
Crop Cycle and Tillage Role in the Outbreak of Late Wilt Disease of Maize Caused by Magnaporthiopsis maydis



Abstract

1. Introduction
2. Materials and Methods
2.1. Fungal Species and Growth Conditions
2.2. The Preparation of Infected Sterilized Wheat Grains
2.3. The Plant Inoculation Methodology
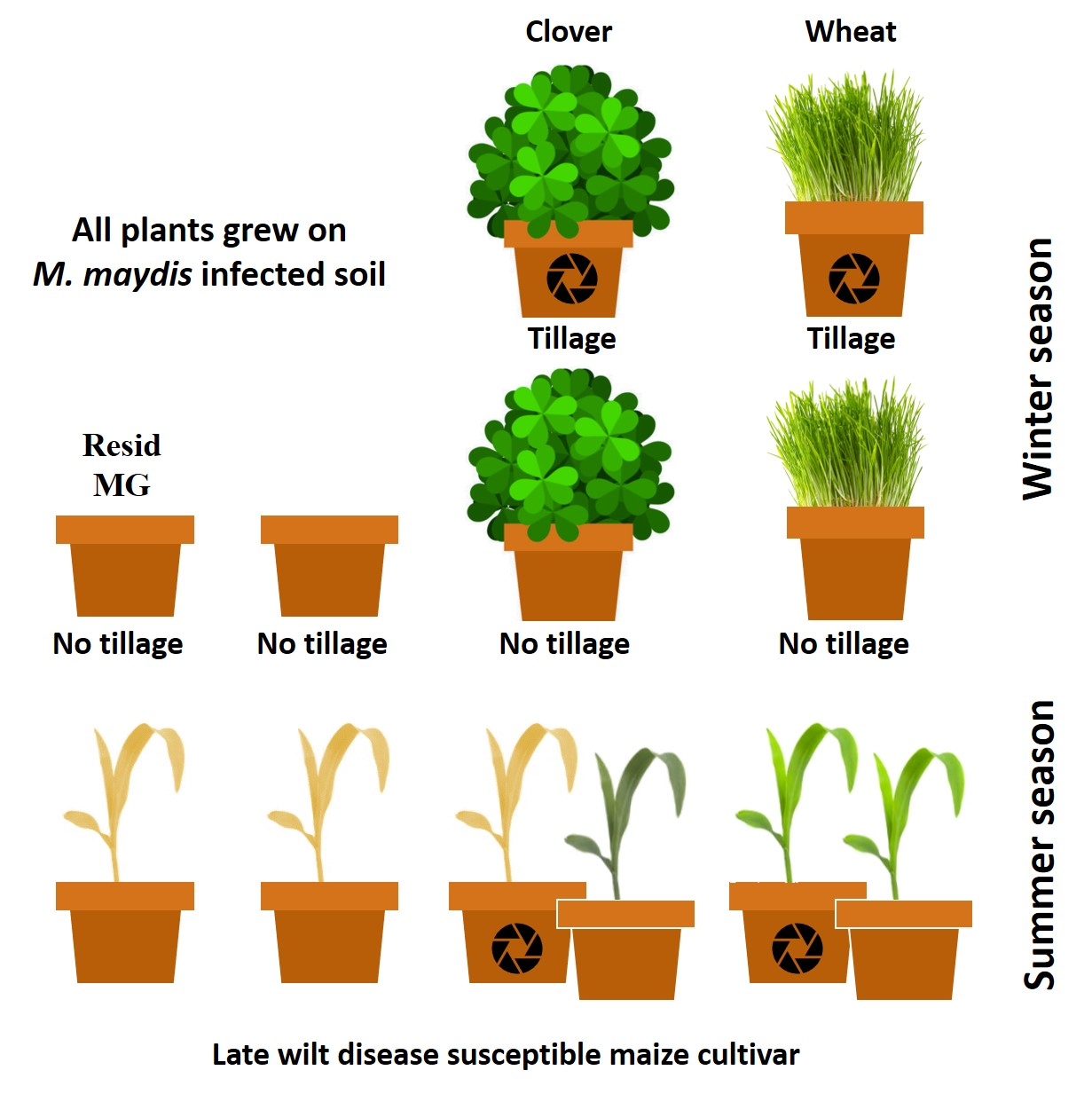
2.4. Crop Rotation and Tillage Effect in Pots over a Full Growing Season—The 2019 Experiment
2.4.1. Winter Season
2.4.2. Summer Season
2.5. Evaluation of Commercial Field Soil after Wheat or Clover Growth—The 2020 Experiment
2.6. Molecular Diagnosis of the Late Wilt Pathogen
2.7. Statistical Analyses
3. Results
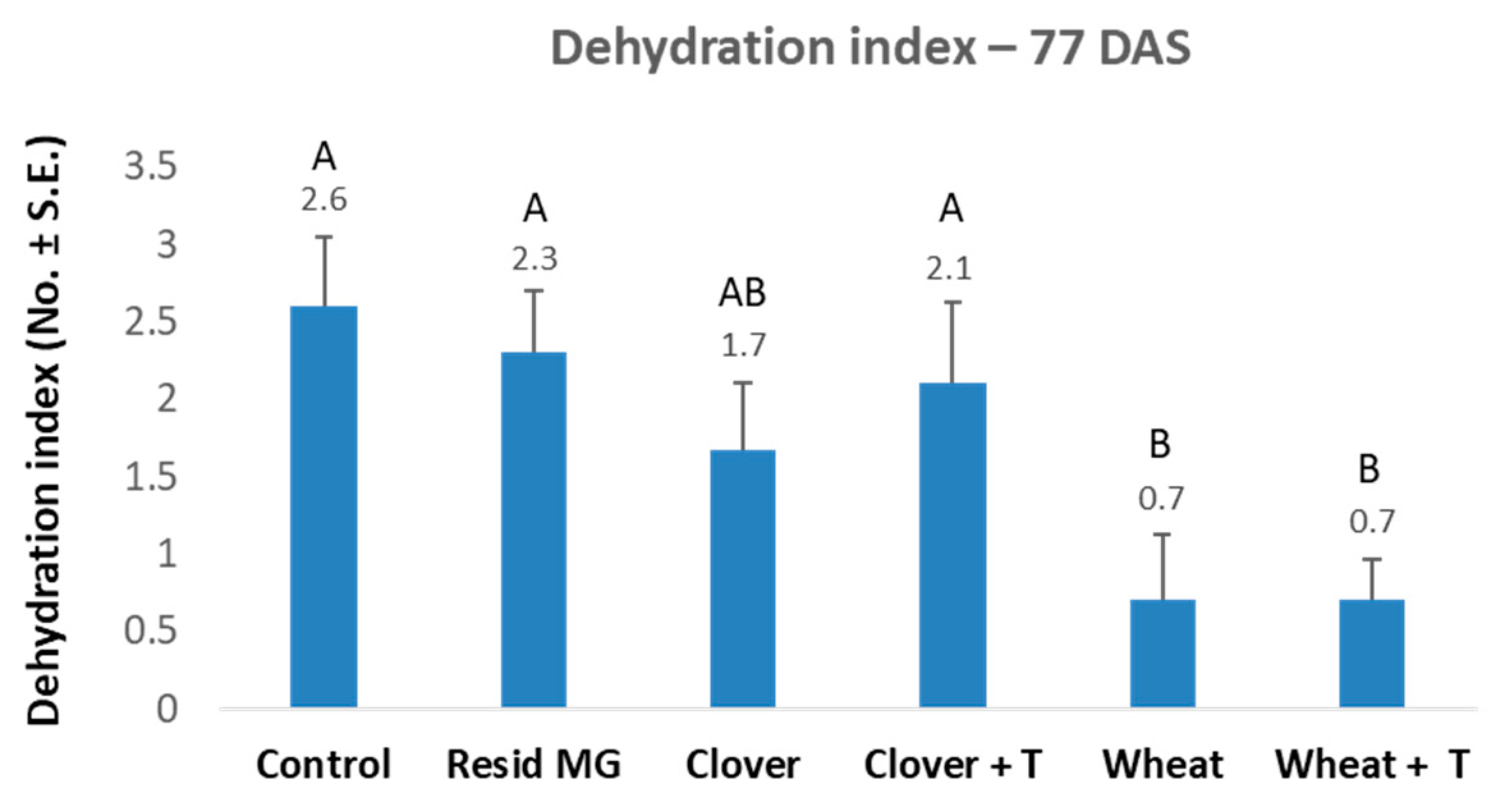
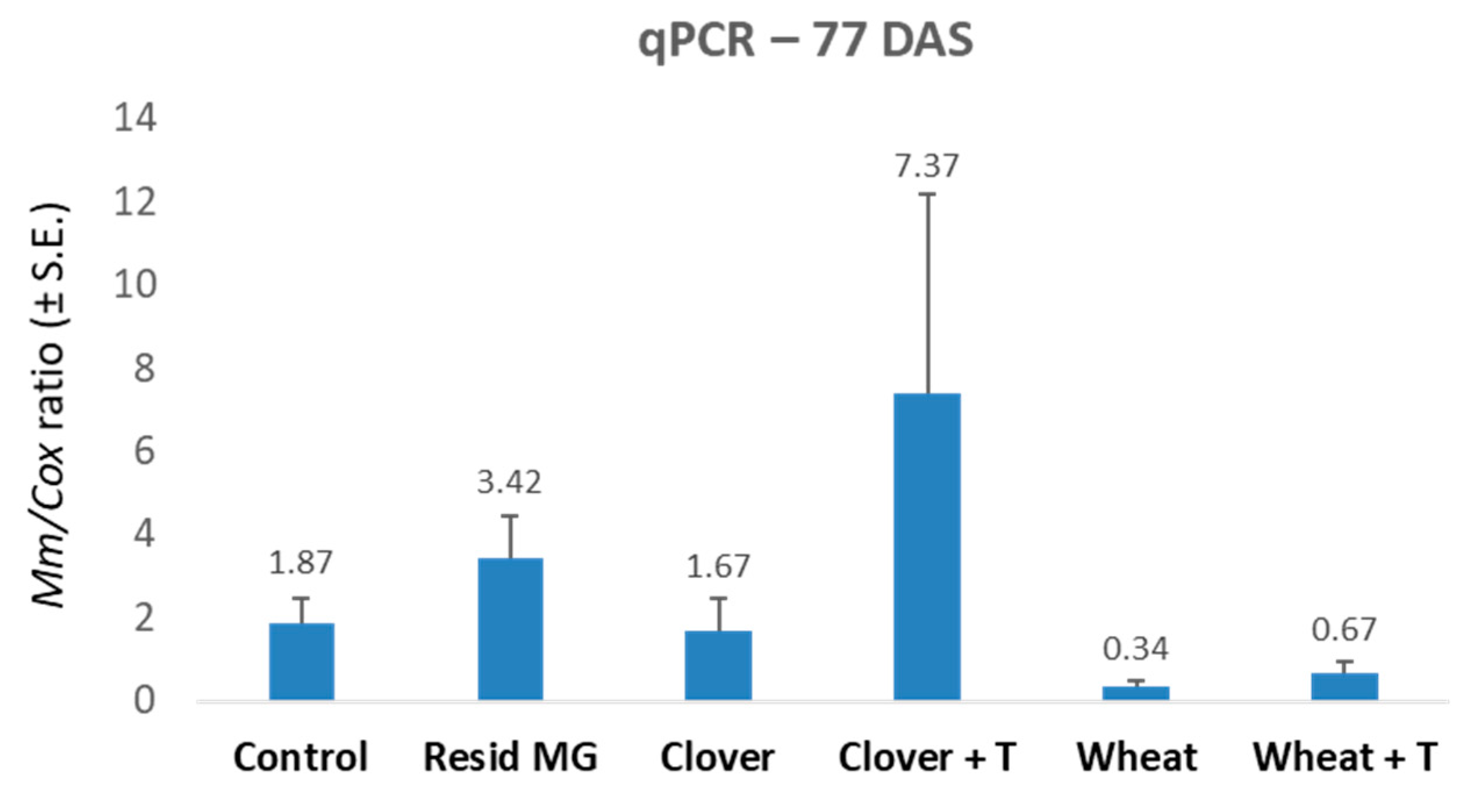
3.1. Crop Rotation and Tillage Effect in Pots over a Full Growing Season—The 2019 Experiment
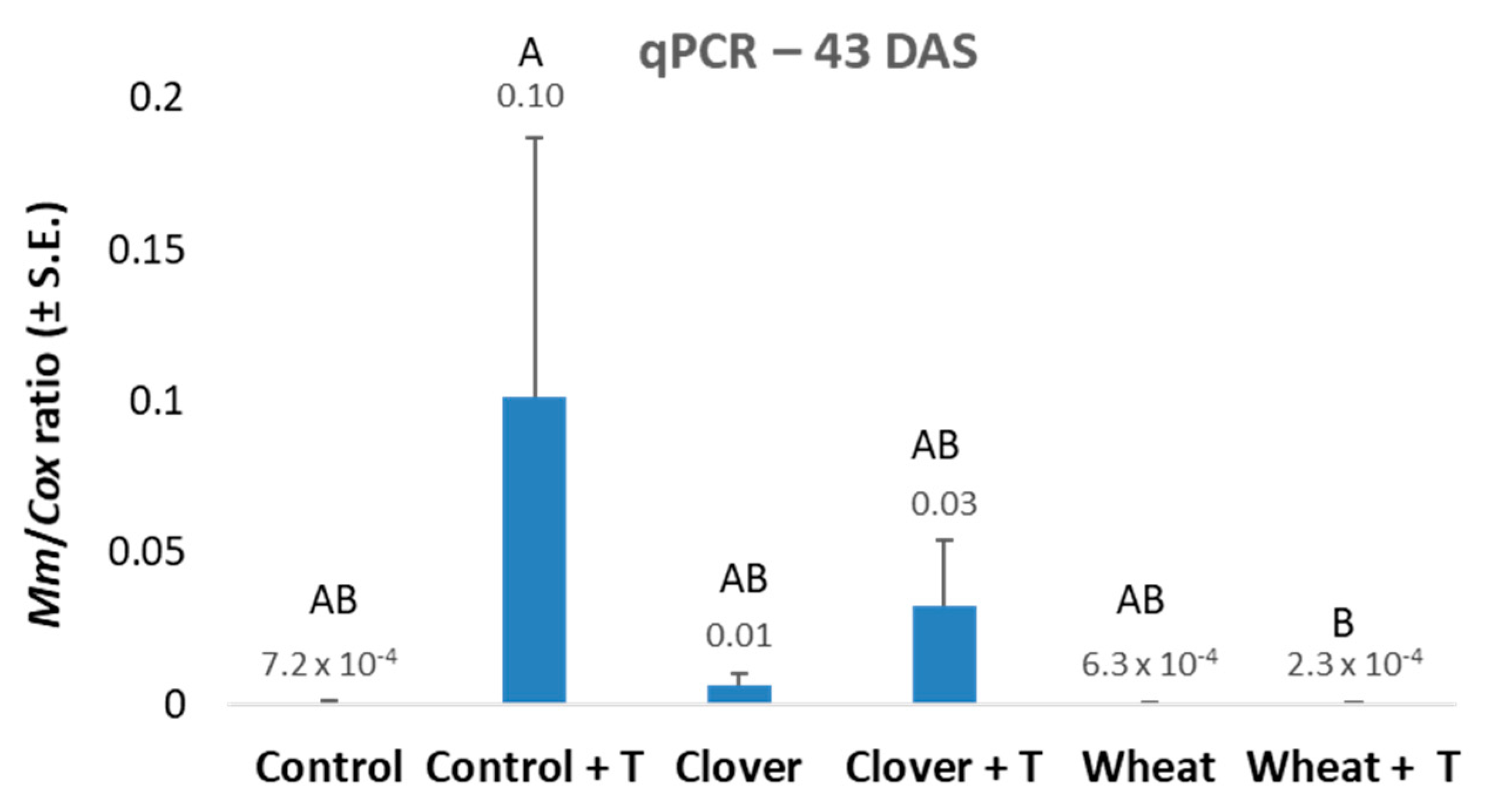
3.2. Evaluation of Commercial Field Soil after Wheat or Clover Growth—The 2020 Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olaniyan, A.B. Maize: Panacea for hunger in Nigeria. Afr. J. Plant Sci. 2015, 9, 155–174. [Google Scholar]
- Tanklevska, N.; Petrenko, V.; Karnaushenko, А.; Melnykova, К. World corn market: Analysis, trends and prospects of its deep processing. Int. Sci. E-J. Agric. Resour. Econ. 2020, 6, 96–111. [Google Scholar] [CrossRef]
- Drori, R.; Sharon, A.; Goldberg, D.; Rabinovitz, O.; Levy, M.; Degani, O. Molecular diagnosis for Harpophora maydis, the cause of maize late wilt in Israel. Phytopathol. Mediterr. 2013, 52, 16–29. [Google Scholar]
