
Extraction, Structural Characterisation, and Immunomodulatory Properties of Edible Amanita hemibapha subspecies javanica (Corner and Bas) Mucilage Polysaccharide as a Potential of Functional Food

 ,
,  ,
, 


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials, and Reagents
2.2. Mucilage Polysaccharide Extraction
2.3. Fractionation of MPs
2.4. Chemical Composition of Polysaccharides
2.5. Analysis of Monosaccharide Composition, and Absolute Configuration
2.6. Methylation Analysis
2.7. NMR Spectroscopy
2.8. RAW264.7 Macrophage Proliferation, and Nitric Oxide (NO) Production Assay
2.9. Gene Expression by RT-PCR
2.10. Western Blot Analysis
2.11. Investigation of Cell Membrane Binding Receptors
2.12. Statistical Analysis
3. Results and Discussion
3.1. Yield, and Chemical Composition
3.2. Molecular Characteristics of MPs, and MPFs
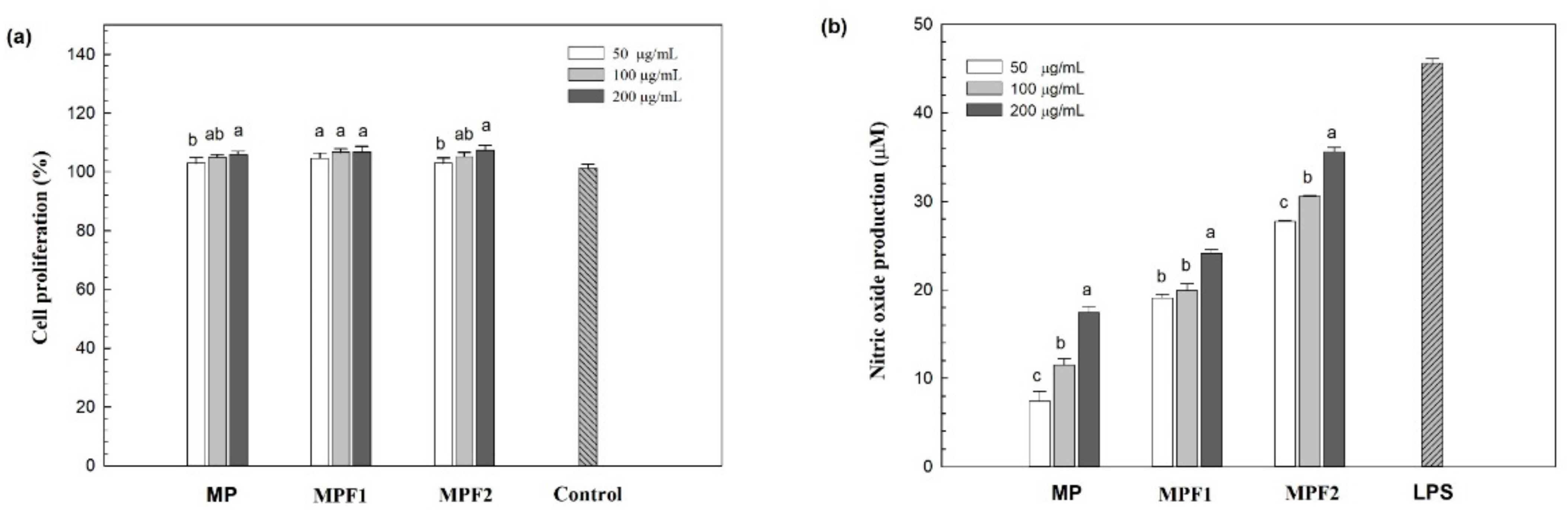
3.3. Immunomodulatory Activity of MPs, and MPFs
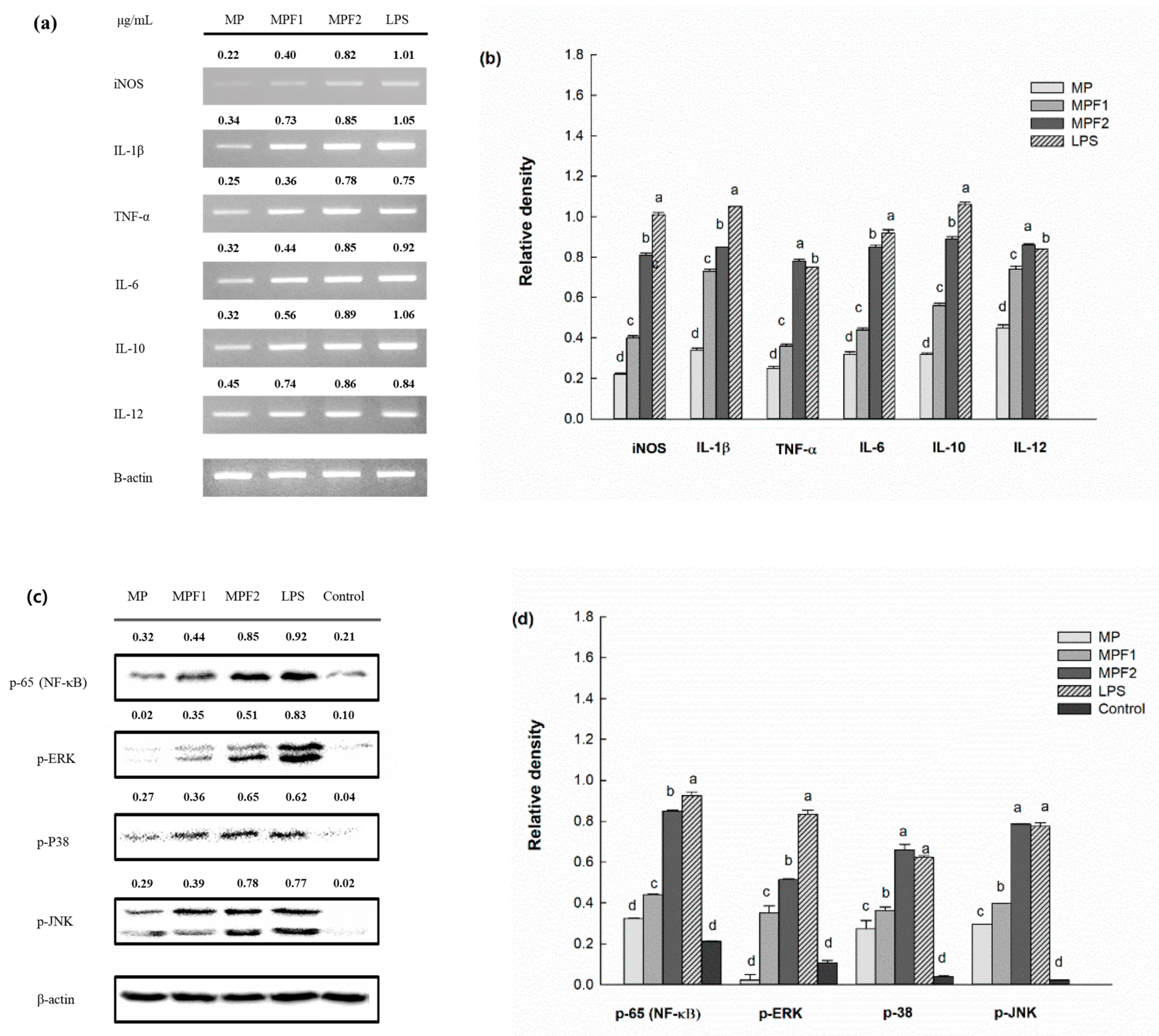
3.4. Effect of MP, and MPF Stimulation on Cytokine mRNA Expression
3.5. Immunomodulatory Mechanism by Western Blot Analysis
3.6. MPF2 on Binding Receptors
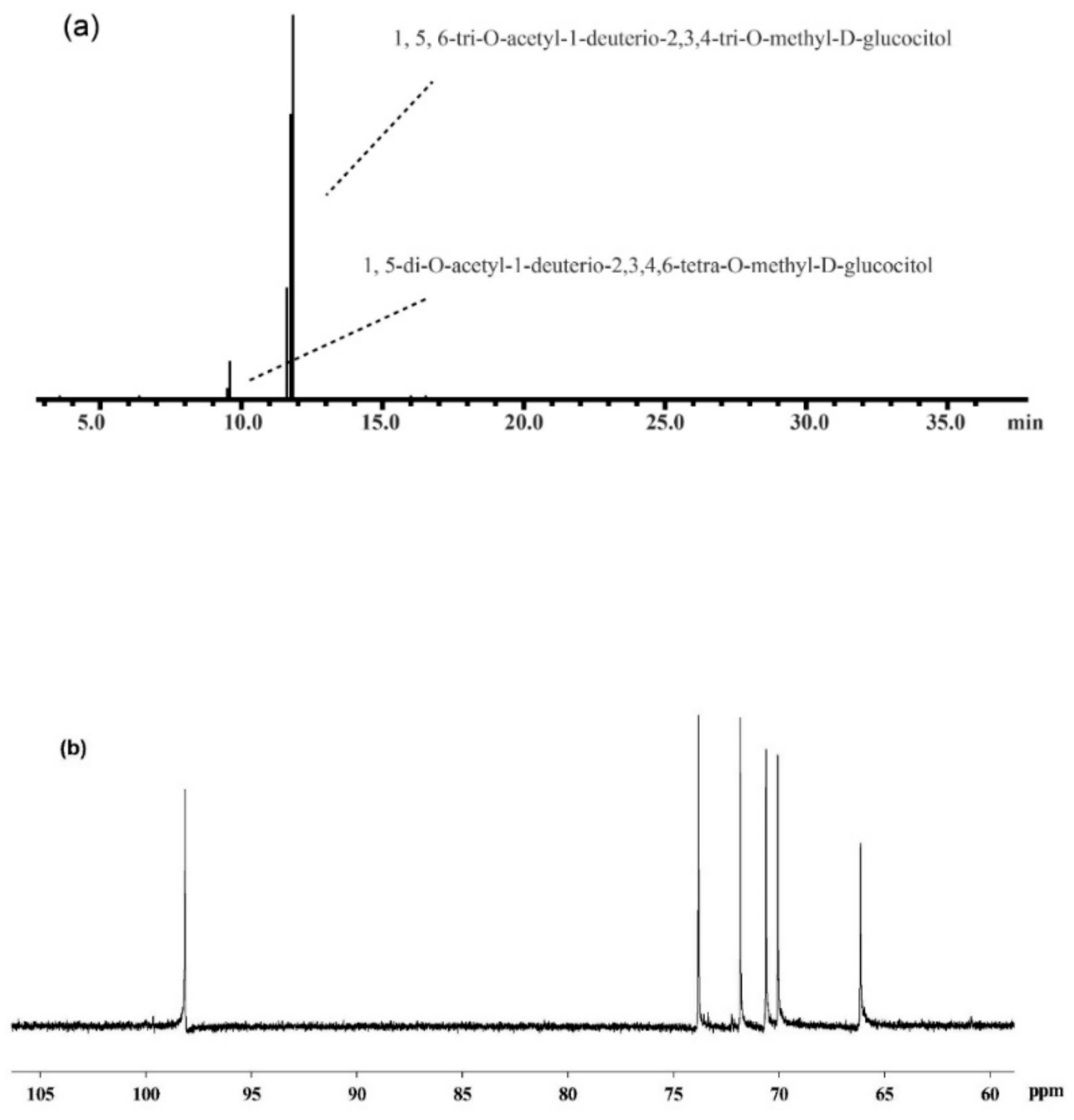
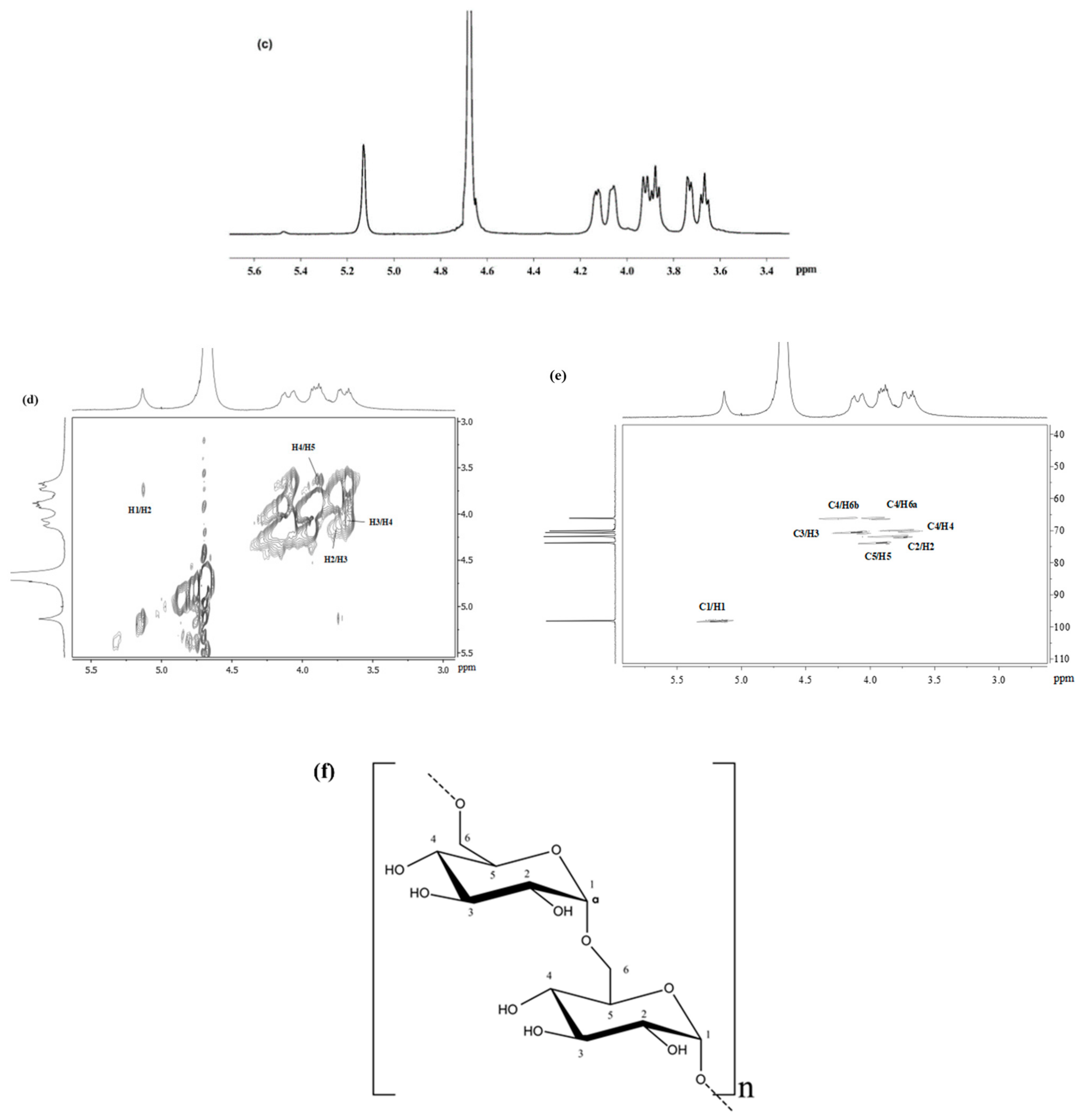
3.7. Structural Analysis of MPF2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roselló-Soto, E.; Parniakov, O.; Deng, Q.; Patras, A.; Koubaa, M.; Grimi, N.; Boussetta, N.; Tiwari, B.K.; Vorobiev, E.; Lebovka, N.; et al. Application of non-conventional extraction methods: Toward a sustainable and green production of valuable compounds from mushrooms. Food. Eng. Rev. 2016, 8, 214–234. [Google Scholar] [CrossRef]
- Lu, X.; Brennan, M.A.; Seventi, L.; Mason, S.; Brennan, C.S. How the inclusion of mushroom powder can affect the physicochemical characteristics of pasta. Int. J. Food Sci. Technol. 2016, 51, 2433–2439. [Google Scholar] [CrossRef]
- Novakovic, S.; Djekic, I.; Vunduk, J.; Klaus, A.; Lorenzo, J.M.; Barba, F.J.; Tomasevic, I. An insight into in vitro antioxidant activity of Cantharellus cibarius hot water extract for the potential application in meat products. IOP Conf. Ser. Earth. Environ. Sci. 2019, 333, 012089. [Google Scholar] [CrossRef]
- Lu, X.; Brennan, M.A.; Guan, W.; Zhang, J.; Yuan, L.; Brennan, C.S. Enhancing the nutritional properties of bread by incorporating mushroom bioactive compounds: The manipulation of the predictive glycaemic response and the phenolic properties. Foods. 2021, 10, 731. [Google Scholar] [CrossRef]
