
Gene Mining for Conserved, Non-Annotated Proteins of Podosphaera xanthii Identifies Novel Target Candidates for Controlling Powdery Mildews by Spray-Induced Gene Silencing

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants, Microbes and Culture Conditions
2.2. Sequence Analysis, Protein Modeling and Protein Function Prediction
2.3. Isolation of Nucleic Acids and cDNA Synthesis
2.4. In Vitro Production of dsRNA
2.5. dsRNA-Mediated Gene Silencing
2.5.1. Leaf Disc Assay
2.5.2. Cotyledon Infiltration Assay
2.6. Quantitative Reverse Transcription RT-qPCR and qPCR
2.7. Haustorial Counts
2.8. SIGS Assay
2.9. Statistical Analysis
3. Results
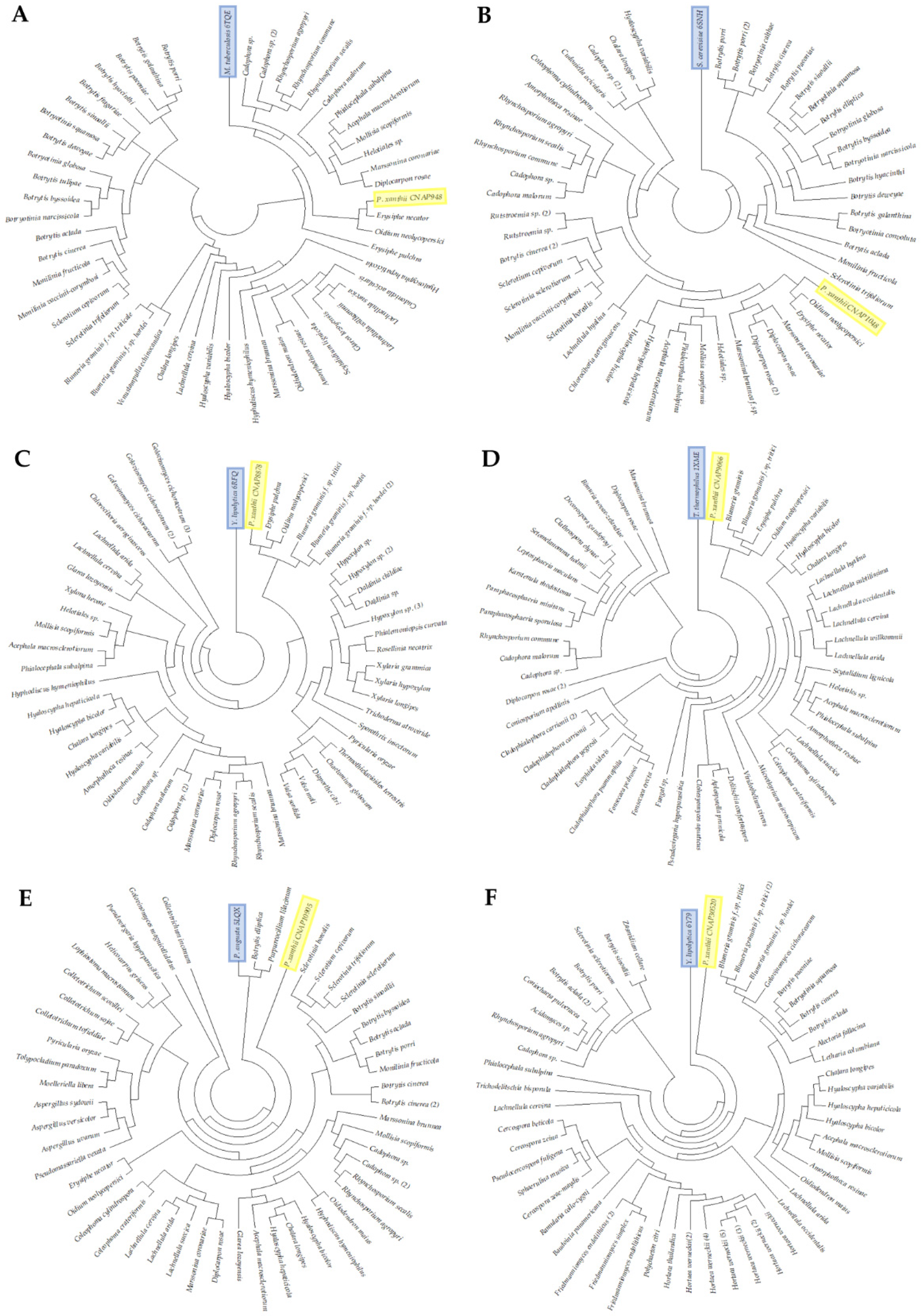
3.1. Identification and Selection of CNAPs
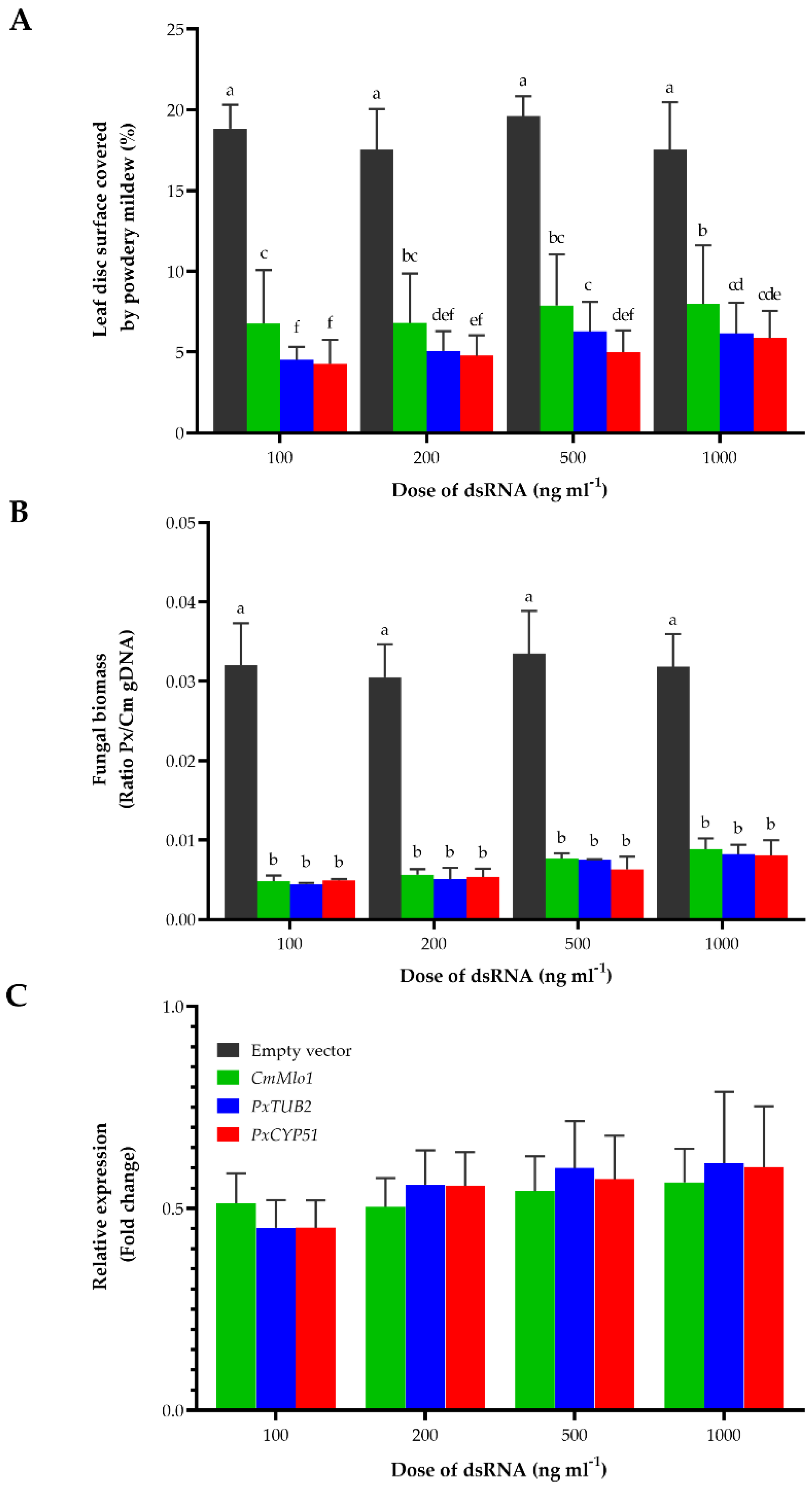
3.2. Development of a dsRNA-Mediated Gene-Silencing Assay for P. xanthii
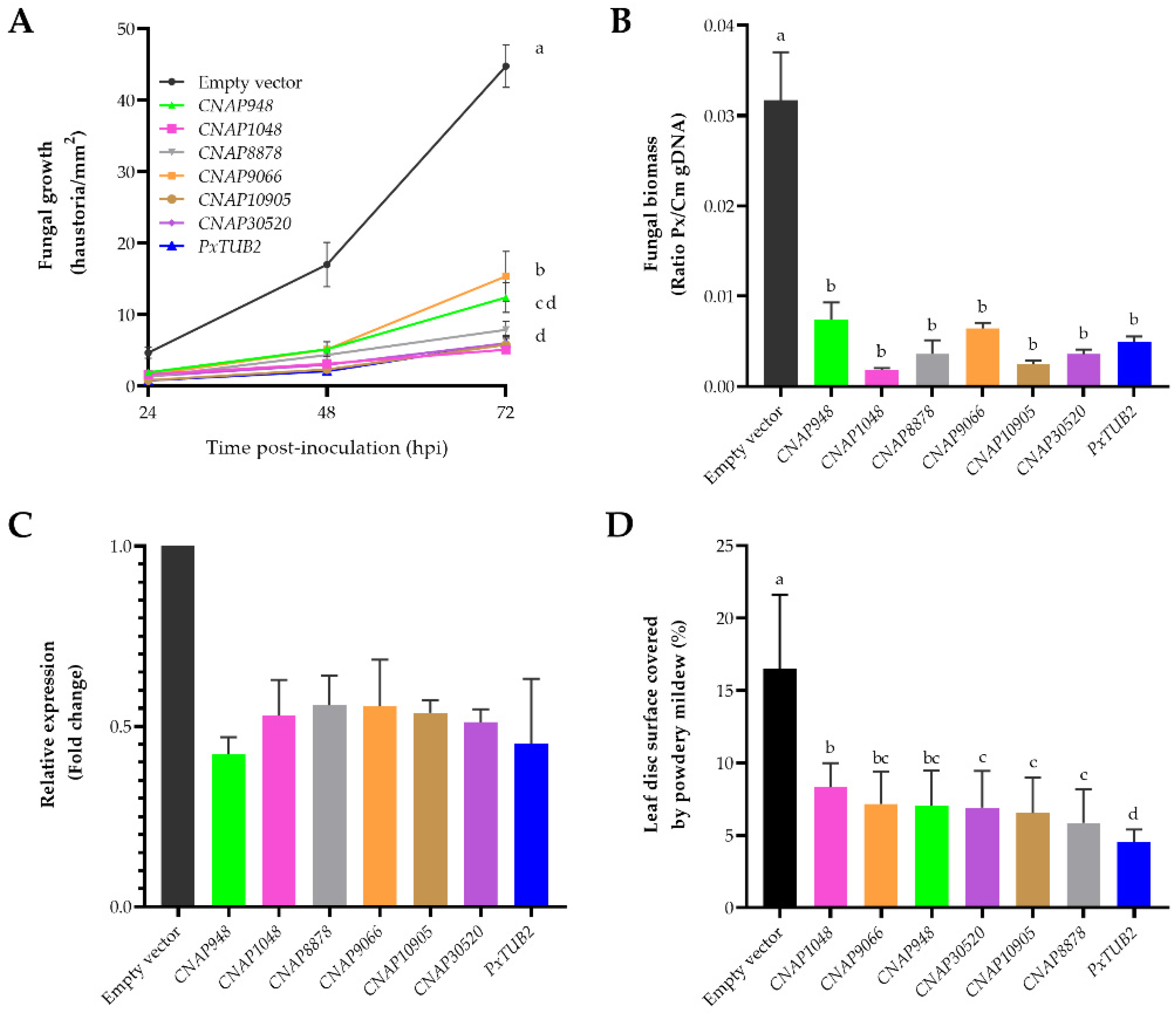
3.3. CNAP948, CNAP1048, CNAP8878, CNAP9066, CNAP10905 and CNAP30520 Are Essential Proteins for P. xanthii
3.4. Efficient Control of Cucurbit Powdery Mildew by SIGS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. Mechanisms of resistance to QoI fungicides in phytopathogenic fungi. Int. Microbiol. 2008, 11, 1–9. [Google Scholar] [CrossRef]
- Micali, C.; Göllner, K.; Humphry, M.; Consonni, C.; Panstruga, R. The powdery mildew disease of Arabidopsis: A paradigm for the interaction between plants and biotrophic fungi. Arab. Book 2008, 6, e0115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Pedersen, C.; Kwaaitaal, M.; Gregersen, P.L.; Mørch, S.M.; Hanisch, S.; Kristensen, A.; Fuglsang, A.T.; Collinge, D.B.; Thordal-Christensen, H. Interaction of barley powdery mildew effector candidate CSEP0055 with the defence protein PR17c. Mol. Plant Pathol. 2012, 13, 1110–1119. [Google Scholar] [CrossRef]
- Martínez-Cruz, J.; Romero, D.; de Vicente, A.; Pérez-García, A. Transformation of the cucurbit powdery mildew pathogen Podosphaera xanthii by Agrobacterium tumefaciens. New Phytol. 2017, 213, 1961–1973. [Google Scholar] [CrossRef] [Green Version]
- Micali, C.O.; Neumann, U.; Grunewald, D.; Panstruga, R.; O’Connell, R. Biogenesis of a specialized plant–fungal interface during host cell internalization of Golovinomyces orontii haustoria. Cell. Microbiol. 2011, 13, 210–226. [Google Scholar] [CrossRef]
