
The Role of the Cell Integrity Pathway in Septum Assembly in Yeast


{kind=link}
{kind=link}
Abstract
1. Maintaining the Shape: The Cell Integrity Signaling Pathways
2. S. pombe, the Fission Yeast
2.1. Before Septum Assembly: Actomyosin Ring Positioning and Assembly
2.2. Anchoring the Ring to the Plasma Membrane and the Growing Septum
2.3. Triggering Septation: The Role of the Septation Initiation Network (SIN) in CAR Maintenance and Constriction
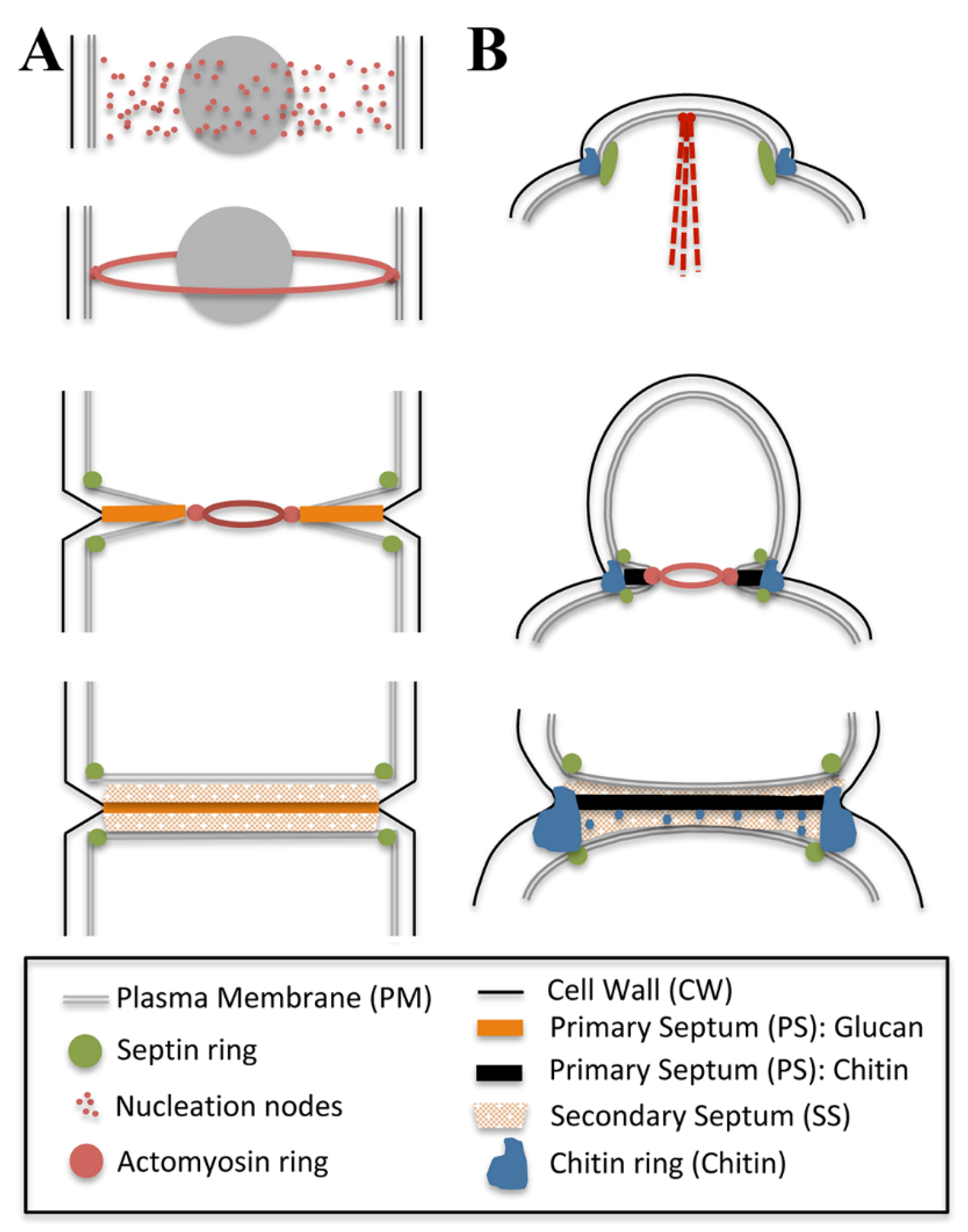
2.4. Furrow Ingression and Septum Deposition
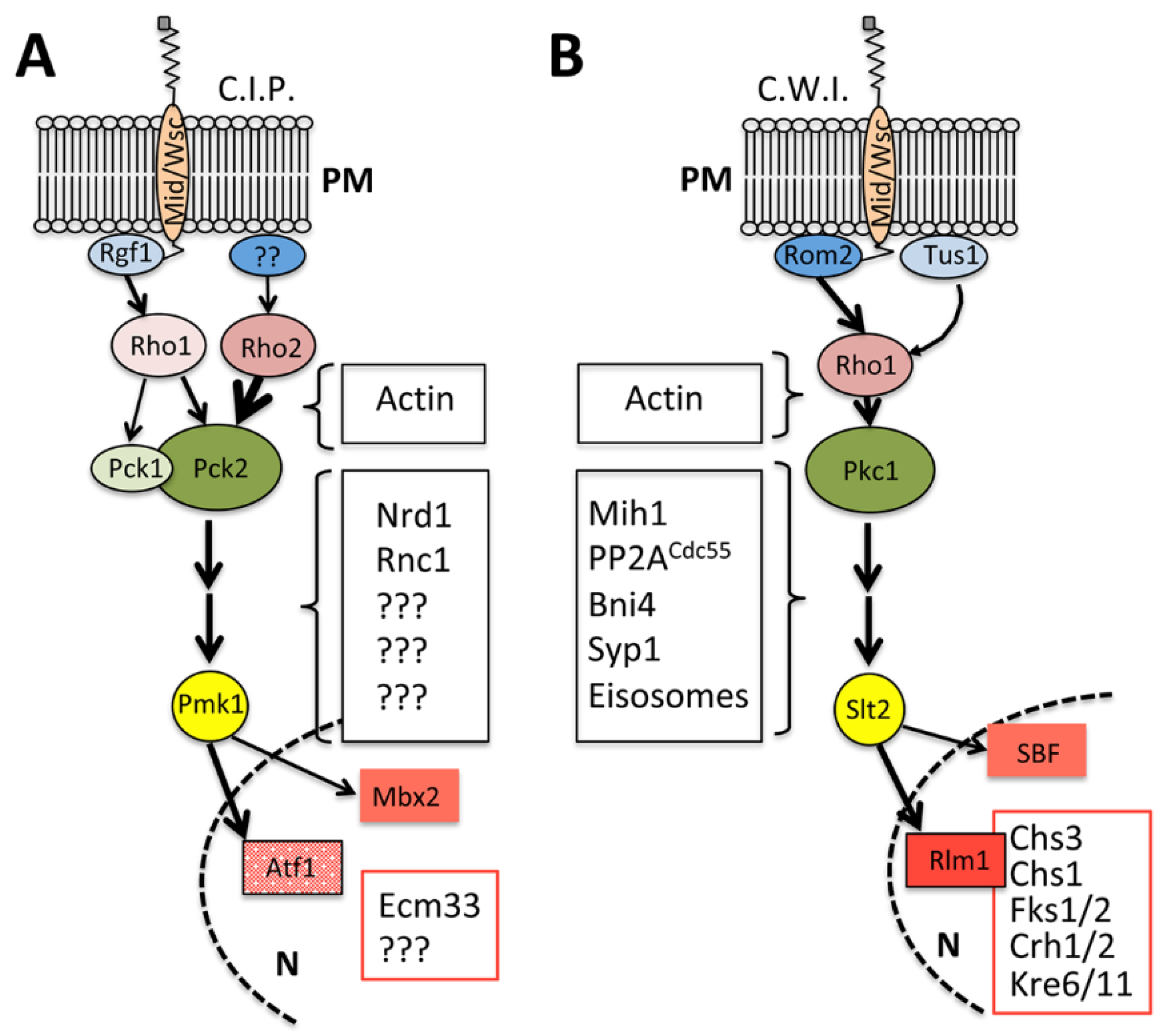
2.5. Integrating Septum Assembly and the Cell Integrity Pathway during Cytokinesis in Fission Yeast
2.6. The Role of the CIP beyond Septum Assembly
3. S. cerevisiae, the Budding Yeast
3.1. Before Septum Assembly: Shaping the Yeast Cell through the Synthesis and Assembly of the Cell Wall
3.2. Starting the Separation: The Building of a Primary Septum
3.3. At the End of the Process, Secondary Septa Synthesis and Cell Separation
3.4. Beyond Septal Assembly: The Generation of a Remedial Septum
3.5. The Action of CWI beyond Septum Assembly
4. Concluding Remarks and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Elhasi, T.; Blomberg, A. Integrins in disguise—Mechanosensors in Saccharomyces cerevisiae as functional integrin analogues. Microb. Cell 2019, 6, 335–355. [Google Scholar] [CrossRef] [PubMed]
- Kock, C.; Dufrene, Y.F.; Heinisch, J.J. Up against the wall: Is yeast cell wall integrity ensured by mechanosensing in plasma membrane microdomains? Appl. Environ. Microbiol. 2015, 81, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef]
- Perez, P.; Cortes, J.C.G.; Cansado, J.; Ribas, J.C. Fission yeast cell wall biosynthesis and cell integrity signalling. Cell Surf. 2018, 4, 1–9. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Soto, T.; Madrid, M.; Núñez, A.; Vicente, J.; Gacto, M.; Cansado, J. Activation of the cell integrity pathway is channelled through diverse signalling elements in fission yeast. Cell. Signal. 2008, 20, 748–757. [Google Scholar] [CrossRef]
- Sánchez-Mir, L.; Soto, T.; Franco, A.; Madrid, M.; Viana, R.A.; Vicente, J.; Gacto, M.; Perez, P.; Cansado, J. Rho1 GTPase and PKC Ortholog Pck1 Are Upstream Activators of the Cell Integrity MAPK Pathway in Fission Yeast. PLoS ONE 2014, 9, e88020. [Google Scholar] [CrossRef]
- Garcia, R.; Bermejo, C.; Grau, C.; Pérez, R.; Peña, J.M.R.; François, J.M.; Nombela, C.; Arroyo, J. The Global Transcriptional Response to Transient Cell Wall Damage in Saccharomyces cerevisiae and Its Regulation by the Cell Integrity Signaling Pathway. J. Biol. Chem. 2004, 279, 15183–15195. [Google Scholar] [CrossRef]
- Lagorce, A.; Hauser, N.C.; Labourdette, D.; Rodriguez, C.; Martin-Yken, H.; Arroyo, J.; Hoheisel, J.D.; François, J.M. Genome-wide Analysis of the Response to Cell Wall Mutations in the Yeast Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 20345–20357. [Google Scholar] [CrossRef]
- Madden, K.; Snyder, M. Cell polarity and morphogenesis in budding yeast. Annu. Rev. Microbiol. 1998, 52, 687–744. [Google Scholar] [CrossRef]
- Pollard, T.D.; O’Shaughnessy, B. Molecular Mechanism of Cytokinesis. Annu. Rev. Biochem. 2019, 88, 661–689. [Google Scholar] [CrossRef]
- Gerien, K.S.; Wu, J.Q. Molecular mechanisms of contractile-ring constriction and membrane trafficking in cytokinesis. Biophys. Rev. 2018, 10, 1649–1666. [Google Scholar] [CrossRef] [PubMed]
- Mangione, M.C.; Gould, K.L. Molecular form and function of the cytokinetic ring. J. Cell Sci. 2019, 132, 12. [Google Scholar] [CrossRef]
- Rincon, S.A.; Paoletti, A. Molecular control of fission yeast cytokinesis. Semin. Cell Dev. Biol. 2016, 53, 28–38. [Google Scholar] [CrossRef]
- Cortés, J.C.G.; Ramos, M.; Osumi, M.; Pérez, P.; Ribas, J.C. Fission yeast septation. Commun. Integr. Biol. 2016, 9, e1189045. [Google Scholar] [CrossRef]
- Pérez, P.; Cortés, J.C.G.; Martín-García, R.; Ribas, J.C. Overview of fission yeast septation. Cell. Microbiol. 2016, 18, 1201–1207. [Google Scholar] [CrossRef]
- Martín-García, R.; Santos, B. The price of independence: Cell separation in fission yeast. World J. Microbiol. Biotechnol. 2016, 32, 1–7. [Google Scholar] [CrossRef]
- Cheffings, T.H.; Burroughs, N.J.; Balasubramanian, M.K. Actomyosin Ring Formation and Tension Generation in Eukaryotic Cytokinesis. Curr. Biol. 2016, 26, R719–R737. [Google Scholar] [CrossRef]
