
Novel Cyclic Peptides from Lethal Amanita Mushrooms through a Genome-Guided Approach

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Preservation of Samples
2.2. Genome and Transcriptome Sequencing
2.3. Extraction of Cyclic Peptides
2.4. HPLC Analysis
2.5. Mining MSDIN Genes
2.6. Predicted Molecular Masses of Candidate Peptide Sequences
2.7. LC–MS and LC–MS/MS Analyses
2.8. Cyclic Peptide Synthesis
3. Results
3.1. Amanita Genomes
3.2. MSDIN Genes for Candidate Cyclic Peptides
3.3. Transcriptome of As
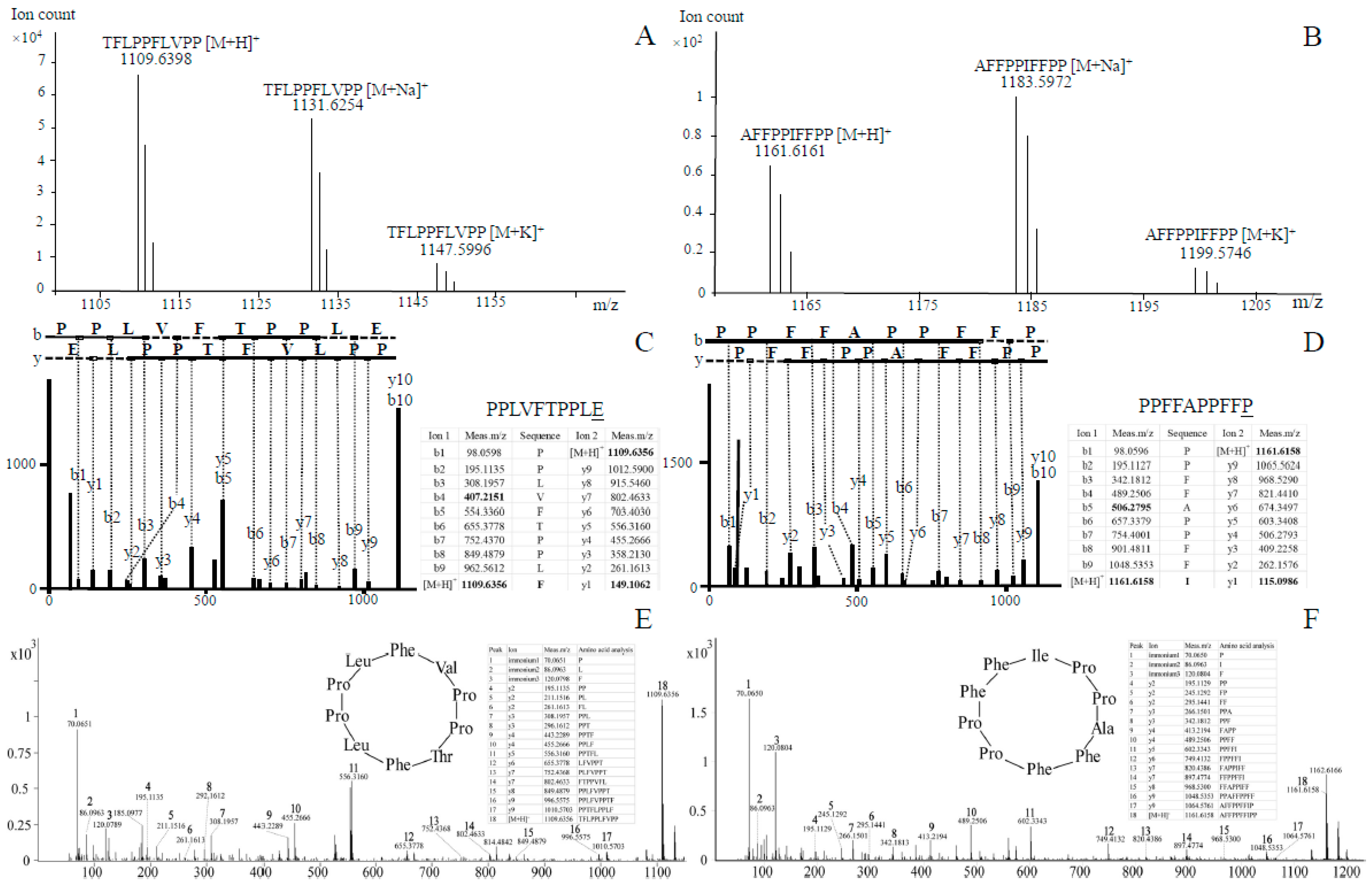
3.4. LC–HRMS and LC–MS/MS Analyses of Novel Cyclic Peptides in As
3.5. Novel Cyclic Peptides in Other Amanita Species
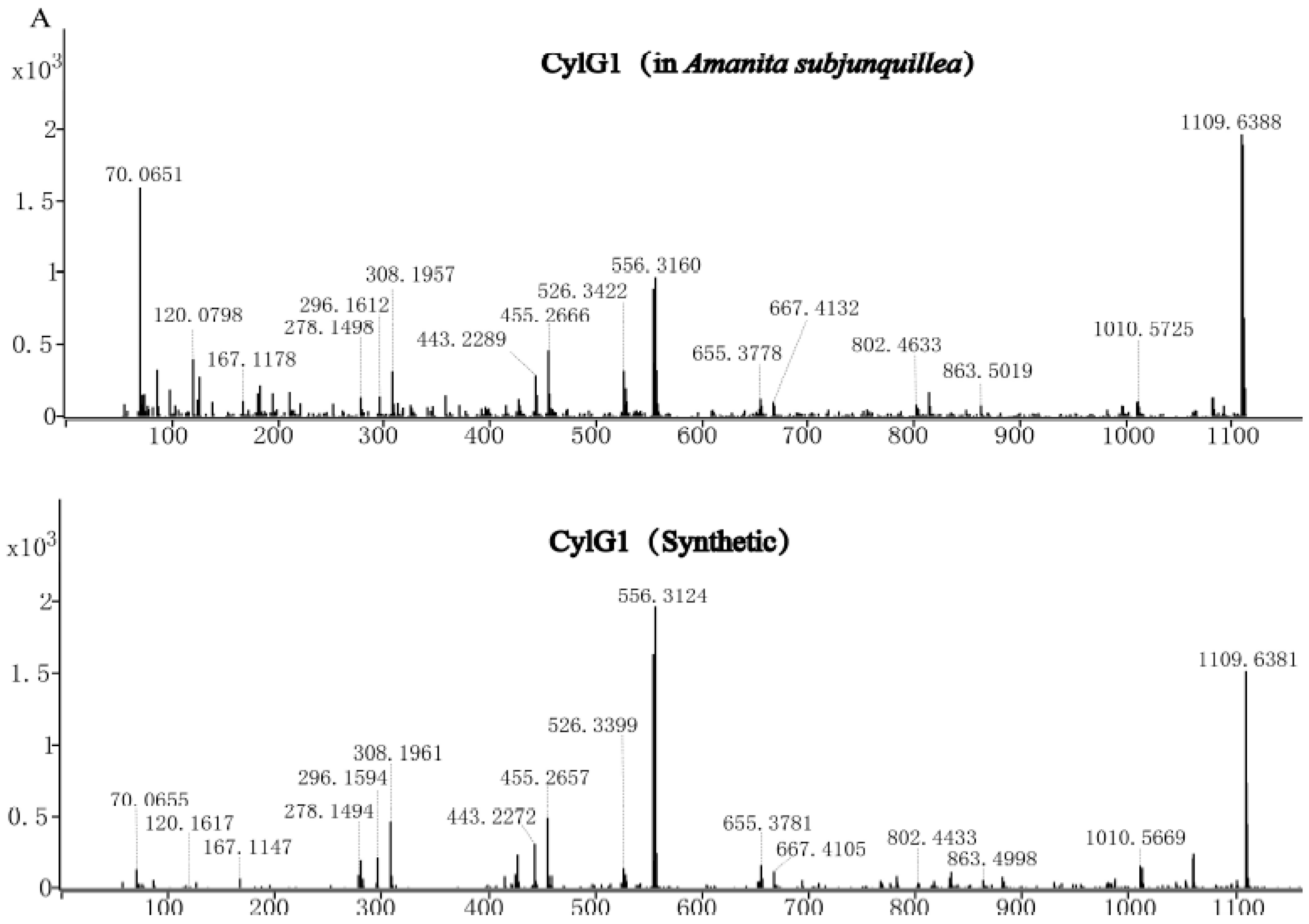
3.6. Confirmation of Three Cyclic Peptides via Synthetic Cyclic Peptides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wieland, T. Peptides of Poisonous Amanita Mushrooms; Springer: Berlin/Heidelberg, Germany, 1986. [Google Scholar]
- Bresinsky, A.; Besl, H. A Colour Atlas of Poisonous Fungi: A Handbook for Pharmacists, Doctors, and Biologists; CRC Press: London, UK, 1990. [Google Scholar]
- Yang, Z.L. Atlas of the Chinese Species of Amanitaceae; Science Press: Beijing, China, 2015. [Google Scholar]
- Chen, Z.H.; Yang, Z.L.; Bau, T.; Li, T.H. Poisonous Mushrooms: Recognition and Poisoning Treatment; Science Press: Beijing, China, 2016. [Google Scholar]
- Cui, Y.Y.; Cai, Q.; Tang, L.P.; Liu, J.W.; Yang, Z.L. The family Amanitaceae: Molecular phylogeny, higher-rank taxonomy and the species in China. Fungal Divers. 2018, 91, 5–230. [Google Scholar] [CrossRef]
- Walton, J.D. The Cyclic Peptide Toxins of Amanita and Other Poisonous Mushrooms; Springer International Publishing AG: Cham, Switzerland, 2018. [Google Scholar]
- Lynen, F.; Wieland, T. Über die Giftstoffe des Knollenblätterpilzes. IV. Liebigs Ann. Chem. 1938, 533, 93–117. [Google Scholar] [CrossRef]
- Wieland, T.; Dudensing, C. Über die Giftstoffe des grünen Knollenblätterpilzes, XI. γ-Amanitin, eine weitere Giftkomponente. Justus Liebigs Ann. Chem. 1941, 600, 156–160. [Google Scholar] [CrossRef]