- Martínez-Cruz, J.; Romero, D.; Dávila, J.C.; Pérez-García, A. The Podosphaera xanthii haustorium, the fungal Trojan horse of cucurbit-powdery mildew interactions. Fungal Genet. Biol. 2014, 71, 21–31. [Google Scholar] [CrossRef]
- del Pino, D.; Olalla, L.; Pérez-García, A.; Rivera, M.E.; García, S.; Moreno, R.; de Vicente, A.; Torés, J.A. Occurrence of races and pathotypes of cucurbit powdery mildew in southeastern Spain. Phytoparasitica 2002, 30, 459–466. [Google Scholar] [CrossRef]
- Pérez-García, A.; Romero, D.; Fernández-Ortuño, D.; López-Ruiz, F.; de Vicente, A.; Torés, J.A. The powdery mildew fungus Podosphaera fusca (synonym Podosphaera xanthii), a constant threat to cucurbits. Mol. Plant Pathol. 2009, 10, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Bellón-Gómez, D.; Vela-Corcía, D.; Pérez-García, A.; Torés, J.A. Sensitivity of Podosphaera xanthii populations to anti-powdery-mildew fungicides in Spain. Pest Manag. Sci. 2015, 71, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Vielba-Fernández, A.; Polonio, Á.; Ruiz-Jiménez, L.; de Vicente, A.; Pérez-García, A.; Fernández-Ortuño, D. Fungicide resistance in powdery mildew fungi. Microorganisms 2020, 8, 1431. [Google Scholar] [CrossRef]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.H.J.; Huang, H.D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef]
- McLoughlin, A.G.; Wytinck, N.; Walker, P.L.; Girard, I.J.; Rashid, K.Y.; de Kievit, T.; Fernando, W.G.D.; Whyard, S.; Belmonte, M.F. Identification and application of exogenous dsRNA confers plant protection against Sclerotinia sclerotiorum and Botrytis cinerea. Sci. Rep. 2018, 8, 7320. [Google Scholar] [CrossRef]
- Song, X.S.; Gu, K.X.; Duan, X.X.; Xiao, X.M.; Hou, Y.P.; Duan, Y.B.; Wang, J.X.; Yu, N.; Zhou, M.G. Secondary amplification of siRNA machinery limits the application of spray-induced gene silencing. Mol. Plant Pathol. 2018, 19, 2543–2560. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Jin, H. Spray-induced gene silencing: A powerful innovative strategy for crop protection. Trends Microbiol. 