- Cheung, P.C.K. Nutritional value and health benefits of mushrooms. In Mushrooms as Functional Foods; Cheung, P., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 71–109. [Google Scholar]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Wei, Y.; Ouyang, Z.; Su, Z. Polysaccharides purified from Cordyceps cicadae protects PC12 cells against glutamate-induced oxidative damage. Carbohydr. Polym. 2016, 153, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Cabello, K.P.; Lugo-Flores, M.A.; Rivera-Palafox, P.; Silva-Espinoza, B.A.; González-Aguilar, G.A.; Esqueda, M.; Gaitán-Hernández, R.; Ayala-Zavala, J.F. Antioxidant Properties and Industrial Uses of Edible Polyporales. J. Fungi. 2021, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yan, X.; Song, Z.; Li, W.; Zhao, W.; Ma, H.; Du, J.; Li, S.; Zhang, D. Two heteropolysaccharides from Isaria cicadae Miquel differ in composition and potentially immunomodulatory activity. Int. J. Biol. Macromol. 2018, 117, 610–616. [Google Scholar] [CrossRef]
- Prasain, J.K. Pharmacological effects of cordyceps and its bioactive compounds. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; Volume 40, pp. 453–468. [Google Scholar]
- Olawuyi, I.F.; Lee, W.Y. Structural characterization, functional properties and antioxidant activities of polysaccharide extract obtained from okra leaves (Abelmoschus esculentus). Food Chem. 2021, 354, 129437. [Google Scholar] [CrossRef]
- Alvarado-Morales, G.; Minjares-Fuentes, R.; Contreras-Esquivel, J.C.; Montañez, J.; Meza-Velázquez, J.A.; Femenia, A. Application of thermosonication for Aloe vera (Aloe barbadensis Miller) juice processing: Impact on the functional properties and the main bioactive polysaccharides. Ultrason. Sonochem. 2019, 56, 125–133. [Google Scholar] [CrossRef]
- Nerkar, P.P.; Gattani, S.G. Oromucosal delivery of venlafaxine by linseed mucilage based gel: In vitro and in vivo evaluation in rabbits. Arch. Pharm. Res. 2013, 36, 846–853. [Google Scholar] [CrossRef]
- Surin, S.; Surayot, U.; Seesuriyachan, P.; You, S.G.; Phimolsiripol, Y. Antioxidant and immunomodulatory activities of sulphated polysaccharides from purple glutinous rice bran (Oryza sativa L.). Int. J. Food Sci. Technol. 2018, 53, 994–1004. [Google Scholar] [CrossRef] [Green Version]
