
Inocybe brijunica sp. nov., a New Ectomycorrhizal Fungus from Mediterranean Croatia Revealed by Morphology and Multilocus Phylogenetic Analysis

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Research Area
2.2. Morphological Study
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Sequence Alignment and Phylogenetic Analysis
3. Results
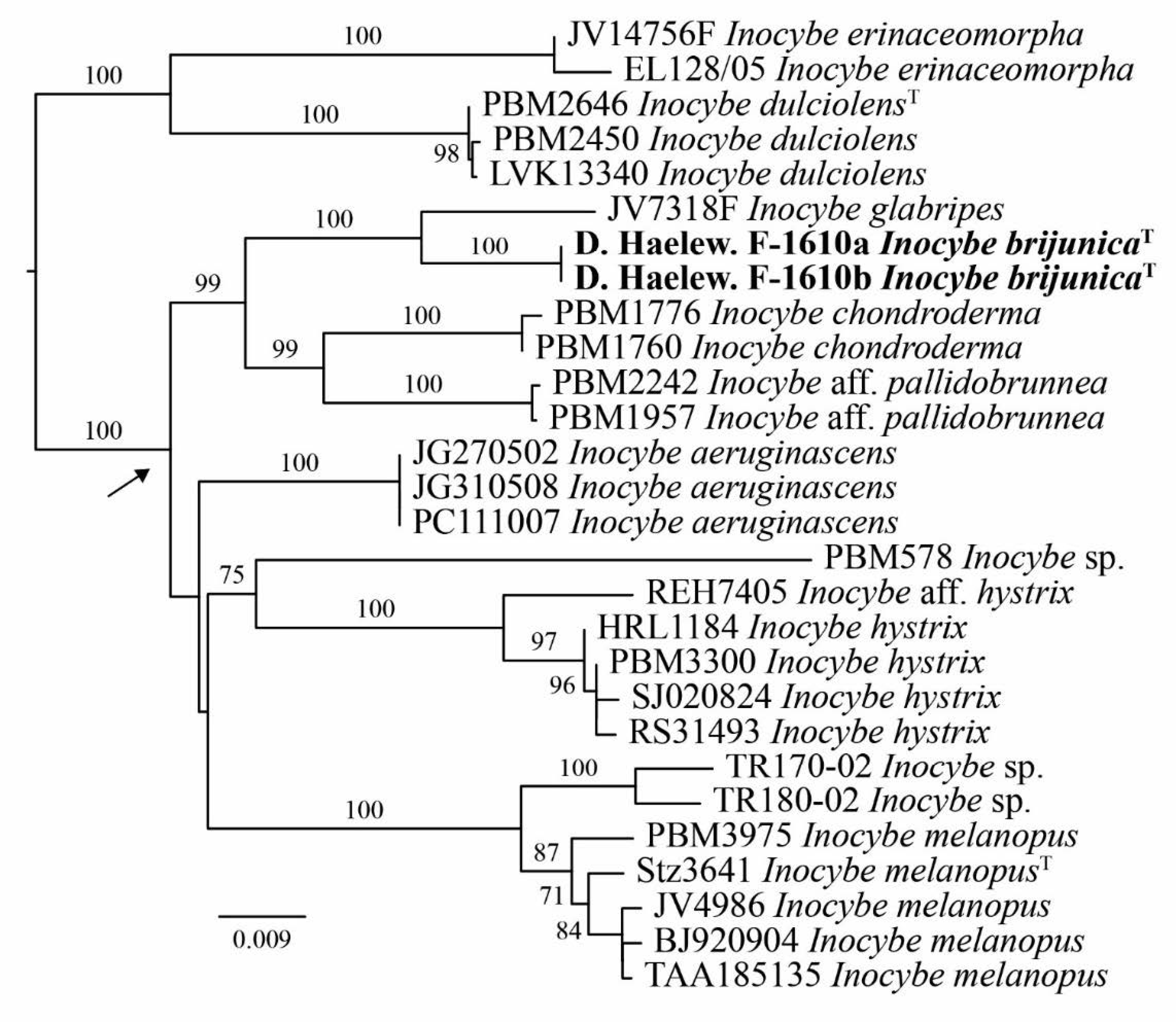
3.1. Phylogenetic Inference
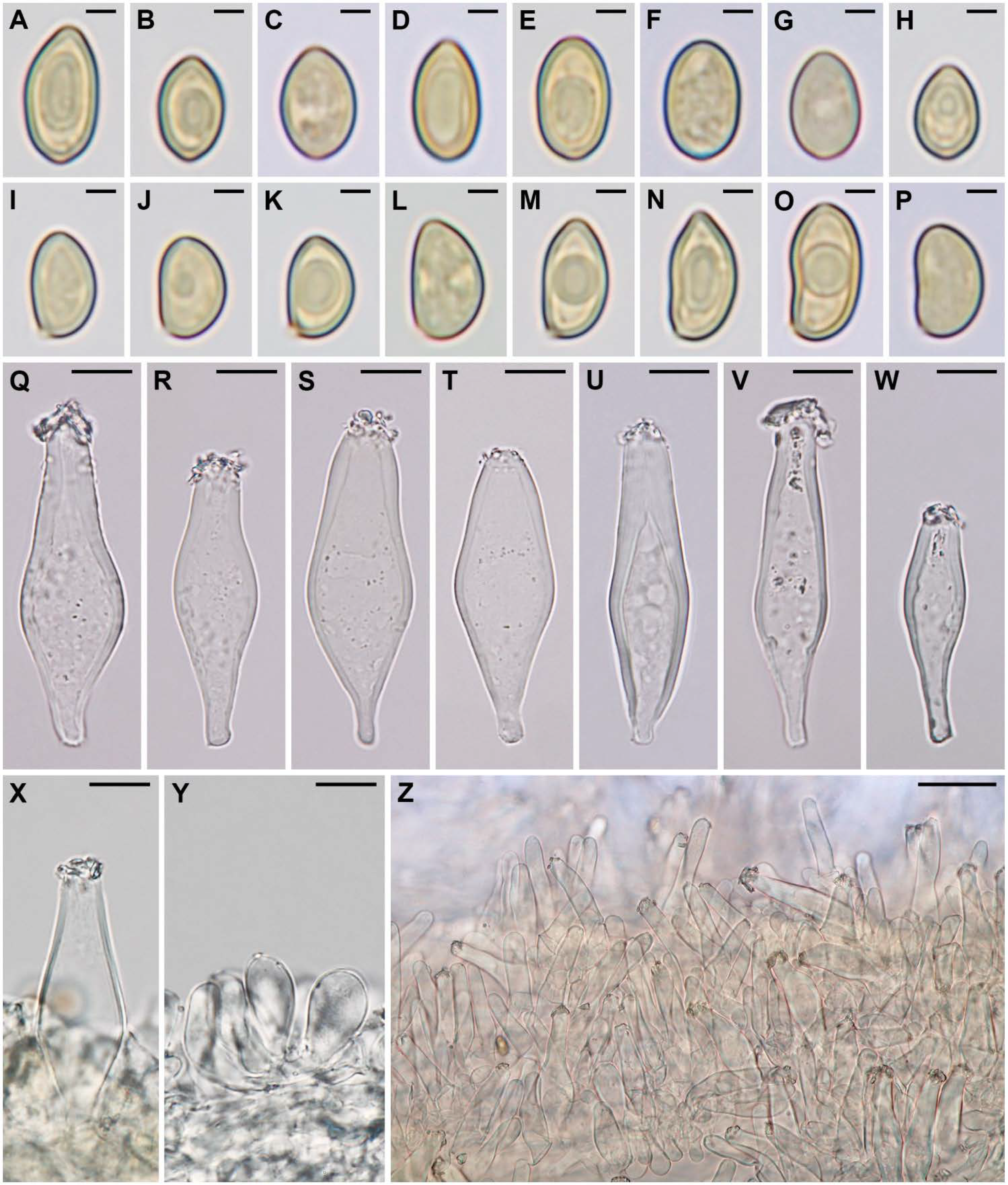
3.2. Taxonomy
Inocybe brijunica Mešić, Tkalčec & Haelew., sp. nov.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brijuni National Park Official Website. Available online: https://www.np-brijuni.hr/en/brijuni (accessed on 14 January 2021).
