
Chronic Occupational Mold Exposure Drives Expansion of Aspergillus-Reactive Type 1 and Type 2 T-Helper Cell Responses

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Blood Collection and PBMC Isolation
2.3. Aspergillus Fumigatus Mycelial Lysate and Recombinant Antigen Generation
2.4. ELISPOT
2.5. Flow Cytometry
2.6. Whole-Blood Stimulation for Multiplex Cytokine Assays
2.7. Statistical Analyses
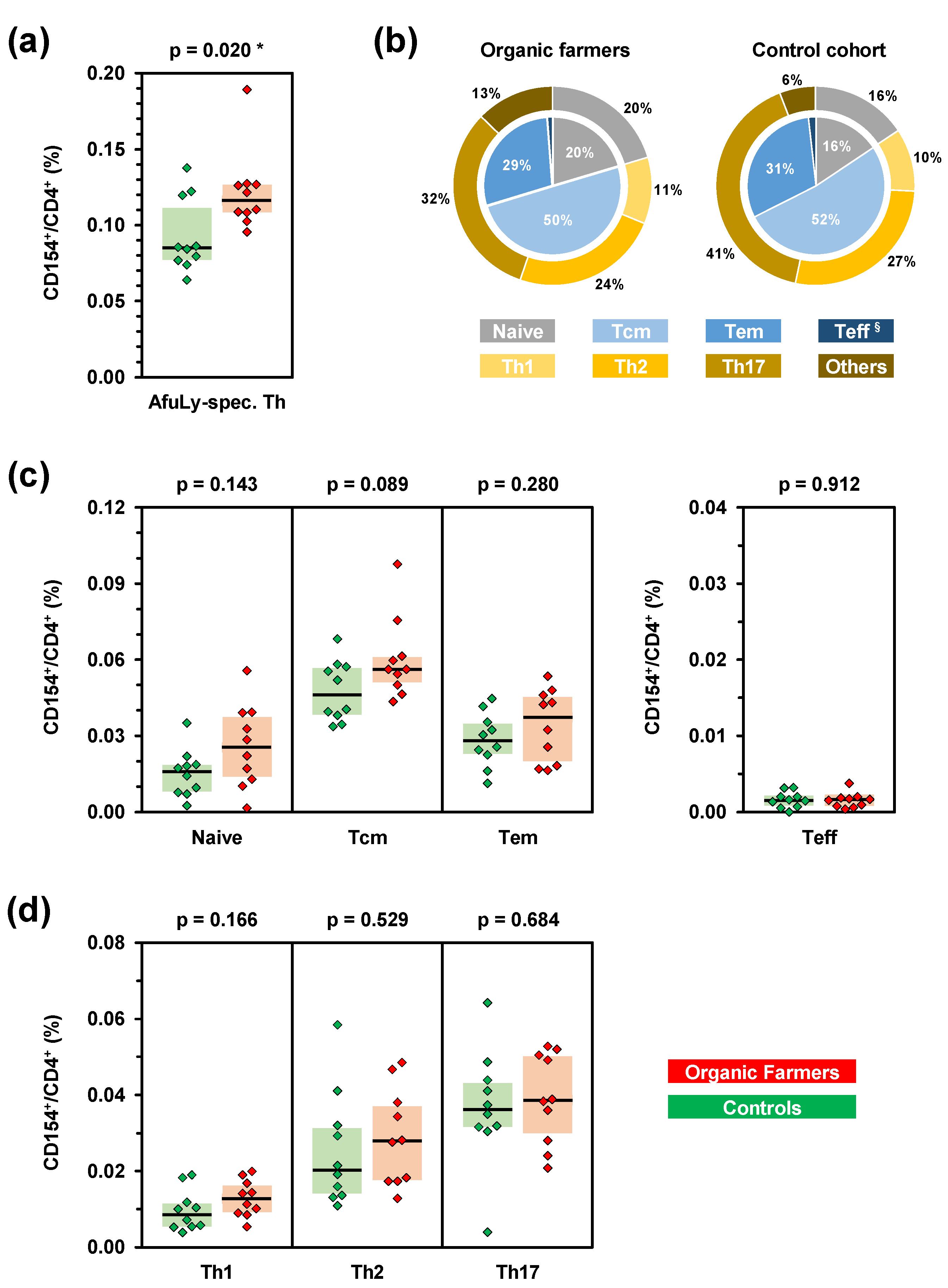
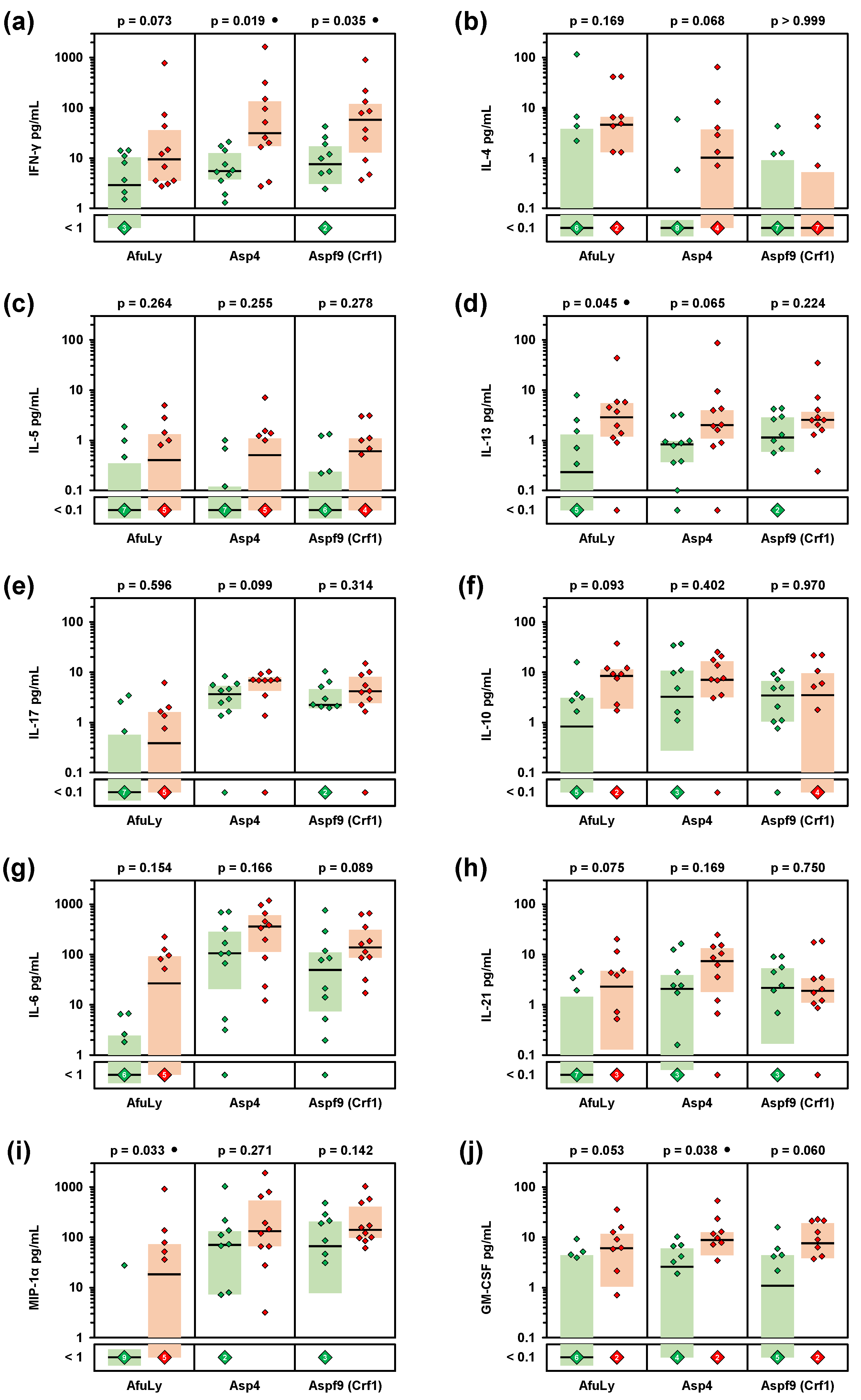
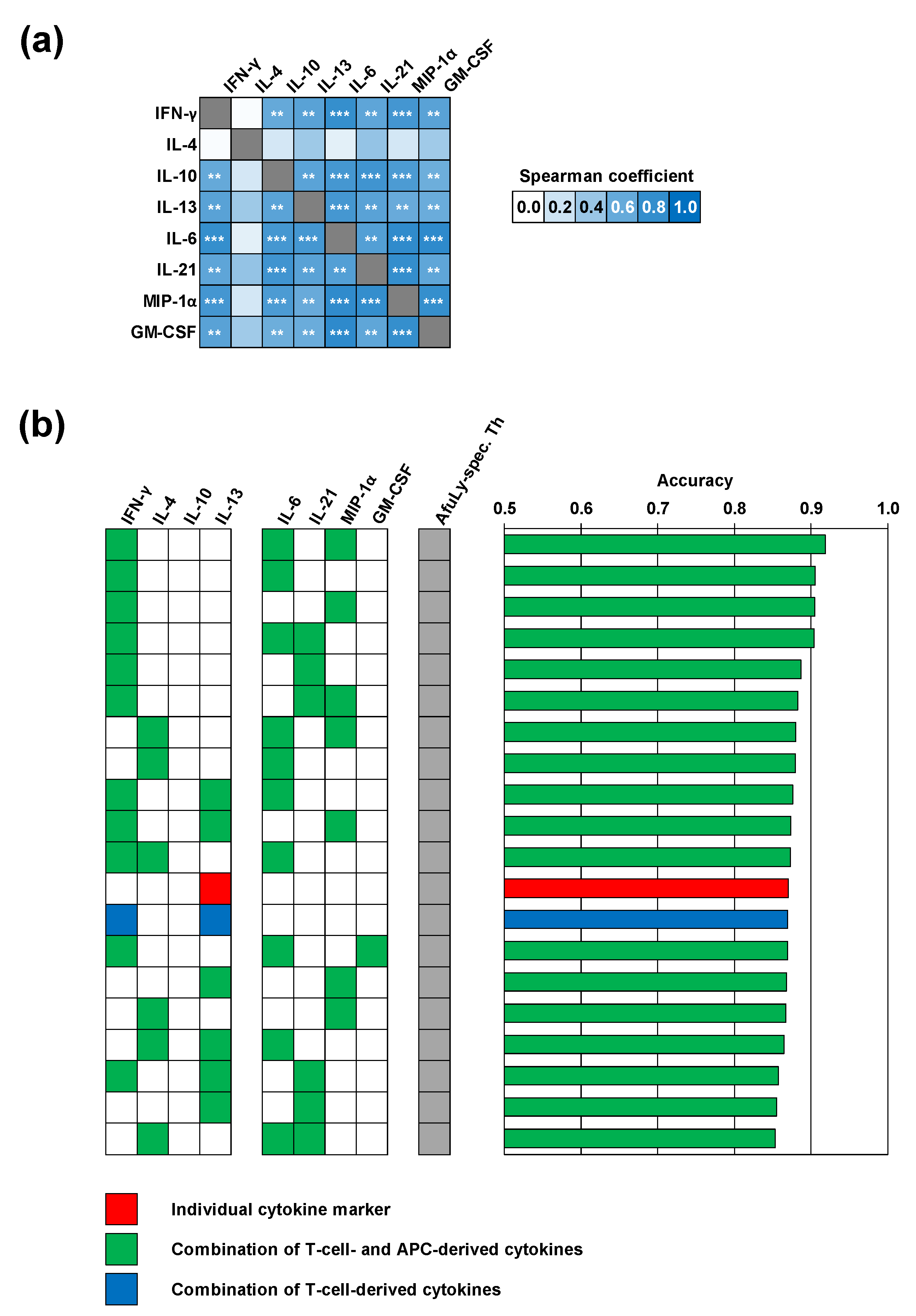
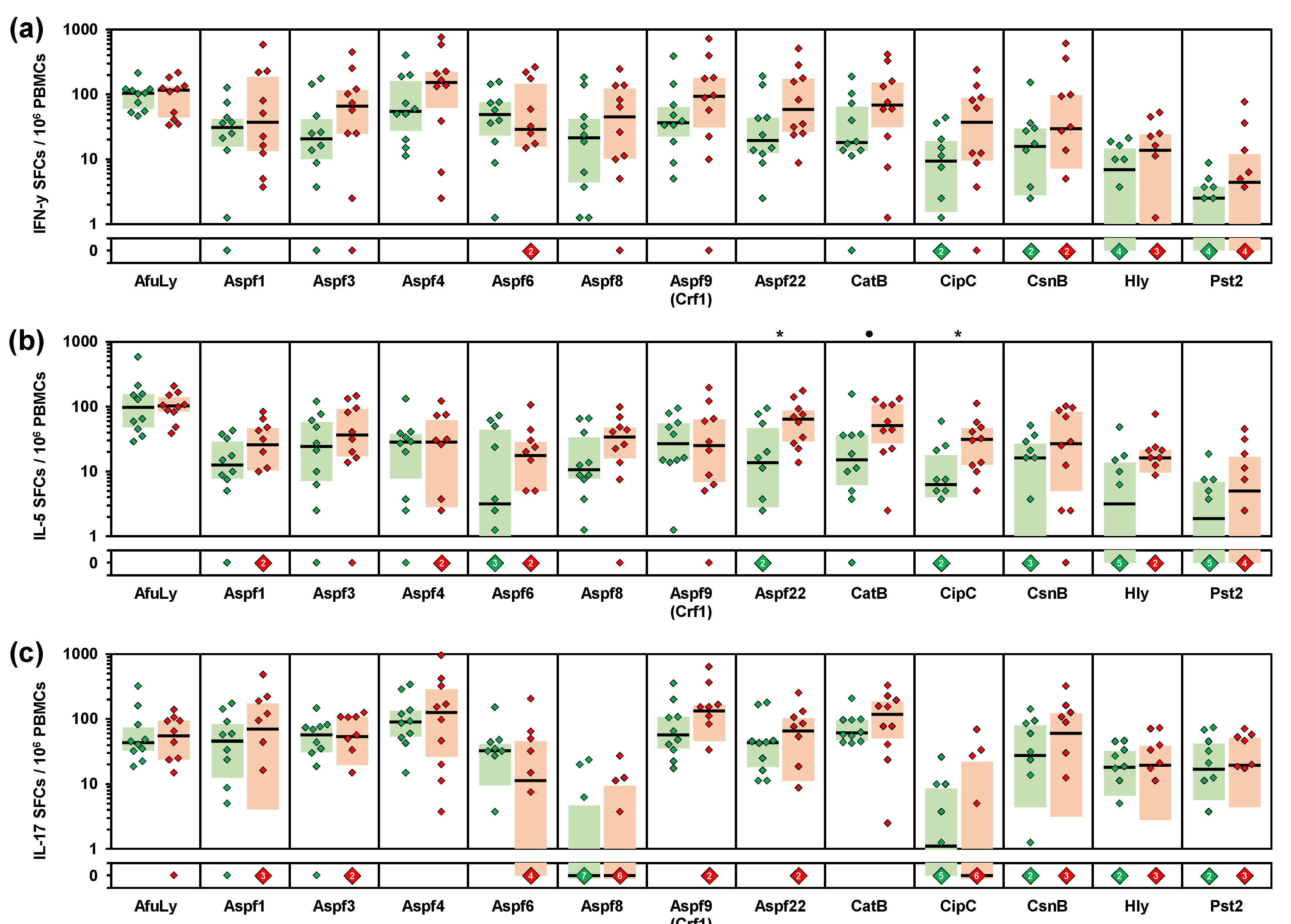
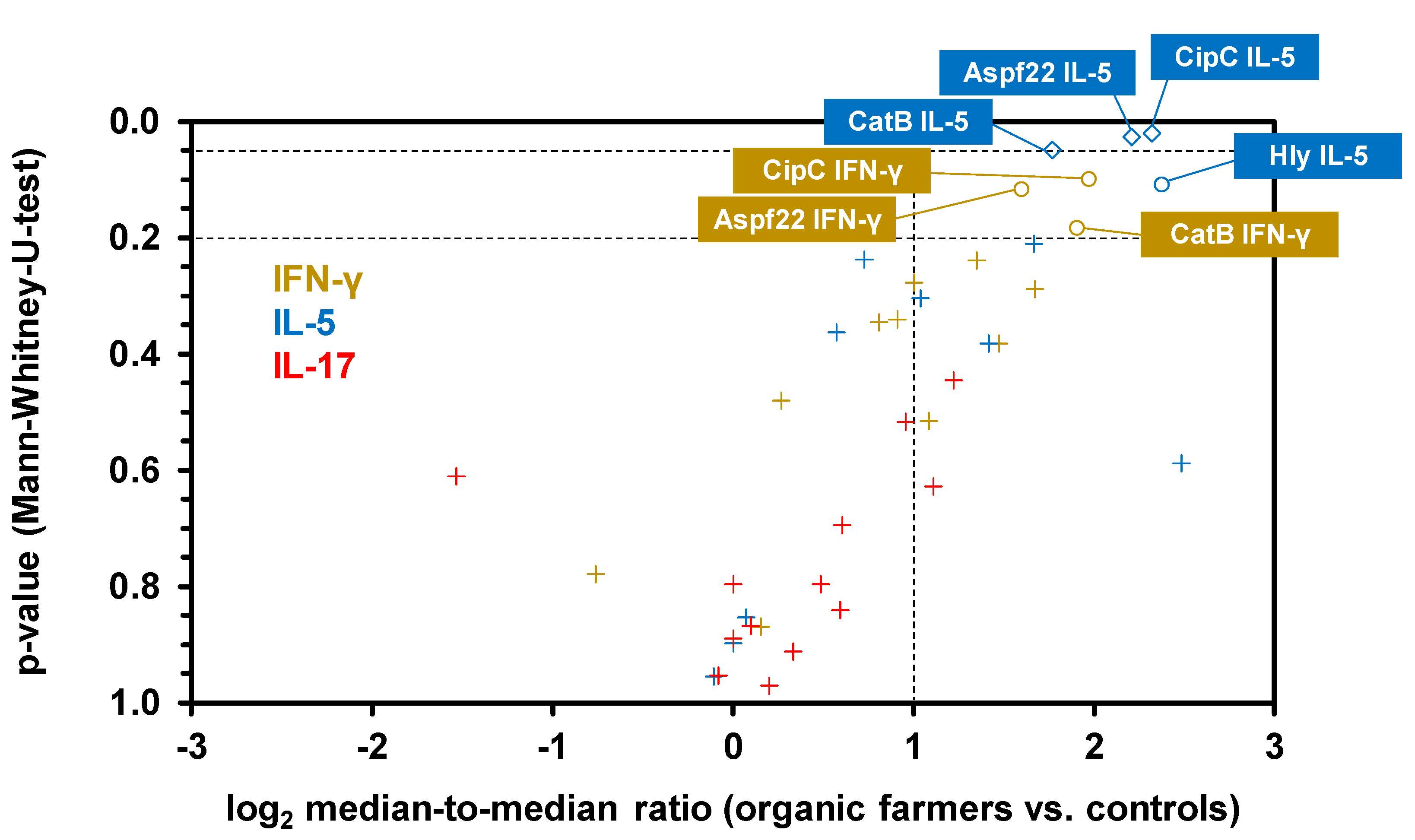
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, S.J.; Mehrad, B. Innate immunity to Aspergillus species. Clin. Microbiol. Rev. 2009, 22, 535–551. [Google Scholar] [CrossRef]
- Baxi, S.N.; Portnoy, J.M.; Larenas-Linnemann, D.; Phipatanakul, W.; Environmental Allergens, W. Exposure and Health Effects of Fungi on Humans. J. Allergy Clin. Immunol. Pract. 2016, 4, 396–404. [Google Scholar] [CrossRef]
- Twaroch, T.E.; Curin, M.; Valenta, R.; Swoboda, I. Mold allergens in respiratory allergy: From structure to therapy. Allergy Asthma Immunol. Res. 2015, 7, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Latge, J.P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33. [Google Scholar] [CrossRef]
- Kotimaa, M.H.; Husman, K.H.; Terho, E.O.; Mustonen, M.H. Airborne molds and actinomycetes in the work environment of farmer’s lung patients in Finland. Scand. J. Work Environ. Health 1984, 10, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, D.; Fu, S.; Ren, Y.; Wang, L.; Zhang, Y.; Zhao, M.; He, X.; Wang, X. Prevalence and risk factors for farmer’s lung in greenhouse farmers: An epidemiological study of 5880 farmers from Northeast China. Cell Biochem. Biophys. 2015, 71, 1051–1057. [Google Scholar] [CrossRef]
- Halpin, D.M.; Graneek, B.J.; Turner-Warwick, M.; Newman Taylor, A.J. Extrinsic allergic alveolitis and asthma in a sawmill worker: Case report and review of the literature. Occup. Environ. Med. 1994, 51, 160–164. [Google Scholar] [CrossRef]