- Akamatsu, M.; Berro, J.; Pu, K.-M.; Tebbs, I.R.; Pollard, T.D. Cytokinetic nodes in fission yeast arise from two distinct types of nodes that merge during interphase. J. Cell Biol. 2014, 204, 977–988. [Google Scholar] [CrossRef]
- Steever, A.; Pringle, J.; Bähler, J.; Wang, Y.-L.; Gould, K.; McCollum, D.; Wheatley, S. Role of Polo Kinase and Mid1p in Determining the Site of Cell Division in Fission Yeast. J. Cell Biol. 1998, 143, 1603–1616. [Google Scholar] [CrossRef]
- Mccollum, D. Cytokinesis: ER keeps Mid1 in the middle. Curr. Biol. 2010, 20, R484–R486. [Google Scholar] [CrossRef]
- Magliozzi, J.O.; Sears, J.; Cressey, L.; Brady, M.; Opalko, H.E.; Kettenbach, A.N.; Moseley, J.B. Fission yeast Pak1 phosphorylates anillin-like Mid1 for spatial control of cytokinesis. J. Cell Biol. 2020, 219, 8. [Google Scholar] [CrossRef]
- Willet, A.H.; McDonald, N.A.; Gould, K.L. Regulation of contractile ring formation and septation in Schizosaccharomyces pombe. Curr. Opin. Microbiol. 2015, 28, 46–52. [Google Scholar] [CrossRef]
- Roberts-Galbraith, R.H.; Chen, J.S.; Wang, J.; Gould, K.L. The SH3 domains of two PCH family members cooperate in assembly of the Schizosaccharomyces pombe contractile ring. J. Cell Biol. 2009, 184, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Laplante, C.; Berro, J.; Karatekin, E.; Hernandez-Leyva, A.; Lee, R.; Pollard, T.D. Three Myosins Contribute Uniquely to the Assembly and Constriction of the Fission Yeast Cytokinetic Contractile Ring. Curr. Biol. 2015, 25, 1955–1965. [Google Scholar] [CrossRef]
- Arasada, R.; Pollard, T.D. A role for F-BAR protein Rga7p during cytokinesis in S. pombe. J. Cell Sci. 2015, 128, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Goss, J.W.; Kim, S.; Bledsoe, H.; Pollard, T.D. Characterization of the roles of Blt1p in fission yeast cytokinesis. Mol. Biol. Cell 2014, 25, 1946–1957. [Google Scholar] [CrossRef]
- McDonald, N.A.; Lind, A.L.; Smith, S.E.; Li, R.; Gould, K.L. Nanoscale architecture of the Schizosaccharomyces pombe contractile ring. eLife 2017, 6, e28865. [Google Scholar] [CrossRef]
- Sun, L.; Guan, R.; Lee, I.-J.; Liu, Y.; Chen, M.; Wang, J.; Wu, J.-Q.; Chen, Z. Mechanistic Insights into the Anchorage of the Contractile Ring by Anillin and Mid1. Dev. Cell 2015, 33, 413–426. [Google Scholar] [CrossRef]
- Snider, C.E.; Willet, A.H.; Chen, J.-S.; Arpağ, G.; Zanic, M.; Gould, K.L. Phosphoinositide-mediated ring anchoring resists perpendicular forces to promote medial cytokinesis. J. Cell Biol. 2017, 216, 3041–3050. [Google Scholar] [CrossRef]
- Arasada, R.; Pollard, T.D. Contractile Ring Stability in S. pombe Depends on F-BAR Protein Cdc15p and Bgs1p Transport from the Golgi Complex. Cell Rep. 2014, 8, 1533–1544. [Google Scholar] [CrossRef]
- McDonald, N.A.; Kooi, C.W.V.; Ohi, M.D.; Gould, K.L. Oligomerization but Not Membrane Bending Underlies the Function of Certain F-BAR Proteins in Cell Motility and Cytokinesis. Dev. Cell 2015, 35, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Cortés, J.C.G.; Pujol, N.; Sato, M.; Pinar, M.; Ramos, M.; Moreno, M.B.; Osumi, M.; Ribas, J.; Perez, P. Cooperation between Paxillin-like Protein Pxl1 and Glucan Synthase Bgs1 Is Essential for Actomyosin Ring Stability and Septum Formation in Fission Yeast. PLoS Genet. 2015, 11, e1005358. [Google Scholar] [CrossRef]
- Mishra, M.; Huang, Y.; Srivastava, P.; Srinivasan, R.; Sevugan, M.; Shlomovitz, R.; Gov, N.; Rao, M.; Balasubramanian, M. Cylindrical Cellular Geometry Ensures Fidelity of Division Site Placement in Fission Yeast. J. Cell Sci. 2012, 125, 3850–3857. [Google Scholar] [CrossRef]
- Muñoz, J.; Cortes, J.G.; Sipiczki, M.; Ramos, M.; Clemente-Ramos, J. Ángel; Moreno, M.B.; Martins, I.; Perez, P.; Ribas, J.C. Extracellular cell wall β(1,3)glucan is required to couple septation to actomyosin ring contraction. J. Cell Biol. 2013, 203, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Pardo, M.; Nurse, P. Equatorial retention of the contractile actin ring by microtubules during cytokinesis. Science 2003, 300, 1569–1574. [Google Scholar] [CrossRef] [PubMed]
- Venkatram, S.; Jennings, J.L.; Link, A.; Gould, K.L. Mto2p, a novel fission yeast protein required for cyto-plasmic microtubule organization and anchoring of the cytokinetic actin ring. Mol. Biol. Cell 2005, 16, 3052–3063. [Google Scholar] [CrossRef]