- Degani, O.; Dor, S.; Movshowitz, D.; Fraidman, E.; Rabinovitz, O.; Graph, S. Effective chemical protection against the maize late wilt causal agent, Harpophora maydis, in the field. PLoS ONE 2018, 13, e0208353. [Google Scholar] [CrossRef]
- Al Taweel, H. Mapping of QTLs for Resistance to Late Wilt of Maize in Egypt Using SNPs. Master’s Thesis, The American University in Cairo Digital Archive and Research Repository, Cairo, Egypt, 2013. [Google Scholar]
- Rakesh, B.; Gangappa, E.; Sonali, G.; Gowda, R.P.V.; Swamy, S.D.; Ramesh, S.; Hemareddy, H.B.; Nagaraju, N. Modified method of screening maize inbred lines to late wilt disease caused by Harpophora maydis. Mysore J. Agric. Sci. 2016, 50, 684–690. [Google Scholar]
- Sunitha, N.; Gangappa, E.; Gowda, R.V.; Ramesh, S.; Biradar, S.; Swamy, D.; Hemareddy, H.J. Assessment of impact of late wilt caused by Harpophora maydis (Samra, Sabet and Hing) on grain yield and its attributing traits in maize (Zea mays L.). Mysore J. Agric. Sci. 2020, 54, 30–36. [Google Scholar]
- Molinero-Ruiz, M.L.; Melero-Vara, J.M.; Mateos, A. Cephalosporium maydis, the Cause of Late Wilt in Maize, a Pathogen New to Portugal and Spain. Plant Dis. 2010, 94, 379. [Google Scholar] [CrossRef] [PubMed]
- Klaubauf, S.; Tharreau, D.; Fournier, E.; Groenewald, J.Z.; Crous, P.W.; de Vries, R.P.; Lebrun, M.H. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol 2014, 79, 85–120. [Google Scholar] [CrossRef] [PubMed]
- Samra, A.S.; Sabet, K.A.; Hingorani, M.K. Late wilt disease of maize caused by Cephalosporium maydis. Phytopathology 1963, 53, 402–406. [Google Scholar]
