
Low Glucose Mediated Fluconazole Tolerance in Cryptococcus neoformans

 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Media and Growth Conditions
2.2. Analysis of Drug Resistance Gene and Super-Oxide dismutase (SOD) Expression
2.3. Nile Red Efflux Assay
2.4. Measurement of Mitochondrial Reactive Oxygen Species (mROS), Mitochondrial Membrane Potential (MMP) and Mitochondrial Mass
2.5. Fluorescence Microscopy and Transmission Electron Microscopy
2.6. Measurement of Cellular ATP Levels
2.7. Minimum Inhibitory Concentration
2.8. Analysis of Gene Copy Number
2.9. Statistics
3. Results
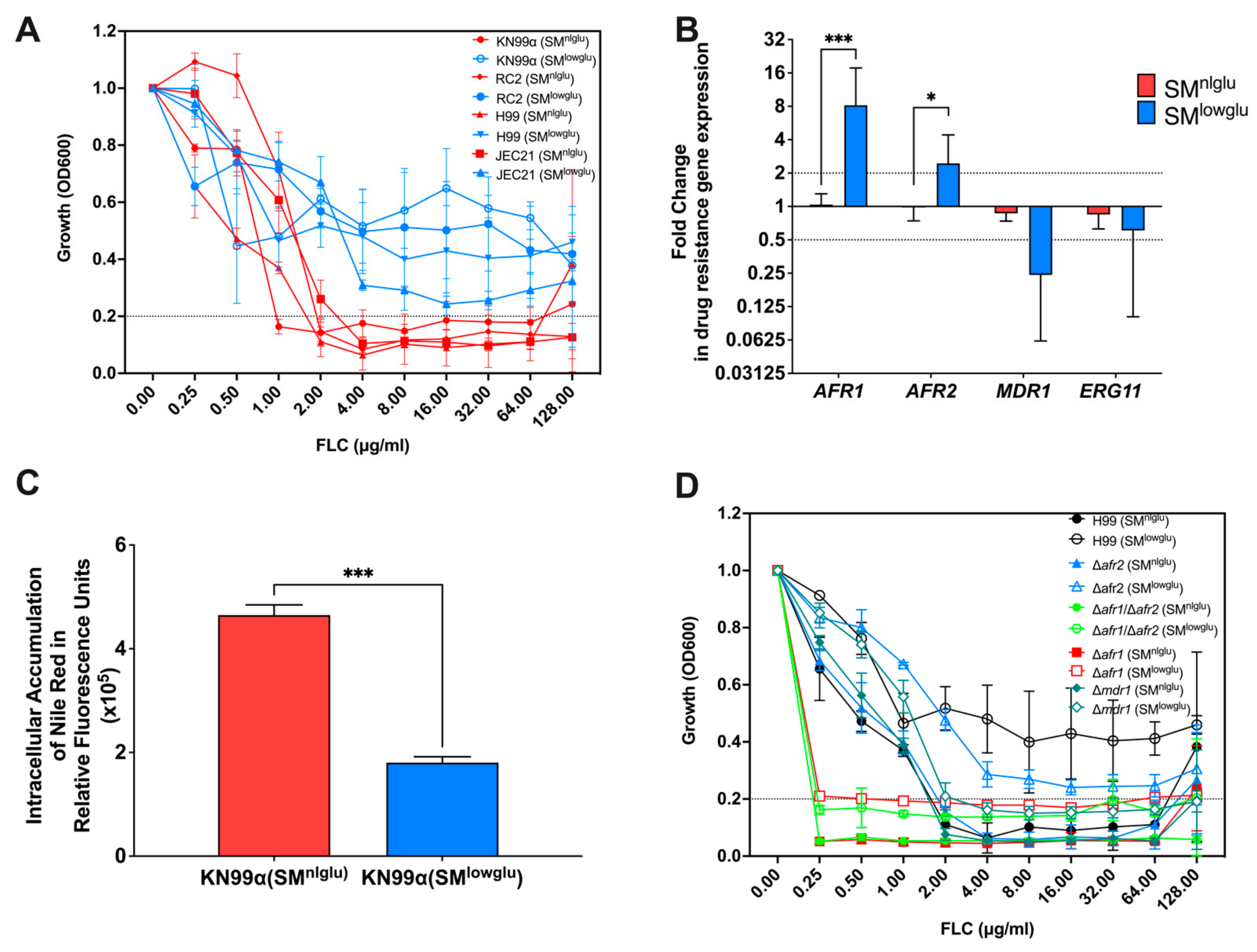
3.1. Low Glucose Increases FLC Resistance and Efflux Pump Activity
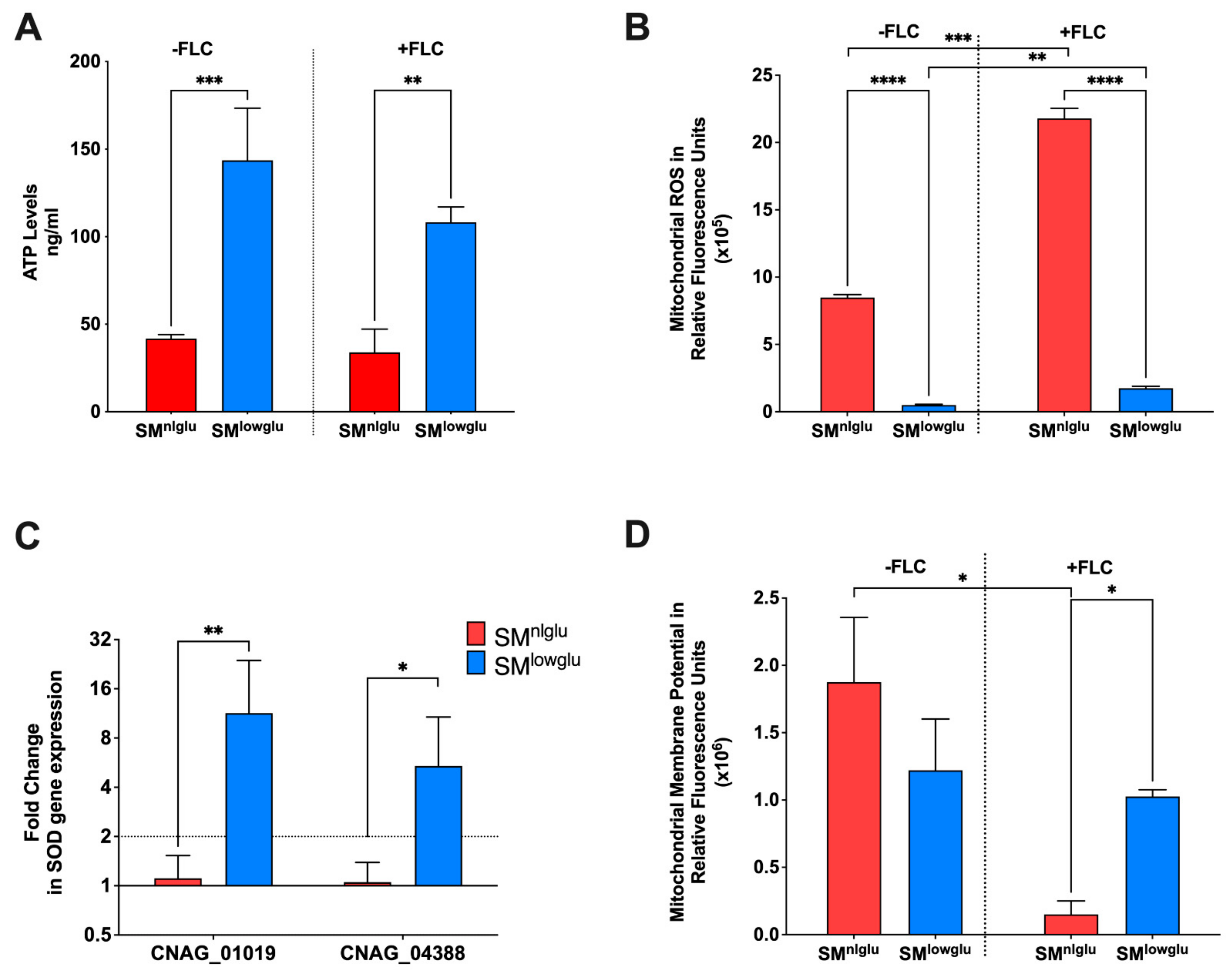
3.2. Effects of Low Glucose on Mitochondrial Function
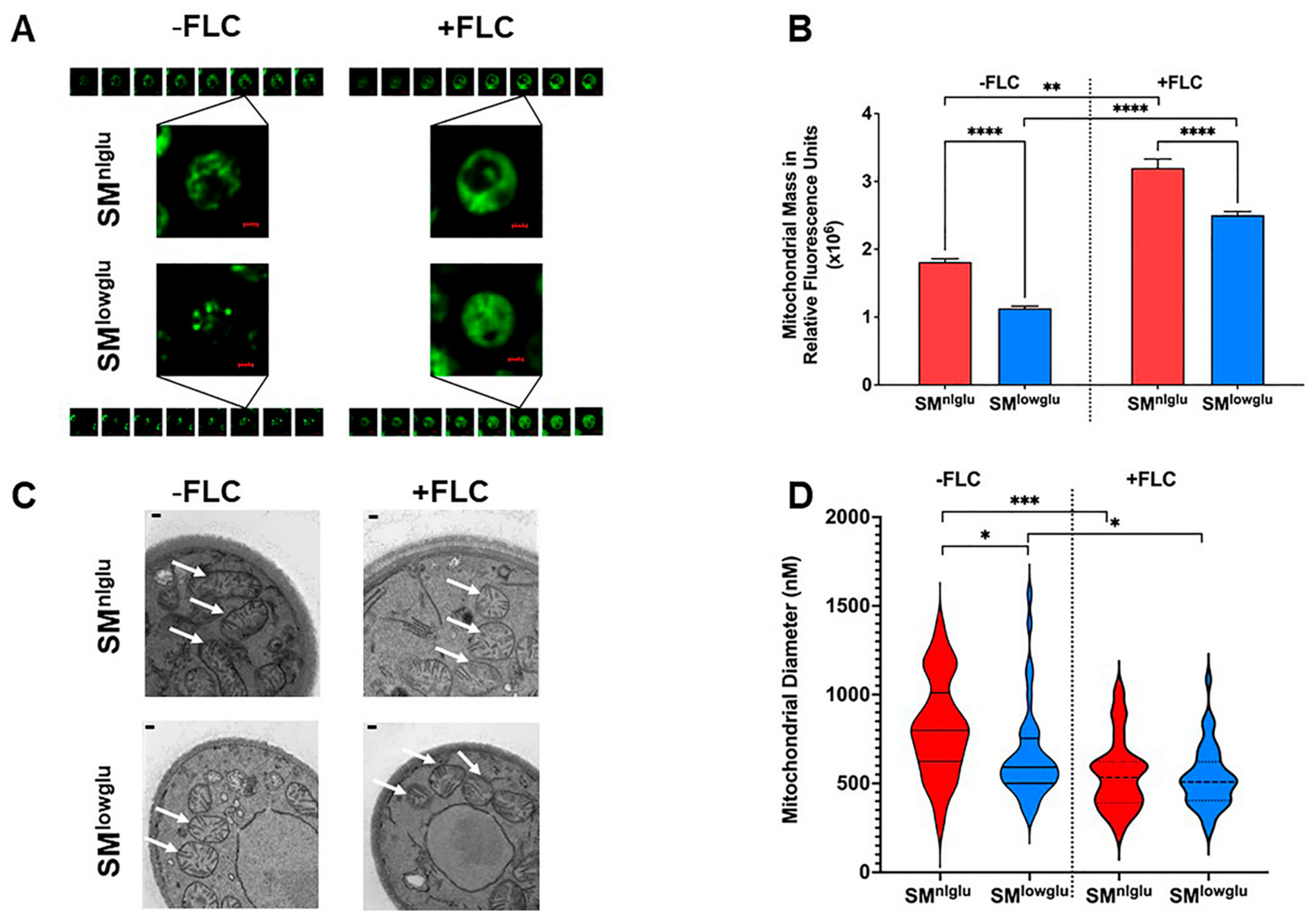
3.3. Low Glucose and FLC Exposure Alter Mitochondrial Morphology
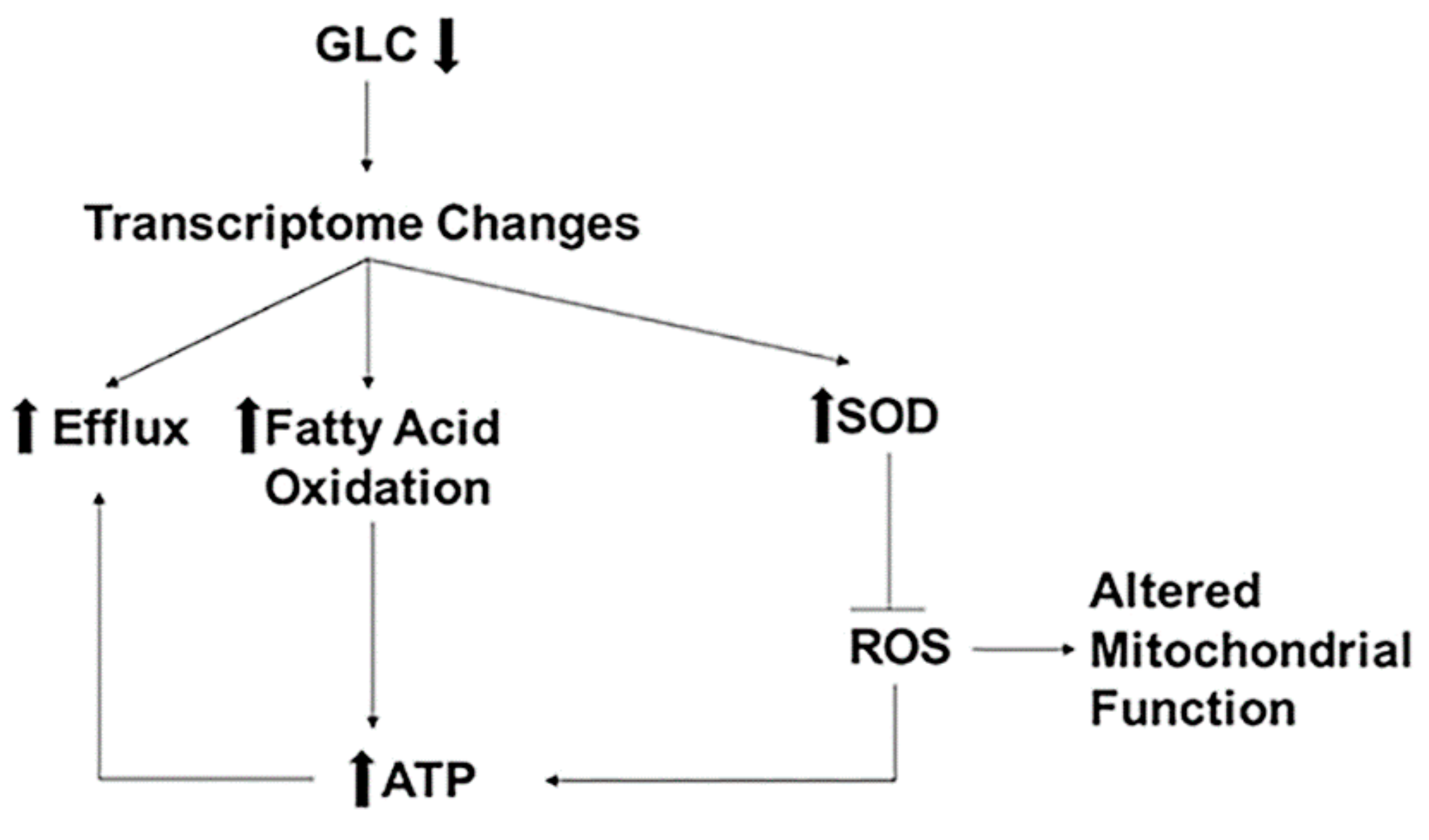
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouklas, T.; Masone, L.; Fries, B.C. Differences in sirtuin regulation in response to calorie restriction in Cryptococcus neoformans. J. Fungi 2018, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Park, B.J.; Wannemuehler, K.A.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. Aids 2009, 23, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Mada, P.; Nowack, B.; Cady, B.; Joel Chandranesan, A.S. Disseminated cryptococcosis in an immunocompetent patient. BMJ Case Rep. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Bollam, R.; Yassin, M.; Phan, T. Disseminated cryptococcosis in an immunocompetent patient. Respir Med. Case Rep. 2020, 30, 101034. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Desnos-Ollivier, M.; Dromer, F. Dynamics of Cryptococcus neoformans-macrophage interactions reveal that fungal background influences outcome during cryptococcal meningoencephalitis in humans. mBio 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Longley, N.; Muzoora, C.; Taseera, K.; Mwesigye, J.; Rwebembera, J.; Chakera, A.; Wall, E.; Andia, I.; Jaffar, S.; Harrison, T.S. Dose response effect of high-dose fluconazole for HIV-associated cryptococcal meningitis in southwestern Uganda. Clin. Infect. Dis 2008, 47, 1556–1561. [Google Scholar] [CrossRef]
- Gaskell, K.M.; Rothe, C.; Gnanadurai, R.; Goodson, P.; Jassi, C.; Heyderman, R.S.; Allain, T.J.; Harrison, T.S.; Lalloo, D.G.; Sloan, D.J.; et al. A prospective study of mortality from cryptococcal meningitis following treatment induction with 1200 mg oral fluconazole in Blantyre, Malawi. PLoS ONE 2014, 9, e110285. [Google Scholar] [CrossRef] [PubMed]
