
Enhanced Outcrossing, Directional Selection and Transgressive Segregation Drive Evolution of Novel Phenotypes in Hybrid Swarms of the Dutch Elm Disease Pathogen Ophiostoma novo-ulmi

Abstract
:
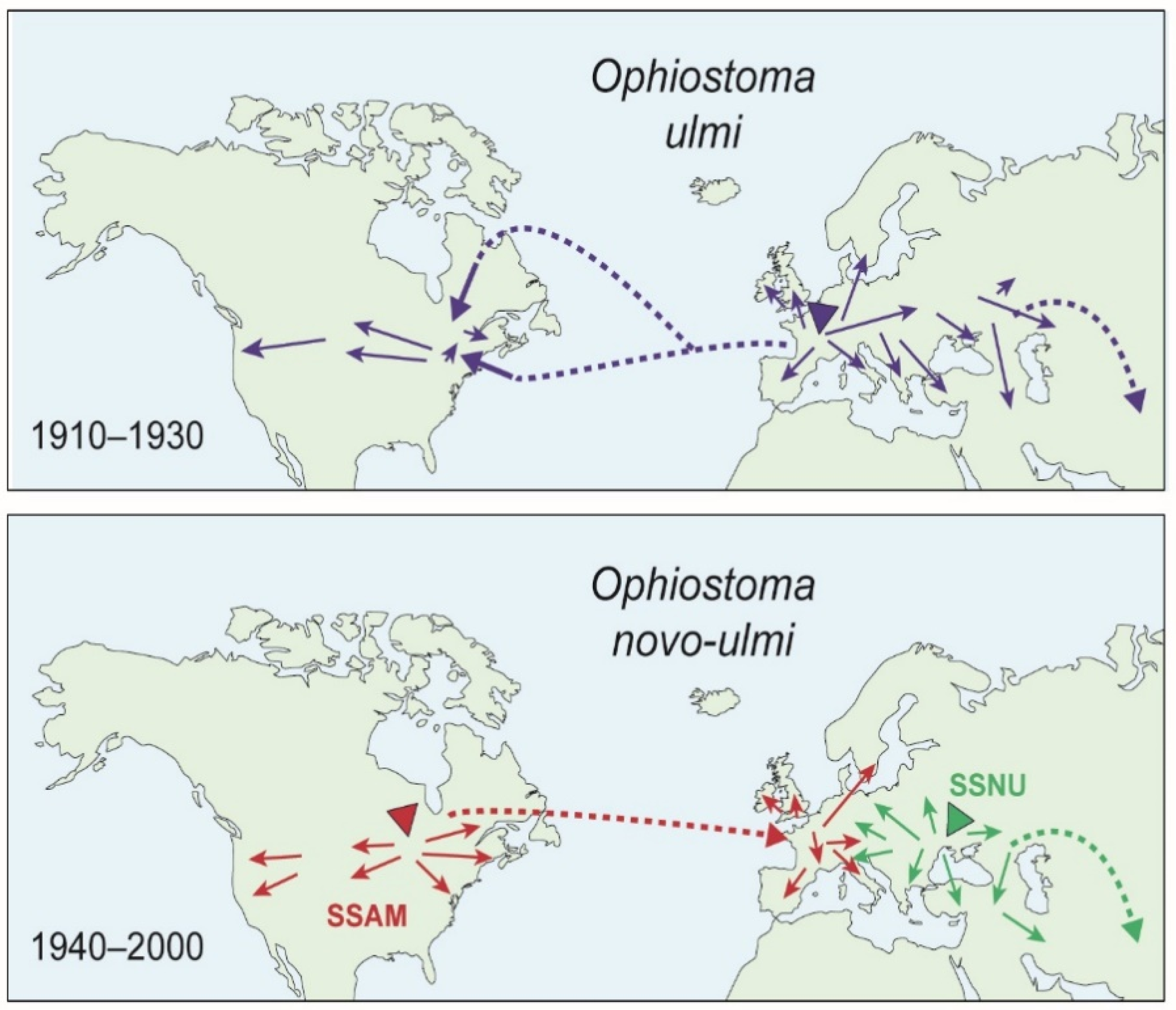
1. Introduction
2. Materials and Methods
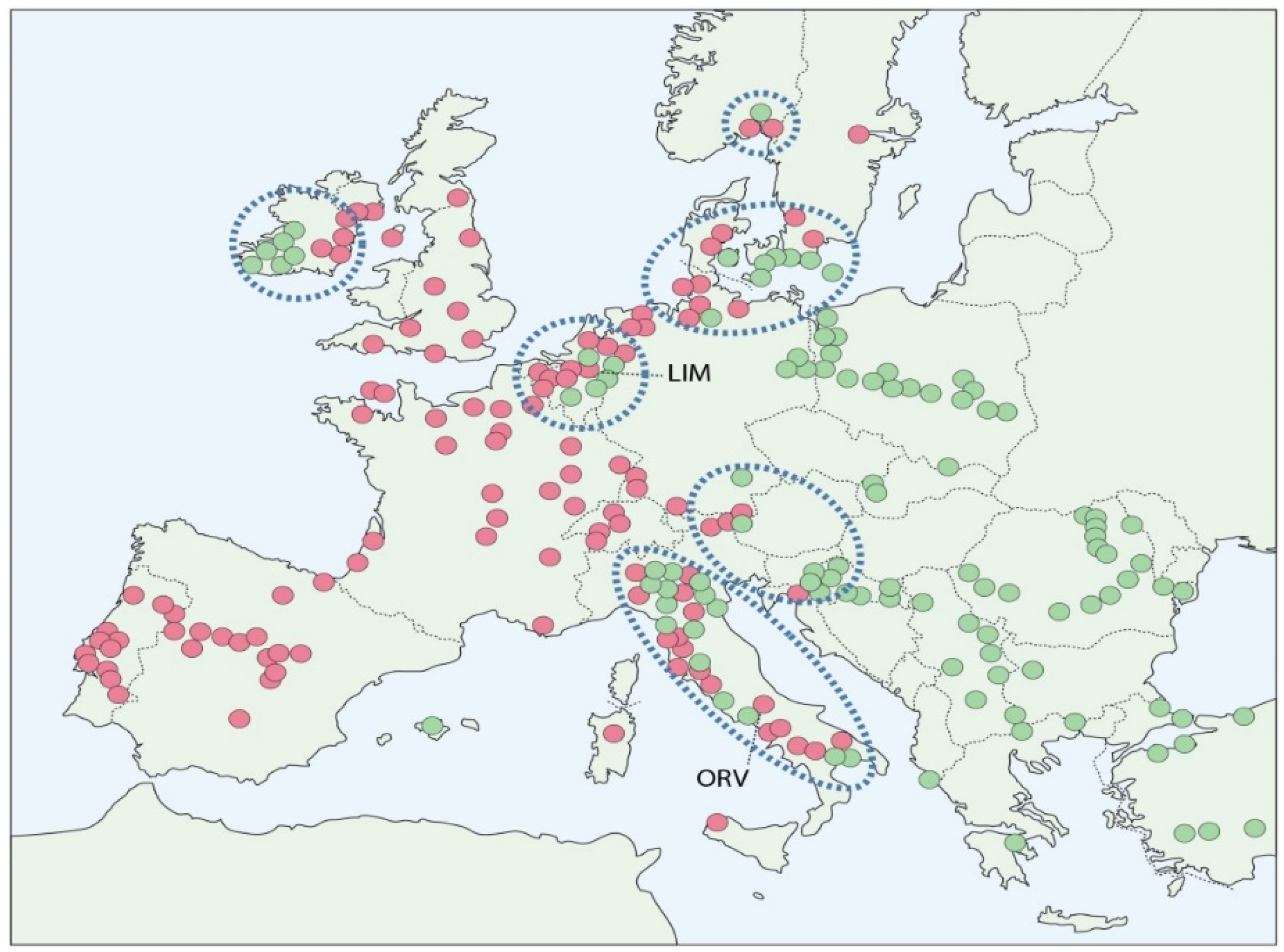
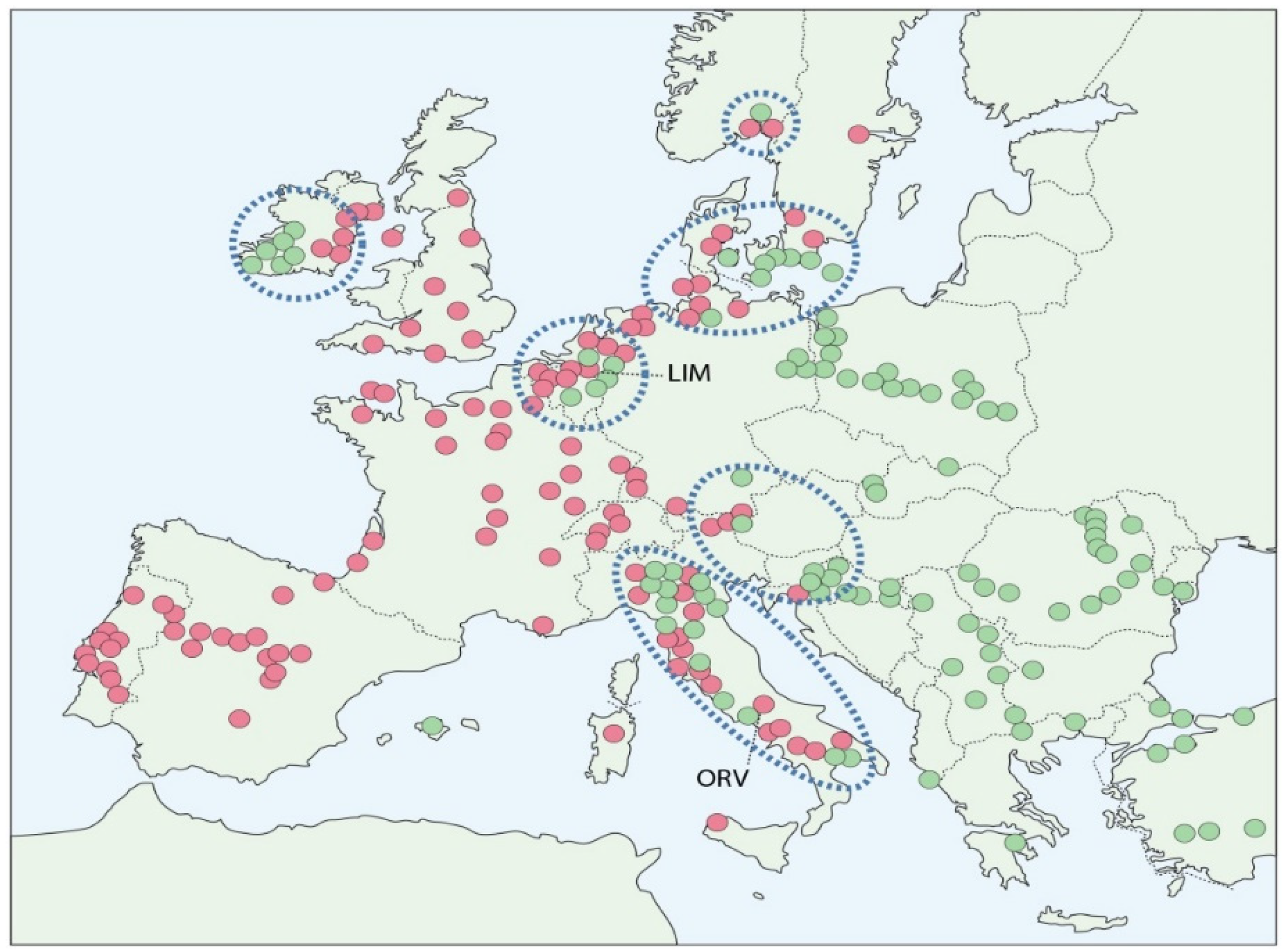
2.1. Comparative Site Conditions
2.2. Isolation Methods and Isolates Used in Tests
2.3. Media and Culture Storage
2.4. Sexual Compatibility Types
2.5. Vegetative Compatibility Type Tests
2.6. Growth Rate, Colony Morphology and Pathogenicity Tests
2.7. Production of Perithecia for Morphological Comparisons
2.8. DNA Extraction and RFLP Analysis of Cu and Col-1 Loci
3. Results
3.1. Increase in MAT-1 Frequency at the Limburg and Orvieto Sites
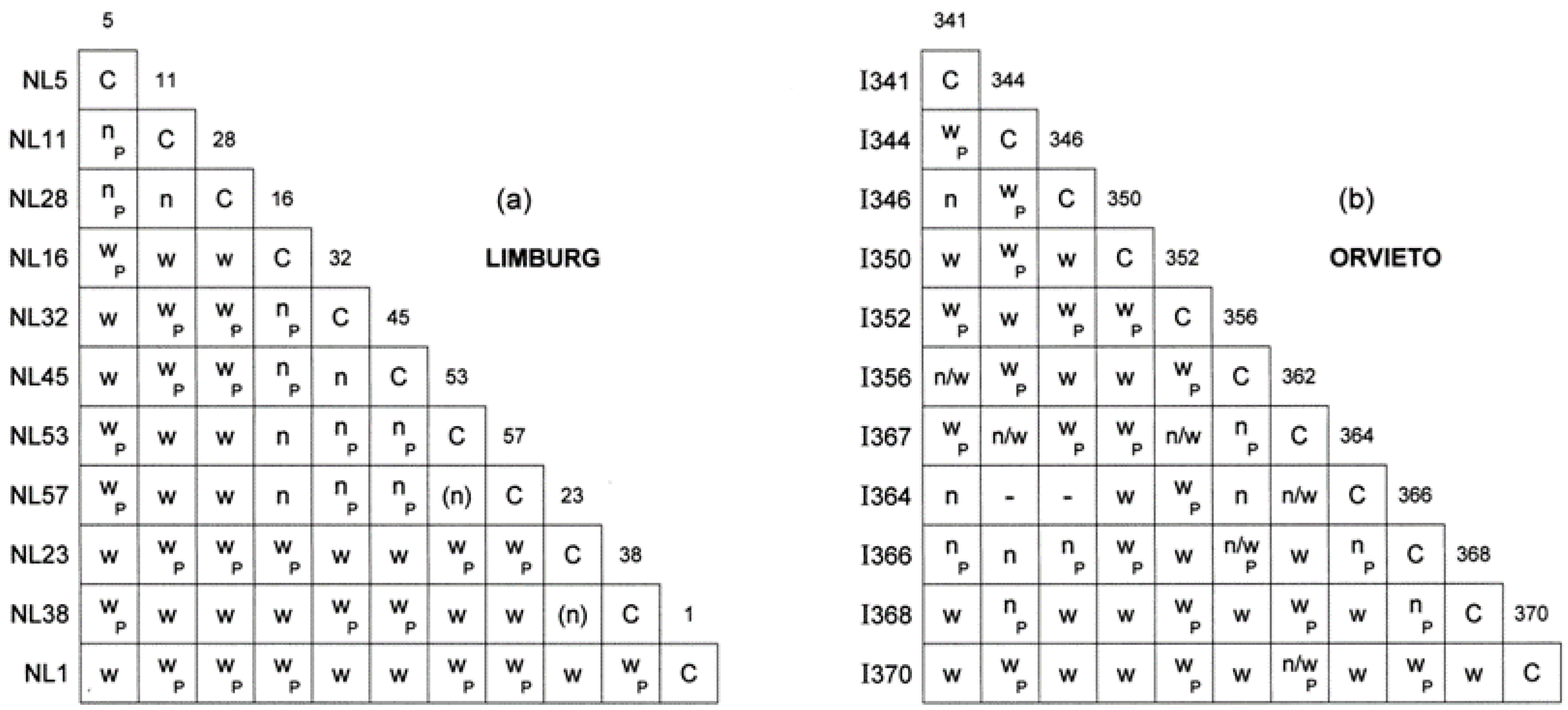
3.2. Vegetative Compatibility Type Diversity in the 2008 Limburg and Orvieto Populations
3.3. Growth Rates, Colony Patterns, Up-Mut Dimorphism and Virus Infection in the 2008 Limburg and Orvieto Hybrids
3.4. Comparative Pathogenicity of the 2008 Limburg and Orvieto Hybrids
3.5. Perithecial Form in the Limburg and Orvieto Hybrids
3.6. Frequency of the SSAM and SSNU Cu and Col-1 Genes
4. Discussion
4.1. Trends in Characters Associated with O. ulmi > O. novo-ulmi Introgression
4.2. Trends in Characters Associated with SSAM × SSNU Hybridisation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brasier, C.M. Episodic selection as a force in fungal microevolution with special reference to clonal speciation and hybrid introgression. Can. J. Bot. 1995, 73, 1213–1221. [Google Scholar] [CrossRef]
- Brasier, C.M. Rise of the hybrid fungi. Nature 2000, 405, 134–135. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M. Rapid evolution of introduced plant pathogens via interspecific hybridization. Bioscience 2001, 51, 123–133. [Google Scholar] [CrossRef]
- Depotter, R.L.; Seidl, R.F.; Wood, T.A.; Thomma, B.P.H.J. Interspecific hybrization impacts host range and pathogenicity of filamentous microbes. Curr. Op. Microbiol. 2016, 32, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Stukenbrock, E.H. The Role of Hybridization in the evolution and emergence of new fungal plant pathogens. Phytopathology 2016, 106, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M.; Cooke, D.; Duncan, J.M. Origins of a new Phytophthora pathogen through interspecific hybridisation. Proc. Natl. Acad. Sci. USA 1999, 96, 5878–5883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newcombe, G.; Stirling, B.; McDonald, S.; Bradshaw, H.D. Melampsora × columbiana, a natural hybrid of M. medusae and M. ocidentalis. Mycol. Res. 2000, 104, 261–274. [Google Scholar] [CrossRef]
- Gonthier, P.; Nicolotti, G.; Linzer, R.; Guglielmo, F.; Garbelotto, M. Invasion of European pine stands by a North American forest pathogen and its hybridization with a native interfertile taxon. Mol. Ecol. 2007, 16, 1389–1400. [Google Scholar] [CrossRef]
- Fabiano, S.; Garbelotto, M.; Giordano, L.; Gonthier, P. Genic introgression from an invasive exotic fungal forest pathogen increases the establishment of potential odf a sibling native pathogen. NeoBiota 2021, 65, 109–136. [Google Scholar]
- Stukenbrock, E.H.; Christiansen, F.B.; Hansen, T.T.; Dutheil, J.Y. Fusion of two divergent fungal individuals led to the recent emergence of a unique widespread pathogen species. Proc. Natl. Acad. Sci. USA 2012, 109, 10954–10959. [Google Scholar] [CrossRef] [Green Version]
