
Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Specimens, Isolation, Morphology, and Culture Characteristics
2.2. DNA Extraction and Molecular Based Amplification
2.3. Sequence Alignment and Phylogenetic Analyses
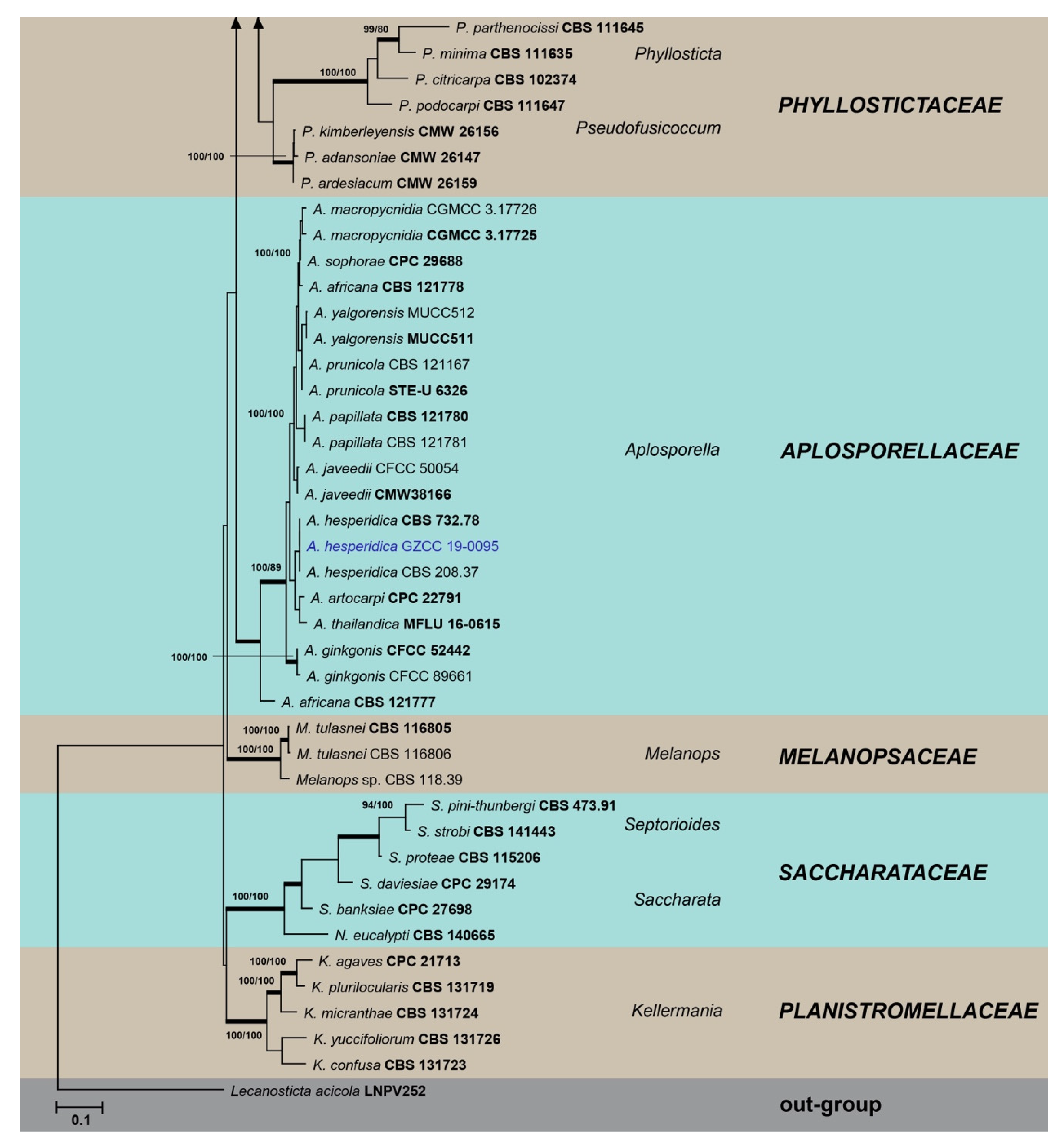
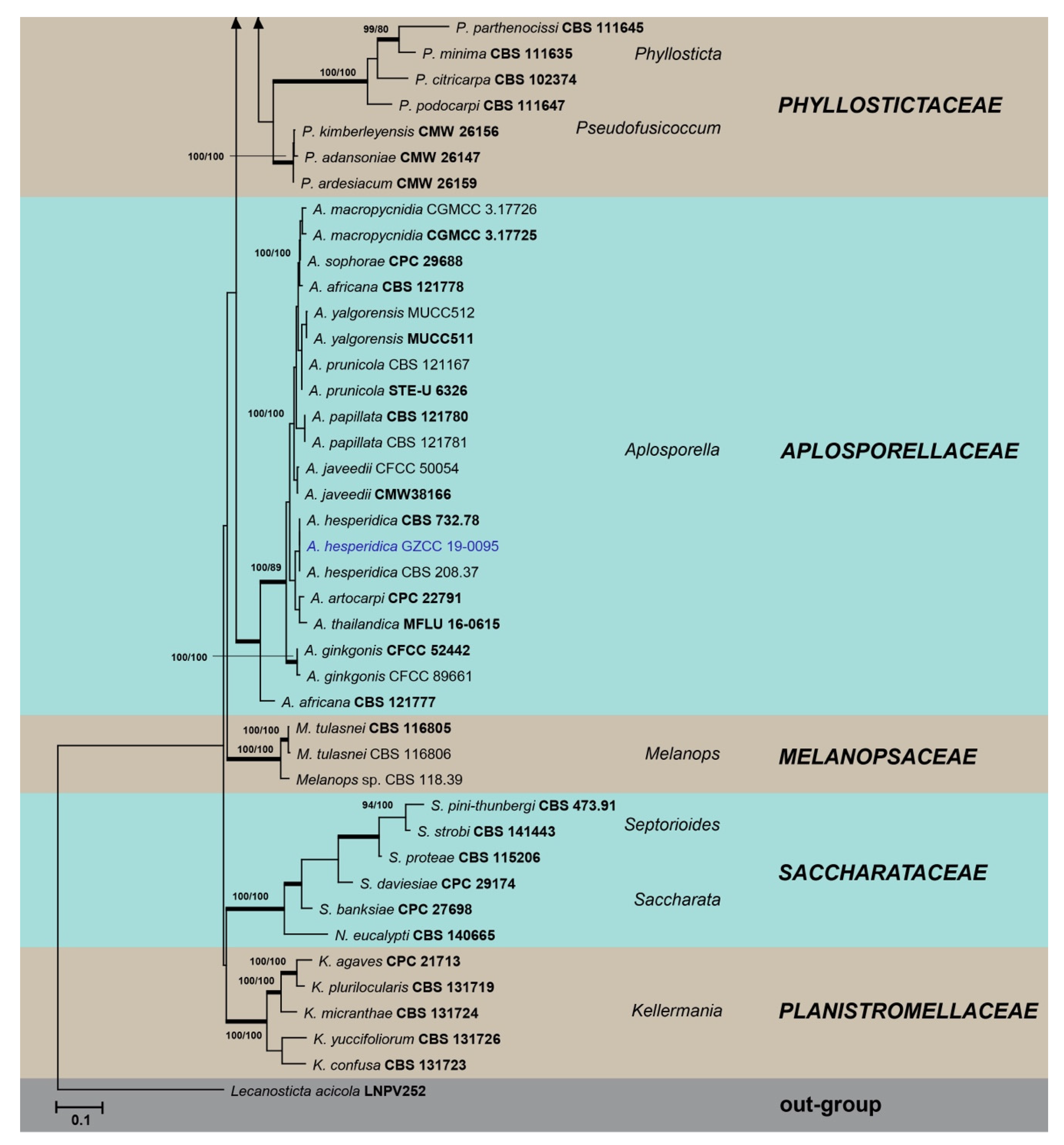