- Blondel, J.; Aronson, J.; Bodiou, J.-Y.; Boeuf, G. The Mediterranean Region—Biological Diversity in Space and Time, 2nd ed.; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, 1–4. [Google Scholar] [CrossRef]
- Tuel, A.; Eltahir, E.A.B. Why is the Mediterranean a climate change hot spot? J. Clim. 2020, 33, 5829–5843. [Google Scholar] [CrossRef]
- Mariotti, A.; Zeng, N.; Yoon, J.-H.; Artale, V.; Navarra, A.; Alpert, P.; Li, L.Z.X. Mediterranean water cycle changes: Transition to drier 21st century conditions in observations and CMIP3 simulations. Environ. Res. Lett. 2008, 3, 044001. [Google Scholar] [CrossRef]
- Brogli, R.; Sørland, S.L.; Kröner, N.; Schär, C. Causes of future Mediterranean precipitation decline depend on the season. Environ. Res. Lett. 2019, 14, 114017. [Google Scholar] [CrossRef]
- Antonelli, A.; Fry, C.; Smith, R.J.; Simmonds, M.S.J.; Kersey, P.J.; Pritchard, H.W.; Abbo, M.S.; Acedo, C.; Adams, J.; Ainsworth, A.M.; et al. State of the World’s Plants and Fungi 2020; Royal Botanic Gardens: Kew, UK, 2020. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Cheek, M.; Nic Lughadha, E.; Kirk, P.; Lindon, H.; Carretero, J.; Looney, B.; Douglas, B.; Haelewaters, D.; Gaya, E.; Llewellyn, T.; et al. New scientific discoveries: Plants and fungi. Plants People Planet 2020, 2, 371–388. [Google Scholar] [CrossRef]
- Saba, M.; Haelewaters, D.; Pfister, D.H.; Khalid, A.N. New species of Pseudosperma (Agaricales, Inocybaceae) from Pakistan revealed by morphology and multi-locus phylogenetic reconstruction. MycoKeys 2020, 69, 1–31. [Google Scholar] [CrossRef] [PubMed]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef]
- Bandini, D.; Oertel, B.; Ploch, S.; Ali, T.; Vauras, J.; Schneider, A.; Scholler, M.; Eberhardt, U.; Thines, M. Revision of some central European species of Inocybe (Fr.: Fr.) Fr. subgenus Inocybe, with the description of five new species. Mycol. Prog. 2018, 18, 247–294. [Google Scholar] [CrossRef]
- Bandini, D.; Oertel, B.; Moreau, P.-A.; Thines, M.; Ploch, S. Three new hygrophilous species of Inocybe, subgenus Inocybe. Mycol. Prog. 2019, 18, 1101–1119. [Google Scholar] [CrossRef]
- Bandini, D.; Oertel, B.; Ploch, S.; Thines, M. Inocybe heidelbergensis, eine neue Risspilz-Art der Untergattung Inocybe. Z. Mykol. 2019, 85, 195–213. [Google Scholar]
- Bandini, D.; Oertel, B.; Schüssler, C.; Eberhardt, U. Noch mehr Risspilze: Fünzehn neue und zwei wenig bekannte Arten der Gattung Inocybe. Mycol. Bavarica 2020, 20, 13–101. [Google Scholar]
- Bandini, D.; Sesli, E.; Oertel, B.; Krisai-Greilhuber, I. Inocybe antoniniana, a new species of Inocybe section Marginatae with nodulose spores. Sydowia 2020, 72, 95–106. [Google Scholar] [CrossRef]
- Bandini, D.; Vauras, J.; Weholt, Ø.; Oertel, B.; Eberhardt, U. Inocybe woglindeana, a new species of the genus Inocybe, thriving in exposed habitats with calcareous sandy soil. Karstenia 2020, 58, 41–59. [Google Scholar] [CrossRef]
- Cripps, C.L.; Larsson, E.; Vauras, J. Nodulose-spored Inocybe from the Rocky Mountain alpine zone molecularly linked to European and type specimens. Mycologia 2019, 112, 133–153. [Google Scholar] [CrossRef]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef]
- Krieglsteiner, L.G. Inocybe calosporoides—Ein neuer Risspilz aus Portugal. Südwestdeutsche Pilzrundsch. 2019, 55, 68–72. [Google Scholar]
- Krieglsteiner, L.G. Nomenclatural novelties. Index Fungorum 2019, 411, 1. [Google Scholar]
- Matheny, P.B.; Hobbs, A.M.; Esteve-Raventós, F. Genera of Inocybaceae: New skin for the old ceremony. Mycologia 2019, 112, 83–120. [Google Scholar] [CrossRef] [PubMed]
- Clémençon, H. Cytology and Plectology of the Hymenomycetes, 2nd ed.; Cramer: Stuttgart, Germany, 2012. [Google Scholar]
- Erb, B.; Matheis, W. Pilzmikroskopie; Kosmos: Stuttgart, Germany, 1982. [Google Scholar]