- Husson, C.; Aguayo, J.; Revellin, C.; Frey, P.; Marcais, B. Evidence for homoploid speciation in Phytophthora alni supports taxonomic reclassification in this species complex. Fungal Genet. Biol. 2015, 77, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J.N. Intercontinental epidemiology of Dutch elm disease. Ann. Rev. Phytopathol. 1978, 16, 287–307. [Google Scholar] [CrossRef]
- Brasier, C.M. The population biology of Dutch elm disease: Its principal features and some implications for other host-pathogen systems. Adv. Plant Pathol. 1986, 5, 55–118. [Google Scholar]
- Brasier, C.M.; Mitchell, A.G.; Kirk, S.A. Decline of the non-aggressive subgrouop at current epidemic fronts. In Report on Forest Research; HMSO: London, UK, 1990; p. 53. [Google Scholar]
- Brasier, C.M. China and the origins of Dutch elm disease: An appraisal. Plant Pathol. 1990, 39, 5–16. [Google Scholar] [CrossRef]
- Brasier, C.M. Intercontinental spread and continuing evolution of the Dutch elm disease pathogens. In The Elms: Breeding, Conservation and Disease Management; Dunne, C.P., Ed.; Kluwer Academic Publishers: Boston, MA, USA, 2000; pp. 61–72. [Google Scholar]
- Brasier, C.M.; Buck, K.W. Rapid evolutionary changes in a globally invading pathogen, the causal agent of Dutch elm disease. Biol. Invasions 2002, 3, 223–233. [Google Scholar] [CrossRef]
- Brasier, C.M. Ophiostoma novo-ulmi sp. nov., causative agent of the current Dutch elm disease pandemic. Mycopathologia 1991, 115, 151–161. [Google Scholar] [CrossRef]
- Bates, M.R.; Buck, K.W.; Brasier, C.M. Molecular relationships between Ophiostoma ulmi and the EAN and NAN races of O. novo-ulmi determined by restriction fragment length polymorphisms of nuclear DNA. Mycol. Res. 1993, 97, 449–455. [Google Scholar] [CrossRef]
- Bates, M.R.; Buck, K.W.; Brasier, C.M. Molecular relationships of the mitochondrial DNA of Ophiostoma ulmi and the NAN and EAN races of O. novo-ulmi determined by restriction fragment length polymorphisms. Mycol. Res. 1993, 97, 1093–1100. [Google Scholar] [CrossRef]
- Pipe, N.D.; Brasier, C.M.; Buck, K.W. Evolutionary relationship of the Dutch elm disease fungus Ophiostoma novo-ulmi to other Ophiostoma species investigated by RFLP analysis of the rDNA region. J. Phytopathol. 2000, 148, 533–539. [Google Scholar] [CrossRef]
- Brasier, C.M.; Mehrotra, M.D. Ophiostoma himal-ulmi sp. nov., a new species of Dutch elm disease fungus endemic to the Himalayas. Mycol. Res. 1995, 99, 205–215. [Google Scholar] [CrossRef]
- Masuya, M.; Brasier, C.; Ichihara, Y.; Kubono, T.; Kanzaki, N. First report of the Dutch elm disease pathogens Ophiostoma ulmi and O. novo-ulmi in Japan. Plant Pathol. New Dis. Reps. 2009, 20, 6. [Google Scholar] [CrossRef]
- Brasier, C.M.; Kirk, S.A. Designation of the EAN and NAN races of Ophiostoma novo-ulmi as subspecies. Mycol. Res. 2001, 105, 547–554. [Google Scholar] [CrossRef]
- Brasier, C.M. Dual origin of recent Dutch elm disease outbreaks in Europe. Nature 1979, 281, 78–79. [Google Scholar] [CrossRef]
- Brasier, C.M. A comparison of pathogenicity and cultural characteristics in the EAN and NAN aggressive sub-groups of Ophiostoma ulmi. Trans. Brit. Mycol. Soc. 1986, 87, 1–13. [Google Scholar] [CrossRef]
- Brasier, C.M.; Gibbs, J.N. Origin of the Dutch elm disease epidemic in Britain. Nature 1973, 242, 607–609. [Google Scholar] [CrossRef]
