
Sterol Composition Modulates the Response of Saccharomyces cerevisiae to Iron Deficiency



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Plasmids, and Growth Conditions
2.2. RNA Analyses
2.3. Determination of Total Fatty Acids and Sterols
2.4. β-Galactosidase Assays
2.5. Fluorescence Microscopy
2.6. Principal Component Analysis and Statistical Analyses
3. Results
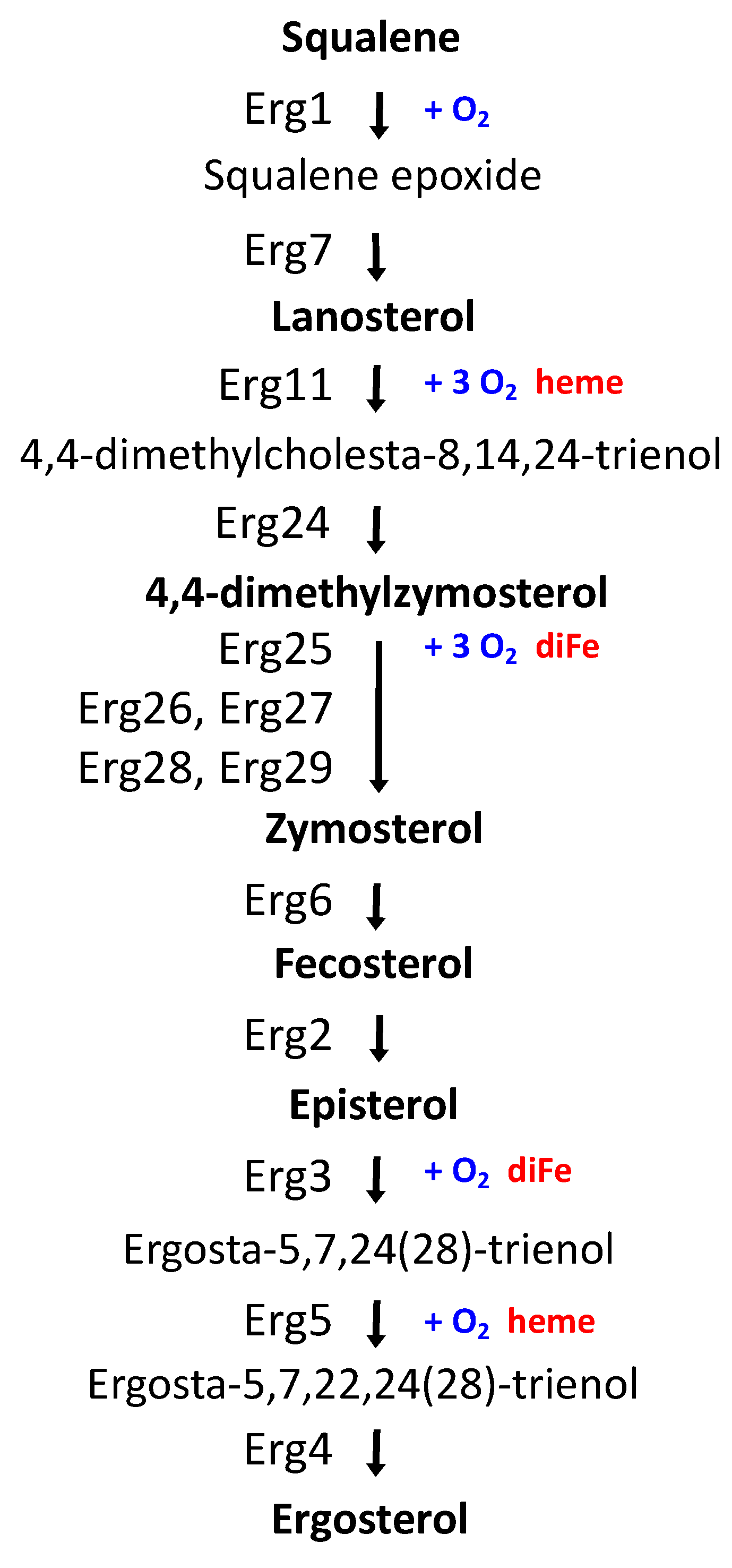
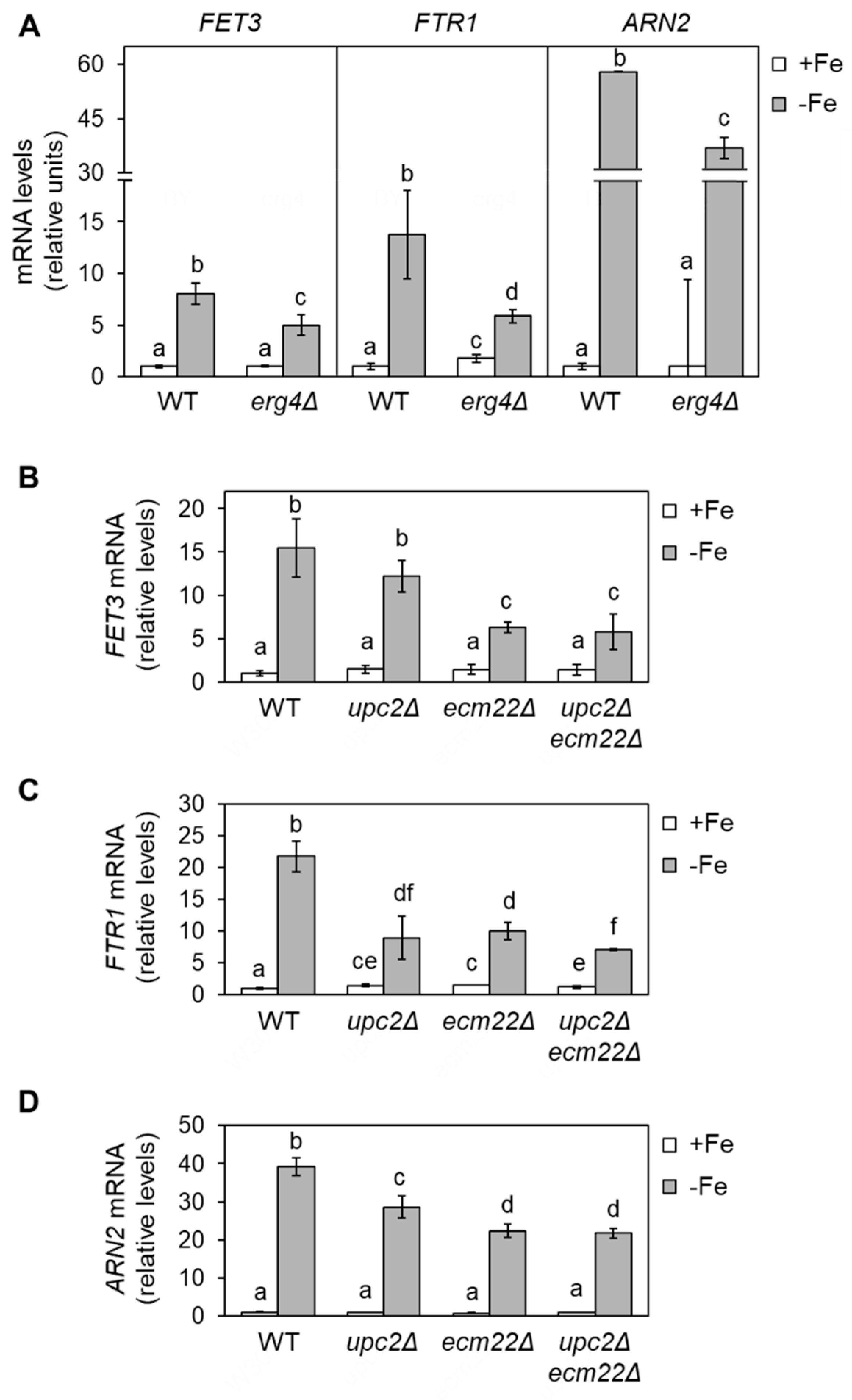
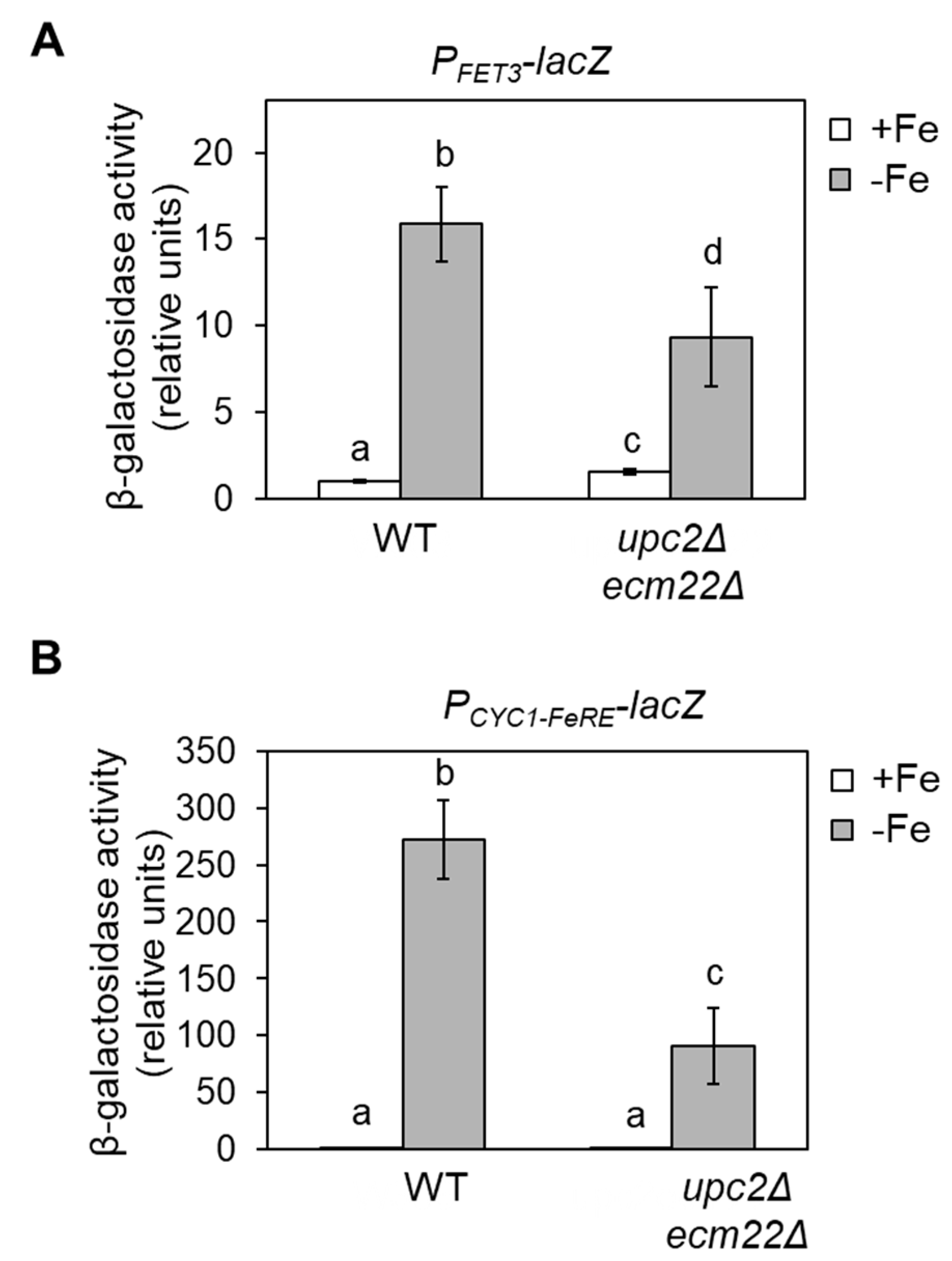
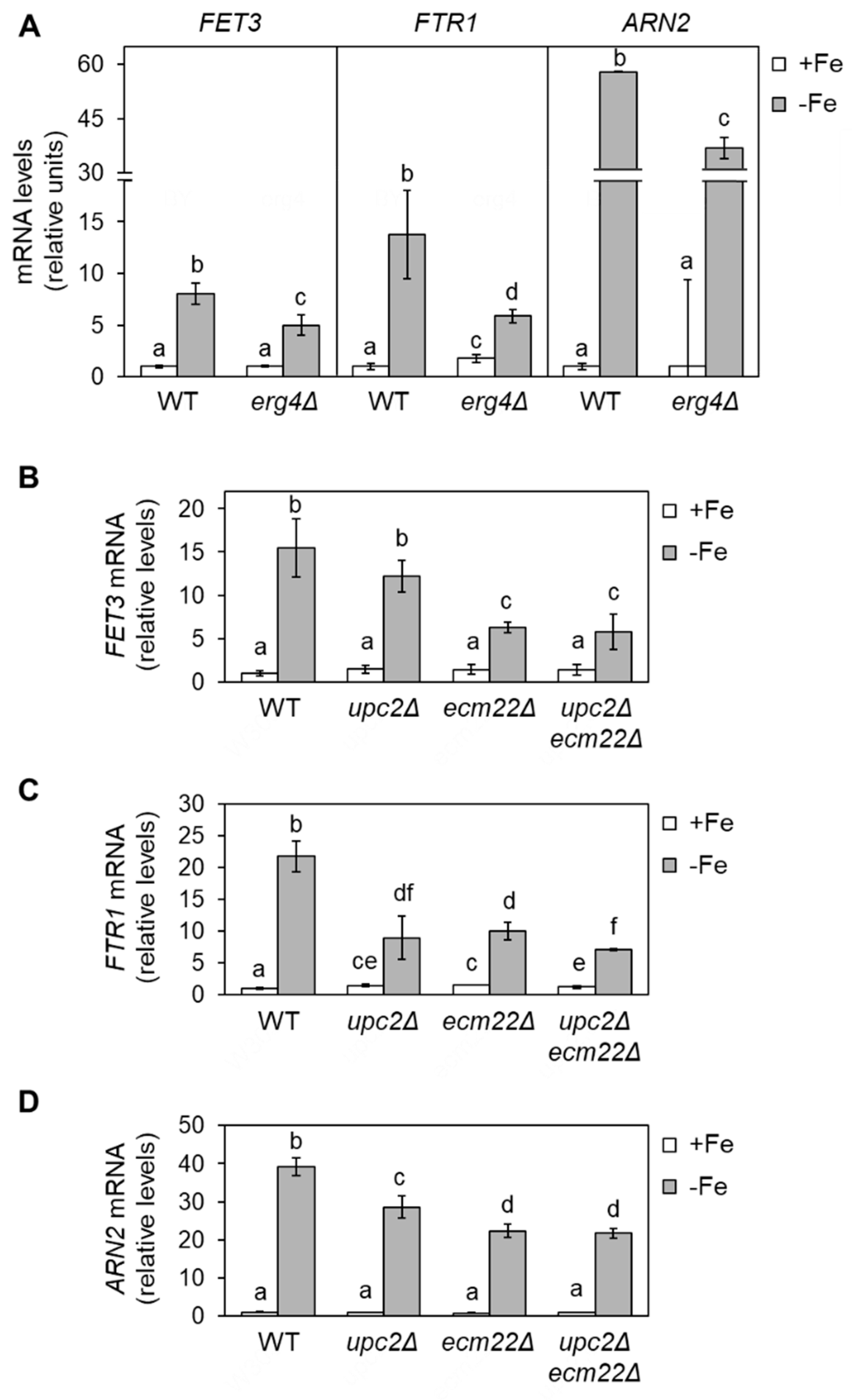
3.1. Full Activation of the Iron Regulon Requires Ergosterol Biosynthesis
3.2. Iron Deficiency Modifies the Sterol Profile of Deletion Mutants in UPC2 and ECM22 Genes
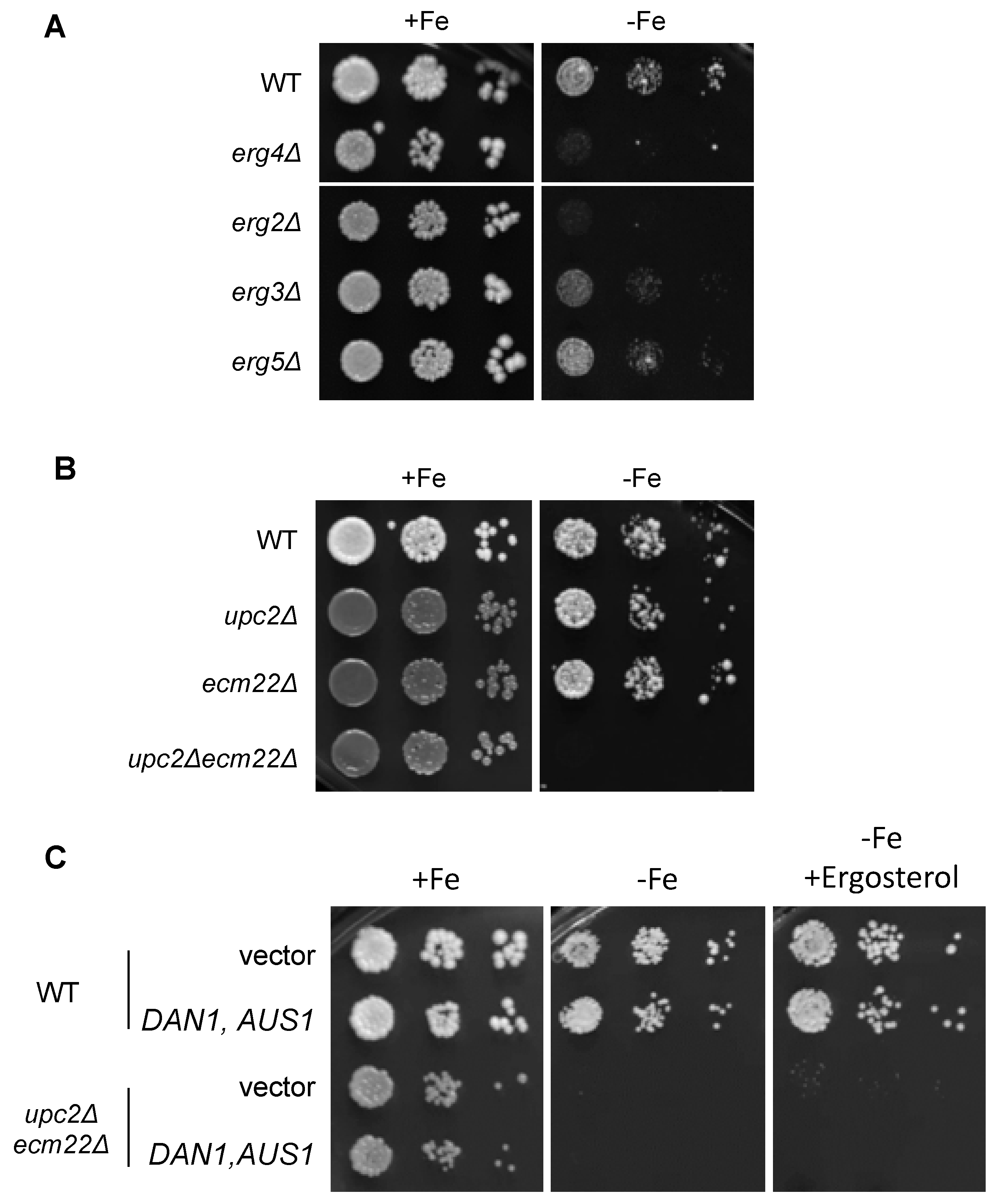
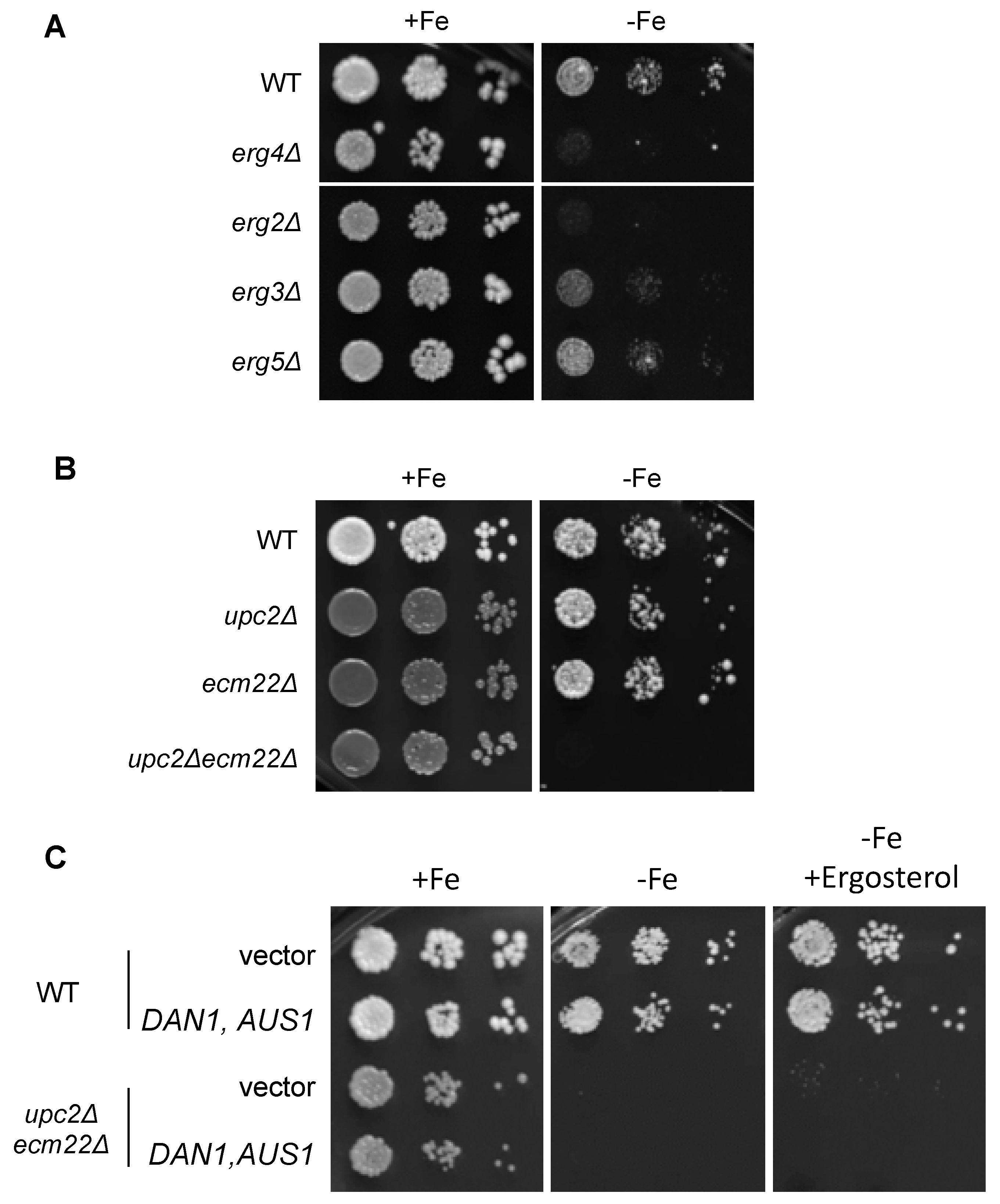
3.3. Growth in Iron-Deficient Conditions Requires Ergosterol Biosynthesis
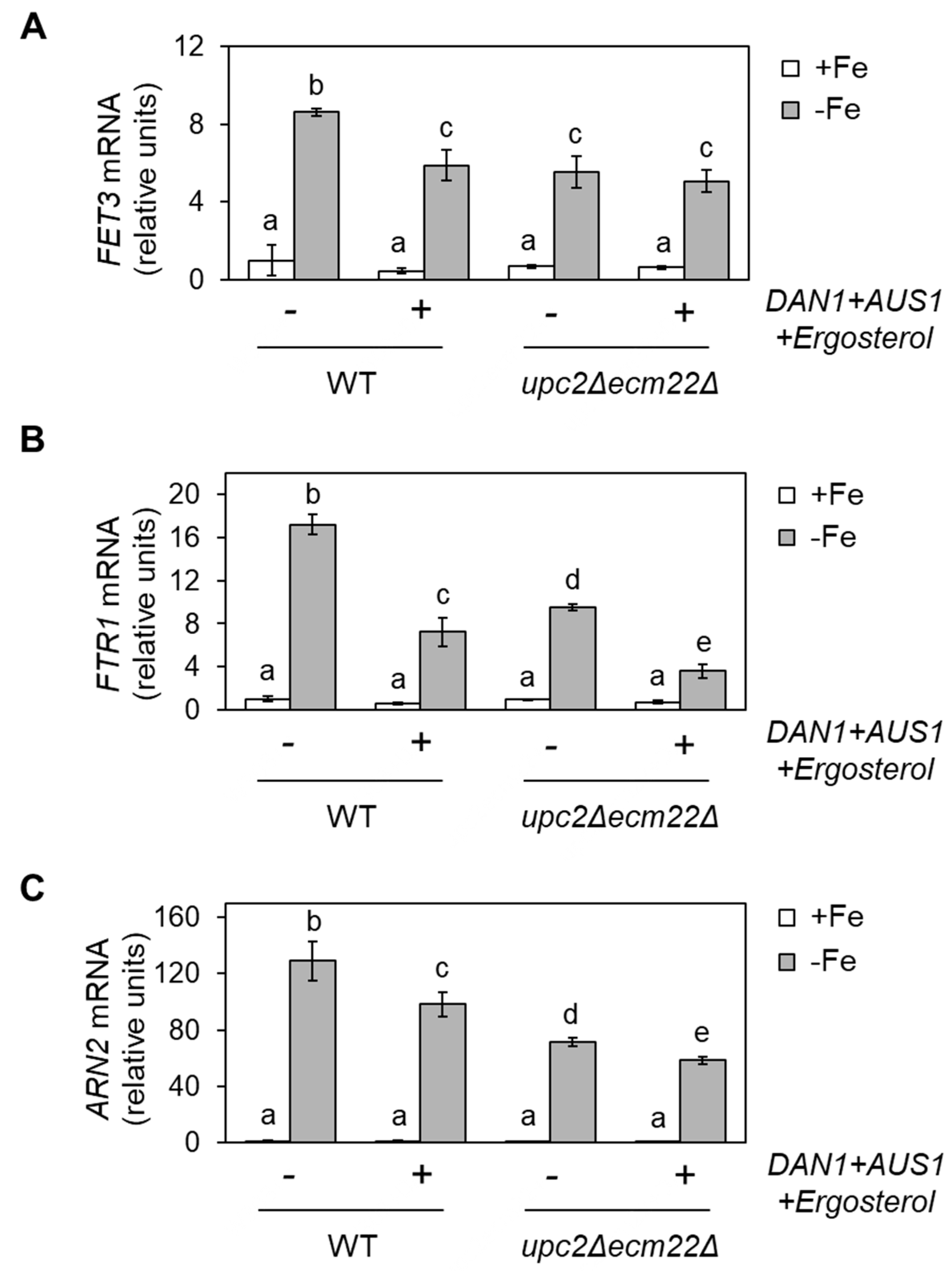
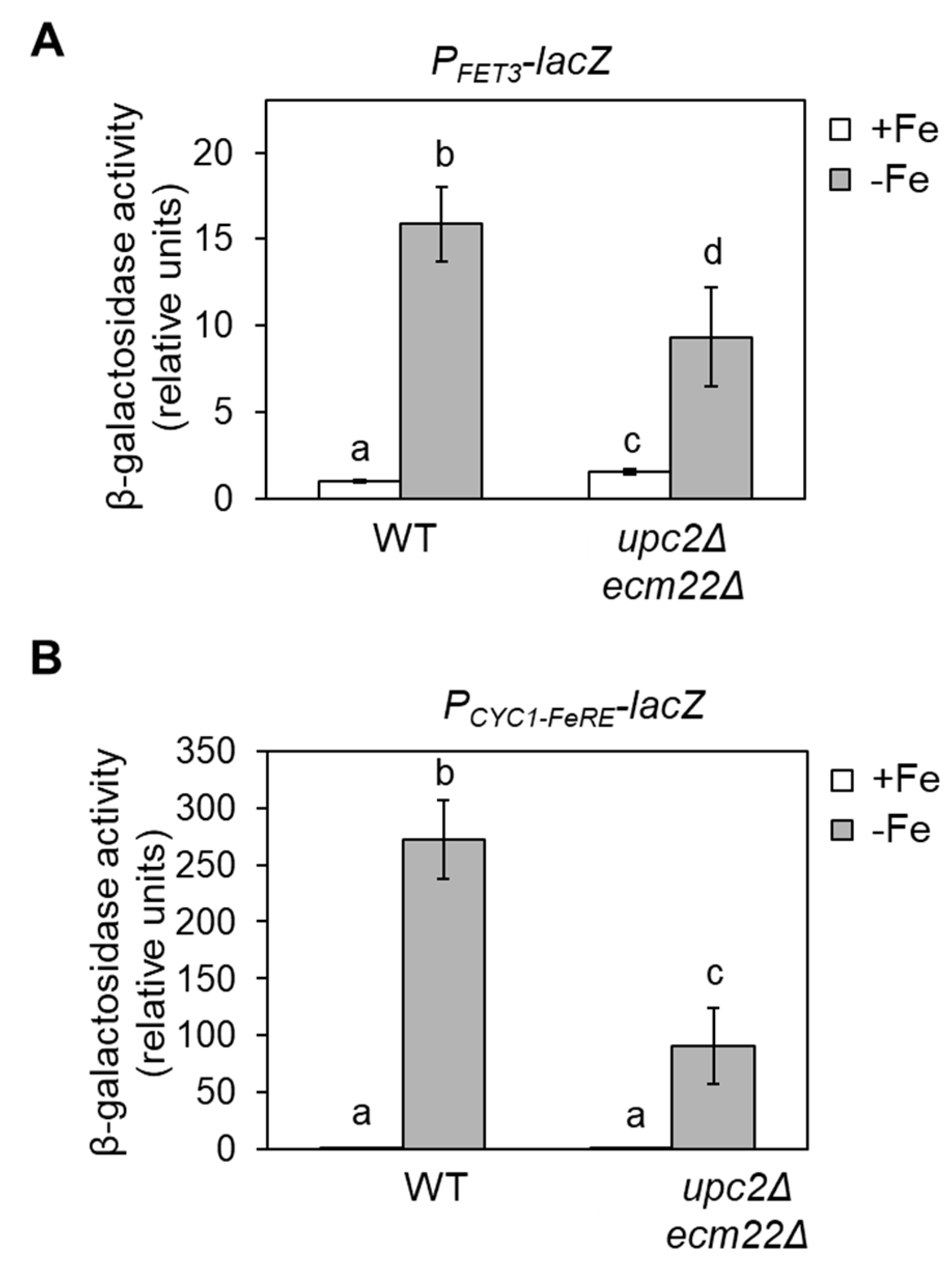
3.4. Upc2 and Ecm22 Facilitate Transcription upon Iron Deficiency through Iron Regulatory Elements
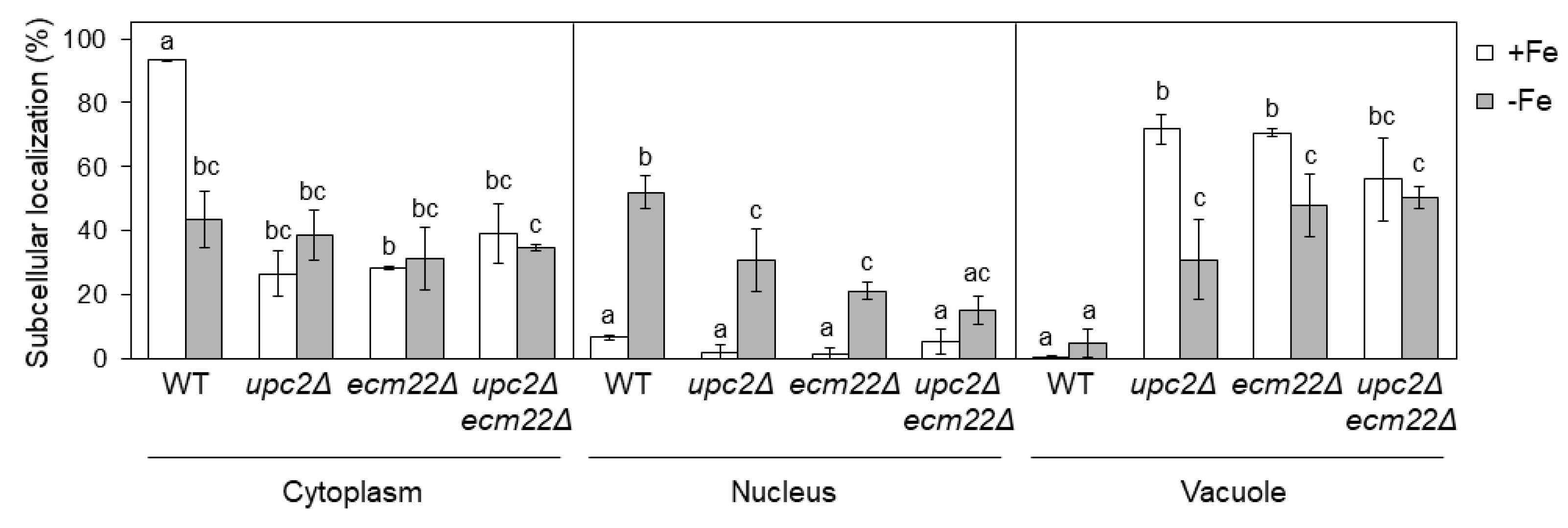
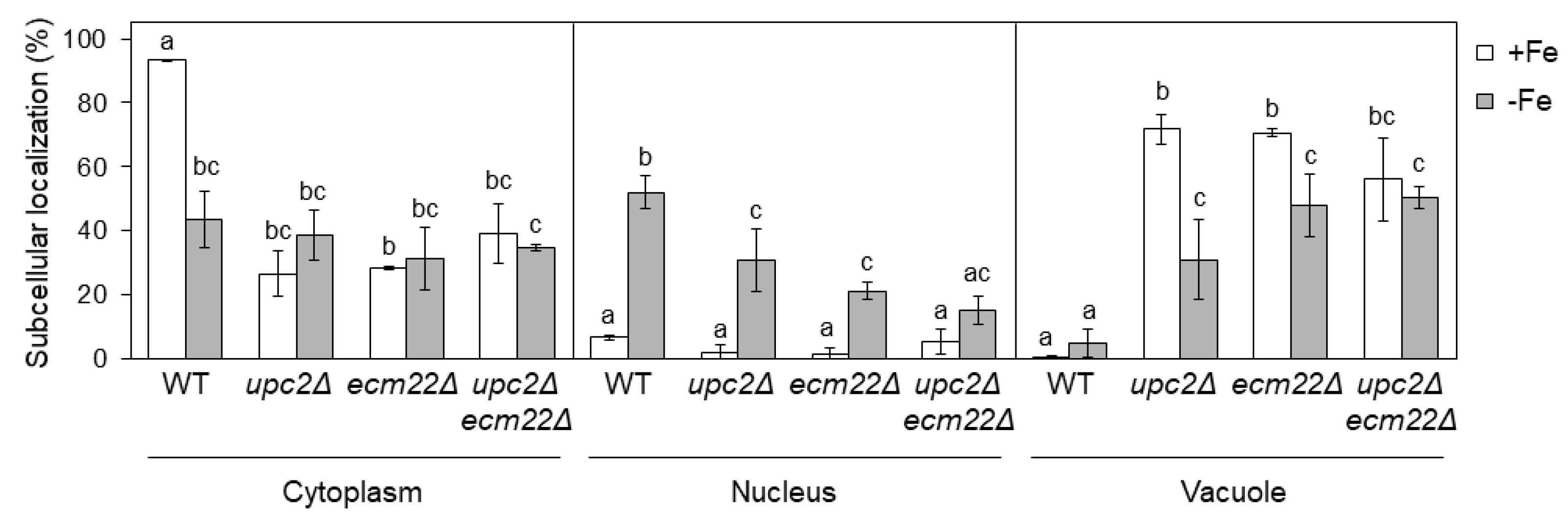
3.5. Upc2 and Ecm22 Are Required for Appropriate Aft1 Protein Localization
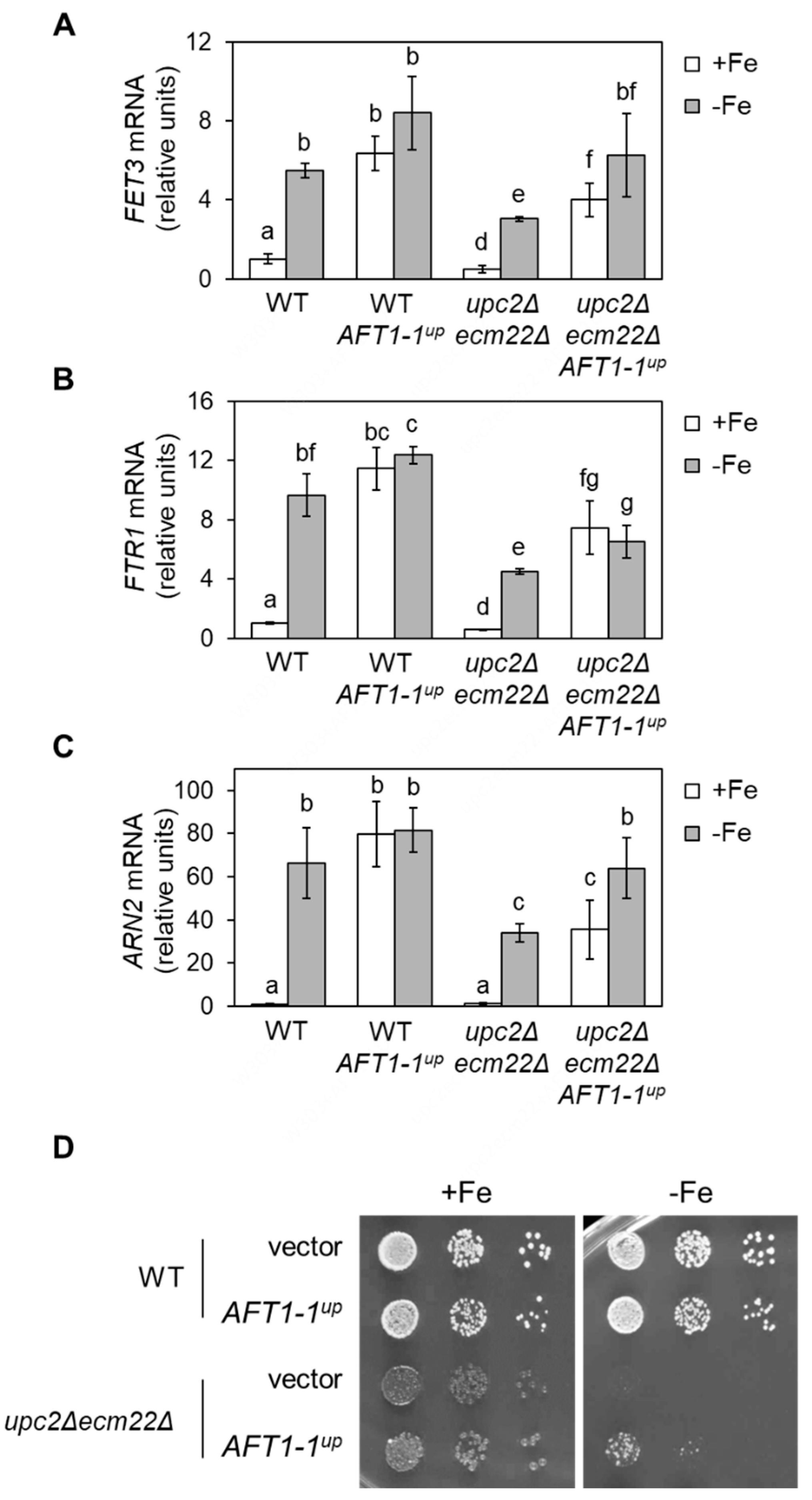
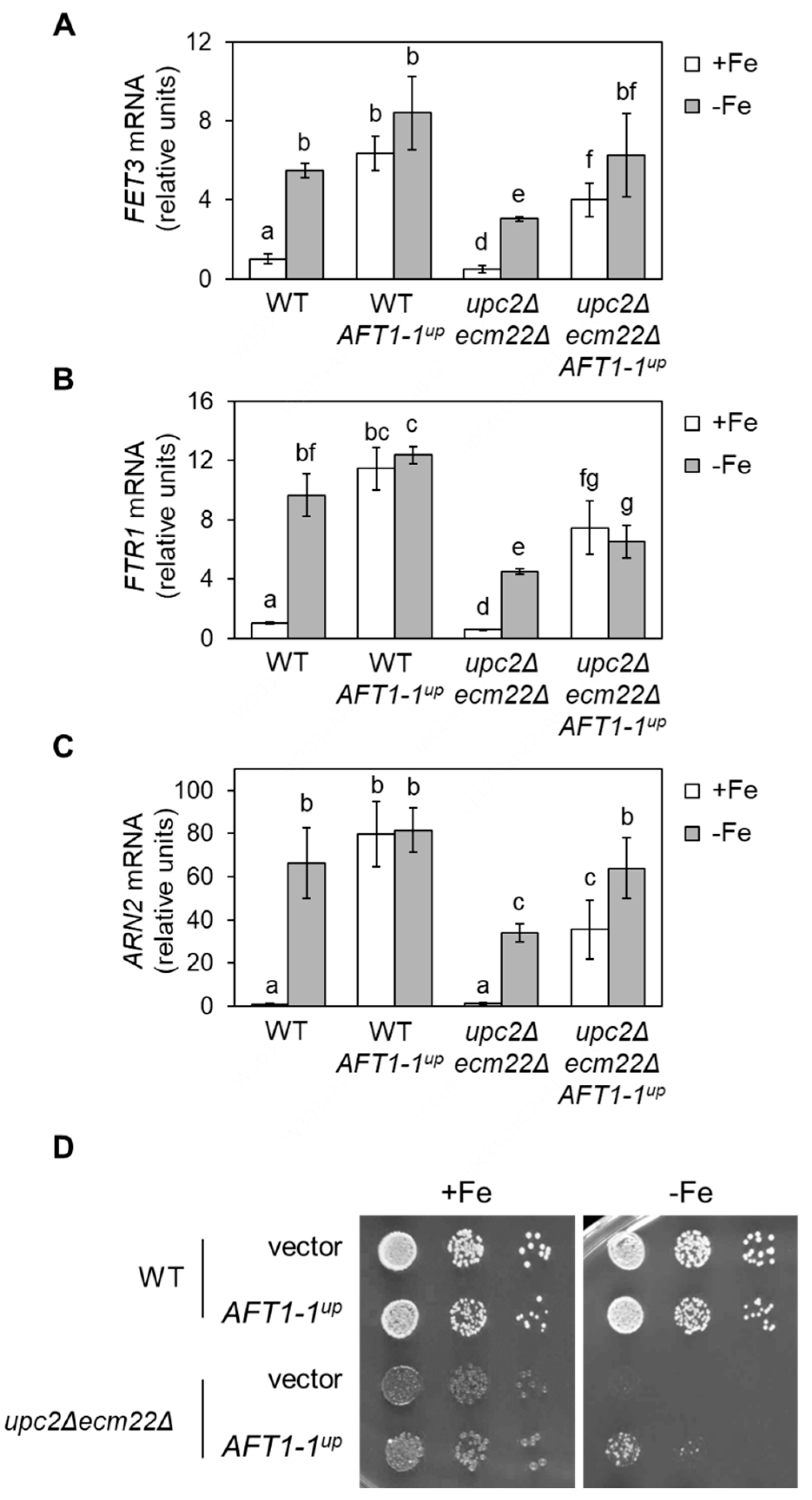