- Rothe, C.; Sloan, D.J.; Goodson, P.; Chikafa, J.; Mukaka, M.; Denis, B.; Harrison, T.; van Oosterhout, J.J.; Heyderman, R.S.; Lalloo, D.G.; et al. A prospective longitudinal study of the clinical outcomes from cryptococcal meningitis following treatment induction with 800 mg oral fluconazole in Blantyre, Malawi. PLoS ONE 2013, 8, e67311. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or deletion of ergosterol biosynthesis genes alters doubling time, response to stress agents, and drug susceptibility in Saccharomyces cerevisiae. mBio 2018, 9. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Boyken, L.; Hollis, R.J.; Kroeger, J.; Messer, S.A.; Tendolkar, S.; Diekema, D.J. Wild-type MIC distributions and epidemiological cutoff values for posaconazole and voriconazole and Candida spp. as determined by 24-hour CLSI broth microdilution. J. Clin. Microbiol. 2011, 49, 630–637. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Fries, B.C. Enhanced efflux pump activity in old Candida glabrata cells. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Holowka, T.; Orner, E.P.; Fries, B.C. Gene duplication associated with increased fluconazole tolerance in Candida auris cells of advanced generational Age. Sci. Rep. 2019, 9, 5052. [Google Scholar] [CrossRef]
- Chang, M.; Sionov, E.; Khanal Lamichhane, A.; Kwon-Chung, K.J.; Chang, Y.C. Roles of three Cryptococcus neoformans and Cryptococcus gattii efflux pump-coding genes in response to drug treatment. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sobel, J.D.; White, T.C. A Combination Fluorescence Assay Demonstrates increased efflux pump activity as a resistance mechanism in azole-resistant vaginal Candida albicans isolates. Antimicrob. Agents Chemother. 2016, 60, 5858–5866. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and mechanisms of antifungal resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Basso, L.R., Jr.; Gast, C.E.; Bruzual, I.; Wong, B. Identification and properties of plasma membrane azole efflux pumps from the pathogenic fungi Cryptococcus gattii and Cryptococcus neoformans. J. Antimicrob. Chemother. 2015, 70, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Sanguinetti, M.; De Bernardis, F.; Torelli, R.; Posteraro, B.; Vandeputte, P.; Sanglard, D. Loss of mitochondrial functions associated with azole resistance in Candida glabrata results in enhanced virulence in mice. Antimicrob. Agents Chemother. 2011, 55, 1852–1860. [Google Scholar] [CrossRef]