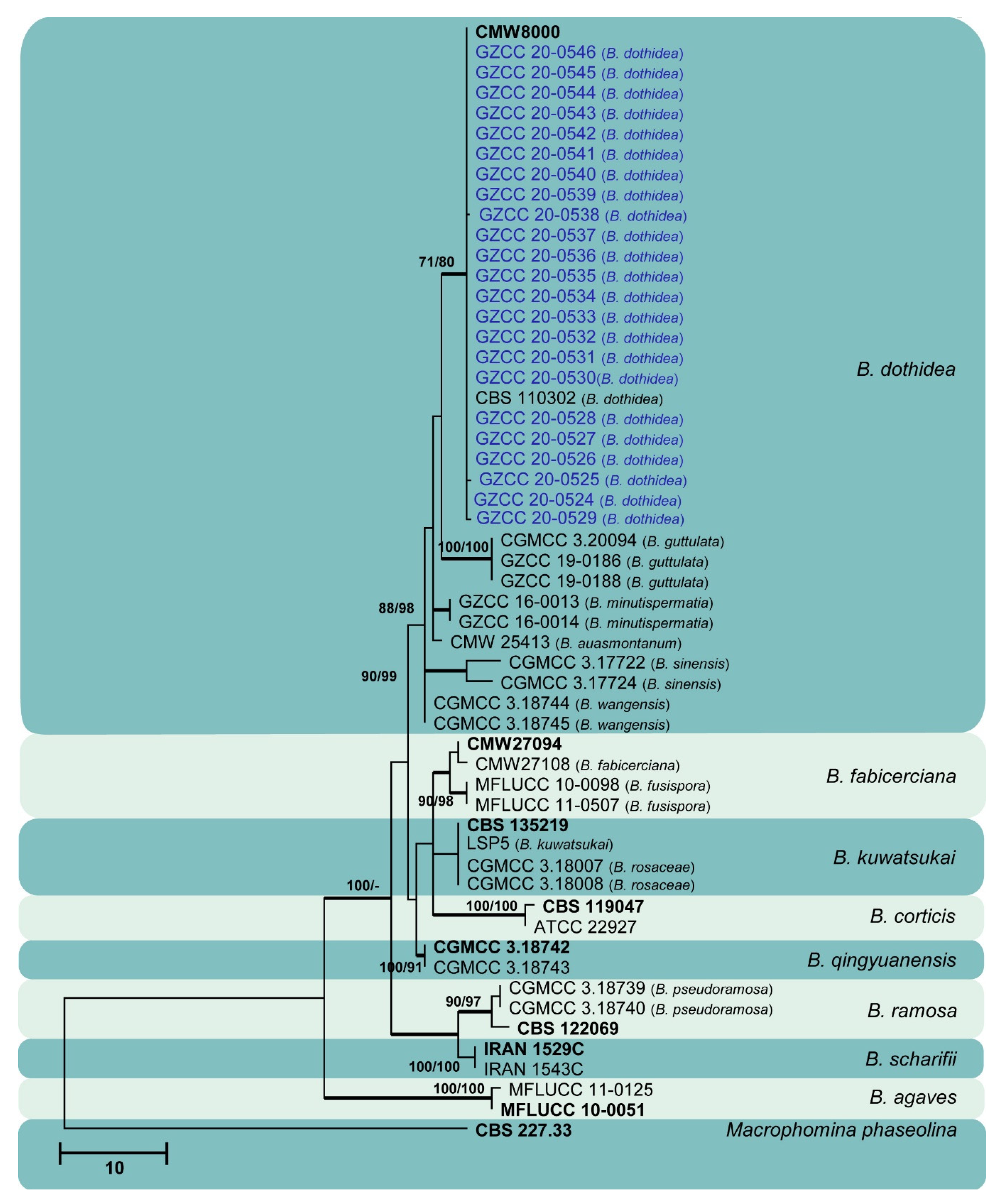
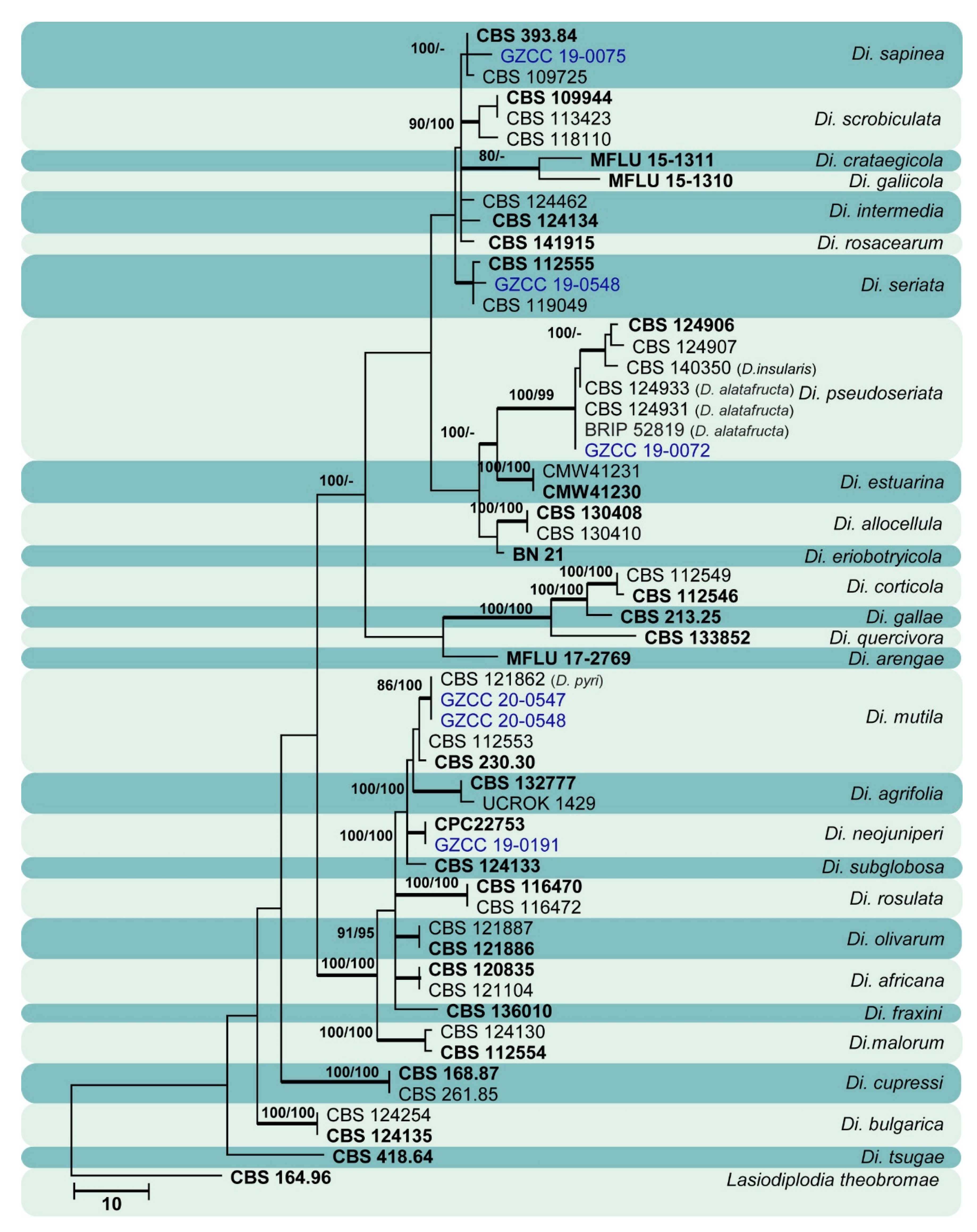