- Surin, S.; You, S.G.; Seesuriyachan, P.; Muangrat, R.; Wangtueai, S.; Jambrak, A.R.; Phongthai, S.; Jantanasakulwong, K.; Chaiyaso, T.; Phimolsiripol, Y. Optimization of ultrasonic-assisted extraction of polysaccharides from purple glutinous rice bran (Oryza sativa L.) and their antioxidant activities. Sci. Rep. 2020, 10, 10410. [Google Scholar] [CrossRef] [PubMed]
- Rattana, T.; Saengsanga, T. Antioxidant and Anti-food Pathogen Properties in Food from Mycelium Culture of Wild Mushroom. Thai. Sc. Technol. J. 2020, 28, 2250–2260. [Google Scholar]
- Chen, L.; Peng, X.; Lv, J.; Liao, S.; Ou, S.; Shen, Y. Purification and structural characterization of a novel water-soluble neutral polysaccharide from Cantharellus cibarius and its immunostimulating activity in RAW264.7 cells. Int. J. Polym. Sci. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Fang, D.; Zhao, R.; Gao, J.; Kimatu, B.M.; Hu, Q.; Chen, G.; Zhao, L. Effects of ultrasound-assisted extraction on antioxidant activity and bidirectional immunomodulatory activity of Flammulina velutipes polysaccharide. Int. J. Biol. Macromol. 2019, 140, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Zhou, X.; Wang, J.; Zhang, K.; Zhou, Y.; Chen, S.; Nie, S.; Xie, M. Cordyceps sinensis polysaccharide inhibits colon cancer cells growth by inducing apoptosis and autophagy flux blockage via mTOR signaling. Carbohydr. Polym. 2020, 237, 116113. [Google Scholar] [CrossRef]
- Shu, X.; Zhang, Y.; Jia, J.; Ren, X.; Wang, Y. Extraction, purification and properties of water-soluble polysaccharides from mushroom Lepista nuda. Int. J. Biol. Macromol. 2019, 128, 858–869. [Google Scholar] [CrossRef]
- Song, X.; Ren, Z.; Wang, X.; Jia, L.; Zhang, C. Antioxidant, anti-inflammatory and renoprotective effects of acidic-hydrolytic polysaccharides by spent mushroom compost (Lentinula edodes) on LPS-induced kidney injury. Int. J. Biol. Macromol. 2020, 151, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Nie, S.; Kan, L.; Chen, H.; Cui, S.W.; Phillips, A.O.; Phillips, G.O.; Xie, M. Comparison of structural features and antioxidant activity of polysaccharides from natural and cultured Cordyceps sinensis. Food Sci. Biotechnol. 2017, 26, 55–62. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, Y.; Shao, J.; Shu, X.; Jia, J.; Ren, X.; Guan, Y. Macrophage immunomodulatory activity of the polysaccharide isolated from Collybia radicata mushroom. Int. J. Biol. Macromol. 