- Lu, Z.; Jia, X.; Chen, Y.; Han, X.; Chen, F.; Tian, S.; Su, X.; Li, Z.; Zhao, J.; Zhang, X.; et al. Identification and characterization of key charged residues in the cofilin protein involved in azole susceptibility, apoptosis, and virulence of Aspergillus fumigatus. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Sionov, E.; Chang, Y.C.; Garraffo, H.M.; Kwon-Chung, K.J. Heteroresistance to fluconazole in Cryptococcus neoformans is intrinsic and associated with virulence. Antimicrob. Agents Chemother. 2009, 53, 2804–2815. [Google Scholar] [CrossRef]
- Stone, N.R.; Rhodes, J.; Fisher, M.C.; Mfinanga, S.; Kivuyo, S.; Rugemalila, J.; Segal, E.S.; Needleman, L.; Molloy, S.F.; Kwon-Chung, J.; et al. Dynamic ploidy changes drive fluconazole resistance in human cryptococcal meningitis. J. Clin. Investig. 2019, 129, 999–1014. [Google Scholar] [CrossRef]
- Williams, V.; Del Poeta, M. Role of glucose in the expression of Cryptococcus neoformans antiphagocytic protein 1, App1. Eukaryot. Cell 2011, 10, 293–301. [Google Scholar] [CrossRef]
- Hu, G.; Cheng, P.Y.; Sham, A.; Perfect, J.R.; Kronstad, J.W. Metabolic adaptation in Cryptococcus neoformans during early murine pulmonary infection. Mol. Microbiol. 2008, 69, 1456–1475. [Google Scholar] [CrossRef]
- Lee, H.; Khanal Lamichhane, A.; Garraffo, H.M.; Kwon-Chung, K.J.; Chang, Y.C. Involvement of PDK1, PKC and TOR signalling pathways in basal fluconazole tolerance in Cryptococcus neoformans. Mol. Microbiol. 2012, 84, 130–146. [Google Scholar] [CrossRef]
- Yu, C.H.; Chen, Y.; Desjardins, C.A.; Tenor, J.L.; Toffaletti, D.L.; Giamberardino, C.; Litvintseva, A.; Perfect, J.R.; Cuomo, C.A. Landscape of gene expression variation of natural isolates of Cryptococcus neoformans in response to biologically relevant stresses. Microb. Genom. 2020, 6. [Google Scholar] [CrossRef]
- Park, Y.D.; Jarvis, J.N.; Hu, G.; Davis, S.E.; Qiu, J.; Zhang, N.; Hollingsworth, C.; Loyse, A.; Gardina, P.J.; Valyi-Nagy, T.; et al. Transcriptional profiling of patient isolates identifies a novel TOR/starvation regulatory pathway in cryptococcal virulence. mBio 2018, 9. [Google Scholar] [CrossRef]
- Wierman, M.B.; Maqani, N.; Strickler, E.; Li, M.; Smith, J.S. caloric restriction extends yeast chronological life span by optimizing the Snf1 (AMPK) signaling pathway. Mol. Cell. Biol. 2017, 37. [Google Scholar] [CrossRef]
- Bouklas, T.; Pechuan, X.; Goldman, D.L.; Edelman, B.; Bergman, A.; Fries, B.C. Old Cryptococcus neoformans cells contribute to virulence in chronic cryptococcosis. mBio 2013, 4. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bouklas, T.; Jain, N.; Fries, B.C. Modulation of Replicative Lifespan in Cryptococcus neoformans: Implications for Virulence. Front. Microbiol. 2017, 8, 98. [Google Scholar] [CrossRef]