- Faerden, K.; Lund, M.B.; Mogens Aalokken, T.; Eduard, W.; Sostrand, P.; Langard, S.; Kongerud, J. Hypersensitivity pneumonitis in a cluster of sawmill workers: A 10-year follow-up of exposure, symptoms, and lung function. Int. J. Occup Environ. Health 2014, 20, 167–173. [Google Scholar] [CrossRef]
- Page, L.; Weis, P.; Müller, T.; Dittrich, M.; Lazariotou, M.; Dragan, M.; Waaga-Gasser, A.M.; Helm, J.; Dandekar, T.; Einsele, H.; et al. Evaluation of Aspergillus and Mucorales specific T-cells and peripheral blood mononuclear cell cytokine signatures as biomarkers of environmental mold exposure. Int. J. Med Microbiol. IJMM 2018, 308, 1018–1026. [Google Scholar] [CrossRef]
- Wurster, S.; Weis, P.; Page, L.; Helm, J.; Lazariotou, M.; Einsele, H.; Ullmann, A.J. Intra- and inter-individual variability of Aspergillus fumigatus reactive T-cell frequencies in healthy volunteers in dependency of mould exposure in residential and working environment. Mycoses 2017, 60, 668–675. [Google Scholar] [CrossRef]
- Rosenblum Lichtenstein, J.H.; Hsu, Y.H.; Gavin, I.M.; Donaghey, T.C.; Molina, R.M.; Thompson, K.J.; Chi, C.L.; Gillis, B.S.; Brain, J.D. Environmental mold and mycotoxin exposures elicit specific cytokine and chemokine responses. PLoS ONE 2015, 10, e0126926. [Google Scholar] [CrossRef]
- Lauruschkat, C.D.; Page, L.; White, P.L.; Etter, S.; Davies, H.E.; Duckers, J.; Ebel, F.; Schnack, E.; Backx, M.; Dragan, M.; et al. Development of a Simple and Robust Whole Blood Assay with Dual Co-Stimulation to Quantify the Release of T-Cellular SignatureCytokines in Response to Aspergillus fumigatus Antigens. J. Fungi 2021, 7, 462. [Google Scholar] [CrossRef]
- Sabino, R.; Faisca, V.M.; Carolino, E.; Verissimo, C.; Viegas, C. Occupational exposure to Aspergillus by swine and poultry farm workers in Portugal. J. Toxicol. Environ. Health A 2012, 75, 1381–1391. [Google Scholar] [CrossRef]
- Barber, A.E.; Riedel, J.; Sae-Ong, T.; Kang, K.; Brabetz, W.; Panagiotou, G.; Deising, H.B.; Kurzai, O. Effects of Agricultural Fungicide Use on Aspergillus fumigatus Abundance, Antifungal Susceptibility, and Population Structure. mBio 2020, 11. [Google Scholar] [CrossRef]