2018, 108, 300–306. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Y.; Zhao, X.; Tang, Q.; Dernedde, J.; Zhang, J.; Fan, H. Anti-inflammatory properties of GLPss58, a sulfated polysaccharide from Ganoderma lucidum. Int. J. Biol. Macromol. 2018, 107, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Mapoung, S.; Umsumarng, S.; Semmarath, W.; Arjsri, P.; Thippraphan, P.; Yodkeeree, S. Skin Wound-Healing Potential of Polysaccharides from Medicinal Mushroom Auricularia auricula-judae (Bull.). J. Fungi. 2021, 7, 247. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, I.; Sen, I.K.; Mondal, S.; Rout, D.; Bhanja, S.K.; Maity, G.N.; Maity, P. Bioactive polysaccharides from natural sources: A review on the antitumor and immunomodulating activities. Biocatal. Agric. Biotechnol. 2019, 22, 101425. [Google Scholar] [CrossRef]
- Maity, P.; Sen, I.K.; Maji, P.K.; Paloi, S.; Devi, K.S.P.; Acharya, K.; Maiti, T.K.; Islam, S.S. Structural, immunological, and antioxidant studies of β-glucan from edible mushroom Entoloma lividoalbum. Carbohydr. Polym. 2015, 123, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Jiang, L.; Qian, L.; Sun, W.; Tao, T.; Zan, X.; Yang, Y.; Wu, D.; Zhao, X. A macromolecular α-glucan from fruiting bodies of Volvariella volvacea activating RAW264.7 macrophages through MAPKs pathway. Carbohydr. Polym. 2020, 230, 115674. [Google Scholar] [CrossRef]
- Ruthes, A.C.; Carbonero, E.R.; Córdova, M.M.; Baggio, C.H.; Sassaki, G.L.; Gorin, P.A.; Santos, A.R.; Iacomini, M. Fucomannogalactan and glucan from mushroom Amanita muscaria: Structure and inflammatory pain inhibition. Carbohydr. Polym. 2013, 98, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Vetvicka, V.; Yvin, J.C. Effects of marine β−1,3 glucan on immune reactions. Int. Immunopharmacol. 2004, 4, 721–730. [Google Scholar] [CrossRef]
- Tabarsa, M.; Karnjanapratum, S.; Cho, M.; Kim, J.K.; You, S.G. Molecular characteristics and biological activities of anionic macromolecules from Codium fragile. Int. J. Biol. Macromol. 2013, 59, 1–12. [Google Scholar] [CrossRef]
- Cao, R.A.; Lee, Y.; You, S.G. Water soluble sulfated-fucans with immune-enhancing properties from Ecklonia cava. Int. J. Biol. Macromol. 2014, 67, 303–311. [Google Scholar] [CrossRef]