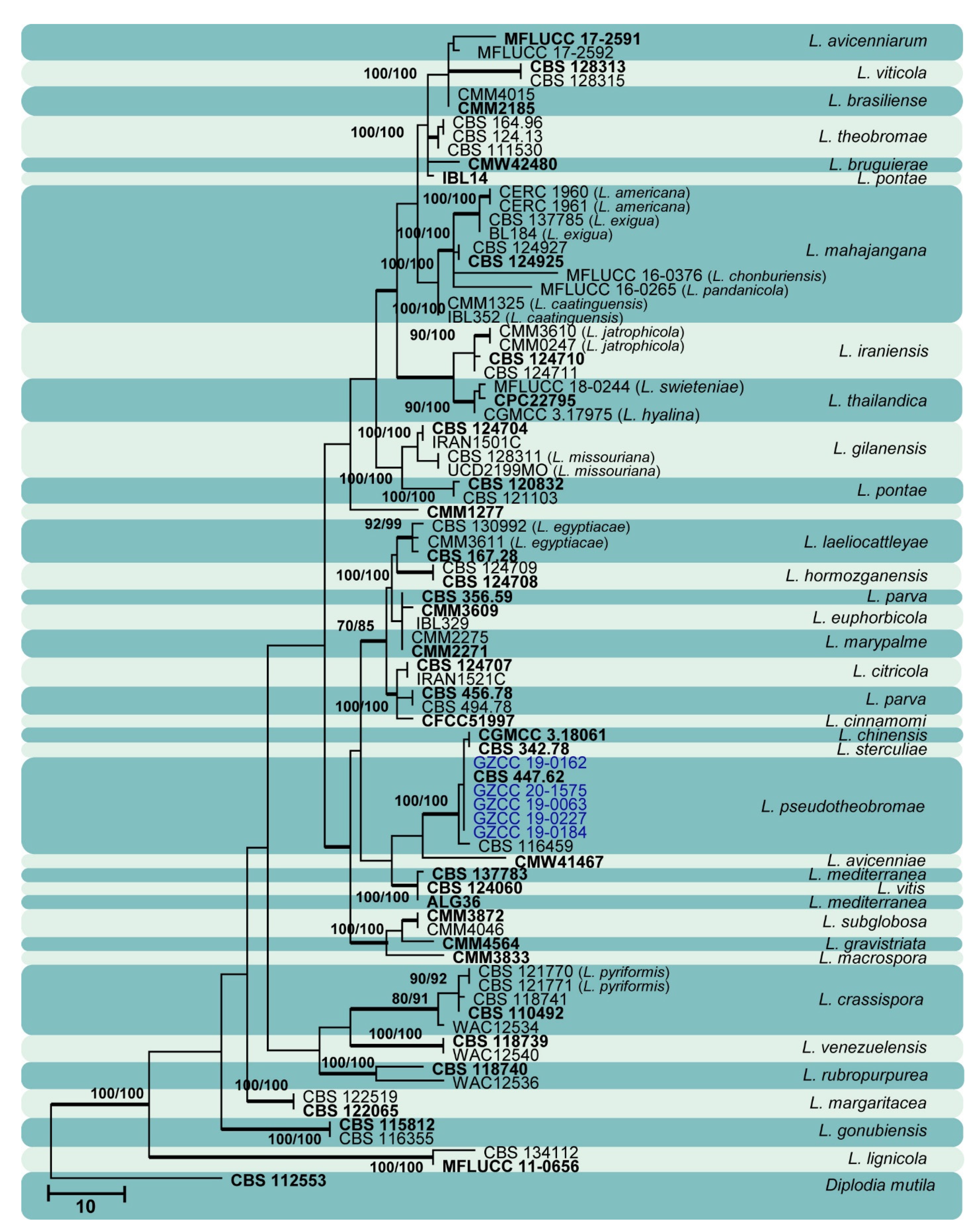
3. Results