2017, 25, 4–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Chen, Z.Y.; Zhang, C.; Ganiger, M. Reduction of Phakopsora pachyrhizi infection on soybean through host- and spray-induced gene silencing. Mol. Plant Pathol. 2020, 21, 794–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, L.; Lan, C.; Capriotti, L.; Ah-Fong, A.; Sanchez, J.N.; Hamby, R.; Heller, J.; Zhao, H.; Glass, N.L.; Judelson, H.S.; et al. Spray-induced gene silencing for disease control is dependent on the efficiency of pathogen RNA uptake. Plant Biotechnol. J. 2021. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Bautista, R.; de Vicente, A.; Spanu, P.D.; Pérez-García, A. De novo analysis of the epiphytic transcriptome of the cucurbit powdery mildew fungus Podosphaera xanthii and identification of candidate secreted effector proteins. PLoS ONE 2016, 11, e0163379. [Google Scholar] [CrossRef] [Green Version]
- De Miccolis Angelini, R.M.; Pollastro, S.; Rotondo, P.R.; Laguardia, C.; Abate, D.; Rotolo, C.; Faretra, F. Transcriptome sequence resource for the cucurbit powdery mildew pathogen Podosphaera xanthii. Sci. Data 2019, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Polonio, Á.; Seoane, P.; Claros, M.G.; Pérez-García, A. The haustorial transcriptome of the cucurbit pathogen Podosphaera xanthii reveals new insights into the biotrophy and pathogenesis of powdery mildew fungi. BMC Genom. 2019, 20, 543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonio, Á.; Díaz-Martínez, L.; Fernández-Ortuño, D.; de Vicente, A.; Romero, D.; López-Ruiz, F.J.; Pérez-García, A. A hybrid genome assembly resource for Podosphaera xanthii, the main causal agent of powdery mildew disease in cucurbits. Mol. Plant-Microbe Interact. 2021, 34, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Subramaniyam, S.; Jung, M.; Oh, E.A.; Kim, T.H.; Kim, J.G. Genome resource of Podosphaera xanthii, the host-specific fungal pathogen that causes cucurbit powdery mildew. Mol. Plant-Microbe Interact. 2021, 34, 457–459. [Google Scholar] [CrossRef]