- Goyal, A.; Takaine, M.; Simanis, V.; Nakano, K. Dividing the spoils of growth and the cell cycle: The fission yeast as a model for the study of cytokinesis. Cytoskeleton 2011, 68, 69–88. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.E.; Mccollum, D.; Gould, K.L. Polar opposites: Fine-tuning cytokinesis through SIN asymmetry. Cytoskeleton 2012, 69, 686–699. [Google Scholar] [CrossRef]
- Simanis, V. Pombe’s thirteen—Control of fission yeast cell division by the septation initiation network. J. Cell Sci. 2015, 128, 1465–1474. [Google Scholar] [CrossRef]
- Baro, B.; Queralt, E.; Monje-Casas, F. Regulation of Mitotic Exit in Saccharomyces cerevisiae. Methods Mol. Biol. 2017, 1505, 3–17. [Google Scholar] [CrossRef]
- Ma, S.; Meng, Z.; Chen, R.; Guan, K.-L. The Hippo Pathway: Biology and Pathophysiology. Annu. Rev. Biochem. 2019, 88, 577–604. [Google Scholar] [CrossRef]
- Hachet, O.; Simanis, V. Mid1p/anillin and the septation initiation network orchestrate contractile ring assembly for cytokinesis. Genes Dev. 2008, 22, 3205–3216. [Google Scholar] [CrossRef]
- Schmidt, S.; Sohrmann, M.; Hofmann, K.; Woollard, A.; Simanis, V. The Spg1p GTPase is an essential, dosage-dependent inducer of septum formation in Schizosaccharomyces pombe. Genes Dev. 1997, 11, 1519–1534. [Google Scholar] [CrossRef]
- Sparks, C.A.; Morphew, M.; McCollum, D. Sid2p, a spindle pole body kinase that regulates the onset of cytokinesis. J. Cell Biol. 1999, 146, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.X.; Domingo-Sananes, M.R.; Huzarska, M.; Novak, B.; Gould, K.L. Multisite phosphoregulation of Cdc25 activity refines the mitotic entrance and exit switches. Proc. Natl. Acad. Sci. USA 2012, 109, 9899–9904. [Google Scholar] [CrossRef]
- Roberts-Galbraith, R.H.; Ohi, M.D.; Ballif, B.A.; Chen, J.S.; McLeod, I.; McDonald, W.H.; Gygi, S.P.; Yates, J.R., 3rd; Gould, K.L. Dephosphorylation of F-BAR protein Cdc15 modulates its conformation and stimulates its scaffolding activity at the cell division site. Mol. Cell 2010, 39, 86–99. [Google Scholar] [CrossRef]
- Bohnert, K.A.; Grzegorzewska, A.P.; Willet, A.; Kooi, C.W.V.; Kovar, D.R.; Gould, K.L. SIN-dependent phosphoinhibition of formin multimerization controls fission yeast cytokinesis. Genes Dev. 2013, 27, 2164–2177. [Google Scholar] [CrossRef]
- Rincon, S.A.; Estravis, M.; Dingli, F.; Loew, D.; Tran, P.T.; Paoletti, A. SIN-Dependent Dissociation of the SAD Kinase Cdr2 from the Cell Cortex Resets the Division Plane. Curr. Biol. 2017, 27, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Willet, A.; DeWitt, A.K.; Beckley, J.R.; Clifford, D.M.; Gould, K.L. NDR Kinase Sid2 Drives Anillin-like Mid1 from the Membrane to Promote Cytokinesis and Medial Division Site Placement. Curr. Biol. 2019, 29, 1055–1063. [Google Scholar] [CrossRef]
- Gavilan, M.A.; Lahoz, A.; Daga, R.; Jimenez, J. Feedback Regulation of SIN by Etd1 and Rho1 in Fission Yeast. Genetics 2014, 196, 455–470. [Google Scholar] [CrossRef]
- Jin, Q.-W.; Zhou, M.; Bimbo, A.; Balasubramanian, M.; McCollum, D. A Role for the Septation Initiation Network in Septum Assembly Revealed by Genetic Analysis of sid2-250 Suppressors. Genetics 2006, 172, 2101–2112. [Google Scholar] [CrossRef]
- Gupta, S.; Mana-Capelli, S.; McLean, J.R.; Chen, C.-T.; Ray, S.; Gould, K.; McCollum, D. Identification of SIN Pathway Targets Reveals Mechanisms of Crosstalk between NDR Kinase Pathways. Curr. Biol. 2013, 23, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Thiyagarajan, S.; Munteanu, E.L.; Arasada, R.; Pollard, T.D.; O’Shaughnessy, B. The fission yeast cytokinetic contractile ring regulates septum shape and closure. J. Cell Sci. 2015, 128, 3672–3681. [Google Scholar] [CrossRef]
- Zhou, Z.; Munteanu, E.L.; He, J.; Ursell, T.; Bathe, M.; Huang, K.C.; Chang, F. The contractile ring coordinates curvature-dependent septum assembly during fission yeast cytokinesis. Mol. Biol. Cell 2015, 26, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Tebbs, I.R.; Pollard, T.D. Separate roles of IQGAP Rng2p in forming and constricting the Schizosaccharomyces pombe cytokinetic contractile ring. Mol. Biol. Cell 2013, 24, 1904–1917. [Google Scholar] [CrossRef]
- Chang, F. Forces that shape fission yeast cells. Mol. Biol. Cell 2017, 28, 1819–1824. [Google Scholar] [CrossRef]
- Proctor, S.A.; Minc, N.; Boudaoud, A.; Chang, F. Contributions of Turgor Pressure, the Contractile Ring, and Septum Assembly to Forces in Cytokinesis in Fission Yeast. Curr. Biol. 2012, 22, 1601–1608. [Google Scholar] [CrossRef]
