
Aequorin as a Useful Calcium-Sensing Reporter in Candida albicans


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Aequorin Expression in C. albicans
2.3. Luminescence Measurements
2.4. Mutant Construction
2.5. Yeast Transformation
2.6. Immunodetection of Aequorin
2.7. Statistics
3. Results
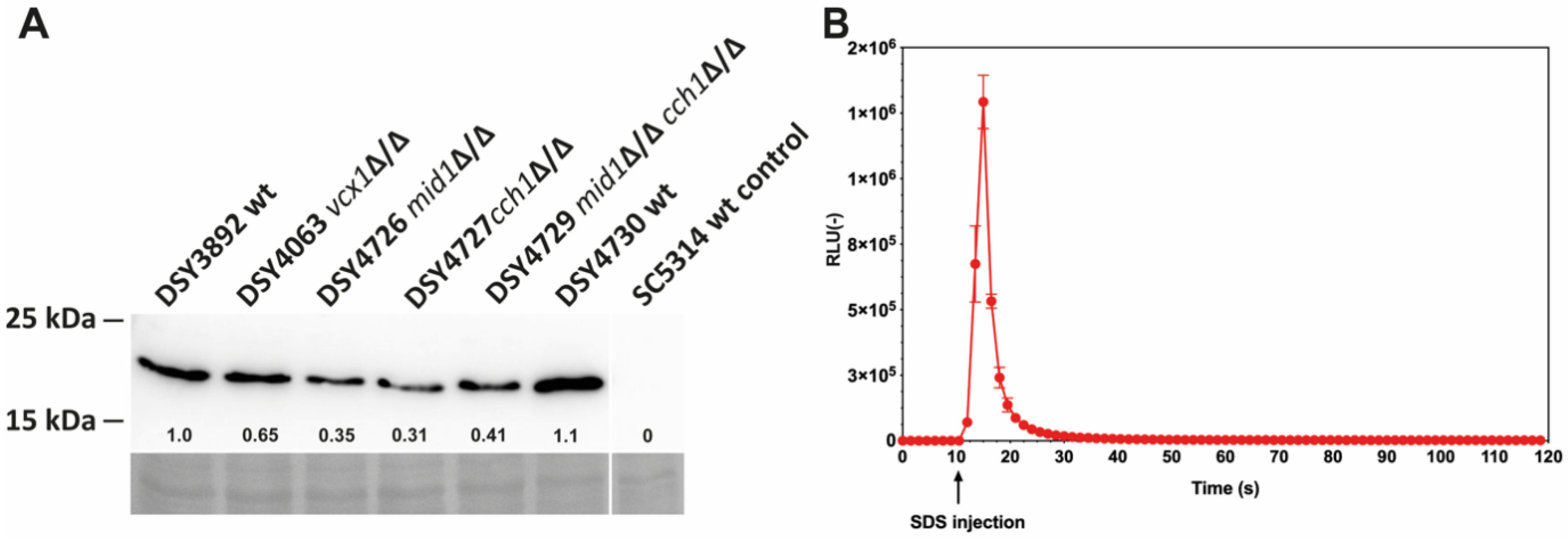
3.1. Establishing Aequorin Expression in C. albicans
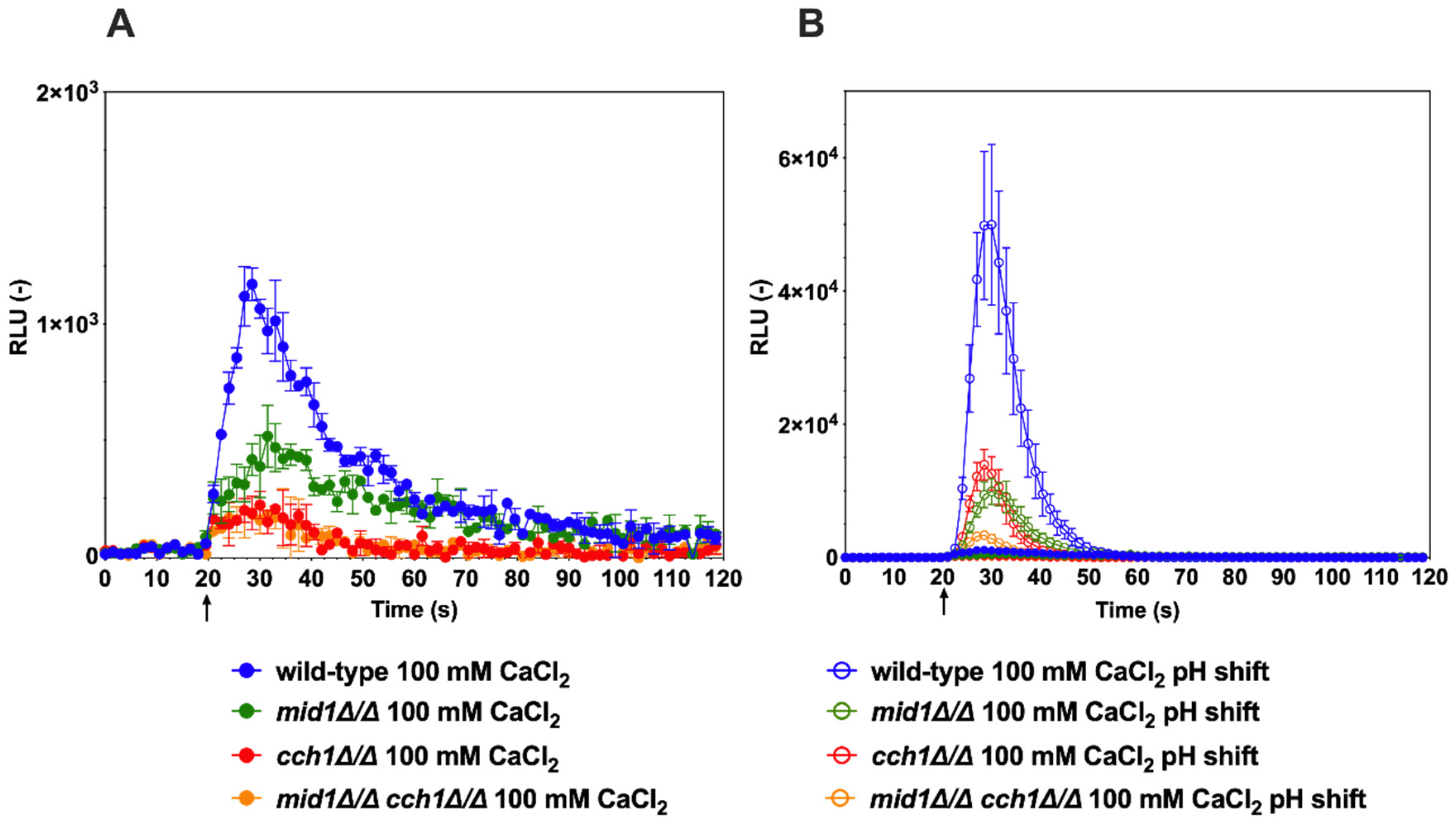
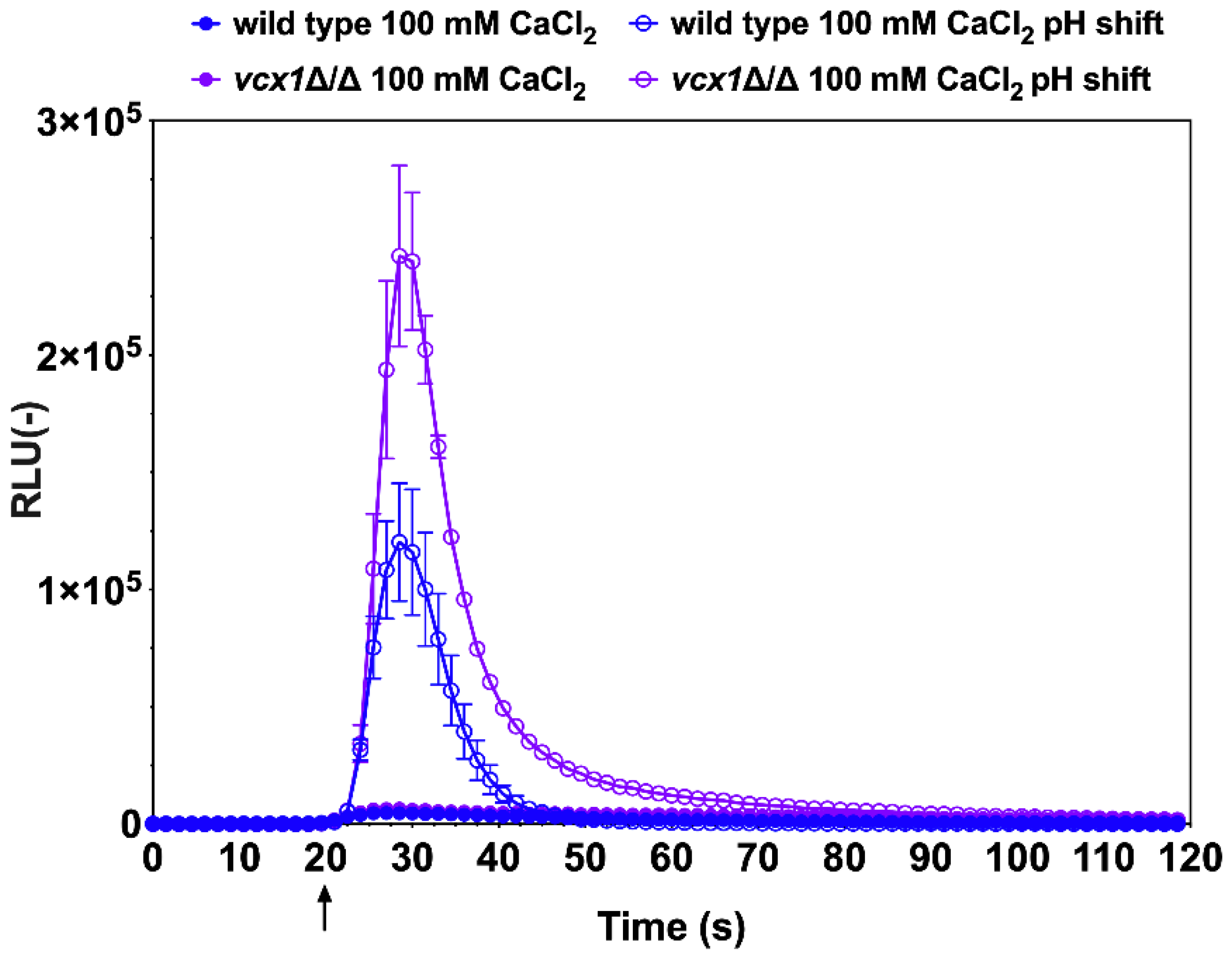
3.2. Probing of Ca2+ Flux by Alkaline Stress
3.3. Probing of Ca2+ Homeostasis by Amiodarone-Induced Stress
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lange, M.; Peiter, E. Calcium Transport Proteins in Fungi: The Phylogenetic Diversity of Their Relevance for Growth, Virulence, and Stress Resistance. Front. Microbiol. 2020, 10, 3100. [Google Scholar] [CrossRef]
- Tisi, R.; Rigamonti, M.; Groppi, S.; Belotti, F. Calcium homeostasis and signaling in fungi and their relevance forpathogenicity of yeasts and filamentous fungi. AIMS Mol. Sci. 2016, 3, 505–549. [Google Scholar] [CrossRef]
- Cottier, F.; Hall, R.A. Face/Off: The Interchangeable Side of Candida Albicans. Front. Cell. Infect. Microbiol. 2020, 9, 471. [Google Scholar] [CrossRef]
- Brand, A.; Lee, K.; Veses, V.; Gow, N.A.R. Calcium homeostasis is required for contact-dependent helical and sinusoidal tip growth in Candida albicanshyphae. Mol. Microbiol. 2009, 71, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.; Gow, N.A.R. Mechanisms of hypha orientation of fungi. Curr. Opin. Microbiol. 2009, 12, 350–357. [Google Scholar] [CrossRef]
- Karababa, M.; Valentino, E.; Pardini, G.; Coste, A.T.; Bille, J.; Sanglard, D. CRZ1, a target of the calcineurin pathway in Candida albicans. Mol. Microbiol. 2006, 59, 1429–1451. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Ischer, F.; Marchetti, O.; Entenza, J.; Bille, J. Calcineurin A of Candida albicans: Involvement in antifungal tolerance, cell morphogenesis and virulence. Mol. Microbiol. 2003, 48, 959–976. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.; Goldstein, A.L.; Blankenship, J.R.; Del Poeta, M.; Davis, D.; Cardenas, M.E.; Perfect, J.R.; McCusker, J.H.; Heitman, J. Calcineurin is essential for survival during membrane stress in Candida albicans. EMBO J. 2002, 21, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hou, Y.; Liu, W.; Lu, C.; Wang, W.; Sun, S. Components of the Calcium-Calcineurin Signaling Pathway in Fungal Cells and Their Potential as Antifungal Targets. Eukaryot. Cell 2015, 14, 324–334. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Brand, A.; Morrison, E.L.; Silao, F.G.S.; Bigol, U.G.; Malbas, F.F.; Nett, J.E.; Andes, D.R.; Solis, N.V.; Filler, S.G.; et al. Calcineurin Controls Drug Tolerance, Hyphal Growth, and Virulence in Candida dubliniensis. Eukaryot. Cell 2011, 10, 803–819. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, H.; Cheng, X.; Xu, N.; Ding, X.; Xing, L.; Li, M. Roles of Cch1 and Mid1 in Morphogenesis, Oxidative Stress Response and Virulence in Candida albicans. Mycopathol. 2012, 174, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Alber, J.; Wang, J.; Du, W.; Yang, X.; Li, X.; Sanglard, D.; Geyer, J. The Candida albicans plasma membrane protein Rch1p, a member of the vertebrate SLC10 carrier family, is a novel regulator of cytosolic Ca2+ homoeostasis. Biochem. J. 2012, 444, 497–502. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, F.; Zhao, Q.; Chen, J.; Zhang, B.; Ding, X.; Wang, H.; Yang, B.; Lu, G.; Zhang, B.; et al. A novel role of the vacuolar calcium channel Yvc1 in stress response, morphogenesis and pathogenicity of Candida albicans. Int. J. Med Microbiol. 2014, 304, 339–350. [Google Scholar] [CrossRef]
- Peng, L.; Yu, Q.; Wei, H.; Zhu, N.; Ren, T.; Liang, C.; Xu, J.; Tian, L.; Li, M. The TRP Ca2+ channel Yvc1 regulates hyphal reactive oxygen species gradient for maintenance of polarized growth in Candida albicans. Fungal Genet. Biol. 2019, 133, 103282. [Google Scholar] [CrossRef]
- Bates, S.; MacCallum, D.M.; Bertram, G.; Munro, C.A.; Hughes, H.B.; Buurman, E.T.; Brown, A.J.; Odds, F.C.; Gow, N.A. Candida albicans Pmr1p, a Secretory Pathway P-type Ca2+/Mn2+-ATPase, Is Required for Glycosylation and Virulence. J. Biol. Chem. 2005, 280, 23408–23415. [Google Scholar] [CrossRef] [PubMed]