- Richardson, M.D.; Page, I.D. Aspergillus serology: Have we arrived yet? Med. Mycol. 2017, 55, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Lauruschkat, C.D.; Page, L.; Etter, S.; Schnack, E.; Einsele, H.; Ebel, F.; Loeffler, J.; Wurster, S. Establishment of a whole blood ELISA to quantify T-cellular cytokine release in response to Aspergillus fumigatus antigens. In Proceedings of the Advances Against Aspergillosis and Mucormycosis, Lugano, Switzerland, 27–29 February 2020. [Google Scholar]
- Lauruschkat, C.D.; Wurster, S.; Page, L.; Lazariotou, M.; Dragan, M.; Weis, P.; Ullmann, A.J.; Einsele, H.; Loffler, J. Susceptibility of A. fumigatus-specific T-cell assays to pre-analytic blood storage and PBMC cryopreservation greatly depends on readout platform and analytes. Mycoses 2018, 61, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Weis, P.; Helm, J.; Page, L.; Lauruschkat, C.D.; Lazariotou, M.; Einsele, H.; Loeffler, J.; Ullmann, A.J.; Wurster, S. Development and evaluation of a whole blood-based approach for flow cytometric quantification of CD154+ mould-reactive T cells. Med. Mycol. 2020, 58, 187–196. [Google Scholar] [CrossRef]
- Banerjee, B.; Kurup, V.P.; Greenberger, P.A.; Johnson, B.D.; Fink, J.N. Cloning and expression of Aspergillus fumigatus allergen Asp f 16 mediating both humoral and cell-mediated immunity in allergic bronchopulmonary aspergillosis (ABPA). Clin. Exp. Allergy 2001, 31, 761–770. [Google Scholar] [CrossRef]
- Ramadan, G.; Davies, B.; Kurup, V.P.; Keever-Taylor, C.A. Generation of cytotoxic T cell responses directed to human leucocyte antigen Class I restricted epitopes from the Aspergillus f16 allergen. Clin. Exp. Immunol. 2005, 140, 81–91. [Google Scholar] [CrossRef]
- Jolink, H.; de Boer, R.; Willems, L.N.; van Dissel, J.T.; Falkenburg, J.H.; Heemskerk, M.H. T helper 2 response in allergic bronchopulmonary aspergillosis is not driven by specific Aspergillus antigens. Allergy 2015, 70, 1336–1339. [Google Scholar] [CrossRef]