- Munshi, M.A.; Gardin, J.M.; Singh, A.; Luberto, C.; Rieger, R.; Bouklas, T.; Fries, B.C.; Del Poeta, M. The role of ceramide synthases in the pathogenicity of Cryptococcus neoformans. Cell Rep. 2018, 22, 1392–1400. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Dounce, A.L.; Rothstein, A. A detailed procedure for the preparation of highly purified adenosine triphosphate. J. Biol. Chem. 1948, 174, 361–370. [Google Scholar] [CrossRef]
- Bertout, S.; Dunyach, C.; Drakulovski, P.; Reynes, J.; Mallie, M. Comparison of the Sensititre YeastOne(R) dilution method with the Clinical Laboratory Standards Institute (CLSI) M27-A3 microbroth dilution reference method for determining MIC of eight antifungal agents on 102 yeast strains. Pathol. Biol. 2011, 59, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.; Semchyshyn, H.; Mandryk, S.; Lushchak, O. Possible role of superoxide dismutases in the yeast Saccharomyces cerevisiae under respiratory conditions. Arch. Biochem. Biophys. 2005, 441, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Vyssokikh, M.Y.; Holtze, S.; Averina, O.A.; Lyamzaev, K.G.; Panteleeva, A.A.; Marey, M.V.; Zinovkin, R.A.; Severin, F.F.; Skulachev, M.V.; Fasel, N.; et al. Mild depolarization of the inner mitochondrial membrane is a crucial component of an anti-aging program. Proc. Natl. Acad. Sci. USA 2020, 117, 6491–6501. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Deng, X.; Zhou, W.; Tan, E.K. Flow cytometry-based assessment of mitophagy using mitotracker. Front. Cell. Neurosci. 2016, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.L.; Doering, T.L. Maintenance of mitochondrial morphology in Cryptococcus neoformans is critical for stress resistance and virulence. mBio 2018, 9. [Google Scholar] [CrossRef]
- Monteiro, L.D.; Davanzo, G.G.; de Aguiar, C.F.; Moraes-Vieira, P.M.M. Using flow cytometry for mitochondrial assays. Methodsx 2020, 7. [Google Scholar] [CrossRef]
- Brown, S.M.; Campbell, L.T.; Lodge, J.K. Cryptococcus neoformans, a fungus under stress. Curr. Opin. Microbiol. 2007, 10, 320–325. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Kojima, K.; Cox, G.M.; Heitman, J. A unique fungal two-component system regulates stress responses, drug sensitivity, sexual development, and virulence of Cryptococcus neoformans. Mol. Biol. Cell 2006, 17, 3122–3135. [Google Scholar] [CrossRef]