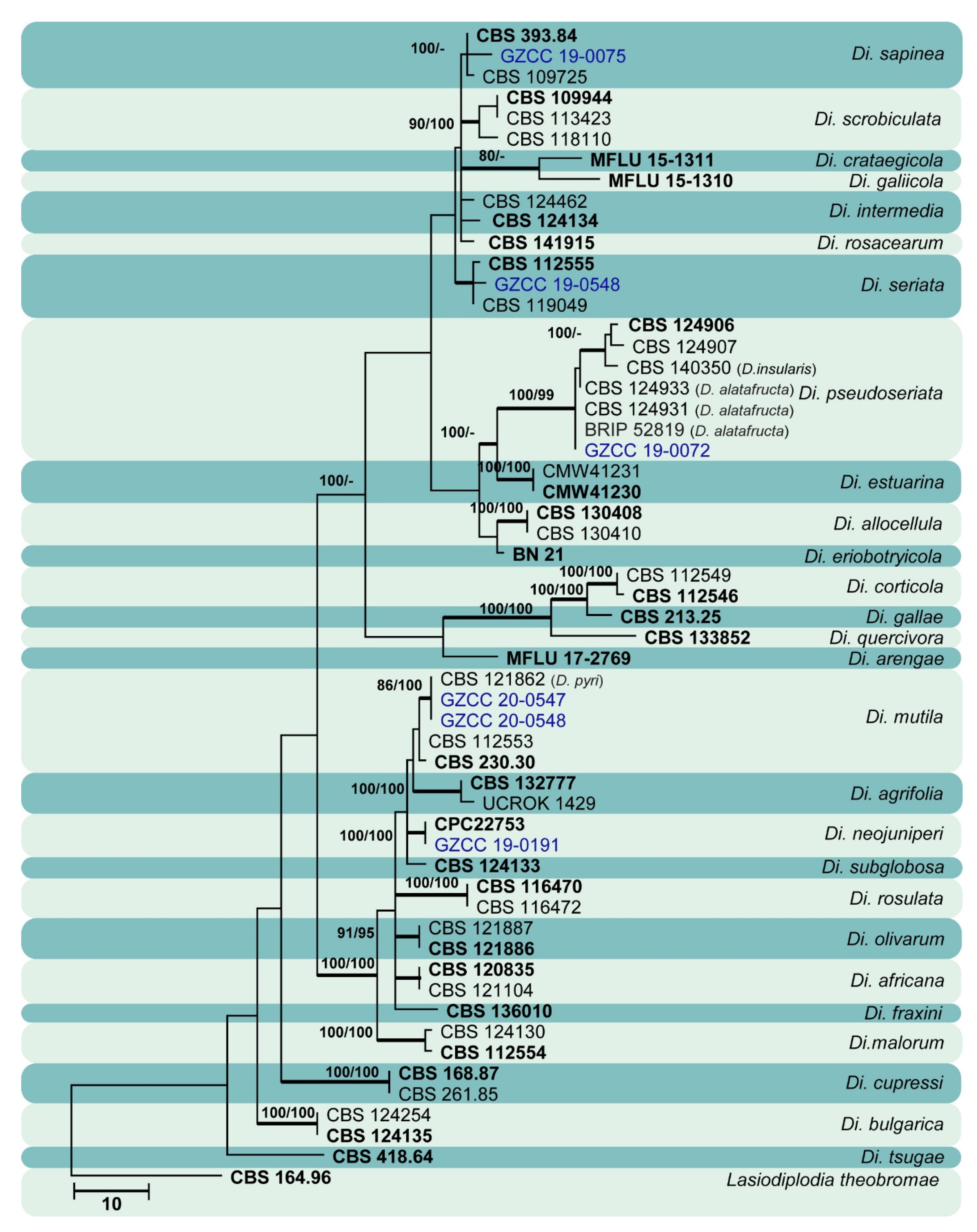
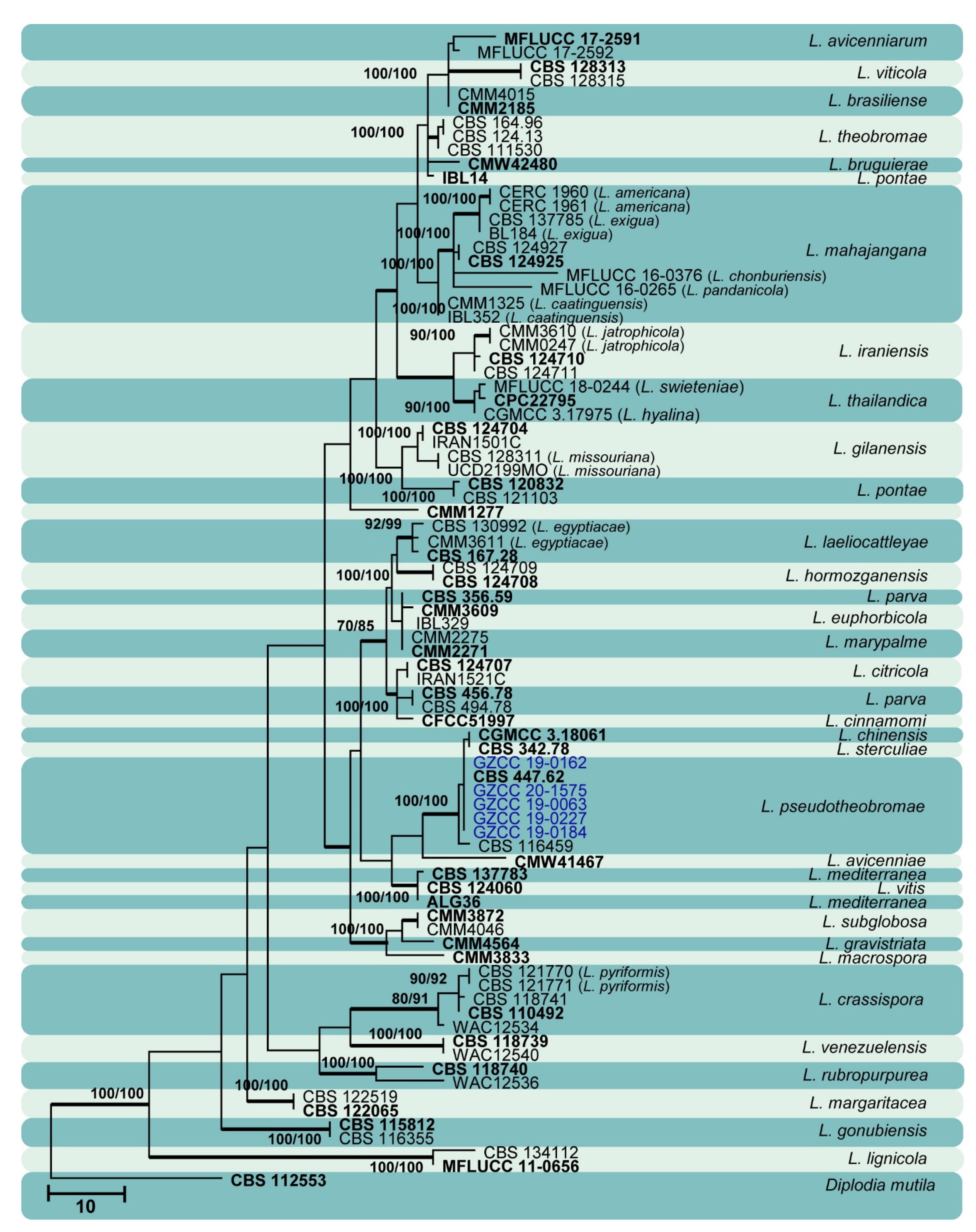