- Hemmann, S.; Nikolaizik, W.H.; Schoni, M.H.; Blaser, K.; Crameri, R. Differential IgE recognition of recombinant Aspergillus fumigatus allergens by cystic fibrosis patients with allergic bronchopulmonary aspergillosis or Aspergillus allergy. Eur. J. Immunol. 1998, 28, 1155–1160. [Google Scholar] [CrossRef]
- Kurup, V.P.; Xia, J.Q.; Crameri, R.; Rickaby, D.A.; Choi, H.Y.; Fluckiger, S.; Blaser, K.; Dawson, C.A.; Kelly, K.J. Purified recombinant A. fumigatus allergens induce different responses in mice. Clin. Immunol. 2001, 98, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Wingard, J.R.; Cheng, S.; Nguyen, M.H.; Clancy, C.J. Serum IgG responses against Aspergillus proteins before hematopoietic stem cell transplantation or chemotherapy identify patients who develop invasive aspergillosis. Biol. Blood Marrow Transplant. 2012, 18, 1927–1934. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bacher, P.; Heinrich, F.; Stervbo, U.; Nienen, M.; Vahldieck, M.; Iwert, C.; Vogt, K.; Kollet, J.; Babel, N.; Sawitzki, B.; et al. Regulatory T Cell Specificity Directs Tolerance versus Allergy against Aeroantigens in Humans. Cell 2016, 167, 1067–1078. [Google Scholar] [CrossRef]
- Murdock, B.J.; Shreiner, A.B.; McDonald, R.A.; Osterholzer, J.J.; White, E.S.; Toews, G.B.; Huffnagle, G.B. Coevolution of TH1, TH2, and TH17 responses during repeated pulmonary exposure to Aspergillus fumigatus conidia. Infect. Immun. 2011, 79, 125–135. [Google Scholar] [CrossRef]
- Schmidt, S.; Ebner, F.; Rosen, K.; Kniemeyer, O.; Brakhage, A.A.; Loffler, J.; Seif, M.; Springer, J.; Schlosser, J.; Scharek-Tedin, L.; et al. The domestic pig as human-relevant large animal model to study adaptive antifungal immune responses against airborne Aspergillus fumigatus. Eur. J. Immunol. 2020, 50, 1712–1728. [Google Scholar] [CrossRef]
- Bacher, P.; Hohnstein, T.; Beerbaum, E.; Rocker, M.; Blango, M.G.; Kaufmann, S.; Rohmel, J.; Eschenhagen, P.; Grehn, C.; Seidel, K.; et al. Human Anti-fungal Th17 Immunity and Pathology Rely on Cross-Reactivity against Candida albicans. Cell 2019, 176, 1340–1355. [Google Scholar] [CrossRef]