- Surayot, U.; Lee, J.H.; Park, W.; You, S.G. Structural characteristics of polysaccharides extracted from Cladophora glomerata Kützing affecting nitric oxide releasing capacity of RAW264.7 cells. Carbohydr. Diet. Fibre. 2016, 7, 26–31. [Google Scholar] [CrossRef]
- Surayot, U.; Lee, J.H.; Kanongnuch, C.; Peerapornpisal, Y.; Park, W.; You, S.G. Structural characterization of sulfated arabinans extracted from Cladophora glomerata Kützing and their macrophage activation. Biosci. Biotechnol. Biochem. 2016, 80, 972–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surayot, U.; Wang, J.; Lee, J.H.; Kanongnuch, C.; Peerapornpisal, Y.; You, S.G. Characterization and immunomodulatory activities of polysaccharides from Spirogyra neglecta (Hassall) Kützing. Biosci. Biotechnol. Biochem. 2015, 79, 1644–1653. [Google Scholar] [CrossRef]
- Yelithao, K.; Surayot, U.; Lee, C.; Palanisamy, S.; Prabhud, N.M.; Lee, J.H.; You, S.G. Studies on structural properties and immune-enhancing activities of glycomannans from Schizophyllum commune. Carbohydr. Polym. 2019, 218, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Surayot, U.; Yelithao, K.; Tabarsa, M.; Lee, D.H.; Palanisamy, S.; Marimuthu Prabhu, N.; Lee, J.; You, S.G. Structural characterization of a polysaccharide from Certaria islandica and assessment of immunostimulatory activity. Process Biochem. 2019, 83, 214–221. [Google Scholar] [CrossRef]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [Google Scholar] [CrossRef]
- Karaman, M.; Janjušević, L.; Jakovljević, D.; Šibul, F.; Pejin, B. Anti-hydroxyl radical activity, redox potential and anti-AChE activity of Amanita strobiliformis polysaccharide extract. Nat. Prod. Res. 2019, 33, 1522–1526. [Google Scholar] [CrossRef]
- Zhu, Y.; Ding, X.; Wang, M.; Hou, Y.; Hou, W.; Yue, C. Structure and antioxidant activity of a novel polysaccharide derived from Amanita caesarea. Mol. Med. Rep. 2016, 14, 3947–3954. [Google Scholar] [CrossRef] [Green Version]
- Percival, E.L.; McDowell, R.H. Chemistry and Enzymology of Marine Algal Polysaccharides; Academic Press: London, UK; New York, NY, USA, 1967. [Google Scholar]
- Liu, C.; Choi, M.W.; Li, X.; Cheung, P.C.K. Immunomodulatory effect of structurally-characterized mushroom sclerotial polysaccharides isolated from Polyporus rhinocerus on human monoctyes THP-1. J. Funct. Foods. 2018, 41, 90–99. [Google Scholar] [CrossRef]