- Brasier, C.M.; Kirk, S.A. Rapid changes in O. novo-ulmi population structure at current epidemic fronts. In Report on Forest Research; HMSO: London, UK, 1991; pp. 37–38. [Google Scholar]
- Brasier, C.M.; Kirk, S.A. Survival of clones of NAN Ophiostoma novo-ulmi around its probable centre of appearance in North America. Mycol. Res. 2000, 104, 1322–1332. [Google Scholar] [CrossRef]
- Webber, J.F.; Brasier, C.M. The transmission of Dutch elm disease: A study of the processes involved. In Invertebrate-Microbial Interactions; Anderson, J.M., Rayner, A.D.M., Walton, D., Eds.; Cambridge University Press: Cambridge, UK, 1984; pp. 271–306. [Google Scholar]
- Webber, J.F.; Brasier, C.M.; Mitchell, A.G. The role of the saprophytic phase in Dutch elm disease. In Plant Infecting Fungi; Pegg, G.F., Ayres, P.G., Eds.; Cambridge University Press: Cambridge, UK, 1988; pp. 298–313. [Google Scholar]
- Lea, J.; Brasier, C.M. A fruiting succession in Ceratocystis ulmi and its role in Dutch elm disease. Trans. Brit. Mycol. Soc. 1983, 80, 381–387. [Google Scholar] [CrossRef]
- Brasier, C.M. Mites and reproduction in Ceratocystis ulmi and other fungi. Trans. Brit. Mycol. Soc. 1978, 70, 81–89. [Google Scholar] [CrossRef]
- Brasier, C.M. Inter-mycelial recognition systems in Ceratocystis ulmi: Their physiological properties and ecological importance. In The Ecology and Physiology of the Fungal Mycelium; Jennings, D., Rayner, A.D.M., Eds.; Cambridge University Press: Cambridge, UK, 1984; pp. 451–497. [Google Scholar]
- Brasier, C.M. Rapid changes in genetic structure of epidemic populations of Ophiostoma ulmi. Nature 1988, 332, 538–541. [Google Scholar] [CrossRef]
- Brasier, C.M. Low genetic diversity of the Ophiostoma novo-ulmi population in North America. Mycologia 1996, 89, 722–726. [Google Scholar]
- Bates, M.R. DNA Polymorphism in the Dutch Elm Disease Fungus Ophiostoma ulmi. Ph.D. Thesis, Imperial College, London, UK, 1990. Available online: https://spiral.imperial.ac.uk/handle/10044/1/47767 (accessed on 4 June 2021).
- Brasier, C.M.; Buck, K.W.; Paoletti, M.; Crawford, L.; Kirk, S.A. Molecular analysis of evolutionary changes in populations of Ophiostoma novo-ulmi. Investig. Agrar. Sist. Recur. For. 2004, 13, 93–103. [Google Scholar]
- Brasier, C.M. The d-factor in Ceratocystis ulmi: Its biological characteristics and implications for Dutch elm disease. In Fungal Virology; Buck, K.W., Ed.; C.R.C. Press: Boca Raton, FL, USA, 1986; pp. 177–208. [Google Scholar]
- Mitchell, A.G.; Brasier, C.M. Contrasting structure of European and North American populations of Ophiostoma ulmi. Mycol. Res. 1994, 98, 576–582. [Google Scholar] [CrossRef]
- Brasier, C.M. A cytoplasmically transmitted disease of Ceratocystis ulmi. Nature 1983, 305, 220–223. [Google Scholar] [CrossRef]
- Brasier, C.M. Viruses as biological control agents of the Dutch elm disease pathogens. In The Elms: Breeding, Conservation and Disease Management; Dunne, C.P., Ed.; Kluwer Academic Publishers: Boston, MA, USA, 2000; pp. 201–212. [Google Scholar]
- Hong, V.; Dover, S.L.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Multiple mitochondrial viruses in an isolate of the Dutch elm disease fungus Ophiostoma novo-ulmi. Virology 1999, 258, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, K.W.; Brasier, C.M. Viruses of the Dutch elm disease pathogens. In Molecular Variability of Double Stranded RNA. Concepts and Applications in Agriculture and Forestry and Medicine; Tavantzis, S., Ed.; CRC Press: Boca Raton, FL, USA, 2001; pp. 165–190. [Google Scholar]