- Werner, J.L.; Metz, A.E.; Horn, D.; Schoeb, T.R.; Hewitt, M.M.; Schwiebert, L.M.; Faro-Trindade, I.; Brown, G.D.; Steele, C. Requisite role for the dectin-1 beta-glucan receptor in pulmonary defense against Aspergillus fumigatus. J. Immunol. 2009, 182, 4938–4946. [Google Scholar] [CrossRef]
- Rivera, A.; Hohl, T.M.; Collins, N.; Leiner, I.; Gallegos, A.; Saijo, S.; Coward, J.W.; Iwakura, Y.; Pamer, E.G. Dectin-1 diversifies Aspergillus fumigatus-specific T cell responses by inhibiting T helper type 1 CD4 T cell differentiation. J. Exp. Med. 2011, 208, 369–381. [Google Scholar] [CrossRef]
- Gresnigt, M.S.; Bozza, S.; Becker, K.L.; Joosten, L.A.; Abdollahi-Roodsaz, S.; van der Berg, W.B.; Dinarello, C.A.; Netea, M.G.; Fontaine, T.; De Luca, A.; et al. A polysaccharide virulence factor from Aspergillus fumigatus elicits anti-inflammatory effects through induction of Interleukin-1 receptor antagonist. PLoS Pathog. 2014, 10, e1003936. [Google Scholar] [CrossRef]
- Fontaine, T.; Delangle, A.; Simenel, C.; Coddeville, B.; van Vliet, S.J.; van Kooyk, Y.; Bozza, S.; Moretti, S.; Schwarz, F.; Trichot, C.; et al. Galactosaminogalactan, a new immunosuppressive polysaccharide of Aspergillus fumigatus. PLoS Pathog. 2011, 7, e1002372. [Google Scholar] [CrossRef]
- Simonian, P.L.; Roark, C.L.; Born, W.K.; O’Brien, R.L.; Fontenot, A.P. Gammadelta T cells and Th17 cytokines in hypersensitivity pneumonitis and lung fibrosis. Transl Res. 2009, 154, 222–227. [Google Scholar] [CrossRef][Green Version]
- Simonian, P.L.; Roark, C.L.; Wehrmann, F.; Lanham, A.K.; Diaz del Valle, F.; Born, W.K.; O’Brien, R.L.; Fontenot, A.P. Th17-polarized immune response in a murine model of hypersensitivity pneumonitis and lung fibrosis. J. Immunol. 2009, 182, 657–665. [Google Scholar] [CrossRef]
- Simonian, P.L.; Roark, C.L.; Wehrmann, F.; Lanham, A.M.; Born, W.K.; O’Brien, R.L.; Fontenot, A.P. IL-17A-expressing T cells are essential for bacterial clearance in a murine model of hypersensitivity pneumonitis. J. Immunol. 2009, 182, 6540–6549. [Google Scholar] [CrossRef]