- Tabarsa, M.; You, S.G.; Dabaghian, E.H.; Surayot, U. Water-soluble polysaccharides from Ulva intestinalis: Molecular properties, structural elucidation and immunomodulatory activities. J. Food Drug Anal. 2017, 26, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Li, C.; Surayot, U.; Yelithao, K.; Lee, S.M.; Park, W.J.; Tabarsa, M.; You, S.G. Molecular structures, chemical properties and biological activities of polysaccharide from Smilax glabra rhizome. Int. J. Biol. Macromol. 2018, 120, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Alavi, M.; Tabarsa, M.; You, S.G.; Gavlighi, H.A. Structural characteristics, molecular properties and immunostimulatory effects of sulfated polysaccharide from freshwater Myriophyllum spicatum L. Int. J. Biol. Macromol. 2020, 153, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Di, T.; Chen, G.; Sun, Y.; Ou, S.; Zeng, X.; Ye, H. Antioxidant and immunostimulating activities in vitro of sulfated polysaccharides isolated from Gracilaria rubra. J. Funct. Foods. 2017, 28, 64–75. [Google Scholar] [CrossRef]
- Leong, Y.K.; Yang, F.-C.; Chang, J.S. Extraction of polysaccharides from edible mushrooms: Emerging technologies and recent advances. Carbohydr. Polym. 2021, 251, 117006. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xu, X.; Yu, M.; Yang, Z.; Zheng, L. Characterisation and immunostimulatory activity of an α-(1→6)-d-glucan from the cultured Armillariella tabescens mycelia. Food Chem. 2008, 111, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Surayot, U.; Wang, J.; Seesuriyachan, P.; Kuntiya, A.; Tabarsa, M.; Lee, Y.; Kim, J.K.; Park, W.; You, S.G. Exopolysaccharides from lactic acid bacteria: Structural analysis, molecular weight effect on immunomodulation. Int. J. Biol. Macromol. 2014, 68, 233–240. [Google Scholar] [CrossRef]
- Bao, H.; Choi, W.S.; You, S.G. Effect of sulfated modification on the molecular characteristics and biological activities of polysaccharides from Hypsizigus marmoreus. Biosci. Biotechnol. Biochem. 2010, 74, 1408–1414. [Google Scholar] [CrossRef]
- Chaisuwan, W.; Jantanasakulwong, K.; Wangtueai, S.; Phimolsiripol, Y.; Chaiyaso, T.; Techapun, C.; Phongthai, S.; You, S.G.; Regenstein, J.M.; Seesuriyachan, P. Microbial exopolysaccharides for immune enhancement: Fermentation, modifications and bioactivities. Food Biosci. 2020, 35, 100564. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Hua, K.F.; Lin, C.C.; Lin, C.H.; Hsu, J.; Wong, C.H. Extract of Reishi polysaccharides induces cytokine expression via TLR4-modulated protein kinase signaling pathways. J. Immunol. 2004, 173, 5989–5999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Gupta, U.; Asthana, A.; Jain, N.K. Dextran conjugated dendritic nano constructs as potential vectors for anti-cancer agent. Biomaterials. 2009, 30, 3588–3596. [Google Scholar] [CrossRef]
- Zhang, A.; Deng, J.; Yu, S.; Zhang, F.; Linhardt, R.J.; Sun, P. Purification and structural elucidation of a water-soluble polysaccharide from the fruiting bodies of the Grifola frondosa. Int. J. Biol. Macromol. 2018, 115, 221–226. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharide | Bioactivity | Extraction Method | References |
|---|---|---|---|
| Cantharellus cibarius | Immunomodulatory activity | Boiling water extraction | [16] |
| Flammulina velutipes | Immunomodulatory, and antioxidant activity | Hot water extraction (80 °C), and ultrasound-assisted extraction | [17] |
| Cordyceps sinensis | Anticancer, and antioxidant activity | Hot water extraction (95–100 °C) | [18,21] |
| Lepista nuda | Antioxidant, antitumor, and antiviral activities | Water extraction (75–95 °C) | [19] |
| Lentinus edodes | Reno-protective activity | Water extraction (85 °C) | [20] |
| Collybia radicata | Immunomodulatory activity | Water extraction (84 °C) | [22] |
| Ganoderma lucidum | Anti-inflammatory | Boiling water extraction | [23] |
| Auricularia auricula-judae (Bull.) | Skin wound-healing | Water extraction (97 °C) | [24] |
| Polysaccharides | |||
|---|---|---|---|
| Component | MP | MPF1 | MPF2 |
| Yield (%) | 4.90 x | 57.3 y | 42.7 y |
| Total carbohydrate (% db) | 87.8 ± 2.64 b | 93.2 ± 0.12 a | 83.5 ± 0.50 c |
| Protein (% db) | 6.80 ± 0.17 b | 5.40 ± 0.20 c | 7.20 ± 0.09 a |
| Sulphate (% db) | 5.40 ± 0.20 b | 1.40 ± 0.03 c | 9.30 ± 0.10 a |
| Uronic acid (%) | - | - | - |
| Monosaccharide content (%) | |||
| Rhamnose | 1.40 ± 0.05 b | 2.70 ± 0.03 a | 0.80 ± 0.10 c |
| Arabinose | 1.80 ± 0.08 b | 4.70 ± 0.08 a | 0.30 ± 0.02 c |
| Xylose | 1.10 ± 0.10 b | 2.50 ± 0.20 a | - |
| Mannose | 3.30 ± 0.30 a | 1.40 ± 0.10 b | 0.30 ± 0.00 c |
| Glucose | 85.5 ± 0.50 b | 79.6 ± 1.35 c | 98.4 ± 0.46 a |
| Galactose | 6.80 ± 2.64 b | 9.10 ± 0.00 a | 0.20 ± 0.02 c |
| Molecular properties | |||
| Samples | MW × 103 (g/mol) | Rg (nm) | |