3.6. upc2Δecm22Δ Phenotypes Are Rescued by a Constitutively Nuclear and Active Aft1 Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zimmermann, M.B.; Hurrell, R.F. Nutritional iron deficiency. Lancet 2007, 370, 511–520. [Google Scholar] [CrossRef]
- Zimmermann, M.B. Global look at nutritional and functional iron deficiency in infancy. Hematology 2020, 2020, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Sanvisens, N.; Puig, S. Causes and consequences of nutritional iron deficiency in living organisms. In Biology of Starvation in Humans and Other Organisms; Merkin, T.C., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2011; pp. 245–276. [Google Scholar]
- Pas, M.; Piskur, B.; Sustaric, M.; Raspor, P. Iron enriched yeast biomass—A promising mineral feed supplement. Bioresour. Technol. 2007, 98, 1622–1628. [Google Scholar] [CrossRef]
- Kyyaly, M.A.; Powell, C.; Ramadan, E. Preparation of iron-enriched baker′s yeast and its efficiency in recovery of rats from dietary iron deficiency. Nutrition 2015, 31, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.G.; Wang, N.; Ma, G.D.; Liu, Z.Y.; Wei, G.X.; Liu, W.J. Preparation of S-iron-enriched yeast using siderophores and its effect on iron deficiency anemia in rats. Food Chem. 2021, 365, 130508. [Google Scholar] [CrossRef]
- Outten, C.E.; Albetel, A.N. Iron sensing and regulation in Saccharomyces cerevisiae: Ironing out the mechanistic details. Curr. Opin. Microbiol. 2013, 16, 662–668. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Alonso, L.; Romero, A.M.; Martínez-Pastor, M.T.; Puig, S. Iron regulatory mechanisms in Saccharomyces cerevisiae. Front. Microbiol. 2020, 11, 582830. [Google Scholar] [CrossRef]