- Polonio, Á.; Pineda, M.; Bautista, R.; Martínez-Cruz, J.; Pérez-Bueno, M.L.; Barón, M.; Pérez-García, A. RNA-seq analysis and fluorescence imaging of melon powdery mildew disease reveal an orchestrated reprogramming of host physiology. Sci. Rep. 2019, 9, 7978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.L.; Chen, B.H.; Chen, X.J.; Guo, Y.Y.; Yang, H.L.; Li, X.Z.; Wang, G.Y. Transcriptome profiling of pumpkin (Cucurbita moschata Duch.) leaves infected with powdery mildew. PLoS ONE 2018, 13, e0190175. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Gao, P.; Wan, Y.; Cui, H.; Fan, C.; Liu, S.; Luan, F. Comparative transcriptome profiling of genes and pathways related to resistance against powdery mildew in two contrasting melon genotypes. Sci. Hortic. 2018, 227, 169–180. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Romero, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. Transient transformation of Podosphaera xanthii by electroporation of conidia. BMC Microbiol. 2015, 15, 20. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Cruz, J.; Romero, D.; de Vicente, A.; Pérez-García, A. Transformation by growth onto agro-infiltrated tissues (TGAT), a simple and efficient alternative for transient transformation of the cucurbit powdery mildew pathogen Podosphaera xanthii. Mol. Plant Pathol. 2018, 19, 2502–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Cruz, J.; Romero, D.; de la Torre, F.N.; Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. The functional characterization of Podosphaera xanthii candidate effector genes reveals novel target functions for fungal pathogenicity. Mol. Plant-Microbe Interact. 2018, 31, 914–931. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Cruz, J.; Romero, D.; Hierrezuelo, J.; Thon, M.; de Vicente, A.; Pérez-García, A. Effectors with chitinase activity (EWCAs), a family of conserved, secreted fungal chitinases that suppress chitin-triggered immunity. Plant Cell 2021, 33, 1319–1340. [Google Scholar] [CrossRef]
- Polonio, Á.; Fernández-Ortuño, D.; de Vicente, A.; Pérez-García, A. A haustorial-expressed lytic polysaccharide monooxygenase from the cucurbit powdery mildew pathogen Podosphaera xanthii contributes to the suppression of chitin-triggered immunity. Mol. Plant Pathol. 2021, 22, 580–601. [Google Scholar] [CrossRef]
- Álvarez, B.; Torés, J.A. Cultivo in vitro de Sphaerotheca fuliginea (Schlecht, ex Fr.), efecto de diferentes fuentes de carbono sobre su desarrollo. Bol. Sanid. Veg. Plagas 1997, 23, 283–288. [Google Scholar]