| MP | 479.4 ± 14.7 a | 148.2 ± 10.0 a | |
| MPF1 | 219.2 ± 14.8 b | 132.3 ± 6.4 b | |
| MPF2 | 104.0 ± 13.1 c | 173.1 ± 5.9 c | |
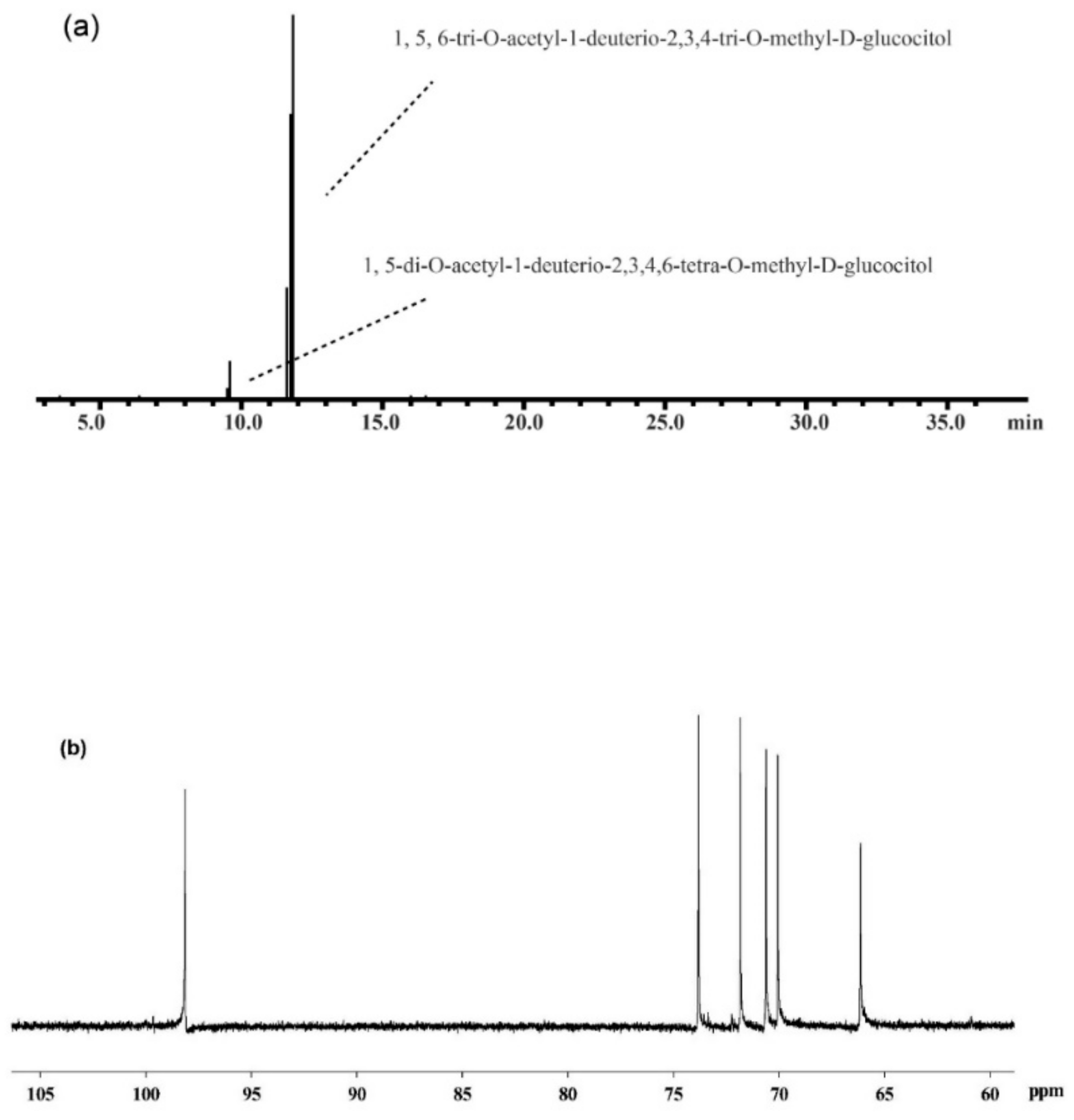
| Methylation Sugar | Retention Time (min) | Mass Fraction (m/z) | Glycosidic Linkage | Peak Area (%) |
|---|---|---|---|---|
| 2,3,4,6-Me4-Glc | 9.52 | 43, 59, 71, 87, 102, 118, 129, 145, 162, 174, 205 | Glc-(1→ | 1.7 |
| 2,3,4-Me3-Glc | 11.8 | 43, 59, 71, 87, 102, 118, 129, 162, 174, 189, 233 | →6)-Glc-(1→ | 98.3 |
| Residue | H-1/C-1 | H-2/C-2 | H-3/C-3 | H-4/C-4 | H-5/C-5 | H-6a, H-6b/C-6 |
|---|---|---|---|---|---|---|
| α-D-(1→6) Glcp | 5.19/97.2 | 3.75/72.3 | 3.98/70.7 | 3.72/70.0 | 4.14/74.1 | 3.95, 4.19/66.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surayot, U.; Wangtueai, S.; You, S.; Palanisamy, S.; Krusong, W.; Brennan, C.S.; Barba, F.J.; Phimolsiripol, Y.; Seesuriyachan, P. Extraction, Structural Characterisation, and Immunomodulatory Properties of Edible Amanita hemibapha subspecies javanica (Corner and Bas) Mucilage Polysaccharide as a Potential of Functional Food. J. Fungi 2021, 7, 683. https://doi.org/10.3390/jof7090683
Surayot U, Wangtueai S, You S, Palanisamy S, Krusong W, Brennan CS, Barba FJ, Phimolsiripol Y, Seesuriyachan P. Extraction, Structural Characterisation, and Immunomodulatory Properties of Edible Amanita hemibapha subspecies javanica (Corner and Bas) Mucilage Polysaccharide as a Potential of Functional Food. Journal of Fungi. 2021; 7(9):683. https://doi.org/10.3390/jof7090683
Chicago/Turabian StyleSurayot, Utoomporn, Sutee Wangtueai, SangGuan You, Subramanian Palanisamy, Warawut Krusong, Charles S. Brennan, Francisco J. Barba, Yuthana Phimolsiripol, and Phisit Seesuriyachan. 2021. "Extraction, Structural Characterisation, and Immunomodulatory Properties of Edible Amanita hemibapha subspecies javanica (Corner and Bas) Mucilage Polysaccharide as a Potential of Functional Food" Journal of Fungi 7, no. 9: 683. https://doi.org/10.3390/jof7090683
APA StyleSurayot, U., Wangtueai, S., You, S., Palanisamy, S., Krusong, W., Brennan, C. S., Barba, F. J., Phimolsiripol, Y., & Seesuriyachan, P. (2021). Extraction, Structural Characterisation, and Immunomodulatory Properties of Edible Amanita hemibapha subspecies javanica (Corner and Bas) Mucilage Polysaccharide as a Potential of Functional Food. Journal of Fungi, 7(9), 683. https://doi.org/10.3390/jof7090683