- Gams, W. Phialophora and some similar morphologically little-differentiated anamorphs of divergent ascomycetes. Stud. Mycol. 2000, 45, 187–200. [Google Scholar]
- Saleh, A.A.; Leslie, J.F. Cephalosporium maydis is a distinct species in the Gaeumannomyces-Harpophora species complex. Mycologia 2004, 96, 1294–1305. [Google Scholar] [CrossRef]
- Tej, R.; Rodríguez-Mallol, C.; Rodríguez-Arcos, R.; Karray-Bouraoui, N.; Molinero-Ruiz, L. Inhibitory effect of Lycium europaeum extracts on phytopathogenic soil-borne fungi and the reduction of late wilt in maize. Eur. J. Plant Pathol. 2018, 152, 249–265. [Google Scholar] [CrossRef]
- Samra, A.S.; Sabet, K.A.; Hingorani, M.K. A new wilt disease of maize in Egypt. Plant Dis. Rep. 1962, 46, 481–483. [Google Scholar]
- Ortiz-Bustos, C.; López-Bernal, A.; Testi, L.; Molinero-Ruiz, L.J. Environmental and irrigation conditions can mask the effect of Magnaporthiopsis maydis on growth and productivity of maize. Plant Pathol. 2019, 68, 1555–1564. [Google Scholar] [CrossRef]
- Degani, O.; Rabinovitz, O.; Becher, P.; Gordani, A.; Chen, A. Trichoderma longibrachiatum and Trichoderma asperellum Confer Growth Promotion and Protection against Late Wilt Disease in the Field. J. Fungi 2021, 7, 444. [Google Scholar] [CrossRef]
- Degani, O.; Cernica, G. Diagnosis and Control of Harpophora maydis, the Cause of Late Wilt in Maize. Adv. Microbiol. 2014, 4, 94–105. [Google Scholar] [CrossRef]
- Degani, O.; Movshowitz, D.; Dor, S.; Meerson, A.; Goldblat, Y.; Rabinovitz, O. Evaluating Azoxystrobin Seed Coating Against Maize Late Wilt Disease Using a Sensitive qPCR-Based Method. Plant Dis. 2019, 103, 238–248. [Google Scholar] [CrossRef]