- Diaz, J.H. Evolving global epidemiology, syndromic classification, general management, and prevention of unknown mushroom poisonings. Crit. Care Med. 2005, 33, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, D.A.; Patrick, C.; Kornberg, R.D. Structural basis of transcription: α-Amanitin-RNA polymerase II cocrystal at 2.8 Å resolution. Proc. Natl. Acad. Sci. USA 2002, 99, 1218–1222. [Google Scholar] [CrossRef] [PubMed]
- Greenleaf, A.L.B.; Leslie, M.; Jiamachello, P.F.; Coulter, D.E. α-Amanitin-resistant D. melanogaster with an altered RNA polymerase II. Cell 1979, 18, 613–622. [Google Scholar] [CrossRef]
- Wieland, T. The Toxic Peptides of Amanita Phalloides; Springer: Vienna, Austria, 1967. [Google Scholar]
- Wieland, T.; Schnabel, H.W. Über die Giftstoffe des grünen Knollenblätterpilzes, XXI: Die Konstitution des Phallacidins. Liebigs Ann. Chem. 1962, 657, 218–225. [Google Scholar] [CrossRef]
- Wieland, T.; Schnabel, H.W. Über die Giftstoffe des grünen Knollenblätterpilzes, XXII: Neue Sequenzanalyse von Phalloidin und Phalloin. Liebigs Ann. Chem. 1962, 657, 225–228. [Google Scholar] [CrossRef]
- Hallen, H.E.; Luo, H.; Scott-Craig, J.S.; Walton, J.D. Gene family encoding the major toxins of lethal Amanita mushrooms. Proc. Natl. Acad. Sci. USA 2007, 104, 19097–19101. [Google Scholar] [CrossRef]
- Luo, H.; Hong, S.Y.; Sgambelluri, R.M.; Angelos, E.; Li, X.; Walton, J.D. Peptide macrocyclization catalyzed by a prolyl oligopeptidase involved in α-amanitin biosynthesis. Chem. Biol. 2014, 21, 1610–1617. [Google Scholar] [CrossRef]
- Sgambelluri, R.M.; Smith, M.O.; Walton, J.D. Versatility of prolyl oligopeptidase B in peptide macrocyclization. ACS Synth. Biol. 2018, 7, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Cai, Q.; Lüli, Y.J.; Li, X.; Sinha, R.; Hallen–Adams, H.E.; Yang, Z.L. The MSDIN family in amanitin-producing mushrooms and evolution of the prolyl oligopeptidase genes. IMA Fungus 2018, 9, 225–242. [Google Scholar] [CrossRef]