- Bloom, A.L.; Solomons, J.T.; Havel, V.E.; Panepinto, J.C. Uncoupling of mRNA synthesis and degradation impairs adaptation to host temperature in Cryptococcus neoformans. Mol. Microbiol. 2013, 89, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Upadhya, R.; Campbell, L.T.; Donlin, M.J.; Aurora, R.; Lodge, J.K. Global transcriptome profile of Cryptococcus neoformans during exposure to hydrogen peroxide induced oxidative stress. PLoS ONE 2013, 8, e55110. [Google Scholar] [CrossRef]
- Chabrier-Rosello, Y.; Gerik, K.J.; Koselny, K.; DiDone, L.; Lodge, J.K.; Krysan, D.J. Cryptococcus neoformans phosphoinositide-dependent kinase 1 (PDK1) ortholog is required for stress tolerance and survival in murine phagocytes. Eukaryot. Cell 2013, 12, 12–22. [Google Scholar] [CrossRef]
- Keniya, M.V.; Fleischer, E.; Klinger, A.; Cannon, R.D.; Monk, B.C. Inhibitors of the Candida albicans major facilitator superfamily transporter Mdr1p responsible for fluconazole resistance. PLoS ONE 2015, 10, e0126350. [Google Scholar] [CrossRef]
- Ivnitski-Steele, I.; Holmes, A.R.; Lamping, E.; Monk, B.C.; Cannon, R.D.; Sklar, L.A. Identification of Nile red as a fluorescent substrate of the Candida albicans ATP-binding cassette transporters Cdr1p and Cdr2p and the major facilitator superfamily transporter Mdr1p. Anal. Biochem 2009, 394, 87–91. [Google Scholar] [CrossRef]
- Whaley, S.G.; Zhang, Q.; Caudle, K.E.; Rogers, P.D. Relative contribution of the ABC transporters Cdr1, Pdh1, and Snq2 to azole resistance in Candida glabrata. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Bencova, A.; Goffa, E.; Morvova, M.; Valachovic, M.; Griac, P.; Toth Hervay, N.; Gbelska, Y. The Absence of PDR16 Gene restricts the overexpression of CaSNQ2 gene in the presence of fluconazole in Candida albicans. Mycopathologia 2020, 185, 455–465. [Google Scholar] [CrossRef]
- Thomas, E.; Roman, E.; Claypool, S.; Manzoor, N.; Pla, J.; Panwar, S.L. Mitochondria influence CDR1 efflux pump activity, Hog1-mediated oxidative stress pathway, iron homeostasis, and ergosterol levels in Candida albicans. Antimicrob. Agents Chemother. 2013, 57, 5580–5599. [Google Scholar] [CrossRef]
- Sionov, E.; Chang, Y.C.; Kwon-Chung, K.J. Azole heteroresistance in Cryptococcus neoformans: Emergence of resistant clones with chromosomal disomy in the mouse brain during fluconazole treatment. Antimicrob. Agents Chemother. 2013, 57, 5127–5130. [Google Scholar] [CrossRef] [PubMed]
- Sionov, E.; Lee, H.; Chang, Y.C.; Kwon-Chung, K.J. Cryptococcus neoformans overcomes stress of azole drugs by formation of disomy in specific multiple chromosomes. PLoS Pathog. 2010, 6, e1000848. [Google Scholar] [CrossRef]