- Andrews, K.; Ghosh, M.C.; Schwingshackl, A.; Rapalo, G.; Luellen, C.; Waters, C.M.; Fitzpatrick, E.A. Chronic hypersensitivity pneumonitis caused by Saccharopolyspora rectivirgula is not associated with a switch to a Th2 response. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 310, L393–L402. [Google Scholar] [CrossRef]
- Hasan, S.A.; Eksteen, B.; Reid, D.; Paine, H.V.; Alansary, A.; Johannson, K.; Gwozd, C.; Goring, K.A.; Vo, T.; Proud, D.; et al. Role of IL-17A and neutrophils in fibrosis in experimental hypersensitivity pneumonitis. J. Allergy Clin. Immunol. 2013, 131, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.L.; Gresnigt, M.S.; Smeekens, S.P.; Jacobs, C.W.; Magis-Escurra, C.; Jaeger, M.; Wang, X.; Lubbers, R.; Oosting, M.; Joosten, L.A.; et al. Pattern recognition pathways leading to a Th2 cytokine bias in allergic bronchopulmonary aspergillosis patients. Clin. Exp. Allergy 2015, 45, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Moss, R.B. Pathophysiology and immunology of allergic bronchopulmonary aspergillosis. Med. Mycol. 2005, 43 (Suppl. S1), S203–S206. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, I.S.; Agarwal, R. Specific IgE is better than skin testing for detecting Aspergillus sensitization and allergic bronchopulmonary aspergillosis in asthma. Chest 2015, 147, e194. [Google Scholar] [CrossRef]
- de Barros Bezerra, G.F.; Haidar, D.M.; da Silva, M.A.; Filho, W.E.; Dos Santos, R.M.; Rosa, I.G.; de Castro Viana, G.M.; Zaror, L.; Soares Brandao Nascimento Mdo, D. IgE serum concentration against airborne fungi in children with respiratory allergies. Allergy Asthma Clin. Immunol. 2016, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Neveu, W.A.; Allard, J.B.; Dienz, O.; Wargo, M.J.; Ciliberto, G.; Whittaker, L.A.; Rincon, M. IL-6 is required for airway mucus production induced by inhaled fungal allergens. J. Immunol. 2009, 183, 1732–1738. [Google Scholar] [CrossRef]
- Rincon, M.; Anguita, J.; Nakamura, T.; Fikrig, E.; Flavell, R.A. Interleukin (IL)-6 directs the differentiation of IL-4-producing CD4+ T cells. J. Exp. Med. 1997, 185, 461–469. [Google Scholar] [CrossRef]