- Seoane, P.; Espigares, M.; Carmona, R.; Polonio, Á.; Quintana, J.; Cretazzo, E.; Bota, J.; Pérez-García, A.; de Dios Alché, J.; Gómez, L.; et al. TransFlow: A modular framework for assembling and assessing accurate de novo transcriptomes in non-model organisms. BMC Bioinform. 2018, 19, 416. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef]
- Cortázar, A.R.; Aransay, A.M.; Alfaro, M.; Oguiza, J.A.; Lavín, J.L. SECRETOOL: Integrated secretome analysis tool for fungi. Amino Acids 2014, 46, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Cortazar, A.R.; Oguiza, J.A.; Aransay, A.M.; Lavín, J.L. PECAS: Prokaryotic and eukaryotic classical analysis of secretome. Amino Acids 2015, 47, 2659–2663. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of protein subcellular localization using deep learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef]
- Pierleoni, A.; Martelli, P.; Casadio, R. PredGPI: A GPI-anchor predictor. BMC Bioinform. 2008, 9, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Pagni, M.; Ioannidis, V.; Cerutti, L.; Zahn-Zabal, M.; Jongeneel, C.V.; Hau, J.; Martin, O.; Kuznetsov, D.; Falquet, L. MyHits: Improvements to an interactive resource for analyzing protein sequences. Nucleic Acids Res. 2007, 35, W433–W437. [Google Scholar] [CrossRef]
- Wass, M.N.; Kelley, L.A.; Sternberg, M.J.E. 3DLigandSite: Predicting ligand-binding sites using similar structures. Nucleic Acids Res. 2010, 38, W469–W473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Roy, A.; Zhang, Y. Protein-ligand binding site recognition using complementary binding-specific substructure comparison and sequence profile alignment. Bioinformatics 2013, 29, 2588–2595. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.D.; Dawson, N.L.; Das, S.; Sillitoe, I.; Ashford, P.; Lee, D.; Lehtinen, S.; Orengo, C.A.; Lees, J.G. Gene3D: Expanding the utility of domain assignments. Nucleic Acids Res. 2016, 44, D404–D409. [Google Scholar] [CrossRef] [PubMed]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Ortuño, D.; Pérez-García, A.; López-Ruiz, F.; Romero, D.; de Vicente, A.; Torés, J.A. Occurrence and distribution of resistance to QoI fungicides in populations of Podosphaera fusca in south central Spain. Eur. J. Plant Pathol. 2006, 115, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Thornton, B.; Basu, C. Real-time PCR (qPCR) primer design using free online software. Biochem. Mol. Biol. Edu. 2011, 39, 145–154. [Google Scholar] [CrossRef]