- D’Hooge, P.; Coun, C.; Van Eyck, V.; Faes, L.; Ghillebert, R.; Mariën, L.; Winderickx, J.; Callewaert, G. Ca 2+ homeostasis in the budding yeast Saccharomyces cerevisiae: Impact of ER/Golgi Ca 2+ storage. Cell Calcium 2015, 58, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, A.; Bertuzzi, M.; Seidel, C.; Thomson, D.; Bignell, E.M.; Read, N.D. Live-cell imaging of rapid calcium dynamics using fluorescent, genetically-encoded GCaMP probes with Aspergillus fumigatus. Fungal Genet. Biol. 2020, 103470, 103470. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.R.; Campbell, A.K.; Smith, S.M.; Trewavas, A.J. Transgenic plant aequorin reports the effects of touch and cold-shock and elicitors on cytoplasmic calcium. Nat. Cell Biol. 1991, 352, 524–526. [Google Scholar] [CrossRef]
- Nelson, G.; Kozlova-Zwinderman, O.; Collis, A.J.; Knight, M.R.; Fincham, J.R.S.; Stanger, C.P.; Renwick, A.; Hessing, J.G.M.; Punt, P.J.; Hondel, C.A.M.J.J.V.D.; et al. Calcium measurement in living filamentous fungi expressing codon-optimized aequorin. Mol. Microbiol. 2004, 52, 1437–1450. [Google Scholar] [CrossRef]
- Batiza, A.F.; Schulz, T.; Masson, P.H. Yeast Respond to Hypotonic Shock with a Calcium Pulse. J. Biol. Chem. 1996, 271, 23357–23362. [Google Scholar] [CrossRef] [PubMed]
- Barelle, C.J.; Manson, C.L.; Maccallum, D.M.; Odds, F.C.; Gow, N.A.R.; Brown, A.J.P. GFP as a quantitative reporter of gene regulation in Candida albicans. Yeast 2004, 21, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Murad, A.M.A.; Lee, P.R.; Broadbent, I.D.; Barelle, C.J.; Brown, A.J.P. CIp10, an efficient and convenient integrating vector forCandida albicans. Yeast 2000, 16, 325–327. [Google Scholar] [CrossRef]
- Coste, A.T.; Karababa, M.; Ischer, F.; Bille, J.; Sanglard, D. TAC1, Transcriptional Activator of CDR Genes, Is a New Transcription Factor Involved in the Regulation of Candida albicans ABC Transporters CDR1 and CDR2. Eukaryot. Cell 2004, 3, 1639–1652. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-L.; Zhao, L.; Li, M.-Q.; Chen, W.-G.; Xu, C.-J. A sensitive and reversible staining of proteins on blot membranes. Anal. Biochem. 2020, 592, 113579. [Google Scholar] [CrossRef]