- Michail, S.H.; Abou-Elseoud, M.S.; Nour Eldin, M.S. Seed health testing of corn for Cephalosporium maydis. Acta Phytopathol. Entomol. Hung. 1999, 34, 35–42. [Google Scholar]
- Sabet, K.A.; Samra, A.S.; Mansour, I.M. Saprophytic behaviour of Cephalosporium maydis and C. acremonium. Ann. Appl. Biol. 1970, 66, 265–271. [Google Scholar] [CrossRef]
- Sahab, A.F.; Osman, A.R.; Soleman, N.K.; Mikhail, M.S. Studies on root-rot of lupin in Egypt and its control. Egypt. J. Phytopathol. 1985, 17, 23–35. [Google Scholar]
- Degani, O.; Dor, S.; Abraham, D.; Cohen, R. Interactions between Magnaporthiopsis maydis and Macrophomina phaseolina, the Causes of Wilt Diseases in Maize and Cotton. Microorganisms 2020, 8, 249. [Google Scholar] [CrossRef]
- Sabet, K.; Samra, A.; Mansour, I. Interaction between Fusarium oxysporum f. vasinfectum and Cephalosporium maydis on cotton and maize. Ann. Appl. Biol. 1966, 58, 93–101. [Google Scholar] [CrossRef]
- Dor, S.; Degani, O. Uncovering the Host Range for Maize Pathogen Magnaporthiopsis maydis. Plants 2019, 8, 259. [Google Scholar] [CrossRef]
- Singh, S.; Siradhana, B. Effect of macro and micronutrients on the development of late wilt of maize induced by Cephalosporium maydis. Summa Phytopathol. 1990, 16, 140–145. [Google Scholar]
- Samra, A.S.; Sabet, K.A.; Abdel-Rahim, M.F. Effect of Soil Conditions and Cultural Practices on Infection with Stalk Rots; U.A.R. Ministry of Agric. Government Printing Offices: Cairo, Egypt, 1966; pp. 117–164.