- Jung, K.W.; Yang, D.H.; Maeng, S.; Lee, K.T.; So, Y.S.; Hong, J.; Choi, J.; Byun, H.J.; Kim, H.; Bang, S.; et al. Systematic functional profiling of transcription factor networks in Cryptococcus neoformans. Nat. Commun. 2015, 6, 6757. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.M.; Hong, S.J.; van Deursen, J.M.; Kim, S.; Kim, K.H.; Lee, C.K. Caloric restriction and rapamycin differentially alter energy metabolism in yeast. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 73, 29–38. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Bille, J. Role of ATP-binding-cassette transporter genes in high-frequency acquisition of resistance to azole antifungals in Candida glabrata. Antimicrob. Agents Chemother. 2001, 45, 1174–1183. [Google Scholar] [CrossRef]
- Zhang, X.; Moye-Rowley, W.S. Saccharomyces cerevisiae multidrug resistance gene expression inversely correlates with the status of the F(0) component of the mitochondrial ATPase. J. Biol. Chem. 2001, 276, 47844–47852. [Google Scholar] [CrossRef] [PubMed]
- Shekhova, E.; Kniemeyer, O.; Brakhage, A.A. Induction of mitochondrial reactive oxygen species production by itraconazole, terbinafine, and amphotericin b as a mode of action against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Kim, M.J.; Hwang, J.W.; Yun, C.K.; Lee, Y.; Choi, Y.S. Delivery of exogenous mitochondria via centrifugation enhances cellular metabolic function. Sci. Rep. 2018, 8, 3330. [Google Scholar] [CrossRef]
- Trevijano-Contador, N.; Rossi, S.A.; Alves, E.; Landin-Ferreiroa, S.; Zaragoza, O. Capsule enlargement in Cryptococcus neoformans is dependent on mitochondrial activity. Front. Microbiol. 2017, 8, 1423. [Google Scholar] [CrossRef] [PubMed]
- Westermann, B. Bioenergetic role of mitochondrial fusion and fission. Biochim. Biophys. Acta 2012, 1817, 1833–1838. [Google Scholar] [CrossRef]
- Horn, A.; Raavicharla, S.; Shah, S.; Cox, D.; Jaiswal, J.K. Mitochondrial fragmentation enables localized signaling required for cell repair. J. Cell Biol. 2020, 219. [Google Scholar] [CrossRef]
- Laporte, D.; Gouleme, L.; Jimenez, L.; Khemiri, I.; Sagot, I. Mitochondria reorganization upon proliferation arrest predicts individual yeast cell fate. eLife 2018, 7. [Google Scholar] [CrossRef]
- Kamogashira, T.; Hayashi, K.; Fujimoto, C.; Iwasaki, S.; Yamasoba, T. Functionally and morphologically damaged mitochondria observed in auditory cells under senescence-inducing stress. NPJ Aging Mech. Dis. 2017, 3, 2. [Google Scholar] [CrossRef]