- Matheny, P.B.; Kudzma, L.V. New species of Inocybe (Inocybaceae) from eastern North America. J. Torrey Bot. Soc. 2019, 146, 213–235. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Egger, K.N. Molecular analysis of ectomycorrhizal fungal communities. Can. J. Bot. 1995, 73, S1415–S1422. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Hopple, J.S. Phylogenetic Investigations in the Genus Coprinus Based on Morphological and Molecular Characters. Ph.D. Thesis, Duke University, Durham, NC, USA, 1994. [Google Scholar]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe, Agaricales). Mol. Phylogenet. Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef]
- Haelewaters, D.; Toome-Heller, M.; Albu, S.; Aime, M.C. Red yeasts from leaf surfaces and other habitats: Three new species and a new combination of Symmetrospora (Pucciniomycotina, Cystobasidiomycetes). Fungal Syst. Evol. 2020, 5, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Matheny, P.B.; Norvell, L.L.; Giles, E.C. A common new species of Inocybe in the Pacific Northwest with a diagnostic PDAB reaction. Mycologia 2013, 105, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Ryberg, M.; Matheny, P.B. Asynchronous origins of ectomycorrhizal clades of Agaricales. Proc. R. Soc. B Biol. Sci. 2012, 279, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Ryberg, M.; Nilsson, R.H.; Kristiansson, E.; Töpel, M.; Jacobsson, S.; Larsson, E. Mining metadata from unidentified ITS sequences in GenBank: A case study in Inocybe (Basidiomycota). BMC Evol. Biol. 2008, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, Louisiana, 14 November 2010; Institute of Electrical and Electronics Engineers: Piscataway, NJ, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Dentinger, B.T.; Didukh, M.Y.; Moncalvo, J.M. Comparing COI and ITS as DNA barcode markers for mushrooms and allies (Agaricomycotina). PLoS ONE 2011, 6, e25081. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef]
- Haelewaters, D.; Dirks, A.C.; Kappler, L.A.; Mitchell, J.K.; Quijada, L.; Vandegrift, R.; Buyck, B.; Pfister, D.H. A preliminary checklist of fungi at the Boston Harbor islands. Northeast. Nat. 2018, 25, 45–76. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Wurzbacher, C.; Larsson, E.; Bengtsson-Palme, J.; Van den Wyngaert, S.; Svantesson, S.; Kristiansson, E.; Kagami, M.; Nilsson, R.H. Introducing ribosomal tandem repeat barcoding for fungi. Mol. Ecol. Resour. 2018, 19, 118–127. [Google Scholar] [CrossRef]
- Stangl, J.; Veselský, J. Risspilze der Section Lilacinae Heim. Česká Mykol. 1982, 36, 85–99. [Google Scholar]
- Kuyper, T.W. A revision of the genus Inocybe in Europe 1. Subgenus Inosperma and the smooth-spored species of subgenus Inocybe. Persoonia 1986, 3, 1–247. [Google Scholar]
- Stangl, J. Die Gattung Inocybe in Bayern. Hoppea 1989, 46, 1–409. [Google Scholar]
- Ludwig, E. Pilzkompendium. Band 4; Fungicon Verlag: Berlin, Germany, 2017. [Google Scholar]
- Ferrari, E.; Bandini, D.; Boccardo, F. Inocybe (Fr.) Fr., Terzo Contributo; Fungi non delineati 73/74; Edizioni Candusso: Alassio, Italy, 2014. [Google Scholar]
- Alessio, C.L. Complemento allo studio del Genere Inocybe: 8° contributo. Riv. Micol. Assoc. Micol. Bresadola 1987, 30, 79–89. [Google Scholar]
- Esteve-Raventós, F. Two new species of Inocybe (Cortinariales) from Spain, with a comparative type study of some related taxa. Mycol. Res. 2001, 105, 1137–1143. [Google Scholar] [CrossRef]
- Smith, A.H.; Stuntz, D.E. New or noteworthy fungi from Mount Rainier National Park. Mycologia 1950, 42, 80–134. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Section | Isolate | Locality | ITS | nrLSU | rpb2 |
|---|---|---|---|---|---|---|
| Inocybe aeruginascens | Hysterices | JG270502 | Germany | GU949590 | JN974970 | |
| Inocybe aeruginascens | Hysterices | JG310508 | Germany | GU949591 | MH220256 | MH249787 |