- Delarze, E.; Ischer, F.; Sanglard, D.; Coste, A.T. Adaptation of aGaussia princepsLuciferase reporter system in Candida albicansforin vivodetection in theGalleria mellonellainfection model. Virulence 2015, 6, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, Y.; Zhang, B.; Zheng, W.; Xing, L.; Li, M. Alkaline stress triggers an immediate calcium fluctuation in Candida albicans mediated by Rim101p and Crz1p transcription factors. FEMS Yeast Res. 2011, 11, 430–439. [Google Scholar] [CrossRef]
- Reedy, J.L.; Filler, S.G.; Heitman, J. Elucidating the Candida albicans calcineurin signaling cascade controlling stress response and virulence. Fungal Genet. Biol. 2010, 47, 107–116. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, M.; Brand, A.; Srikantha, T.; Daniels, K.J.; Soll, D.R.; Gow, N.A.R. Fig1 Facilitates Calcium Influx and Localizes to Membranes Destined To Undergo Fusion during Mating in Candida albicans. Eukaryot. Cell 2011, 10, 435–444. [Google Scholar] [CrossRef]
- Pittman, J.K. Vacuolar Ca2+ uptake. Cell Calcium 2011, 50, 139–146. [Google Scholar] [CrossRef]
- Courchesne, W.E.; Ozturk, S. Amiodarone induces a caffeine-inhibited, MID1 -depedent rise in free cytoplasmic calcium in Saccharomyces cerevisiae. Mol. Microbiol. 2003, 47, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Gamarra, S.; Rocha, E.M.F.; Zhang, Y.-Q.; Park, S.; Rao, R.; Perlin, D.S. Mechanism of the Synergistic Effect of Amiodarone and Fluconazole in Candida albicans. Antimicrob. Agents Chemother. 2010, 54, 1753–1761. [Google Scholar] [CrossRef] [PubMed]
- Tisi, R.; Martegani, E.; Brandão, R.L. Monitoring Yeast Intracellular Ca2+ Levels Using an In Vivo Bioluminescence Assay: Figure 1. Cold Spring Harb. Protoc. 2015, 2015, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Fricker, M.; Plieth, C.; Knight, H.; Blancaflor, E.; Knight, M.; White, N.; Gilroy, S. Fluorescence and Luminescence Techniques to Probe Ion Activities in Living Plant Cells. Fluoresc. Lumin. Probes Biol. Act. 1999, 569–596. [Google Scholar] [CrossRef]