- Vjestica, A.; Tang, X.Z.; Oliferenko, S. The actomyosin ring recruits early secretory compartments to the division site in fission yeast. Mol. Biol. Cell 2008, 19, 1125–1138. [Google Scholar] [CrossRef]
- Wang, N.; Lee, I.J.; Rask, G.; Wu, J.Q. Roles of the TRAPP-II Complex and the Exocyst in Membrane Deposition during Fission Yeast Cytokinesis. PLoS Biol. 2016, 14, e1002437. [Google Scholar] [CrossRef]
- Zhu, Y.; Hyun, J.; Pan, Y.-Z.; Hopper, J.E.; Rizo, J.; Wu, J.-Q. Roles of the fission yeast UNC-13/Munc13 protein Ync13 in late stages of cytokinesis. Mol. Biol. Cell 2018, 29, 2259–2279. [Google Scholar] [CrossRef]
- Frémont, S.; Echard, A. Membrane Traffic in the Late Steps of Cytokinesis. Curr. Biol. 2018, 28, R458–R470. [Google Scholar] [CrossRef] [PubMed]
- Smertenko, A. Phragmoplast expansion: The four-stroke engine that powers plant cytokinesis. Curr. Opin. Plant Biol. 2018, 46, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.; Cortés, J.C.G.; Sato, M.; Rincón, S.A.; Moreno, M.B.; Clemente-Ramos, J.Á.; Osumi, M.; Pérez, P.; Ribas, J.C. Two S. pombe septation phases differ in ingression rate, septum structure, and response to F-actin loss. J. Cell Biol. 2019, 218, 4171–4194. [Google Scholar] [CrossRef] [PubMed]
- Cortes, J.C.; Konomi, M.; Martins, I.M.; Munoz, J.; Moreno, M.B.; Osumi, M.; Duran, A.; Ribas, J.C. The (1,3)beta-D-glucan synthase subunit Bgs1p is responsible for the fission yeast primary septum formation. Mol. Microbiol. 2007, 65, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Humbel, B.M.; Konomi, M.; Takagi, T.; Kamasawa, N.; Ishijima, S.A.; Osumi, M. In situ localization of beta-glucans in the cell wall of Schizosaccharomyces pombe. Yeast 2001, 18, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Garcia Cortes, J.C.; Ramos, M.; Osumi, M.; Perez, P.; Ribas, J.C. The Cell Biology of Fission Yeast Septation. Microbiol. Mol. Biol. Rev. 2016, 80, 779–791. [Google Scholar] [CrossRef]
- Garcia, P.; Garcia, I.; Marcos, F.; de Garibay, G.R.; Sanchez, Y. Fission yeast rgf2p is a rho1p guanine nucleotide exchange factor required for spore wall maturation and for the maintenance of cell integrity in the absence of rgf1p. Genetics 2009, 181, 1321–1334. [Google Scholar] [CrossRef][Green Version]
- García, P.; Tajadura, V.; García, I.; Sanchez, Y. Role of Rho GTPases and Rho-GEFs in the regulation of cell shape and integrity in fission yeast. Yeast 2006, 23, 1031–1043. [Google Scholar] [CrossRef]
- Tajadura, V.; Garcia, B.; Garcia, I.; Garcia, P.; Sanchez, Y. Schizosaccharomyces pombe Rgf3p is a specific Rho1 GEF that regulates cell wall beta-glucan biosynthesis through the GTPase Rho1p. J. Cell Sci. 2004, 117 Pt 25, 6163–6174. [Google Scholar] [CrossRef]
- Davidson, R.; Laporte, D.; Wu, J.-Q. Regulation of Rho-GEF Rgf3 by the arrestin Art1 in fission yeast cytokinesis. Mol. Biol. Cell 2015, 26, 453–466. [Google Scholar] [CrossRef]
- Ren, L.; Willet, A.H.; Roberts-Galbraith, R.H.; McDonald, N.; Feoktistova, A.; Chen, J.-S.; Huang, H.; Guillen, R.; Boone, C.; Sidhu, S.S.; et al. The Cdc15 and Imp2 SH3 domains cooperatively scaffold a network of proteins that redundantly ensure efficient cell division in fission yeast. Mol. Biol. Cell 2015, 26, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Edreira, T.; Celador, R.; Manjon, E.; Sanchez, Y. A novel checkpoint pathway controls actomyosin ring constriction trigger in fission yeast. eLife 2020, 9, e59333. [Google Scholar] [CrossRef]
- Davi, V.; Tanimoto, H.; Ershov, D.; Haupt, A.; De Belly, H.; Le Borgne, R.; Couturier, E.; Boudaoud, A.; Minc, N. Mechanosensation Dynamically Coordinates Polar Growth and Cell Wall Assembly to Promote Cell Survival. Dev. Cell 2018, 45, 170–182. [Google Scholar] [CrossRef]
- Garcia, P.; Tajadura, V.; Sanchez, Y. The Rho1p exchange factor Rgf1p signals upstream from the Pmk1 mitogen-activated protein kinase pathway in fission yeast. Mol. Biol. Cell 2009, 20, 721–731. [Google Scholar] [CrossRef][Green Version]
- Dekker, N.; Speijer, D.; Grun, C.H.; van den Berg, M.; de Haan, A.; Hochstenbach, F. Role of the alpha-glucanase Agn1p in fission-yeast cell separation. Mol. Biol. Cell 2004, 15, 3903–3914. [Google Scholar] [CrossRef]
- García, I.; Jiménez, D.; Martín, V.; Durán, A.; Sánchez, Y. The α-glucanase Agn1p is required for cell separation in Schizosaccharomyces pombe. Biol. Cell 2005, 97, 569–576. [Google Scholar] [CrossRef]