- Eustaquio, T.; Wang, C.; Dugard, C.K.; George, N.I.; Liu, F.; Slikker, W., Jr.; Paule, M.G.; Howard, P.C.; Paredes, A.M. Electron microscopy techniques employed to explore mitochondrial defects in the developing rat brain following ketamine treatment. Exp. Cell Res. 2018, 373, 164–170. [Google Scholar] [CrossRef]
- Sanchez-Alvarez, R.; De Francesco, E.M.; Fiorillo, M.; Sotgia, F.; Lisanti, M.P. Mitochondrial fission factor (MFF) inhibits mitochondrial metabolism and reduces breast cancer stem cell (CSC) Activity. Front. Oncol. 2020, 10, 1776. [Google Scholar] [CrossRef]
- Wei, Y.H.; Lee, C.F.; Lee, H.C.; Ma, Y.S.; Wang, C.W.; Lu, C.Y.; Pang, C.Y. Increases of mitochondrial mass and mitochondrial genome in association with enhanced oxidative stress in human cells harboring 4,977 BP-deleted mitochondrial DNA. Ann. N. Y. Acad. Sci. 2001, 928, 97–112. [Google Scholar] [CrossRef]
- Lee, C.F.; Liu, C.Y.; Hsieh, R.H.; Wei, Y.H. Oxidative stress-induced depolymerization of microtubules and alteration of mitochondrial mass in human cells. Ann. N. Y. Acad. Sci. 2005, 1042, 246–254. [Google Scholar] [CrossRef]
- Apostolova, N.; Gomez-Sucerquia, L.J.; Moran, A.; Alvarez, A.; Blas-Garcia, A.; Esplugues, J.V. Enhanced oxidative stress and increased mitochondrial mass during efavirenz-induced apoptosis in human hepatic cells. Br. J. Pharmacol. 2010, 160, 2069–2084. [Google Scholar] [CrossRef]
- Kaur, R.; Castano, I.; Cormack, B.P. Functional genomic analysis of fluconazole susceptibility in the pathogenic yeast Candida glabrata: Roles of calcium signaling and mitochondria. Antimicrob. Agents Chemother. 2004, 48, 1600–1613. [Google Scholar] [CrossRef]
- Shingu-Vazquez, M.; Traven, A. Mitochondria and fungal pathogenesis: Drug tolerance, virulence, and potential for antifungal therapy. Eukaryot. Cell 2011, 10, 1376–1383. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | FLC (µg/mL) | ||
|---|---|---|---|
| 2% GLC | 0.5% GLC | 0.05% GLC | |
| KN99α | 1 | 1 | >64 |
| H99 | 2 | 2 | >64 |
| RC2 | 2 | 2 | >64 |
| JEC21 | 2 | 2 | >64 |
| H99Δarf1 | 0.25 | NA | 0.5 |
| H99Δarf2 | 2 | NA | >64 |
| H99Δmdr1 | 2 | NA | 2 |
| H99Δarf1/Δarf2 | 0.25 | NA | 0.25 |
| I55 | 0.5 | NA | 2 |
| I114 | 2 | NA | 8 |
| J9 | 1 | NA | 2 |
| J22 | 1 | NA | 4 |
| Gene ID | Fold Change | p-Value | Description | Significant |
|---|---|---|---|---|
| CNAG_03080 | 0.70391347 | 1.77 × 10−2 | fatty acid elongase, putative | YES |
| CNAG_01191 | 1.47264531 | 1.81 × 10−2 | long-chain fatty acid transporter, putative | YES |
| CNAG_02099 | 1.68451118 | 7.76 × 10−3 | fatty-acid synthase complex protein, putative | YES |
| CNAG_03019 | 1.92252186 | 2.61 × 10−4 | long-chain-fatty-acid-CoA ligase, putative | YES |