- Buck, K.W.; Brasier, C.M.; Paoletti, M.; Crawford, L. Virus transmission and gene flow between two species of Dutch elm disease fungi, Ophiostoma ulmi and O.novo-ulmi: Deleterious viruses as selective agents for gene introgression. In Genes in the Environment; Hails, R.S., Beringer, J.E., Godfrey, H.A., Eds.; Blackwells: Oxford, UK, 2002; pp. 26–45. [Google Scholar]
- Paoletti, M.; Buck, K.W.; Brasier, C.M. Selective acquisition of novel mating type and vegetative incompatibility genes via interspecies gene transfer in the globally invading eukaryote Ophiostoma novo-ulmi. Mol. Ecol. 2006, 15, 249–263. [Google Scholar] [CrossRef]
- Brasier, C.M.; Kirk, S.A.; Pipe, N.; Buck, K.W. Rare hybrids in natural populations of the Dutch elm disease pathogens Ophiostoma ulmi and O. novo-ulmi. Mycol. Res. 1998, 102, 45–257. [Google Scholar] [CrossRef]
- Kirisits, T.; Konrad, H. Dutch elm disease in Austria. For. Res. Syst. 2004, 13, 81–92. [Google Scholar]
- Bates, M.R.; Brasier, C.M.; Buck, K.W. Dutch elm disease. Source of the rapid variation in the aggressive subgroup at epidemic fronts. In Report on Forest Research; HMSO: London, UK, 1990; p. 54. [Google Scholar]
- Kile, G.A.; Brasier, C.M. Inheritance and inter-relationship of fitness characters in progeny of aggressive × non-aggressive crosses of Ophiostoma ulmi. Mycol. Res. 1990, 94, 514–522. [Google Scholar] [CrossRef]
- Brasier, C.M.; Kirk, S.A. Rare hybrids may be a genetic bridge between Ophiostoma ulmi and O. novo-ulmi. In Report on Forest Research; HMSO: London, UK, 1993; p. 11. [Google Scholar]
- Abdelali, A.; Brasier, C.M.; Bernier, L. Localization of a pathogenicity gene in Ophiostoma novo-ulmi and evidence that it may be introgressed from O. ulmi. Mol. Plant Microbe Interact. 1999, 12, 6–15. [Google Scholar]
- Pipe, N.D.; Brasier, C.M.; Buck, K.W. Two natural ceratoulmin deficient mutants of Ophiostoma novo-ulmi: One has an introgressed O. ulmi cu gene; the other has an O. novo-ulmi cu gene with a mutation in an intron splice concensus sequence. Mol. Plant Pathol. 2001, 1, 379–382. [Google Scholar] [CrossRef]
- Et-Touil, A.; Dusabenyagasani, M.; Bouvet, G.F.; Brasier, C.M.; Bernier, L. Ophiostoma ulmi DNA naturally introgressed into an isolate of Ophiostoma novo-ulmi is clustered around pathogenicity and mating type loci. Phytoprotection 2019, 99, 1–11. [Google Scholar] [CrossRef]
- Hessenauer, P.; Fijarczyk, A.; Martin, H.; Prunier, J.; Charron, G.; Chapuis, J.; Bernier, L.; Tanguay, P.; Hamelin, R.; Landry, C. Hybridization and introgression drive genome evolution of the Dutch elm disease pathogens. Nat. Ecol. Evol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Millgroom, M.G.; Brasier, C.M. Potential diversity of vegetative compatibility types of Ophiostoma novo-ulmi in North America. Mycologia 1997, 89, 722–726. [Google Scholar] [CrossRef]
- Brasier, C.M.; Gadgil, P. Dutch elm disease outbreak in New Zealand. In Report on Forest Research; HMSO: London, UK, 1992; p. 33. [Google Scholar]
- Hoegger, P.J.; Binz, T.; Heiniger, U. Detection of genetic variation between Ophiostoma ulmi and the EAN and NAN races of O. novo-ulmi determined by restriction fragment length polymorphisms of the nuclear DNA. Mycol. Res. 1996, 97, 449–455. [Google Scholar]
- Jeng, R.S.; Bernier, L.; Brasier, C.M. A comparative study of cultural and electrophoretic characteristics of the Eurasian and North American races of Ophiostoma ulmi. Can. J. Bot. 1988, 66, 1325–1333. [Google Scholar] [CrossRef]