- Martin-Cuadrado, A.B.; Duenas, E.; Sipiczki, M.; Vazquez de Aldana, C.R.; del Rey, F. The en-do-beta-1,3-glucanase eng1p is required for dissolution of the primary septum during cell separation in Schizosaccharomyces pombe. J. Cell Sci. 2003, 116 Pt 9, 1689–1698. [Google Scholar] [CrossRef]
- Koyano, T.; Kume, K.; Konishi, M.; Toda, T.; Hirata, D. Search for Kinases Related to Transition of Growth Polarity in Fission Yeast. Biosci. Biotechnol. Biochem. 2010, 74, 1129–1133. [Google Scholar] [CrossRef]
- Bohnert, K.A.; Gould, K.L. Cytokinesis-Based Constraints on Polarized Cell Growth in Fission Yeast. PLoS Genet. 2012, 8, e1003004. [Google Scholar] [CrossRef]
- Sengar, A.; Markley, N.A.; Marini, N.J.; Young, D. Mkh1, a MEK kinase required for cell wall integrity and proper response to osmotic and temperature stress in Schizosaccharomyces pombe. Mol. Cell. Biol. 1997, 17, 3508–3519. [Google Scholar] [CrossRef]
- Toda, T.; Dhut, S.; Superti-Furga, G.; Gotoh, Y.; Nishida, E.; Sugiura, R.; Kuno, T. The fission yeast pmk1+ gene encodes a novel mitogen-activated protein kinase homolog which regulates cell integrity and functions coordinately with the Protein Kinase C pathway. Mol. Cell. Biol. 1996, 16, 6752–6764. [Google Scholar] [CrossRef]
- Zaitsevskaya-Carter, T.; Cooper, J.A. Spm1, a stress-activated MAP kinase that regulates morphogenesis in S.pombe. EMBO J. 1997, 16, 1318–1331. [Google Scholar] [CrossRef]
- Wilkinson, M.G.; Samuels, M.; Takeda, T.; Toone, W.M.; Shieh, J.C.; Toda, T.; Millar, J.; Jones, N. The Atf1 transcription factor is a target for the Sty1 stress-activated MAP kinase pathway in fission yeast. Genes Dev. 1996, 10, 2289–2301. [Google Scholar] [CrossRef]
- Takada, H.; Nishida, A.; Domae, M.; Kita, A.; Yamano, Y.; Uchida, A.; Ishiwata, S.; Fang, Y.; Zhou, X.; Masuko, T.; et al. The cell surface protein gene ecm33+ is a target of the two transcription factors Atf1 and Mbx1 and negatively regulates Pmk1 MAPK cell integrity signaling in fission yeast. Mol. Biol. Cell 2010, 21, 674–685. [Google Scholar] [CrossRef]
- Takada, H.; Nishimura, M.; Asayama, Y.; Mannse, Y.; Ishiwata, S.; Kita, A.; Doi, A.; Nishida, A.; Kai, N.; Moriuchi, S.; et al. Atf1 Is a Target of the MAP Kinase Pmk1 and Regulates Cell Integrity in Fission Yeast. Mol. Biol. Cell 2007, 18, 4794–4802. [Google Scholar] [CrossRef]
- Levin, D.E. Cell Wall Integrity Signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 262–291. [Google Scholar] [CrossRef]
- Madrid, M.; Nunez, A.; Soto, T.; Vicente-Soler, J.; Gacto, M.; Cansado, J. Stress-activated protein kinase-mediated down-regulation of the cell integrity pathway mitogen-activated protein kinase Pmk1p by protein phosphatases. Mol. Biol. Cell 2007, 18, 4405–4419. [Google Scholar] [CrossRef]
- Madrid, M.; Soto, T.; Khong, H.K.; Franco, A.; Vicente, J.; Perez, P.; Gacto, M.; Cansado, J. Stress-induced response, localization, and regulation of the Pmk1 cell integrity pathway in Schizosaccharomyces pombe. J. Biol. Chem. 2006, 281, 2033–2043. [Google Scholar] [CrossRef]
- Sanchez-Mir, L.; Franco, A.; Martin-Garcia, R.; Madrid, M.; Vicente-Soler, J.; Soto, T.; Gacto, M.; Perez, P.; Cansado, J. Rho2 Palmitoylation Is Required for Plasma Membrane Localization and Proper Signaling to the Fission Yeast Cell Integrity Mitogen-Activated Protein Kinase Pathway. Mol. Cell. Biol. 2014, 34, 2745–2759. [Google Scholar] [CrossRef]
- Satoh, R.; Morita, T.; Takada, H.; Kita, A.; Ishiwata, S.; Doi, A.; Hagihara, K.; Taga, A.; Matsumura, Y.; Tohda, H.; et al. Role of the RNA-binding Protein Nrd1 and Pmk1 Mitogen-activated Protein Kinase in the Regulation of Myosin mRNA Stability in Fission Yeast. Mol. Biol. Cell 2009, 20, 2473–2485. [Google Scholar] [CrossRef] [PubMed]
- Bähler, J. A Transcriptional Pathway for Cell Separation in Fission Yeast. Cell Cycle 2004, 4, 39–41. [Google Scholar] [CrossRef][Green Version]
- Bi, E.; Park, H.-O. Cell Polarization and Cytokinesis in Budding Yeast. Genetics 2012, 191, 347–387. [Google Scholar] [CrossRef]
- Howell, A.S.; Lew, D.J. Morphogenesis and the Cell Cycle. Genetics 2012, 190, 51–77. [Google Scholar] [CrossRef] [PubMed]
- Wloka, C.; Nishihama, R.; Onishi, M.; Oh, Y.; Hanna, J.; Pringle, J.R.; Krauss, M.; Bi, E. Evidence that a septin diffusion barrier is dispensable for cytokinesis in budding yeast. Biol. Chem. 2011, 392, 813–829. [Google Scholar] [CrossRef]