- Patanita, M.; Campos, M.D.; Félix, M.d.R.; Carvalho, M.; Brito, I. Effect of tillage system and cover crop on maize mycorrhization and presence of Magnaporthiopsis maydis. Biology 2020, 9, 46. [Google Scholar] [CrossRef]
- Singh, S.D.; Siradhana, B.S. Influence of some environmental conditions on the development of late wilt of maize induced by Cephalosporium maydis. Indian J. Mycol. Plant Pathol. 1987, 17, 1–5. [Google Scholar]
- Fayzalla, E.; Sadik, E.; Elwakil, M.; Gomah, A. Soil solarization for controlling Cephalosporium maydis, the cause of late wilt disease of maize in Egypt. Egypt J. Phytopathol. 1994, 22, 171–178. [Google Scholar]
- Abd-el-Rahim, M.F.; Sabet, K.A.; El-Shafey, H.A.; El-Assiuty, E.M. Chemical control of the late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. 1982, 60, 31–49. [Google Scholar]
- Degani, O.; Dor, S.; Chen, A.; Orlov-Levin, V.; Stolov-Yosef, A.; Regev, D.; Rabinovitz, O. Molecular Tracking and Remote Sensing to Evaluate New Chemical Treatments against the Maize Late Wilt Disease Causal Agent, Magnaporthiopsis maydis. J. Fungi 2020, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Weinberg, T.; Graph, S. Chemical control of maize late wilt in the field. Phytoparasitica 2014, 42, 559–570. [Google Scholar] [CrossRef]
- Singh, S.D.; Siradhana, B.S. Chemical control of late wilt of maize induced by Cephalosporium maydis. Indian J. Mycol. Plant Path. 1989, 19, 121–122. [Google Scholar]
- Ghazy, N.; El-Gremi, S.; Belal, E.-S. Chemical and Histological Differences of Corn (Zea mays L.) Responsive to Harpophora maydis Infection. Environ. Biodivers. Soil Secur. 2017, 1, 3–7. [Google Scholar] [CrossRef]
- El-Shafey, H.A.; El-Shorbagy, F.A.; Khalil, I.I.; El-Assiuty, E.M. Additional sources of resistance to the late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. Egypt 1988, 66, 221–230. [Google Scholar]
- Ortiz-Bustos, C.M.; Testi, L.; García-Carneros, A.B.; Molinero-Ruiz, L. Geographic distribution and aggressiveness of Harpophora maydis in the Iberian peninsula, and thermal detection of maize late wilt. Eur. J. Plant Pathol. 2015, 144, 383–397. [Google Scholar] [CrossRef]
- Zeller, K.A.; Ismael, A.M.; El-Assiuty, E.M.; Fahmy, Z.M.; Bekheet, F.M.; Leslie, J.F. Relative Competitiveness and Virulence of Four Clonal Lineages of Cephalosporium maydis from Egypt Toward Greenhouse-Grown Maize. Plant Dis. 2002, 86, 373–378. [Google Scholar] [CrossRef]
- Avila-Adame, C.; Koller, W. Characterization of spontaneous mutants of Magnaporthe grisea expressing stable resistance to the Qo-inhibiting fungicide azoxystrobin. Curr. Genet. 2003, 42, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef] [PubMed]
- Ghazy, N.; El-Nahrawy, S. Siderophore production by Bacillus subtilis MF497446 and Pseudomonas koreensis MG209738 and their efficacy in controlling Cephalosporium maydis in maize plant. Arch. Microbiol. 2021, 203, 1195–1209. [Google Scholar] [CrossRef] [PubMed]
- Elshahawy, I.E.; El-Sayed, A.E.-K.B. Maximizing the efficacy of Trichoderma to control Cephalosporium maydis, causing maize late wilt disease, using freshwater microalgae extracts. Egypt. J. Biol. Pest. Control. 2018, 28, 48. [Google Scholar] [CrossRef]