- Pipe, N.D.; Buck, K.W.; Brasier, C.M. Molecular relationships between Ophiostoma ulmi and the EAN and NAN races of O. novo-ulmi determined by RAPD markers. Mycol. Res. 1995, 99, 653–658. [Google Scholar] [CrossRef]
- Konrad, H.; Kirisits, T.; Riegler, M.; Halmschlager, E.; Stauffer, C. Genetic evidence for natural hybridisation between the Dutch elm disease pathogens Ophiostoma novo-ulmi ssp. novo-ulmi and Ophiostoma novo-ulmi ssp. americana. Plant Pathol. 2002, 51, 78–84. [Google Scholar] [CrossRef]
- Santini, A.; Montaghi, A.; Vendramin, G.G.; Capretti, P. Analysis of the Italian Dutch elm disease population. J. Phytopathol. 2005, 153, 73–79. [Google Scholar] [CrossRef]
- Paoletti, M.; Buck, K.W.; Brasier, C.M. Cloning and sequence analysis of the Mat-B (Mat-2) genes from the three Dutch elm disease pathogens Ophiostoma ulmi, O. novo-ulmi and Ophiostoma himal-ulmi. Mycol. Res. 2005, 109, 983–991. [Google Scholar] [CrossRef]
- Brasier, C.M.; Takai, S.; Nordin, J.H.; Richards, W.C. Differences in ceratoulmin production between EAN, NAN and non-aggressive subgroups of Ophiostoma ulmi. Plant Pathol. 1990, 39, 31–236. [Google Scholar] [CrossRef]
- Brasier, C.M. Dutch elm disease—Ophiostoma (Ceratocystis) ulmi. The emergence of EAN and NAN hybrids in Europe. In Report on Forest Research; HMSO: London, UK, 1986; p. 37. [Google Scholar]
- Dvořák, M.; Tomšovský, M.; Jankovský, L.; Novotný, D. Contribution to identify the causal agents of Dutch elm disease in the Czech Republic. Plant Protect. Sci. 2007, 43, 142–145. [Google Scholar] [CrossRef] [Green Version]
- Solla, A.; Dacasa, M.C.; Nasmith, C.; Hubbes, M.; Gil, L. Analysis of Spanish populations of Ophiostoma ulmi and O. novo-ulmi using phenotypic characters and RAPD markers. Plant Pathol. 2008, 57, 33–44. [Google Scholar]
- Brasier, C.M.; Kirk, S.A. Rapid emergence of hybrids between the two subspecies of Ophiostoma novo-ulmi with a high level of pathogenic fitness. Plant Pathol. 2010, 59, 186–199. [Google Scholar] [CrossRef]
- Katović, Z.; Krstin, L.; Ćaleta, B.; Stanĉin, P.; Zebec, M.; Ćurković-Peria, M. Identification and characteriosation of the causal agent of Dutch elm disease in Croatia. Eur. J. For. Res. 2020, 139, 805–815. [Google Scholar]
- Brasier, C.M.; Webber, J.F. Is there evidence for post-epidemic attenuation in the Dutch elm disease pathogen Ophiostoma novo-ulmi? Plant Pathol. 2019. [Google Scholar] [CrossRef]
- Webber, J.F. Relative effectiveness of Scolytus scolytus, S. multistriatus and S. kirschi as vectors of Dutch elm disease. Eur. J. For. Pathol. 1990, 184–192. [Google Scholar] [CrossRef]
- Favaro, A.; Battista, A. Observations on the elm bark beetle Scolytus pygmaeus as a possible vector of the fungus Ophiostoma ulmi. Redia 1993, 76, 459–466. [Google Scholar]
- Brasier, C.M. Laboratory investigation of Ceratocystis ulmi. In Compendium of Elm Diseases; Stipes, R.J., Campana, R.J., Eds.; American Phytopathological Society: St Paul, MN, USA, 1981; pp. 76–79. [Google Scholar]
- Tchernoff, B. Methods for screening and for the rapid selection of elms for resistance to Dutch elm disease. Acta Bot. Neer. 1965, 14, 409–452. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/- (accessed on 20 April 2019).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Core Team: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-137. 2018. Available online: https://CRAN.R-project.org/package=nlme (accessed on 20 April 2019).