- Förster, C.; Kane, P.M. Cytosolic Ca2+ Homeostasis Is a Constitutive Function of the V-ATPase in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 38245–38253. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, J.R.; Wormley, F.L.; Boyce, M.K.; Schell, W.A.; Filler, S.G.; Perfect, J.R.; Heitman, J. Calcineurin Is Essential for Candida albicans Survival in Serum and Virulence. Eukaryot. Cell 2003, 2, 422–430. [Google Scholar] [CrossRef]
- Bader, T.; Bodendorfer, B.; Schröppel, K.; Morschhäuser, J. Calcineurin Is Essential for Virulence in Candida albicans. Infect. Immun. 2003, 71, 5344–5354. [Google Scholar] [CrossRef]
- Blankenship, J.R.; Heitman, J. Calcineurin Is Required for Candida albicans To Survive Calcium Stress in Serum. Infect. Immun. 2005, 73, 5767–5774. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.; Fox, D.S.; Heitman, J. Calcineurin is required for hyphal elongation during mating and haploid fruiting in Cryptococcus neoformans. EMBO J. 2001, 20, 1020–1032. [Google Scholar] [CrossRef]
- Kraus, P.R.; Heitman, J. Coping with stress: Calmodulin and calcineurin in model and pathogenic fungi. Biochem. Biophys. Res. Commun. 2003, 311, 1151–1157. [Google Scholar] [CrossRef]
- Button, D.; Eidsath, A. Aequorin targeted to the endoplasmic reticulum reveals heterogeneity in luminal Ca++ concentration and reports agonist- or IP3-induced release of Ca++. Mol. Biol. Cell 1996, 7, 419–434. [Google Scholar] [CrossRef]
- Kim, H.-S.; Czymmek, K.J.; Patel, A.; Modla, S.; Nohe, A.; Duncan, R.; Gilroy, S.; Kang, S. Expression of the Cameleon calcium biosensor in fungi reveals distinct Ca2+ signatures associated with polarized growth, development, and pathogenesis. Fungal Genet. Biol. 2012, 49, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Viladevall, L.; Serrano, R.; Ruiz, A.; Domenech, G.; Giraldo, J.; Barceló, A.; Ariño, J. Characterization of the Calcium-mediated Response to Alkaline Stress in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 43614–43624. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Zhang, K.; Yu, Q.; Zhang, B.; Xiao, C.; Dong, Y.; Chen, Y.; Zhang, B.; Xing, L.; Li, M. Tfp1 is required for ion homeostasis, fluconazole resistance and N-Acetylglucosamine utilization in Candida albicans. Biochim. Biophys. Acta (BBA) Bioenerg. 2015, 1853, 2731–2744. [Google Scholar] [CrossRef]
- Butts, A.; DiDone, L.; Koselny, K.; Baxter, B.K.; Chabrier-Rosello, Y.; Wellington, M.; Krysan, D.J. A Repurposing Approach Identifies Off-Patent Drugs with Fungicidal Cryptococcal Activity, a Common Structural Chemotype, and Pharmacological Properties Relevant to the Treatment of Cryptococcosis. Eukaryot. Cell 2012, 12, 278–287. [Google Scholar] [CrossRef]
- Muend, S.; Rao, R. Fungicidal activity of amiodarone is tightly coupled to calcium influx. FEMS Yeast Res. 2008, 8, 425–431. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maresova, L.; Muend, S.; Zhang, Y.-Q.; Sychrova, H.; Rao, R. Membrane Hyperpolarization Drives Cation Influx and Fungicidal Activity of Amiodarone. J. Biol. Chem. 2009, 284, 2795–2802. [Google Scholar] [CrossRef]
- Zhang, Y.-Q.; Rao, R. A spoke in the wheel: Calcium spikes disrupt yeast cell cycle. Cell Cycle 2008, 7, 870–873. [Google Scholar] [CrossRef][Green Version]
- Zhang, Y.-Q.; Gamarra, S.; Garcia-Effron, G.; Park, S.; Perlin, D.S.; Rao, R. Requirement for Ergosterol in V-ATPase Function Underlies Antifungal Activity of Azole Drugs. PLoS Pathog. 2010, 6, e1000939. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanglard, D. Aequorin as a Useful Calcium-Sensing Reporter in Candida albicans. J. Fungi 2021, 7, 319. https://doi.org/10.3390/jof7040319
Sanglard D. Aequorin as a Useful Calcium-Sensing Reporter in Candida albicans. Journal of Fungi. 2021; 7(4):319. https://doi.org/10.3390/jof7040319
Chicago/Turabian StyleSanglard, Dominique. 2021. "Aequorin as a Useful Calcium-Sensing Reporter in Candida albicans" Journal of Fungi 7, no. 4: 319. https://doi.org/10.3390/jof7040319
APA StyleSanglard, D. (2021). Aequorin as a Useful Calcium-Sensing Reporter in Candida albicans. Journal of Fungi, 7(4), 319. https://doi.org/10.3390/jof7040319