- Vela-Corcía, D.; Bellón-Gómez, D.; López-Ruiz, F.; Torés, J.A.; Pérez-García, A. The Podosphaera fusca TUB2 gene, a molecular “Swiss Army Knife” with multiple applications in powdery mildew research. Fungal Biol. 2014, 118, 228–241. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- FRAC Code List 2021. Available online: https://www.frac.info/docs/default-source/publications/frac-code-list/frac-code-list-2021--final.pdf?sfvrsn=f7ec499a_2 (accessed on 18 June 2021).
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies. Mol. Plant Pathol. 2012, 13, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, L.; Baraldi, E.; Mezzetti, B.; Limera, C.; Sabbadini, S. Biotechnological approaches: Gene overexpression, gene silencing, and genome editing to control fungal and oomycete diseases in grapevine. Int. J. Mol. Sci. 2020, 21, 5701. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Kogel, K.-H. New wind in the sails: Improving the agronomic value of crop plants through RNAi-mediated gene silencing. Plant Biotechnol. J. 2014, 12, 821–831. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.M.; Palmquist, J.; Huang, S.D.; Jin, H. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Kanaya, S.; Uehara, H.; Ikemura, T. A novel bioinformatics strategy for function prediction of poorly-characterized protein genes obtained from metagenome analyses. DNA Res. 2009, 16, 287–297. [Google Scholar] [CrossRef]
- Hu, W.; Yan, L.; Ma, Z. Cloning and expression analysis of a putative ABC transporter gene BgABC1 from the biotrophic pathogenic fungus Blumeria graminis f. sp. tritici. J. Phytopathol. 2008, 156, 120–124. [Google Scholar] [CrossRef]
- Nagashima, Y.; von Schaewen, A.; Koiwa, H. Function of N-glycosylation in plants. Plant Sci. 2018, 274, 70–79. [Google Scholar] [CrossRef]