- Rogers, H.J.; Buck, K.W.; Brasier, C.M. The molecular nature of the d2 factor in Ceratocystis ulmi. In Fungal Virology; Buck, K.W., Ed.; CRC Press: Boca Raton, FL, USA, 1986; pp. 109–220. [Google Scholar]
- Pipe, N.; Buck, K.W.; Brasier, C.M. Comparison of cerato-ulmin (cu) gene sequences of the recently identified Himalayan Dutch elm disease fungus Ophiostoma himal-ulmi with those of O. ulmi and O. novo-ulmi suggests the cu gene of O. novo-ulmi is unlikely to have been acquired recently from O. himal-ulmi. Mycol. Res. 1997, 101, 415–421. [Google Scholar]
- Pereira, V.; Hintz, W.E.; Royer, J.C.; Field, D.J. A gene associated with filamentous growth in Ophiostoma novo-ulmi has RNA-binding motifs and is similar to a yeast gene involved in RNA splicing. Curr. Genet. 2000, 37, 94–103. [Google Scholar] [CrossRef]
- Takai, S. Pathogenoicity and cerato-ulmin production in Ceratocystis ulmi. Nature 1974, 252, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Webber, J.F. Influence of the d2 factor on survival and infection by the Dutch elm disease pathogen Ophiostoma ulmi. Plant Pathol. 1987, 36, 531–538. [Google Scholar] [CrossRef]
- Sutherland, M.L.; Brasier, C.M. A comparison of thirteen d-factors as potential biological control agents of Ophiostoma novo-ulmi. Plant Pathol. 1997, 46, 680–693. [Google Scholar] [CrossRef]
- Bowden, C.G.; Hintz, W.E.; Jeng, R.; Hubbes, M.; Horgen, P.A. Isolation and characterization of the cerato-ulmin toxin gene of the Dutch elm disease pathogen, Ophiostoma ulmi. Curr. Genet. 1994, 25, 323–329. [Google Scholar] [CrossRef]
- Brasier, C.M. Fitness, continuous variation and selection in fungal populations: An ecological perspective. In The Structure of Fungal Populations; Worrall, J., Ed.; Kluwer Academic Publishers: Boston, MA, USA; New York, NY, USA, 1998; pp. 289–318. [Google Scholar]
- Rieser, L.H.; Archer, A.A.; Wayne, R.K. Transgressive hybridization, adaptation and speciation. Heredity 1999, 83, 363–372. [Google Scholar]
- Goldschmidt, R.B. The Material Basis of Evolution; Yale University Press: New Haven, CT, USA, 1940. [Google Scholar]
- Brasier, C. Some genetical aspects of necrotophy with special reference to Ophiostoma ulmi. In Genetics and Plant Pathogenesis; Day, P.R., Jellis, G.J., Eds.; Blackwells: Oxford, UK, 1987; pp. 297–310. [Google Scholar]
- Brasier, C.M.; Webber, J.F. Positive correlations between in vitro growth rate and pathogenesis in Ophiostoma ulmi. Plant Pathol. 1987, 36, 462–466. [Google Scholar] [CrossRef]
- Santini, A.; Pecori, F.; Pepori, A.; Brookes, A. ‘Morfeo’ Elm: A new variety resistant to Dutch elm disease. For. Pathol. 2012, 42, 171–176. [Google Scholar] [CrossRef]