- Farkašovský, M. Septin architecture and function in budding yeast. Biol. Chem. 2020, 401, 903–919. [Google Scholar] [CrossRef]
- Spiliotis, E.T.; McMurray, M.A. Masters of asymmetry—Lessons and perspectives from 50 years of septins. Mol. Biol. Cell 2020, 31, 2289–2297. [Google Scholar] [CrossRef]
- Miller, K.E.; Kang, P.J.; Park, H.-O. Regulation of Cdc42 for polarized growth in budding yeast. Microb. Cell 2020, 7, 175–189. [Google Scholar] [CrossRef]
- Drgonova, J.; Drgon, T.; Tanaka, K.; Kollár, R.; Chen, G.-C.; Ford, R.A.; Chan, C.S.M.; Takai, Y.; Cabib, E. Rho1p, a Yeast Protein at the Interface between Cell Polarization and Morphogenesis. Science 1996, 272, 277–279. [Google Scholar] [CrossRef]
- Qadota, H.; Ishii, I.; Fujiyama, A.; Ohya, Y.; Anraku, Y. RHO gene products, putative small GTP-binding proteins, are important for activation of the CAL1/CDC43 gene product, a protein geranylgeranyltransferase in Saccharomyces cerevisiae. Yeast 1992, 8, 735–741. [Google Scholar] [CrossRef]
- Heinisch, J.J.; Rodicio, R. Protein kinase C in fungi—More than just cell wall integrity. FEMS Microbiol. Rev. 2017, 42, fux051. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.D.; Stark, M.J. Dynamic, Rho1p-dependent localization of Pkc1p to sites of polarized growth. J. Cell Sci. 2000, 113, 2685–2693. [Google Scholar] [CrossRef] [PubMed]
- Kono, K.; Saeki, Y.; Yoshida, S.; Tanaka, K.; Pellman, D. Proteasomal degradation resolves competition between cell polarization and cellular wound healing. Cell 2012, 150, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Varma, A.; Drgon, T.; Bowers, B.; Cabib, E. Septins, under Cla4p Regulation, and the Chitin Ring Are Required for Neck Integrity in Budding Yeast. Mol. Biol. Cell 2003, 14, 2128–2141. [Google Scholar] [CrossRef]
- Gomez, A.; Perez, J.; Reyes, A.; Duran, A.; Roncero, C. Slt2 and Rim101 Contribute Independently to the Correct Assembly of the Chitin Ring at the Budding Yeast Neck in Saccharomyces cerevisiae. Eukaryot. Cell 2009, 8, 1449–1459. [Google Scholar] [CrossRef][Green Version]
- Lesage, G.; Shapiro, J.; Specht, C.A.; Sdicu, A.-M.; Ménard, P.; Hussein, S.; Tong, A.H.Y.; Boone, C.; Bussey, H. An interactional network of genes involved in chitin synthesis in Saccharomyces cerevisiae. BMC Genet. 2005, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Lenardon, M.D.; Munro, C.; Gow, N.A. Chitin synthesis and fungal pathogenesis. Curr. Opin. Microbiol. 2010, 13, 416–423. [Google Scholar] [CrossRef]
- Valdivia, R.H.; Schekman, R. The yeasts Rho1p and Pkc1p regulate the transport of chitin synthase III (Chs3p) from internal stores to the plasma membrane. Proc. Natl. Acad. Sci. USA 2003, 100, 10287–10292. [Google Scholar] [CrossRef]
- DeMarini, D.J.; Adams, A.E.; Fares, H.; De Virgilio, C.; Valle, G.; Chuang, J.S.; Pringle, J.R. A septin-based hierarchy of proteins required for localized deposition of chitin in the Saccharomyces cerevisiae cell wall. J. Cell Biol. 1997, 139, 75–93. [Google Scholar] [CrossRef]
- Reyes, A.; Sanz, M.; Duran, A.; Roncero, C. Chitin synthase III requires Chs4p-dependent translocation of Chs3p into the plasma membrane. J. Cell Sci. 2007, 120, 1998–2009. [Google Scholar] [CrossRef]
- Sacristan, C.; Reyes, A.; Roncero, C. Neck compartmentalization as the molecular basis for the different endocytic behaviour of Chs3 during budding or hyperpolarized growth in yeast cells. Mol. Microbiol. 2012, 83, 1124–1135. [Google Scholar] [CrossRef]
- Oh, Y.; Schreiter, J.H.; Okada, H.; Wloka, C.; Okada, S.; Yan, D.; Duan, X.; Bi, E. Hof1 and Chs4 Interact via F-BAR Domain and Sel1-like Repeats to Control Extracellular Matrix Deposition during Cytokinesis. Curr. Biol. 2017, 27, 2878–2886. [Google Scholar] [CrossRef]
- Lagorce, A.; Le Berre-Anton, V.; Aguilar-Uscanga, B.; Martin-Yken, H.; Dagkessamanskaia, A.; François, J.M. Involvement ofGFA1, which encodes glutamine-fructose-6-phosphate amidotransferase, in the activation of the chitin synthesis pathway in response to cell-wall defects in Saccharomyces cerevisiae. JBIC J. Biol. Inorg. Chem. 2002, 269, 1697–1707. [Google Scholar] [CrossRef]
- Niwa, M. A cell cycle checkpoint for the endoplasmic reticulum. Biochim. Biophys. Acta Mol. Cell. Res. 2020, 1867, 118825. [Google Scholar] [CrossRef]
- Chin, C.F.; Bennett, A.M.; Ma, W.K.; Hall, M.; Yeong, F.M. Dependence of Chs2 ER export on dephosphorylation by cytoplasmic Cdc14 ensures that septum formation follows mitosis. Mol. Biol. Cell 2012, 23, 45–58. [Google Scholar] [CrossRef]