- Diehl, S.; Rincon, M. The two faces of IL-6 on Th1/Th2 differentiation. Mol. Immunol. 2002, 39, 531–536. [Google Scholar] [CrossRef]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006, 441, 235–238. [Google Scholar] [CrossRef]
- Kimura, A.; Kishimoto, T. IL-6: Regulator of Treg/Th17 balance. Eur. J. Immunol. 2010, 40, 1830–1835. [Google Scholar] [CrossRef]
- Olszewski, M.A.; Huffnagle, G.B.; McDonald, R.A.; Lindell, D.M.; Moore, B.B.; Cook, D.N.; Toews, G.B. The role of macrophage inflammatory protein-1 alpha/CCL3 in regulation of T cell-mediated immunity to Cryptococcus neoformans infection. J. Immunol. 2000, 165, 6429–6436. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S.J.; Pesenacker, A.M.; Wang, A.Y.; Gillies, J.; Mojibian, M.; Morishita, K.; Tan, R.; Kieffer, T.J.; Verchere, C.B.; Panagiotopoulos, C.; et al. T regulatory cell chemokine production mediates pathogenic T cell attraction and suppression. J. Clin. Investig. 2016, 126, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Spolski, R.; Feng, C.G.; Qi, C.F.; Cheng, J.; Sher, A.; Morse, H.C., 3rd; Liu, C.; Schwartzberg, P.L.; Leonard, W.J. A critical role for IL-21 in regulating immunoglobulin production. Science 2002, 298, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Suto, A.; Nakajima, H.; Hirose, K.; Suzuki, K.; Kagami, S.; Seto, Y.; Hoshimoto, A.; Saito, Y.; Foster, D.C.; Iwamoto, I. Interleukin 21 prevents antigen-induced IgE production by inhibiting germ line C(epsilon) transcription of IL-4-stimulated B cells. Blood 2002, 100, 4565–4573. [Google Scholar] [CrossRef]
- Lin, P.Y.; Jen, H.Y.; Chiang, B.L.; Sheu, F.; Chuang, Y.H. Interleukin-21 suppresses the differentiation and functions of T helper 2 cells. Immunology 2015, 144, 668–676. [Google Scholar] [CrossRef]
- Spolski, R.; Kim, H.P.; Zhu, W.; Levy, D.E.; Leonard, W.J. IL-21 mediates suppressive effects via its induction of IL-10. J. Immunol. 2009, 182, 2859–2867. [Google Scholar] [CrossRef] [PubMed]
