
| Th26-52 | 100 a | 23 c | 22 d | 100 a | 100 a | 20 c | 3 f | 11 b | 100 a | 63 b | |
| T. harzianum | Th02-01 | 100 a | 42 c | 0 e | 100 a | 23 d | 15 c | 4 f | 0 c | 98 a | 34 d |
| Th20-07 | 100 a | 6 d | 0 e | 98 a | 66 c | 13 c | 97 a | 0 c | 7 e | 0 h | |
| Th43-14 | 100 a | 66 b | 51 b | 100 a | 100 a | 57 b | 45 c | 66 a | 93 b | 4 g | |
| Th33-59 | 100 a | 21 c | 30 c | 100 a | 72 b | 22 c | 9 e | 9 b | 10 a | 0 h | |
| T. koningiopsis | Th41-11 | 100 a | 38 c | 54 b | 100 a | 100 a | 82 a | 7 e | 1 c | 93 b | 51 c |
| T. simmonsii | Th09-06 | 100 a | 6 d | 37 c | 100 a | 96 a | 53 b | 6 e | 0 c | 14 d | 0 h |
| T. virens | Th05-02 | 100 a | 22 c | 18 d | 100 a | 19 d | 18 c | 8 e | 0 c | 100 a | 10 f |
| Th27-08 | 100 a | 71 b | 0 e | 100 a | 100 a | 64 b | 85 b | 9 b | 100 a | 94 a | |
| Th32-09 | 100 a | 20 c | 100 a | 100 a | 100 a | 83 a | 7 e | 3 c | 86 c | 5 g | |
| Th43-13 | 100 a | 23 c | 31 c | 100 a | 86 b | 49 c | 7 e | 8 b | 95 b | 0 h | |
| Control | PDB | 0 c | 0 d | 0 e | 0 b | 0 d | 0 c | 0 f | 0 c | 0 f | 0 h |
| DW/WDT * | 0 c | 0 d | 0 e* | 0 b | 0 d | 0 c | 0 f | 0 c* | 0 f | 0 h | |
| Blank | 0 c | 0 d | 0 e | 0 b | 0 d | 0 c | 0 f | 0 c | 0 f | 0 h | |
| SD | 0.2 | 125 | 5.7 | 0.5 | 15.2 | 12.9 | 2.4 | 1.4 | 2.6 | 2.4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moo-Koh, F.A.; Cristóbal-Alejo, J.; Andrés, M.F.; Martín, J.; Reyes, F.; Tun-Suárez, J.M.; Gamboa-Angulo, M. In Vitro Assessment of Organic and Residual Fractions of Nematicidal Culture Filtrates from Thirteen Tropical Trichoderma Strains and Metabolic Profiles of Most-Active. J. Fungi 2022, 8, 82. https://doi.org/10.3390/jof8010082
Moo-Koh FA, Cristóbal-Alejo J, Andrés MF, Martín J, Reyes F, Tun-Suárez JM, Gamboa-Angulo M. In Vitro Assessment of Organic and Residual Fractions of Nematicidal Culture Filtrates from Thirteen Tropical Trichoderma Strains and Metabolic Profiles of Most-Active. Journal of Fungi. 2022; 8(1):82. https://doi.org/10.3390/jof8010082
Chicago/Turabian StyleMoo-Koh, Felicia Amalia, Jairo Cristóbal-Alejo, María Fé Andrés, Jesús Martín, Fernando Reyes, Jose María Tun-Suárez, and Marcela Gamboa-Angulo. 2022. "In Vitro Assessment of Organic and Residual Fractions of Nematicidal Culture Filtrates from Thirteen Tropical Trichoderma Strains and Metabolic Profiles of Most-Active" Journal of Fungi 8, no. 1: 82. https://doi.org/10.3390/jof8010082
APA StyleMoo-Koh, F. A., Cristóbal-Alejo, J., Andrés, M. F., Martín, J., Reyes, F., Tun-Suárez, J. M., & Gamboa-Angulo, M. (2022). In Vitro Assessment of Organic and Residual Fractions of Nematicidal Culture Filtrates from Thirteen Tropical Trichoderma Strains and Metabolic Profiles of Most-Active. Journal of Fungi, 8(1), 82. https://doi.org/10.3390/jof8010082