- Jordá, T.; Puig, S. Regulation of ergosterol biosynthesis in Saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef]
- Shakoury-Elizeh, M.; Protchenko, O.; Berger, A.; Cox, J.; Gable, K.; Dunn, T.M.; Prinz, W.A.; Bard, M.; Philpott, C.C. Metabolic response to iron deficiency in Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 14823–14833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, A.M.; Jorda, T.; Rozes, N.; Martinez-Pastor, M.T.; Puig, S. Regulation of yeast fatty acid desaturase in response to iron deficiency. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Zinser, E.; Paltauf, F.; Daum, G. Sterol composition of yeast organelle membranes and subcellular distribution of enzymes involved in sterol metabolism. J. Bacteriol. 1993, 175, 2853–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuller, G.; Nemec, T.; Hrastnik, C.; Daum, G. Lipid composition of subcellular membranes of an FY1679-derived haploid yeast wild-type strain grown on different carbon sources. Yeast 1999, 15, 1555–1564. [Google Scholar] [CrossRef]
- Ernst, R.; Ballweg, S.; Levental, I. Cellular mechanisms of physicochemical membrane homeostasis. Curr. Opin. Cell Biol. 2018, 53, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Johnston, E.J.; Moses, T.; Rosser, S.J. The wide-ranging phenotypes of ergosterol biosynthesis mutants, and implications for microbial cell factories. Yeast 2020, 37, 27–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valachovic, M.; Wilcox, L.I.; Sturley, S.L.; Bard, M. A mutation in sphingolipid synthesis suppresses defects in yeast ergosterol metabolism. Lipids 2004, 39, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Cirigliano, A.; Amelina, A.; Biferali, B.; Macone, A.; Mozzetta, C.; Bianchi, M.M.; Mori, M.; Botta, B.; Pick, E.; Negri, R.; et al. Statins interfere with the attachment of S. cerevisiae mtDNA to the inner mitochondrial membrane. J. Enzyme. Inhib. Med. Chem. 2020, 35, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Demuyser, L.; Van Dijck, P. Can Saccharomyces cerevisiae keep up as a model system in fungal azole susceptibility research? Drug Resist. Updat. 2019, 42, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Jordá, T.; Romero, A.M.; Perea-García, A.; Rozès, N.; Puig, S. The lipid composition of yeast cells modulates the response to iron deficiency. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158707. [Google Scholar] [CrossRef] [PubMed]