- Zhang, G.; Kashimshetty, R.; Ng, K.E.; Tan, H.B.; Yeong, F.M. Exit from mitosis triggers Chs2p transport from the endoplasmic reticulum to mother–daughter neck via the secretory pathway in budding yeast. J. Cell Biol. 2006, 174, 207–220. [Google Scholar] [CrossRef]
- Tolliday, N.; VerPlank, L.; Li, R. Rho1 directs formin-mediated actin ring assembly during budding yeast cytokinesis. Curr. Biol. 2002, 12, 1864–1870. [Google Scholar] [CrossRef]
- Yoshida, S.; Kono, K.; Lowery, D.M.; Bartolini, S.; Yaffe, M.B.; Ohya, Y.; Pellman, D. Polo-Like Kinase Cdc5 Controls the Local Activation of Rho1 to Promote Cytokinesis. Science 2006, 313, 108–111. [Google Scholar] [CrossRef]
- Cabib, E. The septation apparatus, a chitin-requiring machine in budding yeast. Arch. Biochem. Biophys. 2004, 426, 201–207. [Google Scholar] [CrossRef]
- Arellano, M.; Duran, A.; Perez, P. Rho 1 GTPase activates the (1-3)beta-D-glucan synthase and is involved in Schizosaccharomyces pombe morphogenesis. EMBO J. 1996, 15, 4584–4591. [Google Scholar] [CrossRef]
- Schmidt, M.; Bowers, B.; Varma, A.; Roh, D.-H.; Cabib, E. In budding yeast, contraction of the actomyosin ring and formation of the primary septum at cytokinesis depend on each other. J. Cell Sci. 2002, 115, 293–302. [Google Scholar] [CrossRef]
- Ver Plank, L.; Li, R. Cell Cycle-regulated Trafficking of Chs2 Controls Actomyosin Ring Stability during Cytokinesis. Mol. Biol. Cell 2005, 16, 2529–2543. [Google Scholar] [CrossRef] [PubMed]
- Foltman, M.; Filali-Mouncef, Y.; Crespo, D.; Sanchez-Diaz, A. Cell polarity protein Spa2 coordinates Chs2 incorporation at the division site in budding yeast. PLoS Genet. 2018, 14, e1007299. [Google Scholar] [CrossRef] [PubMed]
- Foltman, M.; Molist, I.; Arcones, I.; Sacristan, C.; Filali-Mouncef, Y.; Roncero, C.; Sanchez-Diaz, A. Ingression Progression Complexes Control Extracellular Matrix Remodelling during Cytokinesis in Budding Yeast. PLoS Genet. 2016, 12, e1005864. [Google Scholar] [CrossRef]
- Devrekanli, A.; Foltman, M.; Roncero, C.; Sanchez-Diaz, A.; Labib, K. Inn1 and Cyk3 regulate chitin synthase during cytokinesis in budding yeasts. J. Cell Sci. 2012, 125, 5453–5466. [Google Scholar] [CrossRef]
- Guo, S.; Shen, X.; Yan, G.; Ma, D.; Bai, X.; Li, S.; Jiang, Y. A MAP Kinase Dependent Feedback Mechanism Controls Rho1 GTPase and Actin Distribution in Yeast. PLoS ONE 2009, 4, e6089. [Google Scholar] [CrossRef]
- Prosser, D.C.; Drivas, T.G.; Maldonado-Báez, L.; Wendland, B. Existence of a novel clathrin-independent endocytic pathway in yeast that depends on Rho1 and formin. J. Cell Biol. 2011, 195, 657–671. [Google Scholar] [CrossRef]
- Boettner, D.R.; D’Agostino, J.L.; Torres, O.T.; Daugherty-Clarke, K.; Uygur, A.; Reider, A.; Wendland, B.; Lemmon, S.K.; Goode, B.L. The F-BAR protein Syp1 negatively regulates WASp-Arp2/3 complex activity during endocytic patch formation. Curr. Biol. 2009, 19, 1979–1987. [Google Scholar] [CrossRef]
- Merlini, L.; Bolognesi, A.; Juanes, M.A.; Vandermoere, F.; Courtellemont, T.; Pascolutti, R.; Séveno, M.; Barral, Y.; Piatti, S. Rho1- and Pkc1-dependent phosphorylation of the F-BAR protein Syp1 contributes to septin ring assembly. Mol. Biol. Cell 2015, 26, 3245–3262. [Google Scholar] [CrossRef]
- Mascaraque, V.; Hernaez, M.L.; Jimenez-Sanchez, M.; Hansen, R.; Gil, C.; Martin, H.; Cid, V.J.; Molina, M. Phosphoproteomic analysis of protein kinase C signaling in Saccharomyces cerevisiae reveals Slt2 mitogen-activated protein kinase (MAPK)-dependent phosphorylation of eisosome core components. Mol. Cell Proteom. 2013, 12, 557–574. [Google Scholar] [CrossRef] [PubMed]
- Ziolkowska, N.E.; Karotki, L.; Rehman, M.; Huiskonen, J.T.; Walther, T.C. Eisosome-driven plasma membrane organization is mediated by BAR domains. Nat. Struct. Mol. Biol. 2011, 18, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Chang, K.J.; Orlean, P.; Wloka, C.; Deshaies, R.; Bi, E. Mitotic exit kinase Dbf2 directly phosphorylates chitin synthase Chs2 to regulate cytokinesis in budding yeast. Mol. Biol. Cell 2012, 23, 2445–2456. [Google Scholar] [CrossRef]
- Shaw, J.A.; Mol, P.C.; Bowers, B.; Silverman, S.J.; Valdivieso, M.-H.; Durán, A.; Cabib, E. The function of chitin synthases 2 and 3 in the Saccharomyces cerevisiae cell cycle. J. Cell Biol. 1991, 114, 111–123. [Google Scholar] [CrossRef]