| Inocybe aeruginascens | Hysterices | PC111007 | South Africa | GU949592 | MH220257 | |
| Inocybe chondroderma | Hysterices | PBM1760 | British Columbia | GU949586 | MH220258 | |
| Inocybe chondroderma | Hysterices | PBM1776 | Washington | GU949579 | JN974967 | MH249789 |
| Inocybe brijunicaT | Hysterices | D. Haelew. F-1610a | Croatia | MN749370 | MN749492 | MT878448 |
| Inocybe brijunicaT | Hysterices | D. Haelew. F-1610b | Croatia | MN749371 | MN749493 | MT878449 |
| Inocybe dulciolensT | Lactiferae | PBM2646 | Tennessee | MH216088 | MH220265 | MH249796 |
| Inocybe dulciolens | Lactiferae | PBM2450 | New York | MH216087 | MH220264 | MH249795 |
| Inocybe dulciolens | Lactiferae | LVK13340 | New Jersey | MH216084 | MH220261 | MH249792 |
| Inocybe erinaceomorpha | Lactiferae | EL128/05 | Sweden | AM882735 | AM882735 | |
| Inocybe erinaceomorpha | Lactiferae | JV14756F | Sweden | MH216089 | MH220266 | MH249797 |
| Inocybe glabripes | Hysterices | JV7318F | Finland | MH216096 | MH249803 | |
| Inocybe hystrix | Hysterices | HRL11842 | Quebec | KX897428 | ||
| Inocybe hystrix | Hysterices | PBM3300 | North Carolina | GU949588 | MH220275 | |
| Inocybe hystrix | Hysterices | RS31493 | Finland | AY380380 | AY337381 | |
| Inocybe hystrix | Hysterices | SJ020824 | Sweden | AM882810 | AM882810 | |
| Inocybe aff. hystrix | Hysterices | REH7405 | Costa Rica | GU949589 | JN974969 | MH249806 |
| Inocybe melanopusT | Hysterices | Stz3641 | Washington | HQ201359 | ||
| Inocybe melanopus | Hysterices | BJ920904 | Sweden | AM882725 | AM882725 | |
| Inocybe melanopus | Hysterices | JV4986 | Finland | AM882727 | AM882727 | |
| Inocybe melanopus | Hysterices | PBM3975 | Tennessee | MH220276 | MH249807 | |
| Inocybe melanopus | Hysterices | TAA185135 | Estonia | AM882726 | ||
| Inocybe aff. pallidobrunnea | Hysterices | PBM1957 | Washington | MH216098 | MH220277 | MH249808 |
| Inocybe aff. pallidobrunnea | Hysterices | PBM2242 | Washington | MH216099 | JN974968 | MH249809 |
| Inocybe sp. | Hysterices | PBM578 | Washington | MH216104 | JN974961 | MH249813 |
| Inocybe sp. | Hysterices | TR170-02 | New Guinea | JN974964 | MH249814 | |
| Inocybe sp. | Hysterices | TR180-02 | New Guinea | JN974965 |
| Locus | Sequences | Sites | Informative | Constant | Model | -lnL |
|---|---|---|---|---|---|---|
| ITS1 | 23 | 246 | 92 | 134 | HKY + F + G4 | 1306.690 |
| 5.8S | 23 | 158 | 4 | 153 | TIM3e | 249.420 |
| ITS2 | 23 | 203 | 79 | 113 | TPM3u + F + G4 | 1034.365 |
| nrLSU | 25 | 1379 | 94 | 1257 | TN + F+I | 3068.786 |
| rpb2 | 16 | 766 | 156 | 540 | TN + F+I | 2935.101 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mešić, A.; Haelewaters, D.; Tkalčec, Z.; Liu, J.; Kušan, I.; Aime, M.C.; Pošta, A. Inocybe brijunica sp. nov., a New Ectomycorrhizal Fungus from Mediterranean Croatia Revealed by Morphology and Multilocus Phylogenetic Analysis. J. Fungi 2021, 7, 199. https://doi.org/10.3390/jof7030199
Mešić A, Haelewaters D, Tkalčec Z, Liu J, Kušan I, Aime MC, Pošta A. Inocybe brijunica sp. nov., a New Ectomycorrhizal Fungus from Mediterranean Croatia Revealed by Morphology and Multilocus Phylogenetic Analysis. Journal of Fungi. 2021; 7(3):199. https://doi.org/10.3390/jof7030199
Chicago/Turabian StyleMešić, Armin, Danny Haelewaters, Zdenko Tkalčec, Jingyu Liu, Ivana Kušan, M. Catherine Aime, and Ana Pošta. 2021. "Inocybe brijunica sp. nov., a New Ectomycorrhizal Fungus from Mediterranean Croatia Revealed by Morphology and Multilocus Phylogenetic Analysis" Journal of Fungi 7, no. 3: 199. https://doi.org/10.3390/jof7030199
APA StyleMešić, A., Haelewaters, D., Tkalčec, Z., Liu, J., Kušan, I., Aime, M. C., & Pošta, A. (2021). Inocybe brijunica sp. nov., a New Ectomycorrhizal Fungus from Mediterranean Croatia Revealed by Morphology and Multilocus Phylogenetic Analysis. Journal of Fungi, 7(3), 199. https://doi.org/10.3390/jof7030199