- Sanvisens, N.; Romero, A.M.; An, X.; Zhang, C.; de Llanos, R.; Martínez-Pastor, M.T.; Banó, M.C.; Huang, M.; Puig, S. Yeast Dun1 kinase regulates ribonucleotide reductase inhibitor Sml1 in response to iron deficiency. Mol. Cell Biol. 2014, 34, 3259–3271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrull, A.; Poblet, M.; Rozes, N. New insights into the capacity of commercial wine yeasts to grow on sparkling wine media. Factor screening for improving wine yeast selection. Food Microbiol. 2015, 48, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Borrull, A.; López-Martínez, G.; Poblet, M.; Cordero-Otero, R.; Rozès, N. A simple method for the separation and quantification of neutral lipid species using GC-MS. Eur. J. Lipid Sci. Technol. 2015, 117, 274–280. [Google Scholar] [CrossRef]
- Puig, S.; Lau, M.; Thiele, D.J. Cti6 is an Rpd3-Sin3 histone deacetylase-associated protein required for growth under iron-limiting conditions in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 30298–30306. [Google Scholar] [CrossRef] [Green Version]
- Alimardani, P.; Regnacq, M.; Moreau-Vauzelle, C.; Ferreira, T.; Rossignol, T.; Blondin, B.; Berges, T. SUT1-promoted sterol uptake involves the ABC transporter Aus1 and the mannoprotein Dan1 whose synergistic action is sufficient for this process. Biochem. J. 2004, 381, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Puig, S.; Askeland, E.; Thiele, D.J. Coordinated remodeling of cellular metabolism during iron deficiency through targeted mRNA degradation. Cell 2005, 120, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi-Iwai, Y.; Ueta, R.; Fukunaka, A.; Sasaki, R. Subcellular localization of Aft1 transcription factor responds to iron status in Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 18914–18918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi-Iwai, Y.; Dancis, A.; Klausner, R.D. AFT1: A mediator of iron regulated transcriptional control in Saccharomyces cerevisiae. EMBO J. 1995, 14, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Iwai, Y.; Stearman, R.; Dancis, A.; Klausner, R.D. Iron-regulated DNA binding by the AFT1 protein controls the iron regulon in yeast. EMBO J. 1996, 15, 3377–3384. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.J.; Kim, J.H.; Oh, E.; Jaramillo, D.; Holman, P.; Loguinov, A.V.; Arkin, A.P.; Nislow, C.; Giaever, G.; Vulpe, C.D. Novel insights into iron metabolism by integrating deletome and transcriptome analysis in an iron deficiency model of the yeast Saccharomyces cerevisiae. BMC Genom. 2009, 10, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craven, R.J.; Mallory, J.C.; Hand, R.A. Regulation of iron homeostasis mediated by the heme-binding protein Dap1 (damage resistance protein 1) via the P450 protein Erg11/Cyp51. J. Biol. Chem. 2007, 282, 36543–36551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallory, J.C.; Crudden, G.; Johnson, B.L.; Mo, C.; Pierson, C.A.; Bard, M.; Craven, R.J. Dap1p, a heme-binding protein that regulates the cytochrome P450 protein Erg11p/Cyp51p in Saccharomyces cerevisiae. Mol. Cell Biol. 2005, 25, 1669–1679. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Kaplan, J. Characterization of yeast methyl sterol oxidase (ERG25) and identification of a human homologue. J. Biol. Chem. 1996, 271, 16927–16933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, D.M.; Chen, O.S.; Li, L.; Kaplan, J.; Bhuiyan, S.A.; Natarajan, S.K.; Bard, M.; Cox, J.E. Altered sterol metabolism in budding yeast affects mitochondrial iron-sulfur (Fe-S) cluster synthesis. J. Biol. Chem. 2018, 293, 10782–10795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or deletion of ergosterol biosynthesis genes alters doubling time, response to stress agents, and drug susceptibility in Saccharomyces cerevisiae. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Vasicek, E.M.; Berkow, E.L.; Flowers, S.A.; Barker, K.S.; Rogers, P.D. UPC2 is universally essential for azole antifungal resistance in Candida albicans. Eukaryot. Cell 2014, 13, 933–946. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.; Kastora, S.L.; Gomes-Goncalves, A.; Azevedo, N.; Rodrigues, C.F.; Silva, S.; Demuyser, L.; Van Dijck, P.; Casal, M.; Brown, A.J.P.; et al. Transcriptional responses of Candida glabrata biofilm cells to fluconazole are modulated by the carbon source. NPJ Biofilms Microbiomes 2020, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Cohen, N.; Breker, M.; Bakunts, A.; Pesek, K.; Chas, A.; Argemi, J.; Orsi, A.; Gal, L.; Chuartzman, S.; Wigelman, Y.; et al. Iron affects Ire1 clustering propensity and the amplitude of endoplasmic reticulum stress signaling. J. Cell Sci. 2017, 130, 3222–3233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolov, S.S.; Trushina, N.I.; Severin, F.F.; Knorre, D.A. Ergosterol turnover in yeast: An interplay between biosynthesis and transport. Biochemistry 2019, 84, 346–357. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jordá, T.; Rozès, N.; Puig, S. Sterol Composition Modulates the Response of Saccharomyces cerevisiae to Iron Deficiency. J. Fungi 2021, 7, 901. https://doi.org/10.3390/jof7110901
Jordá T, Rozès N, Puig S. Sterol Composition Modulates the Response of Saccharomyces cerevisiae to Iron Deficiency. Journal of Fungi. 2021; 7(11):901. https://doi.org/10.3390/jof7110901
Chicago/Turabian StyleJordá, Tania, Nicolas Rozès, and Sergi Puig. 2021. "Sterol Composition Modulates the Response of Saccharomyces cerevisiae to Iron Deficiency" Journal of Fungi 7, no. 11: 901. https://doi.org/10.3390/jof7110901
APA StyleJordá, T., Rozès, N., & Puig, S. (2021). Sterol Composition Modulates the Response of Saccharomyces cerevisiae to Iron Deficiency. Journal of Fungi, 7(11), 901. https://doi.org/10.3390/jof7110901