| CNAG_00644 | 1.93712896 | 3.63 × 10−6 | fatty acid desaturase, putative | YES |
| CNAG_04485 | 2.10846051 | 6.80 × 10−5 | long-chain-fatty-acid-CoA-ligase, putative | YES |
| CNAG_00984 | 4.48453997 | 2.07 × 10−5 | fatty acid beta-oxidation-related protein, putative | YES |
| CNAG_03666 | 6.42630743 | 2.31 × 10−7 | acyl-CoA dehydrogenase, long-chain specific precursor, putative | YES |
| CNAG_04688 | 2.67314853 | 1.87 × 10−4 | acyl-CoA dehydrogenase, putative | YES |
| Gene ID | Fold Change | p-Value | Description | Significant |
|---|---|---|---|---|
| CNAG_00896 | 10.4893255 | 3.49 × 10−25 | transcription factor, putative | YES |
| CNAG_07724 | 3.42118669 | 4.18 × 10−5 | ligand-regulated transcription factor, putative | YES |
| CNAG_04263 | 2.65694836 | 4.47 × 10−3 | transcriptional activator, putative | YES |
| CNAG_00156 | 2.58714788 | 1.74 × 102 | conserved hypothetical protein | YES |
| CNAG_02696 | 2.87529851 | 3.56 × 10−4 | exocyst protein, putative | YES |
| CNAG_01431 | 3.67805054 | 2.88 × 10−6 | LIM-homeobox protein, putative | YES |
| Mitochondrial Characterization | SMnlglu | SMlowglu | ||
|---|---|---|---|---|
| (−) FLC | (+) FLC | (−) FLC | (+) FLC | |
| ATP levels | ↓ | ↓ | ↑↑ | ↑ |
| mROS | ↑ | ↑↑ | ↓↓ | ↓ |
| SOD | ↓ | nt | ↑↑ | nt |
| MMP | ↑↑ | ↑ | ↓ | ↑ |
| Mitochondrial morphology | tubular | dispersed | fragmented | dispersed |
| Mitochondrial mass | ↑ | ↑↑ | ↓↓ | ↓ |
| Mitochondrial diameter | ↑ | ↓ | ↓ | ↓↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharya, S.; Oliveira, N.K.; Savitt, A.G.; Silva, V.K.A.; Krausert, R.B.; Ghebrehiwet, B.; Fries, B.C. Low Glucose Mediated Fluconazole Tolerance in Cryptococcus neoformans. J. Fungi 2021, 7, 489. https://doi.org/10.3390/jof7060489
Bhattacharya S, Oliveira NK, Savitt AG, Silva VKA, Krausert RB, Ghebrehiwet B, Fries BC. Low Glucose Mediated Fluconazole Tolerance in Cryptococcus neoformans. Journal of Fungi. 2021; 7(6):489. https://doi.org/10.3390/jof7060489
Chicago/Turabian StyleBhattacharya, Somanon, Natalia Kronbauer Oliveira, Anne G. Savitt, Vanessa K. A. Silva, Rachel B. Krausert, Berhane Ghebrehiwet, and Bettina C. Fries. 2021. "Low Glucose Mediated Fluconazole Tolerance in Cryptococcus neoformans" Journal of Fungi 7, no. 6: 489. https://doi.org/10.3390/jof7060489
APA StyleBhattacharya, S., Oliveira, N. K., Savitt, A. G., Silva, V. K. A., Krausert, R. B., Ghebrehiwet, B., & Fries, B. C. (2021). Low Glucose Mediated Fluconazole Tolerance in Cryptococcus neoformans. Journal of Fungi, 7(6), 489. https://doi.org/10.3390/jof7060489