- Sharma, G.; Aminedi, R.; Saxena, D.; Gupta, A.; Banerjee, P.; Jain, D.; Chandran, D. Effector mining from the Erysiphe pisi haustorial transcriptome identifies novel candidates involved in pea powdery mildew pathogenesis. Mol. Plant Pathol. 2019, 20, 1506–1522. [Google Scholar] [CrossRef] [Green Version]
- Andersson, J.O. Convergent evolution: Gene sharing by eukaryotic plant pathogens. Curr. Biol. 2006, 16, R804–R806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Xiao, G.; Zheng, P.; Cen, K.; Zhan, S.; Wang, C. Divergent and convergent evolution of fungal pathogenicity. Genome Biol. Evol. 2016, 8, 1374–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paldi, F.; Alver, B.; Robertson, D.; Schalbetter, S.A.; Kerr, A.; Kelly, D.A.; Baxter, J.; Neale, M.J.; Marston, A.L. Convergent genes shape budding yeast pericentromeres. Nature 2020, 582, 119–123. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence ID | Protein Name | Protein Length 1 | Subject ID | Description | E-Value | Identity |
|---|---|---|---|---|---|---|
| Pxanthii_948 | CNAP948 | 708 | XP_007290516 | Hypothetical protein Marssonina brunnea f. sp. multigermtubi MB_m1 | 0.0 | 80% (571/711) |
| Pxanthii_1048 | CNAP1048 | 421 | KAE8449429 | Hypothetical protein Helotiales sp. DMI_Dod_QoI | 0.0 | 81% (342/421) |
| Pxanthii_8878 | CNAP8878 | 103 | RKF57155 | Hypothetical protein Oidium neolycopersici | 7 × 10−58 | 81% (83/103) |
| Pxanthii_9066 | CNAP9066 | 122 | CCU82200 | Hypothetical protein Blumeria graminis f. sp. hordei DH14 | 1 × 10−59 | 75% (92/122) |
| Pxanthii_10905 | CNAP10905 | 74 | ESZ98542 | Hypothetical protein Sclerotinia borealis F-4128 | 8 × 10−31 | 77% (51/66) |
| Pxanthii_30520 | CNAP30520 | 80 | TVY46286 | Hypothetical protein Lachnellula occidentalis | 3 × 10−47 | 88% (70/80) |
| Score Values 1 | Structural Analogs 2 | Predicted Features 3 | |||||
|---|---|---|---|---|---|---|---|
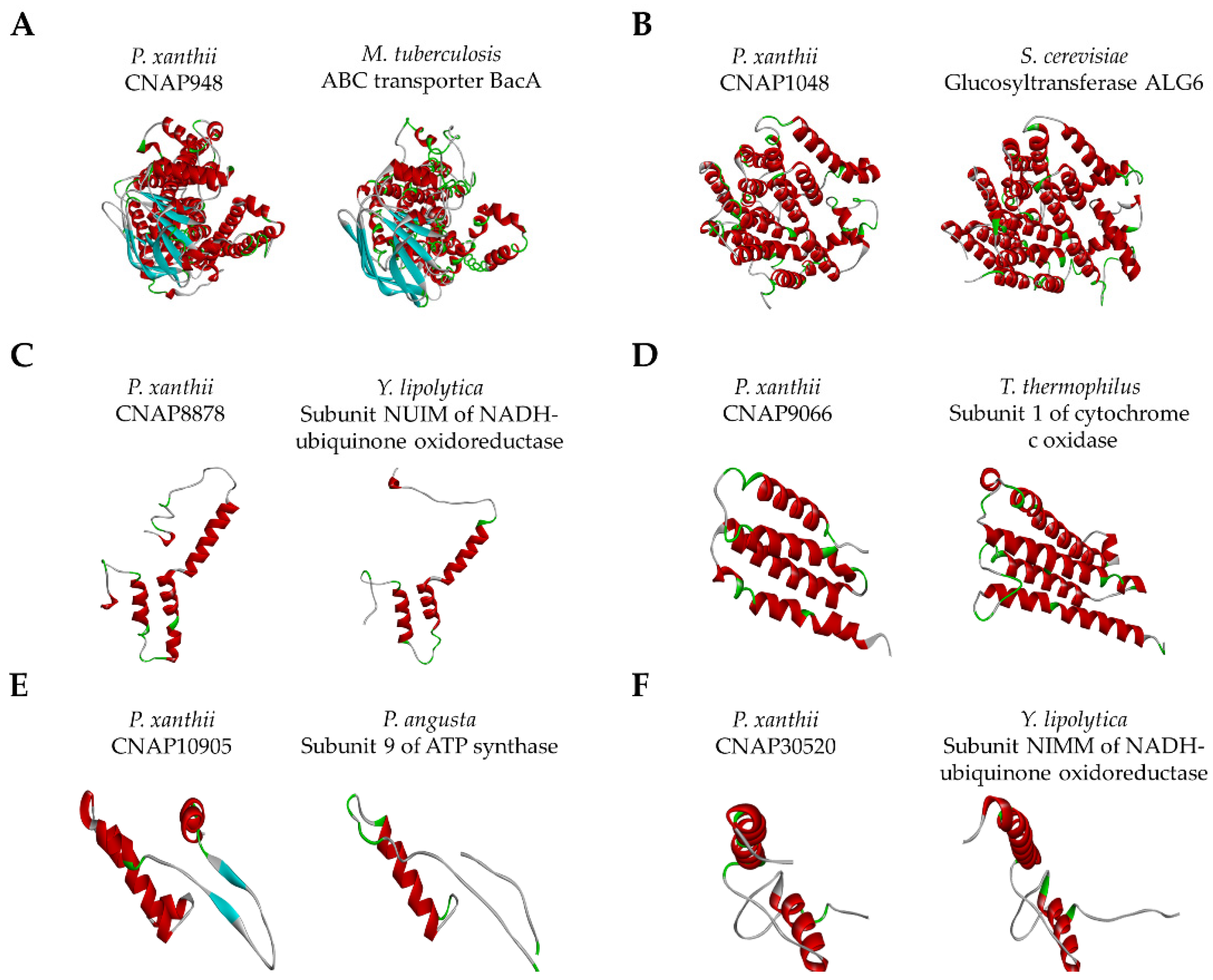
| Protein Model | C Score | TM Score | PDB Code | Species | Activity | Ligands | Putative Function 4 |
| CNAP948 | −0.63 | 0.781 | 6TQE | Mycobacterium tuberculosis | Efflux transporter | ATP | Efflux transport |
| CNAP1048 | −0.82 | 0.873 | 6SNH | Saccharomyces cerevisiae | Glucosyltransferase | Glucose | Glycosylation |
| CNAP8878 | −1.22 | 0.689 | 6RFQ | Yarrowia lipolytica | Oxidoreductase | Iron-sulfur | Respiration |