- Martín, J.A.; Sobrino-Plata, J.; Rodríguez-Calcerrada, J. Breeding and scientific advances in the fight against Dutch elm disease: Will they allow the use of elms in forest restoration? New For. 2019, 50, 183–215. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Brasier, C.M.; Gibbs, J.N. Inheritance of pathogenicity and cultural characters in Ceratocystis ulmi: I. Hybridisation of aggressive and non-aggressive strains. Ann. Appl. Biol. 1976, 83, 31–37. [Google Scholar] [CrossRef]
- Brasier, C.M. Inheritance of pathogenicity and cultural characters in Ceratocystis ulmi: Hybridisation of protoperithecial and non-aggressive strains. Trans. Brit. Mycol. Soc. 1977, 68, 45–52. [Google Scholar] [CrossRef]


{kind=link}
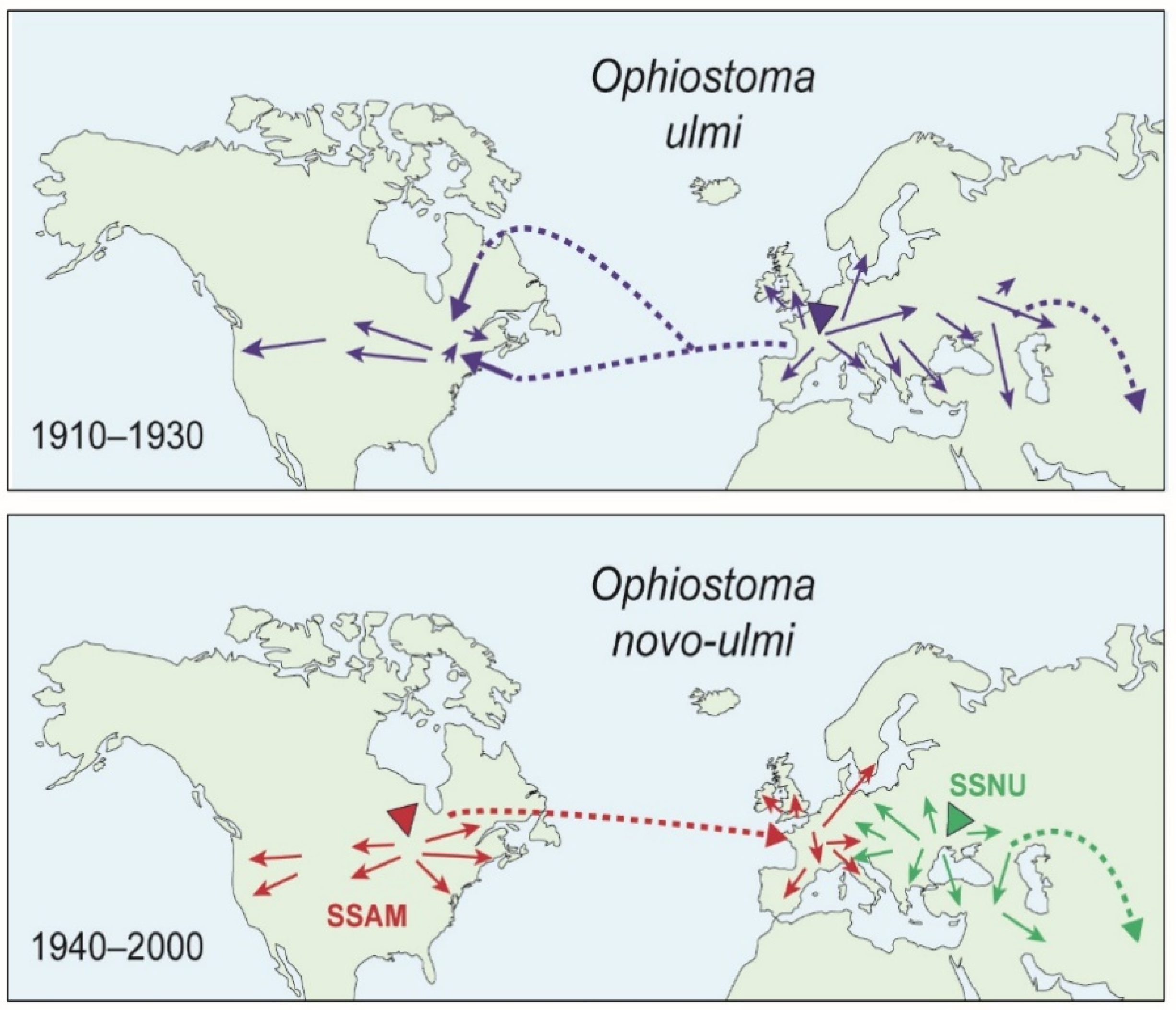{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample and Date | Limburg | Orvieto | ||||
|---|---|---|---|---|---|---|
| No. of Isolates | % Hybrid | % MAT-1 | No. of Isolates | % Hybrid | % MAT-1 | |
| Early subspecies overlap phase, 1979–1980 | 22 | 4.5 1 | 4.5 3 | 15 | ~0 2 | 0 |
| Advanced hybrid phase, 1983–1986 | 96 | >79 4 | 35.4 3 | 108 | >72 4 | 10.5 3 |
| Hybrid swarm phase, 2008 5 | 56 | ~100 | 42.8 | 25 | ~100 | 28.0 |
| Sample 1,2 | Mean Growth Rate and SE mm /Day−1 | Mean Pathogenicity (% Defoliation) and SE | |
|---|---|---|---|
| English Elm SR4 | Commelin Elm 270 | ||
| 2008 Orvieto | 3.24 ± 0.08 (55) 1 | 42.4 ± 4.5 (15) | 17.1 ± 2.2 (15) |
| 2008 Limburg | 3.54 ± 0.05 (26) | 38.2 ± 4.5 (15) | 12.0 ± 1.9 (15) |
| 1986 Limburg | nt 3 | 41.9 ± 4.5 (15) | 18.0 ± 2.2 (15) |
| Sample and Date | Limburg | Orvieto | ||
|---|---|---|---|---|
| No. of Isolates | % Up-Mut | No. of Isolates | % Up-Mut | |
| Early subsp. overlap phase 1979–1980 | 22 | 36.4 1 | 15 | 53.3 1 |
| Advanced hybrid phase 1983–1986 | 107 | 52.3 2 | 92 | 34.8 2 |
| Hybrid swarm phase 2008 | 56 | 0 | 25 | 0 |
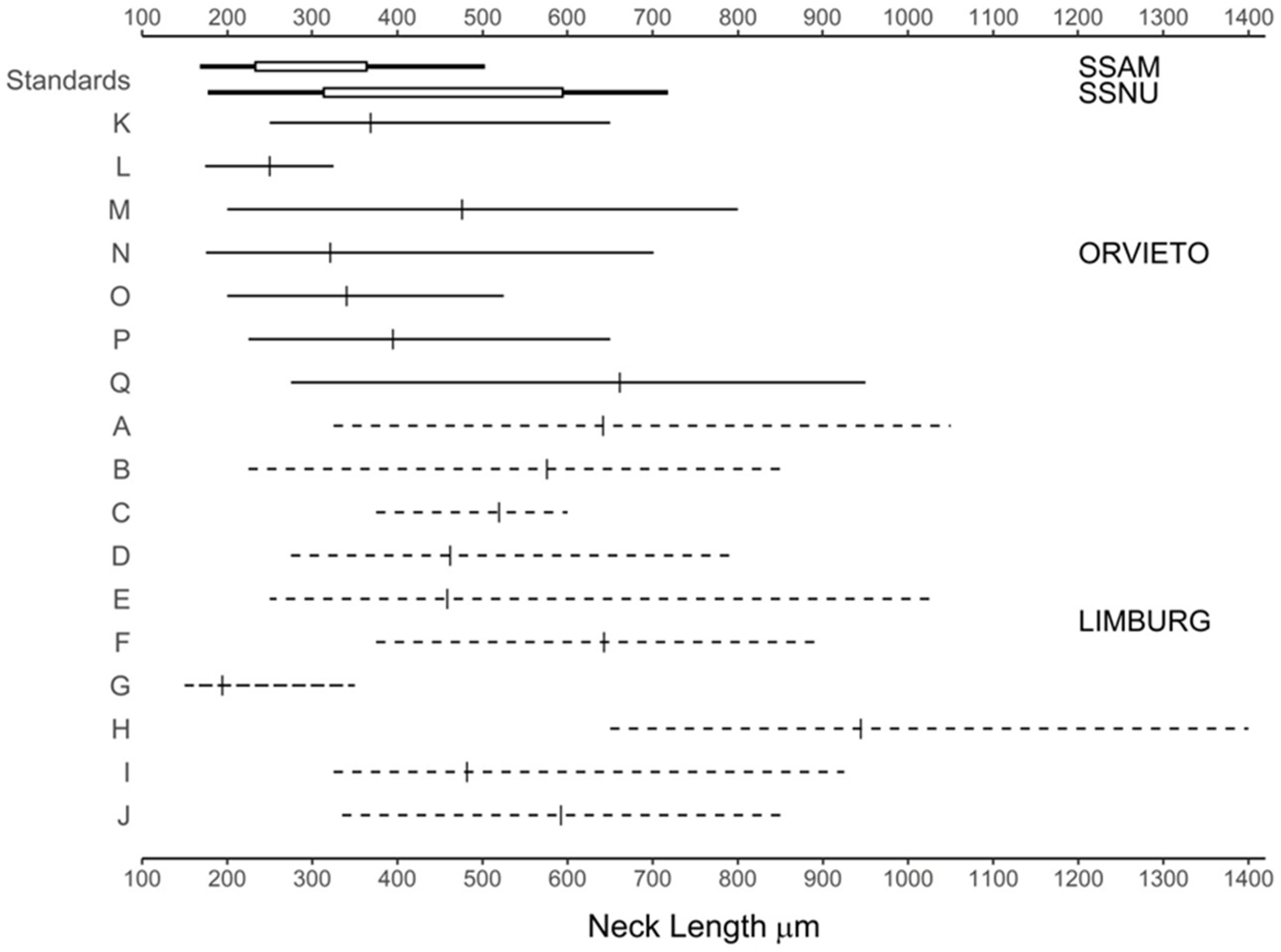
| Groups | No. of Pairings | Neck Length (NL) μm | Bowl Width (BW) μm | NL: BW Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Contrast | Mean | Contrast | Mean | Contrast | |||||
| 1 3 | 2 4 | 1 | 2 4 | 1 | 2 4 | |||||
| All data | ||||||||||
| Orvieto | 7 | 393.1 ± 57.4 | NS | NS | 120.0 ± 4.5 | NS | NS | 3.32 ± 0.40 | NS | NS |
| Limburg | 10 | 535.2 ± 56.3 | 123.2 ± 3.8 | 4.35 ± 0.38 | ||||||
| Adjusted1 | ||||||||||
| Orvieto | 5 | 378.3 ± 26.0 | a | a | 119.7 ± 5.0 | a | a | 3.19 ± 0.31 | ab | ab |
| Limburg | 9 | 556.8 ± 28.4 | b | b | 120.0 ± 4.2 | a | a | 4.66 ± 0.32 | c | b |
| Subspecies2 | ||||||||||
| SSAM | 10 | 294.0 ± 129.7 | a | 115.5 ± 12.5 | a | 2.52 ± 0.34 | a | |||
| SSNU | 10 | 444.0 ± 162.8 | b | 103.4 ± 12.4 | a | 4.30 ± 0.42 | bc | |||
| Subsp. mean | 20 | 366.6 ± 45.03 | a | 109.4 ± 8.1 | a | 3.41 ± 0.24 | a | |||
| Locus Combination | No. of Isolates | Total | of Which | |||
|---|---|---|---|---|---|---|
| cu Locus | col-1 Locus | Limburg | Orvieto | MAT-1 | MAT-2 | |
| SSNU | SSNU | 5 | 5 | 10 | 5 | 5 |
| SSAM | SSAM | 2 | 5 | 7 | 5 | 2 |
| SSNU | SSAM | 7 | 3 | 10 | 4 | 6 |
| SSAM | SSNU | 0 | 1 | 1 | 0 | 1 |