- Pot, C.; Jin, H.; Awasthi, A.; Liu, S.M.; Lai, C.Y.; Madan, R.; Sharpe, A.H.; Karp, C.L.; Miaw, S.C.; Ho, I.C.; et al. Cutting edge: IL-27 induces the transcription factor c-Maf, cytokine IL-21, and the costimulatory receptor ICOS that coordinately act together to promote differentiation of IL-10-producing Tr1 cells. J. Immunol. 2009, 183, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Doganci, A.; Birkholz, J.; Gehring, S.; Puhl, A.G.; Zepp, F.; Meyer, C.U. In the presence of IL-21 human cord blood T cells differentiate to IL-10-producing Th1 but not Th17 or Th2 cells. Int. Immunol. 2013, 25, 157–169. [Google Scholar] [CrossRef]
- Bush, R.K.; Portnoy, J.M.; Saxon, A.; Terr, A.I.; Wood, R.A. The medical effects of mold exposure. J. Allergy Clin. Immunol. 2006, 117, 326–333. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauruschkat, C.D.; Etter, S.; Schnack, E.; Ebel, F.; Schäuble, S.; Page, L.; Rümens, D.; Dragan, M.; Schlegel, N.; Panagiotou, G.; et al. Chronic Occupational Mold Exposure Drives Expansion of Aspergillus-Reactive Type 1 and Type 2 T-Helper Cell Responses. J. Fungi 2021, 7, 698. https://doi.org/10.3390/jof7090698
Lauruschkat CD, Etter S, Schnack E, Ebel F, Schäuble S, Page L, Rümens D, Dragan M, Schlegel N, Panagiotou G, et al. Chronic Occupational Mold Exposure Drives Expansion of Aspergillus-Reactive Type 1 and Type 2 T-Helper Cell Responses. Journal of Fungi. 2021; 7(9):698. https://doi.org/10.3390/jof7090698
Chicago/Turabian StyleLauruschkat, Chris D., Sonja Etter, Elisabeth Schnack, Frank Ebel, Sascha Schäuble, Lukas Page, Dana Rümens, Mariola Dragan, Nicolas Schlegel, Gianni Panagiotou, and et al. 2021. "Chronic Occupational Mold Exposure Drives Expansion of Aspergillus-Reactive Type 1 and Type 2 T-Helper Cell Responses" Journal of Fungi 7, no. 9: 698. https://doi.org/10.3390/jof7090698
APA StyleLauruschkat, C. D., Etter, S., Schnack, E., Ebel, F., Schäuble, S., Page, L., Rümens, D., Dragan, M., Schlegel, N., Panagiotou, G., Kniemeyer, O., Brakhage, A. A., Einsele, H., Wurster, S., & Loeffler, J. (2021). Chronic Occupational Mold Exposure Drives Expansion of Aspergillus-Reactive Type 1 and Type 2 T-Helper Cell Responses. Journal of Fungi, 7(9), 698. https://doi.org/10.3390/jof7090698