- Pulman, J.A.; Childs, K.L.; Sgambelluri, R.M.; Walton, J.D. Expansion and diversification of the MSDIN family of cyclic peptide genes in the poisonous agarics Amanita phalloides and A. bisporigera. BMC Genom. 2016, 17, 1038. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.H.; Wu, P.; Chi, Y.L.; Xu, L.X.; Wei, X.Y. Cyclopeptides from Amanita exitialis. Nat. Prod. Bioprospect. 2011, 1, 52–56. [Google Scholar] [CrossRef]
- Clarke, D.B.; Lloyd, A.S.; Robb, P. Application of liquid chromatography coupled to time-of-flight mass spectrometry separation for rapid assessment of toxins in Amanita mushrooms. Anal. Methods 2012, 4, 1298–1309. [Google Scholar] [CrossRef]
- Letschert, K.; Faulstich, H.; Keller, D.; Keppler, D. Molecular characterization and inhibition of amanitin uptake into human hepatocytes. Toxicol. Sci. 2006, 91, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Meier-Abt, F.; Faulstich, H.; Hagenbuch, B. Identification of phalloidin uptake systems of rat and human liver. Biochim. Biophys. Acta Biomembr. 2004, 1664, 64–69. [Google Scholar] [CrossRef]
- Azzolin, L.; Antolini, N.; Calderan, A.; Ruzza, P.; Sciacovelli, M.; Marin, O.; Mammi, S.; Bernardi, P.; Rasola, A. Antamanide, a derivative of Amanita phalloides, is a novel inhibitor of the mitochondrial permeability transition pore. PLoS ONE 2011, 6, e16280. [Google Scholar] [CrossRef]
- Siemion, I.Z.; Pedyczak, A.; Trojnar, J.; Zimecki, M.; Wieczorek, Z. Immunosuppressive activity of antamanide and some of its analogs. Peptides 1992, 13, 1233–1237. [Google Scholar] [CrossRef]
- Wieczorek, Z.; Siemion, I.Z.; Zimecki, M.; Bolewska-pedyczak, E.; Wieland, T. Immunosuppressive activity in the series of cycloamanide peptides from mushrooms. Peptides 1993, 14, 1–5. [Google Scholar] [CrossRef]