- Yoshida, S.; Bartolini, S.; Pellman, D. Mechanisms for concentrating Rho1 during cytokinesis. Genes Dev. 2009, 23, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Cabib, E.; Blanco, N.; Grau, C.; Rodriguez-Pena, J.M.; Arroyo, J. Crh1p and Crh2p are required for the cross-linking of chitin to beta(1-6)glucan in the Saccharomyces cerevisiae cell wall. Mol. Microbiol. 2007, 63, 921–935. [Google Scholar] [CrossRef]
- Cabib, E.; Arroyo, J. How carbohydrates sculpt cells: Chemical control of morphogenesis in the yeast cell wall. Nat. Rev. Genet. 2013, 11, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Dohrmann, P.R.; Butler, G.; Tamai, K.; Dorland, S.; Greene, J.R.; Thiele, D.J.; Stillman, D.J. Parallel pathways of gene regulation: Homologous regulators SWI5 and ACE2 differentially control transcription of HO and chitinase. Genes Dev. 1992, 6, 93–104. [Google Scholar] [CrossRef][Green Version]
- Kuranda, M.J.; Robbins, P.W. Chitinase is required for cell separation during growth of Saccharomyces cerevisiae. J. Biol. Chem. 1991, 266, 19758–19767. [Google Scholar] [CrossRef]
- Baladron, V.; Ufano, S.; Duenas, E.; Martin-Cuadrado, A.B.; del Rey, F.; Vazquez de Aldana, C.R. Eng1p, an endo-1,3-beta-glucanase localized at the daughter side of the septum, is involved in cell separation in Saccharomyces cerevisiae. Eukaryot. Cell 2002, 1, 774–786. [Google Scholar] [CrossRef]
- Colman-Lerner, A.; Chin, T.E.; Brent, R. Yeast Cbk1 and Mob2 Activate Daughter-Specific Genetic Programs to Induce Asymmetric Cell Fates. Cell 2001, 107, 739–750. [Google Scholar] [CrossRef]
- Cabib, E.; Silverman, S.J.; Shaw, J.A. Chitinase and chitin synthase 1: Counterbalancing activities in cell separation of Saccharomyces cerevisiae. J. Gen. Microbiol. 1992, 138, 97–102. [Google Scholar] [CrossRef]
- Brace, J.L.; Doerfler, M.; Weiss, E.L. A cell separation checkpoint that enforces the proper order of late cytokinetic events. J. Cell Biol. 2019, 218, 150–170. [Google Scholar] [CrossRef]
- Madden, K.; Sheu, Y.-J.; Baetz, K.; Andrews, B.; Snyder, M. SBF Cell Cycle Regulator as a Target of the Yeast PKC-MAP Kinase Pathway. Science 1997, 275, 1781–1784. [Google Scholar] [CrossRef]
- Jonasson, E.M.; Rossio, V.; Hatakeyama, R.; Abe, M.; Ohya, Y.; Yoshida, S. Zds1/Zds2-PP2ACdc55 com-plex specifies signaling output from Rho1 GTPase. J. Cell Biol. 2016, 212, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Perez, J.; Arcones, I.; Gomez, A.; Casquero, V.; Roncero, C. Phosphorylation of Bni4 by MAP kinases contributes to septum assembly during yeast cytokinesis. FEMS Yeast Res. 2016, 16, fow060. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Friesen, H.; Larson, J.; Huang, D.; Cox, M.; Tatchell, K.; Andrews, B. Regulation of Cell Polarity through Phosphorylation of Bni4 by Pho85 G1 Cyclin-dependent Kinases in Saccharomyces cerevisiae. Mol. Biol. Cell 2009, 20, 3239–3250. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sanz, M.; Castrejón, F.; Durán, A.; Roncero, C. Saccharomyces cerevisiae Bni4p directs the formation of the chitin ring and also participates in the correct assembly of the septum structure. Microbiology 2004, 150, 3229–3241. [Google Scholar] [CrossRef]
- Kozubowski, L.; Larson, J.; Tatchell, K. Role of the Septin Ring in the Asymmetric Localization of Proteins at the Mother-Bud Neck in Saccharomyces cerevisiae. Mol. Biol. Cell 2005, 16, 3455–3466. [Google Scholar] [CrossRef]
- Dichtl, K.; Samantaray, S.; Wagener, J. Cell wall integrity signalling in human pathogenic fungi. Cell. Microbiol. 2016, 18, 1228–1238. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roncero, C.; Celador, R.; Sánchez, N.; García, P.; Sánchez, Y. The Role of the Cell Integrity Pathway in Septum Assembly in Yeast. J. Fungi 2021, 7, 729. https://doi.org/10.3390/jof7090729
Roncero C, Celador R, Sánchez N, García P, Sánchez Y. The Role of the Cell Integrity Pathway in Septum Assembly in Yeast. Journal of Fungi. 2021; 7(9):729. https://doi.org/10.3390/jof7090729
Chicago/Turabian StyleRoncero, Cesar, Rubén Celador, Noelia Sánchez, Patricia García, and Yolanda Sánchez. 2021. "The Role of the Cell Integrity Pathway in Septum Assembly in Yeast" Journal of Fungi 7, no. 9: 729. https://doi.org/10.3390/jof7090729
APA StyleRoncero, C., Celador, R., Sánchez, N., García, P., & Sánchez, Y. (2021). The Role of the Cell Integrity Pathway in Septum Assembly in Yeast. Journal of Fungi, 7(9), 729. https://doi.org/10.3390/jof7090729