- Gal-Hemed, I.; Atanasova, L.; Komon-Zelazowska, M.; Druzhinina, I.S.; Viterbo, A.; Yarden, O. Marine isolates of Trichoderma spp. as potential halotolerant agents of biological control for arid-zone agriculture. Appl. Environ. Microbiol. 2011, 77, 5100–5109. [Google Scholar] [CrossRef]
- Degani, O.; Danielle, R.; Dor, S. The microflora of maize grains as a biological barrier against the late wilt causal agent, Magnaporthiopsis maydis. Agronomy 2021, 11, 965. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S. Trichoderma Biological Control to Protect Sensitive Maize Hybrids against Late Wilt Disease in the Field. J. Fungi 2021, 7, 315. [Google Scholar] [CrossRef] [PubMed]
- Brígido, C.; van Tuinen, D.; Brito, I.; Alho, L.; Goss, M.J.; Carvalho, M. Management of the biological diversity of AM fungi by combination of host plant succession and integrity of extraradical mycelium. Soil Biol. Biochem. 2017, 112, 237–247. [Google Scholar] [CrossRef]
- Sharma, M.; Sudheer, S.; Usmani, Z.; Rani, R.; Gupta, P. Deciphering the omics of plant-microbe interaction: Perspectives and new insights. Curr. Genom. 2020, 21, 343–362. [Google Scholar] [CrossRef]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Martín, F.F.; Molina, J.J.; Nicolás, E.N.; Alarcón, J.J.; Kirchmair, M.; García, F.J.; Garcia, A.J.B.; Bernal, C. Application of Arbuscular Mycorrhizae Glomus iranicum var. tenuihypharum var. nova in Intensive Agriculture: A study case. J. Agric. Sci. Technol. B 2017, 7, 221–247. [Google Scholar]
- Degani, O.; Goldblat, Y. Ambient Stresses Regulate the Development of the Maize Late Wilt Causing Agent, Harpophora maydis. Agric. Sci. 2014, 5, 571–582. [Google Scholar] [CrossRef]
- Degani, O.; Regev, D.; Dor, S.; Rabinovitz, O. Soil bioassay for detecting Magnaporthiopsis maydis infestation using a hyper susceptible maize hybrid. J. Fungi 2020, 6, 107. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Dor, S.; Movshovitz, D.; Rabinovitz, O. Methods for Studying Magnaporthiopsis maydis, the Maize Late Wilt Causal Agent. Agronomy 2019, 9, 181. [Google Scholar] [CrossRef]
- Abendroth, L.J.; Elmore, R.W.; Boyer, M.J.; Marlay, S.K. Corn Growth and Development; PM&R: Ames, IA, USA, 2011. [Google Scholar]
- Saleh, A.A.; Zeller, K.A.; Ismael, A.S.; Fahmy, Z.M.; El-Assiuty, E.M.; Leslie, J.F. Amplified Fragment Length Polymorphism Diversity in Cephalosporium maydis from Egypt. Phytopathology 2003, 93, 853–859. [Google Scholar] [CrossRef]
- Zeller, K.A.; Jurgenson, J.E.; El-Assiuty, E.M.; Leslie, J.F. Isozyme and amplified fragment length polymorphisms from Cephalosporium maydis in Egypt. Phytoparasitica 2000, 28, 121–130. [Google Scholar] [CrossRef]
- Li, W.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus huanglongbing. J. Microbiol. Methods 2006, 66, 104–115. [Google Scholar] [CrossRef]