| Character | Trend | Inferred Evolutionary Process |
|---|---|---|
| Substantially O. ulmi > O. novo-ulmi Introgression-Related | ||
| MAT-1 frequency | Increase from near zero to 28% (Orvieto) and 43% (Limburg) | Directional selection (under virus pressure) |
| Diversity of vc types | Change from near clonal to very high diversity | Directional selection (under virus pressure) |
| Overt virus infection | Greatly reduced | Suppression by high vc diversity and ascospore production |
| Subspecies Hybridisation-Related | ||
| Growth rate | Fast, SSAM-like; slower at Orvieto | Directional selection |
| Up-mut dimorphism | Loss | Directional selection |
| Colony pattern | Loss of original subspecies patterns; highly variable | Uncertain |
| Pathogenicity | High, SSAM-like | Directional selection |
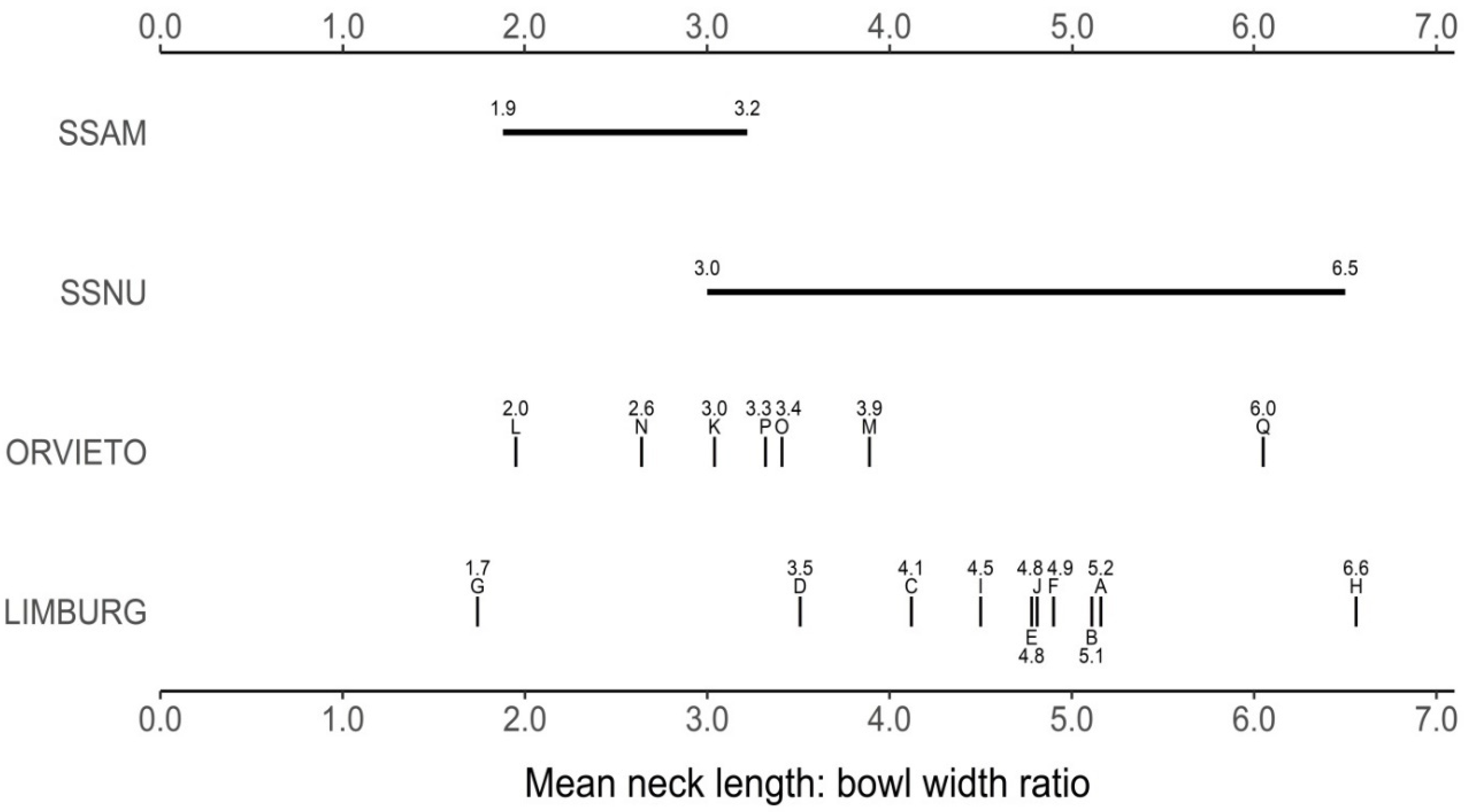
| Perithecial neck length | Limburg: extreme phenotypes; mean length significantly above combined subspecies mean | Transgressive segregation |
| Orvieto: from SSAM-like to shorter necked SSNU-like, mean not significantly different from combined subspecies mean | Transgressive segregation followed by stabilising selection | |
| Perithecial neck length: bowl width ratio | Limburg: mean ratio significantly higher than combined subspecies mean | Transgressive segregation |
| Orvieto: mean not significantly different from combined subspecies mean | Transgressive segregation followed by stabilising selection | |
| SSSAM and SSNU cu loci | Co-occurrence | Neutral |
| SSSAM and SSNU col-1 loci | Co-occurrence | Neutral |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brasier, C.; Franceschini, S.; Forster, J.; Kirk, S. Enhanced Outcrossing, Directional Selection and Transgressive Segregation Drive Evolution of Novel Phenotypes in Hybrid Swarms of the Dutch Elm Disease Pathogen Ophiostoma novo-ulmi. J. Fungi 2021, 7, 452. https://doi.org/10.3390/jof7060452
Brasier C, Franceschini S, Forster J, Kirk S. Enhanced Outcrossing, Directional Selection and Transgressive Segregation Drive Evolution of Novel Phenotypes in Hybrid Swarms of the Dutch Elm Disease Pathogen Ophiostoma novo-ulmi. Journal of Fungi. 2021; 7(6):452. https://doi.org/10.3390/jof7060452
Chicago/Turabian StyleBrasier, Clive, Selma Franceschini, Jack Forster, and Susan Kirk. 2021. "Enhanced Outcrossing, Directional Selection and Transgressive Segregation Drive Evolution of Novel Phenotypes in Hybrid Swarms of the Dutch Elm Disease Pathogen Ophiostoma novo-ulmi" Journal of Fungi 7, no. 6: 452. https://doi.org/10.3390/jof7060452
APA StyleBrasier, C., Franceschini, S., Forster, J., & Kirk, S. (2021). Enhanced Outcrossing, Directional Selection and Transgressive Segregation Drive Evolution of Novel Phenotypes in Hybrid Swarms of the Dutch Elm Disease Pathogen Ophiostoma novo-ulmi. Journal of Fungi, 7(6), 452. https://doi.org/10.3390/jof7060452