- Thell, K.; Hellinger, R.; Schabbauer, G.; Gruber, C.W. Immunosuppressive peptides and their therapeutic applications. Drug Discov. Today 2014, 19, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Landmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Hallen, H.E.; Watling, R.; Adams, G.C. Taxonomy and toxicity of Conocybe lactea and related species. Mycol. Res. 2003, 107, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Enjalbert, F.; Gallion, C.; Jehl, F.; Monteil, H.; Faulstich, H. Simultaneous assay for amatoxins and phallotoxins in Amanita phalloides Fr. by high-performance liquid chromatography. J. Chromatogr. 1992, 598, 227–236. [Google Scholar] [CrossRef]
- Luo, H.; Hallen-Adams, H.E.; Scott-Craig, J.S.; Walton, J.D. Ribosomal biosynthesis of α-amanitin in Galerina marginata. Fungal Genet. Biol. 2012, 49, 123–129. [Google Scholar] [CrossRef] [PubMed]
- May, J.P.; Perrin, D.M. Tryptathionine bridges in peptide synthesis. Biopolymers 2007, 88, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Pevzner, P.A. MS-GF+ makes progress towards a universal database search tool for proteomics. Nat. Commun. 2014, 5, 5277. [Google Scholar] [CrossRef]
- Yang, H.; Chi, H.; Zhou, W.J.; Zeng, W.F.; He, K.; Liu, C.; Sun, R.X.; He, S.M. Open-pNovo: De novo peptide sequencing with thousands of protein modifications. J. Proteome Res. 2017, 16, 645–654. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef]
- Li, H.J.; Zhang, H.S.; Zhang, Y.Z.; Zhang, K.P.; Sun, C.Y. Mushroom poisoning outbreaks—China, 2019. China CDC Wkly. 2020, 2, 19–24. [Google Scholar] [CrossRef]
- Li, P.; Deng, W.Q.; Li, T.H.; Song, B.; Shen, Y.H. Illumina-based de novo transcriptome sequencing and analysis of Amanita exitialis basidiocarps. Gene 2013, 532, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Deng, W.Q.; Li, T.H. The molecular diversity of toxin gene families in lethal Amanita mushrooms. Toxicon 2014, 83, 59–68. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Han, C.; Wan, G.; Huang, X.; Ivan, C.; Jiang, D.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Rao, P.H.; et al. TP53 loss creates therapeutic vulnerability in colorectal cancer. Nature 2015, 520, 697–701. [Google Scholar] [CrossRef]