| CNAP9066 | −1.49 | 0.710 | 1XME | Thermus thermophilus | Oxidoreductase | Copper | Respiration |
| CNAP10905 | 0.37 | 0.624 | 5LQX | Pichia angusta | ATP synthase | ADP | Respiration |
| CNAP30520 | 0.80 | 0.839 | 6Y79 | Yarrowia lipolytica | Oxidoreductase | Iron-sulfur | Respiration |
| Treatment | Dose of dsRNA (µg mL−1) | Leaf Surface Covered by Powdery Mildew (%) 1 | Disease Reduction (%) 2 |
|---|---|---|---|
| Water | - | 59.35 ± 3.78 a | - |
| Empty vector-dsRNA | 30 | 55.60 ± 4.75 a | 6.32 |
| CNAP1048-dsRNA | 30 | 5.62 ± 1.01 ef | 90.53 |
| 20 | 5.25 ± 0.72 ef | 91.15 | |
| 10 | 10.04 ± 0.58 cde | 83.08 | |
| 5 | 21.26 ± 1.28 b | 64.18 | |
| CNAP30520-dsRNA | 30 | 3.92 ± 0.62 f | 93.40 |
| 20 | 3.74 ± 0.46 f | 93.70 | |
| 10 | 9.11 ± 0.78 cdef | 84.65 | |
| 5 | 14.83 ± 1.26 c | 75.01 | |
| CNAP10905-dsRNA | 30 | 3.41 ± 0.28 f | 94.25 |
| 20 | 3.53 ± 0.26 f | 94.05 | |
| 10 | 8.24 ± 1.51 def | 86.12 | |
| 5 | 12.96 ± 1.49 cd | 78.16 | |
| PxTUB2-dsRNA | 30 | 3.86 ± 0.23 f | 93.50 |
| 20 | 4.51 ± 0.45 ef | 92.40 | |
| 10 | 11.94 ± 1.89 cd | 79.88 | |
| 5 | 22.83 ± 0.60 b | 61.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Jiménez, L.; Polonio, Á.; Vielba-Fernández, A.; Pérez-García, A.; Fernández-Ortuño, D. Gene Mining for Conserved, Non-Annotated Proteins of Podosphaera xanthii Identifies Novel Target Candidates for Controlling Powdery Mildews by Spray-Induced Gene Silencing. J. Fungi 2021, 7, 735. https://doi.org/10.3390/jof7090735
Ruiz-Jiménez L, Polonio Á, Vielba-Fernández A, Pérez-García A, Fernández-Ortuño D. Gene Mining for Conserved, Non-Annotated Proteins of Podosphaera xanthii Identifies Novel Target Candidates for Controlling Powdery Mildews by Spray-Induced Gene Silencing. Journal of Fungi. 2021; 7(9):735. https://doi.org/10.3390/jof7090735
Chicago/Turabian StyleRuiz-Jiménez, Laura, Álvaro Polonio, Alejandra Vielba-Fernández, Alejandro Pérez-García, and Dolores Fernández-Ortuño. 2021. "Gene Mining for Conserved, Non-Annotated Proteins of Podosphaera xanthii Identifies Novel Target Candidates for Controlling Powdery Mildews by Spray-Induced Gene Silencing" Journal of Fungi 7, no. 9: 735. https://doi.org/10.3390/jof7090735