- Weller, S.; Elphinstone, J.; Smith, N.; Boonham, N.; Stead, D. Detection of Ralstonia solanacearumstrains with a quantitative, multiplex, real-time, fluorogenic PCR (TaqMan) assay. Appl. Environ. Microbiol. 2000, 66, 2853–2858. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N., Jr. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef]
- Balota, E.L.; Colozzi-Filho, A.; Andrade, D.S.; Dick, R.P. Microbial biomass in soils under different tillage and crop rotation systems. Biol. Fertil. Soils 2003, 38, 15–20. [Google Scholar] [CrossRef]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular mycorrhizal fungi as natural biofertilizers: Let’s benefit from past successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef] [PubMed]
- Tahat, M.; Sijam, K.; Othman, R. Mycorrhizal fungi as a biocontrol agent. Plant Pathol. J. 2010, 9, 198–207. [Google Scholar] [CrossRef]
- Brito, I.; Goss, M.; Alho, L.; Brígido, C.; van Tuinen, D.; Félix, M.; Carvalho, M. Agronomic management of AMF functional diversity to overcome biotic and abiotic stresses-The role of plant sequence and intact extraradical mycelium. Fungal Ecol. 2019, 40, 72–81. [Google Scholar] [CrossRef]
- El-Shabrawy, E.-S.; Shehata, H. Controlling maize late-wilt and enhancing plant salinity tolerance by some rhizobacterial strains. Egypt. J. Phytopathol. 2018, 46, 235–255. [Google Scholar] [CrossRef]
- Sabet, K.A.; Zaher, A.M.; Samra, A.S.; Mansour, I.M. Pathogenic behaviour of Cephalosporium maydis and C. acremonium. Ann. Appl. Biol. 1970, 66, 257–263. [Google Scholar] [CrossRef]
- Smit, E.; Van Rensburg, G.D.J.; Rijkenberg, F. Number of isolates of maize root fungi in different crop rotation systems. South Afr. J. Plant Soil 1997, 14, 127–130. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
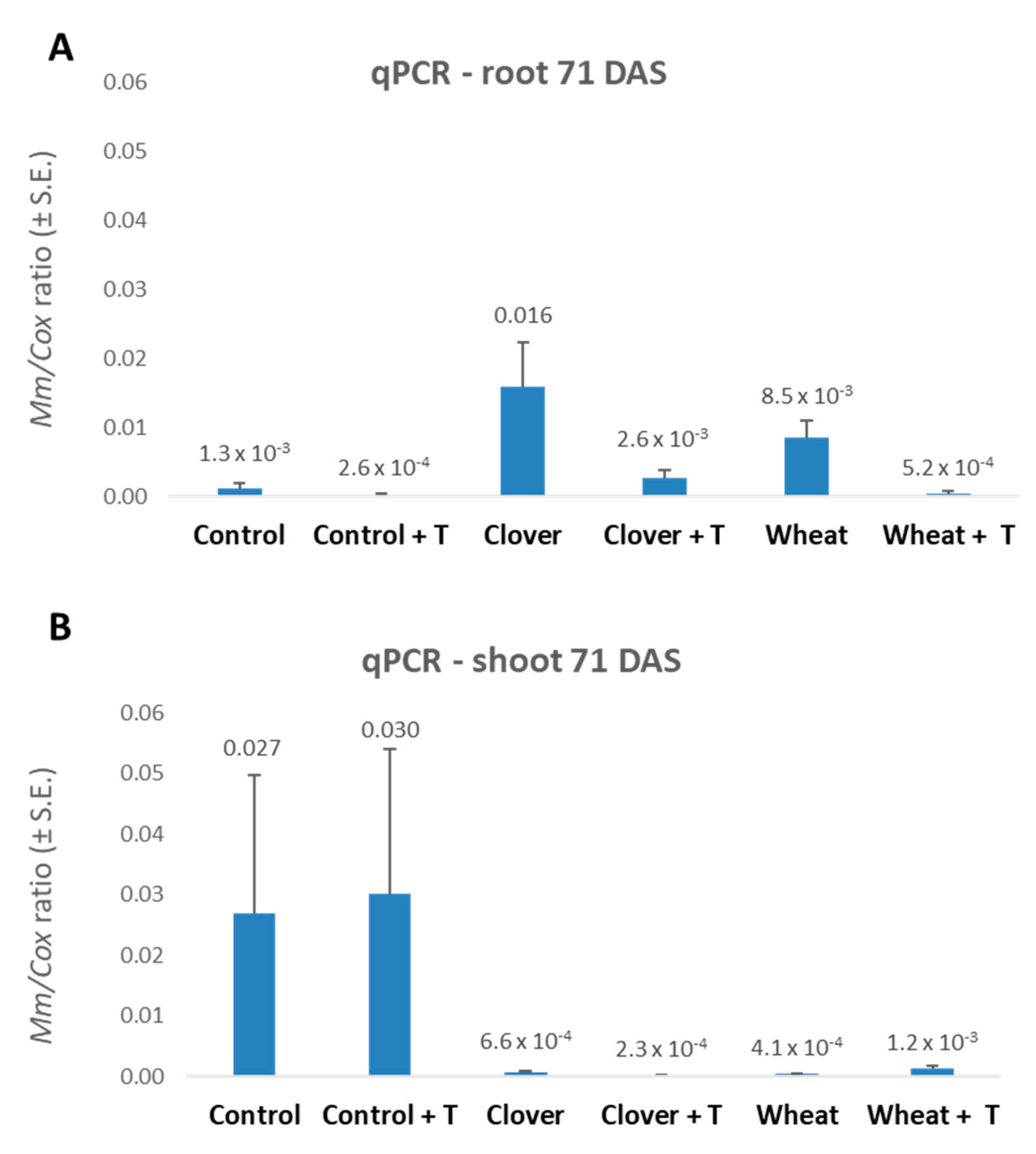{kind=link}
| Tillage | Emergence (%) | Root Weight (g) | Shoot Weight (g) | Leaves (No.) | Height (cm) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | − | 78% ± 7% | B | 0.80 ± 0.07 | A | 10.8 ± 0.8 | AB | 6.7 ± 0.1 | A | 7.1 ± 0.3 | B |
| Resid MG | − | 94% ± 4% | A | 0.88 ± 0.12 | A | 10.2 ± 1.5 | B | 5.8 ± 1.0 | B | 10.4 ± 0.7 | A |
| Clover | − | 84% ± 7% | AB | 0.79 ± 0.08 | A | 13.2 ± 0.9 | A | 6.9 ± 0.2 | A | 7.4 ± 0.3 | B |
| + | 94% ± 3% | A | 0.72 ± 0.06 | A | 9.5 ± 1.0 | B | 7.3 ± 0.3 | A | 7.6 ± 0.3 | B | |
| Wheat | − | 88% ± 5% | AB | 0.85 ± 0.06 | A | 8.9 ± 0.6 | B | 5.8 ± 0.2 | B | 11.2 ± 0.5 | A |
| + | 96% ± 4% | A | 0.66 ± 0.08 | A | 10.4 ± 0.8 | AB | 7.0 ± 0.4 | A | 7.0 ± 0.4 | B |
| Tillage | Phenological Development | Height (cm) | ||
|---|---|---|---|---|
| Control | − | Dis.–1, Vt–1, R1–8 | 140.0 ± 9.0 | AB |
| Resid MG | − | Dis.–2, Vt–5, R1–3 | 125.5 ± 9.9 | BC |
| Clover | − | Vt–4, R1–6 | 117.5 ± 6.2 | C |
| + | Vt–1, R1–9 | 125.5 ± 3.2 | BC | |
| Wheat | − | R1–10 | 127.0 ± 3.4 | BC |
| + | Dead–1, Dis.–1, R1–8 | 147.2 ± 3.1 | A |
| Tillage | Phenological Development | Shoot Wet Weight (g) | Height (cm) | Cob Wet Weight (g) | ||||
|---|---|---|---|---|---|---|---|---|
| Control | − | Vt–1, R5–9 | 98.1 ± 9.8 | C | 132.5 ± 8.6 | AB | 75.7 ± 6.3 | B |
| Resid MG | − | R5–10 | 129.5 ± 9.4 | ABC | 148.8 ± 4.6 | A | 95.2 ± 10.6 | AB |
| Clover | − | R5–10 | 116.2 ± 15.5 | BC | 122.5 ± 9.8 | B | 87.4 ± 8.5 | AB |