- Kume, K.; Ikeda, M.; Miura, S.; Ito, K.; Sato, K.A.; Ohmori, Y.; Endo, F.; Katagiri, H.; Ishida, K.; Ito, C.; et al. α-Amanitin restrains cancer relapse from drug-tolerant cell subpopulations via TAF15. Sci. Rep. 2016, 6, 25895. [Google Scholar] [CrossRef] [PubMed]
- Craig, E.W.; Avasthi, P. Visualizing filamentous actin using phalloidin in chlamydomonas reinhardtii. Bio Protocol. 2019, 9, e3274. [Google Scholar] [CrossRef] [PubMed]
- Raymond, G.; Potreau, D.; Cognard, C.; Jahn, W.; Wieland, T. Antamanide antagonizes the phalloidin-induced negative inotropic effect and blocks voltage dependently the fast outward K+ current in voltage-clamped frog muscle fibres. Eur. J. Pharmacol. 1987, 138, 21–27. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leader Peptide | Core Peptide | Recognition Sequence | Monoisotopic Mass |
|---|---|---|---|
| MSDINSTRLP | IWGIGCNP | SVGDEVTALLTRGEA | 918.3541 (α-amanitin) |
| MSDINATRLP | IWGIGCNP | SVGDEVTALLASGEA | 918.3541 (α-amanitin) |
| MSDINATRLP | IWGIGCDP | CVGDDVAALTTRGEA | 919.3382 (β-amanitin) |
| MSDINATRVP | AWLVDCP | CVGDDISRLLTRGEK | 846.3217 (phallacidin) |
| MSDINATRLP | AWDSKHP | CVGDDVSRLLTRGE | 821.3820 |
| MSDINATRLP | AWDSKHP | CVGDDISRLLTRGE | 821.3820 |
| MSDINATRVP | AWLAECP | CVGDDISHLLTRGE | 770.3421 |
| MSDINASRLP | FFIIIVKP | CGNPYVSDDVNSTLTRGE | 957.6052 |
| MSDINTSRLP | FIPLGIITILP ★ | CVSDDVNTTITRGD | 1177.7475 |
| MSDINTACLP | FLFPVIPP | CLSEDANVVVLNSGE | 910.5317 |
| MSDINVTRLP | FFPIVFIPP | CI | 1057.6000 |
| MSDINIARLP | IFWFIYFP | CVGDDVDNTLSRGE | 1113.5688 |
| MSDINVTRLP | IFLIMFIPP | CIGDDAASILKQGE | 1071.6191 |
| MSDINTSCLP | IFIAFPIPP | CVSDDIQTVLTRGE | 995.5844 |
| MSDTNTACLP | IFIAFPIPP | CVSDDIQTVLTRGE | 995.5844 |
| MSDINASRLP | ILKKPWAP | SVCDDVNSTLTRGE | 933.5800 |
| MSDINVARLP | ISDPTAYP ★ | CVGDDIQAVVKRGE | 844.3967 |
| MSDINATRLP | IIIVLGLIIP | LCVSDIEMILTRGE | 1044.7311 |
| MSDINASRLP | IILAPIIP | CISDDVNTTLTCAE | 830.5630 |
| MSDINTTGLP | HFYNLMPP | CFSDDTGMVLVRGE | 999.4637 |
| MSDINATRLP | HPFPLGLQP | CAGDVDNFTLIKGE | 986.5338 |
| MSDINASCLP | LILVANGMAYV | --SDDVSPTLTRGE | 1144.6315 |
| MSDINTARLP | SYIPFPPP | CLSEDTNAVLMLGE | 898.4589 |
| MSDINTARLP | SYIPFPPP | CLSEDTNAVLMLGE | 898.4589 |
| MSDINTSRFP | SYGYRAFP | CVGDDVEMVLMHGE | 941.4396 |
| MSDINVTRLP | VLVFIFFLP | CISDDAASIIKLGE | 1075.6470 |
| MSDIDTTRLP | LILFTLQP | SIGDDVNPTLTRGEK | 925.5637 |
| MSDIHAARLP | FPTRPVFP ★ | SAGDDMIEVVLGRGE | 941.5123 |
| MSDNNAARLP | FYFYLGIP | SDDAHPILTRGERLA | 1000.5058 |
| MSDTNTARLP | ILFIQLEIP | CISDDVHPVLTRGE | 1066.6427 |
| MSDVNTTRLP | FNFFRFPYP | CICDDSEKVLELGE | 1215.5866 |
| MSEINTARFP | NHGHRTIP | CVGDDIEMVLMHGE | 912.4678 |
| MSEINTSRLP | LVFIPPYFAP | CVSDDIQMVLTLGE | 1144.6321 |
| MFDMNTTCLP | GFIIYAYV | --GDDVNHTLTRGE | 926.4902 |
| MLDINTARLP | FSLPTFPP | CVSDEIDVVLKRGE | 886.4589 |
| MLDINATRFP | LGRPTHLP | CVGDDVNYIL | 871.5028 |
| MTDINDARLP | ILLLIFFWIP | CANDDDENILNRG | 1255.7733 |
| MTDINDTRLP | FVWILWLWLA | CVGDDTSILNRGE | 1327.7481 |
| MPDINVTRLP | LLIIVLLTP | CISDDNNILNRGK | 975.6732 |
| Leader Peptide | Core Peptide | Recognition Sequence | Monoisotopic Mass |
|---|---|---|---|
| MSDINATRLP | IWGIGCNP | CVGDDVTSVLTRGEA | 918.3541(α-amanitin) |
| MSDINATRLP | IWGIGCDP | CVGDDVTALLTRGEA | 919.3382(β-amanitin) |
| MSDINATRLP | AWLVDCP | CVGDDVNRLLTRGE | 846.3217(phallacidin) |
| MSDINATRLP | AWLTDCP | CVGDDVNRLLTRGE | 786.3371 |
| MSDINTTRLP | FVFVASPP ★ | CVGDDIAMVLTRGE | 844.4483 |
| MSDINTARLP | FIWVFGIP | --GDDIGTVLTRGEK | 959.5269 |
| MSDINLTRLP | GIIAIIP | CVGDDDDVNSTLTRGQ | 677.4476 |
| MSDINATRLP | IILAPVIP | CISDDNDP--TLTRGQ | 816.5473 |
| MSDINTARLP | IPIPPFFFP | FVSDDIEIVLRRGEK | 1055.5844 |
| MSDINTARLP | IPIPPFFFP | FVSDDIEIVLRRGEK | 1055.5844 |
| MSDINATRLP | IGRPQLLP | CVGGDVNYILISGEK | 874.5389 |
| MSDINPTRLP | IFWFIYFP | CVSDVDST-LTRGE | 1113.5688 |
| MSDINTARLP | IYRPPFYALP | CVGDDIQAVLTRGE | 1217.6597 |
| MSDINTARLP | IIWIIGNP | CVSDDVERILTRGE | 906.5327 |
| MSDINVIRAP | LLILSILP | CVGDDIEV-LRRGE | 862.5892 |
| MSDINATRLP | LFFPPDFRPP ★ | CVGDADNFTLTRGEK | 1213.6284 |
| MSDINATRLP | LFFPPDFRPP ★ | CVGDADNFTLTRGE | 1213.6284 |
| MSDINVIRLP | SMLTILPP | CVSDDASNTLTRGE | 852.4779 |