| + | V12–1, R5–9 | 111.7 ± 13.4 | BC | 122.5 ± 10.7 | B | 92.2 ± 11.8 | AB | |
| Wheat | − | Vt–1, R5–9 | 144.1 ± 10.5 | AB | 132.9 ± 6.0 | AB | 110.9 ± 9.0 | A |
| + | R5–10 | 151.4 ± 7.0 | A | 134.6 ± 7.0 | AB | 103.1 ± 11.5 | AB |
| Tillage | Shoot Dry Weight (g) | Cob Dry Weight (g) | |
|---|---|---|---|
| Control | − | 59.1 ± 6.3 | 47.0 ± 3.4 |
| Resid MG | − | 59.6 ± 6.1 | 60.8 ± 7.7 |
| Clover | − | 61.3 ± 8.3 | 53.9 ± 6.2 |
| + | 47.5 ± 9.4 | 48.3 ± 7.6 | |
| Wheat | − | 65.5 ± 3.8 | 61.7 ± 7.9 |
| + | 60.4 ± 5.8 | 65.5 ± 6.5 |
| Tillage | Emergence (%) | Shoot Wet Weight (g) | Leaves (no.) | Height (cm) | |
|---|---|---|---|---|---|
| Control | − | 58% ± 5.5% | 317.2 ± 22.4 | 8.2 ± 0.4 | 78.3 ± 1.2 |
| + | 36% ± 7.8% | 307.8 ± 24.4 | 7.8 ± 0.4 | 84.0 ± 3.0 | |
| Clover | − | 50% ± 10.0% | 329.2 ± 20.4 | 8.1 ± 0.3 | 79.7 ± 2.3 |
| + | 44% ± 8.3% | 303.0 ± 25.5 | 8.1 ± 0.4 | 81.9 ± 2.5 | |
| Wheat | − | 52% ± 9.5% | 333.4 ± 17.5 | 8.1 ± 0.3 | 83.9 ± 3.5 |
| + | 50% ± 5.4% | 345.3 ± 17.6 | 7.9 ± 0.3 | 84.9 ± 1.8 |
| Tillage | Shoot Weight (g) | Leaves (No.) | Height (cm) | Cob Wet Weight (g) | ||
|---|---|---|---|---|---|---|
| Control | − | 137.1 ± 9.4 | 8.7 ± 0.4 | 95.1 ± 4.6 | 118.1 ± 11.0 | AB |
| + | 153.6 ± 9.4 | 8.8 ± 0.3 | 99.2 ± 4.2 | 120.3 ± 4.5 | AB | |
| Clover | − | 153.4 ± 16.6 | 8.9 ± 0.6 | 100.4 ± 7.7 | 125.2 ± 7.7 | A |
| + | 146.8 ± 6.6 | 8.1 ± 0.3 | 102.4 ± 2.5 | 122.8 ± 7.3 | A | |
| Wheat | − | 147.3 ± 8.2 | 8.1 ± 0.2 | 96.9 ± 4.1 | 125.3 ± 9.0 | A |
| + | 158.8 ± 10.3 | 8.0 ± 0.3 | 96.4 ± 3.1 | 95.5 ± 12.1 | B |
| Shoot Wet Weight (g) | Cob’s Wet Weight (g) | ||||
|---|---|---|---|---|---|
| Tillage | 2019 | 2020 | 2019 | 2020 | |
| Clover | − | 118% | 112% | 115% | 106% |
| + | 114% | 107% | 122% | 104% | |
| Wheat | − | 147% | 107% | 146% | 106% |
| + | 154% | 116% | 136% | 81% | |
| Diseased Plants | M. maydis DNA | ||||
|---|---|---|---|---|---|
| Tillage | 2019 | 2020 | 2019 | 2020 | |
| Clover | − | 65% | 109% | 89% | 2% |
| + | 81% | 78% | 394% | 1% | |
| Wheat | − | 27% | 87% | 18% | 2% |
| + | 27% | 43% | 36% | 4% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Degani, O.; Gordani, A.; Becher, P.; Dor, S. Crop Cycle and Tillage Role in the Outbreak of Late Wilt Disease of Maize Caused by Magnaporthiopsis maydis. J. Fungi 2021, 7, 706. https://doi.org/10.3390/jof7090706
Degani O, Gordani A, Becher P, Dor S. Crop Cycle and Tillage Role in the Outbreak of Late Wilt Disease of Maize Caused by Magnaporthiopsis maydis. Journal of Fungi. 2021; 7(9):706. https://doi.org/10.3390/jof7090706
Chicago/Turabian StyleDegani, Ofir, Asaf Gordani, Paz Becher, and Shlomit Dor. 2021. "Crop Cycle and Tillage Role in the Outbreak of Late Wilt Disease of Maize Caused by Magnaporthiopsis maydis" Journal of Fungi 7, no. 9: 706. https://doi.org/10.3390/jof7090706
APA StyleDegani, O., Gordani, A., Becher, P., & Dor, S. (2021). Crop Cycle and Tillage Role in the Outbreak of Late Wilt Disease of Maize Caused by Magnaporthiopsis maydis. Journal of Fungi, 7(9), 706. https://doi.org/10.3390/jof7090706