| MSDINTARLP | VFSLPVFFP ★ | --SDDIQAVLTRGE | 1033.5637 |
| MSDINVTRLP | VFIFFFIPP | CVGDGTADIVRKGEK | 1107.6157 |
| MSDINATRLP | VWIGYSP | CVGDDCIALLTRGE | 802.4014 |
| MSDINATRLP | VWIGYSP | CVGDDCIALLTRGE | 802.4014 |
| MTDINDTRLP | FIWLLWIWLP | SVGDD-NNILNRGEE | 1367.7794 |
| Leader Peptide | Core Peptide | Recognition Sequence | Monoisotopic Mass | Expressed |
|---|---|---|---|---|
| MSDINATCLP | IWGIGCNP | CVGDEVAALLTRGEALC | 918.3541(α-amanitin) | √ |
| MSDINATRLP | IWGIGCDP | CVGDEVTALLTRGEALC | 919.3382 (β-amanitin) | √ |
| MSDINATRLP | IWGIGCDP | CIGDDVTALLTRGEALC | 919.3382 (β-amanitin) | √ |
| MSDINATRLP | AWLATCP | CAGDDVNPTLTRGESLC | 788.3160 (phalloidin) | √ |
| MSDINATRLP | AWLVDCP | CVGDDINRRVVSAFA-C | 846.3217 (phallacidin) | √ |
| MSDMNATRLP | LIQRPFAP | CVSDDVDFALIRRCALVYAESSV | 922.5389 | √ |
| MSDINTARLP | HFASFIPP | CIGDDIEMVLKRGESLC | 896.4545 | √ |
| MSDINTARLP | TFLPPLFVPP ★ | CVSDDIEMVLTRGESLC | 1108.6321 | √ |
| MSDINATRLP | LNILPFMLPP | CVGDDVNPTLTRGEDLC | 1135.6464 | √ |
| MSDMNATRLP | LIQRPYAP | CVSDDVNSPLTRGESLC | 938.5338 | √ |
| MSDINTARLP | IGRPESIP | CVGDDIEMILERGQKLC | 849.4709 | √ |
| MSDINTARLP | LRLPPFMIPP | CVGDDIGMVLTRGENLC | 1161.6733 | √ |
| MSDVNATRLP | FNFFRFPYP | CIGDDSASVLGLGESLC | 1215.5866 | √ |
| MSDINATRLP | SSVLPRP | CVGDVDNIILTSREKLC | 736.4232 | √ |
| MSDINTARLP | AFFPPFFIPP ★ | CVSDDIEMVLTRGESLC | 1160.6059 | √ |
| MSDINATRLP | IPILPIPP | YCSDDANTTLTLGESLC | 840.5473 | √ |
| MSDINATRLP | LFLLAALGIP | --SDDADSTLTRGESLC | 1008.6372 | √ |
| MSDTNDARLP | LFFWFWFLWP | SVSDDIDSVLNRGEDLC | 1469.7325 | √ |
| MSDMNVARLP | ISDPTAYP | CVGGDIHAVLRRGE | 844.3966 | × |
| MSDMNVARLP | ISDPTAYP | CVGGDIHAVLRRGE | 844.3966 | × |
| MSDINVTCLP | FIFWFFWPP | CVGDDAASIIK-GK | 1267.6218 | × |
| MSDINAARLP | FIFPPFFIPP | CVSDDIEMVLTRGE | 1202.6528 | × |
| MSDINTVCLP | LQKPWSRP | CVGDDIEMILERGE | 992.5556 | × |
| MFDINITRLP | IFWFIYFP | CVGDDVTALLTRGE | 1113.5689 | × |
| Species | Cyclopeptide Sequence | Molecular Formula | Theoretical (m/z) | Measured (m/z) | δ (ppm) |
|---|---|---|---|---|---|
| A. subjunquillea | TFLPPLFVPP (CylG1) | C59H84N10O11 | 1109.6394 | 1109.6398 | 0.36 |
| AFFPPFFIPP (CylG2) | C65H80N10O10 | 1161.6132 | 1161.6161 | 2.50 | |
| A. rimosa | ISDPTAYP * (CylH1) | C39H56N8O13 | 845.4039 | 845.4040 | 0.12 |
| FIPLGIITILP (CylH2) | C61H99N11O12 | 1178.7547 | 1178.7555 | 0.68 | |
| FPTRPVFP (CylH3) | C48H67N11O9 | 942.5196 | 942.5191 | 0.53 | |
| A. pallidorosea | EFIVFGIFP (CylI1) | C56H75N9O11 | 1050.5658 | 1050.5694 | 3.43 |
| FVIIPPFIFP (CylI2) | C65H90N10O10 | 1171.6914 | 1171.6941 | 2.31 | |
| YFFNDHPP (CylI3) | C51H59N11O12 | 1018.4417 | 1018.4421 | 0.39 | |
| TIHLFSAP (CylI4) | C42H62N10O10 | 867.4723 | 867.4733 | 1.15 | |
| MHILAPPP (CylI5) | C41H64N10O8S | 857.4702 | 857.4714 | 1.40 | |
| A. exitialis | FVFVASPP (CylJ1) | C44H60N8O9 | 845.4556 | 845.4582 | 3.08 |
| LFFPPDFRPP # (CylJ2) | C63H83N13O12 | 1214.6357 | 1214.6357 | 0.00 | |
| VFSLPVFFP ^ | C56H75N9O10 | 1034.5709 | 1034.5734 | 2.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Li, X.; Lüli, Y.; Li, X.; Chen, Z.H.; Yuan, P.; Yang, Z.L.; Li, G.; Luo, H. Novel Cyclic Peptides from Lethal Amanita Mushrooms through a Genome-Guided Approach. J. Fungi 2021, 7, 204. https://doi.org/10.3390/jof7030204
Zhou S, Li X, Lüli Y, Li X, Chen ZH, Yuan P, Yang ZL, Li G, Luo H. Novel Cyclic Peptides from Lethal Amanita Mushrooms through a Genome-Guided Approach. Journal of Fungi. 2021; 7(3):204. https://doi.org/10.3390/jof7030204
Chicago/Turabian StyleZhou, Shengwen, Xincan Li, Yunjiao Lüli, Xuan Li, Zuo H. Chen, Pengcheng Yuan, Zhu L. Yang, Guohong Li, and Hong Luo. 2021. "Novel Cyclic Peptides from Lethal Amanita Mushrooms through a Genome-Guided Approach" Journal of Fungi 7, no. 3: 204. https://doi.org/10.3390/jof7030204
APA StyleZhou, S., Li, X., Lüli, Y., Li, X., Chen, Z. H., Yuan, P., Yang, Z. L., Li, G., & Luo, H. (2021). Novel Cyclic Peptides from Lethal Amanita Mushrooms through a Genome-Guided Approach. Journal of Fungi, 7(3), 204. https://doi.org/10.3390/jof7030204