3.1. Phylogenetic Analyses
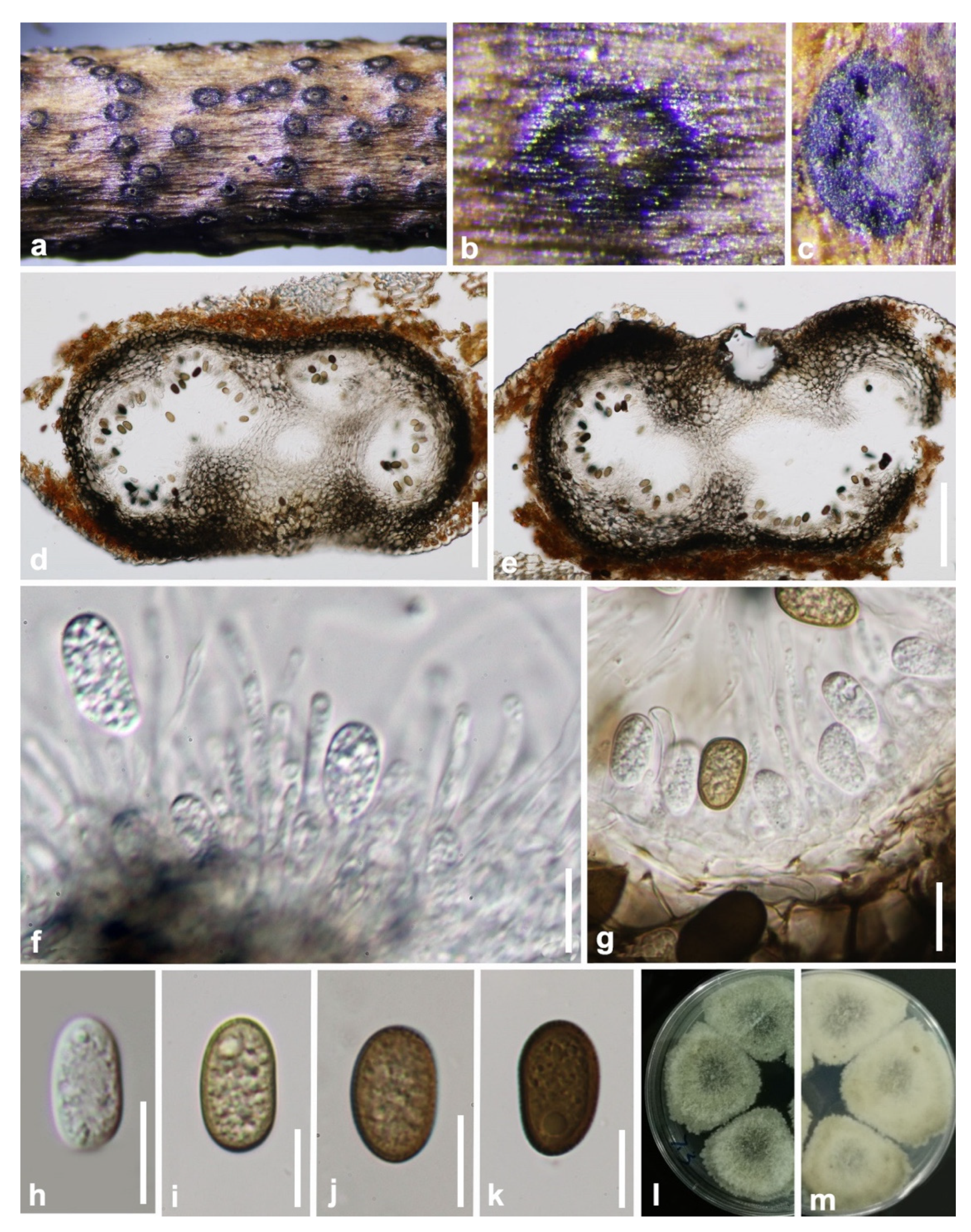
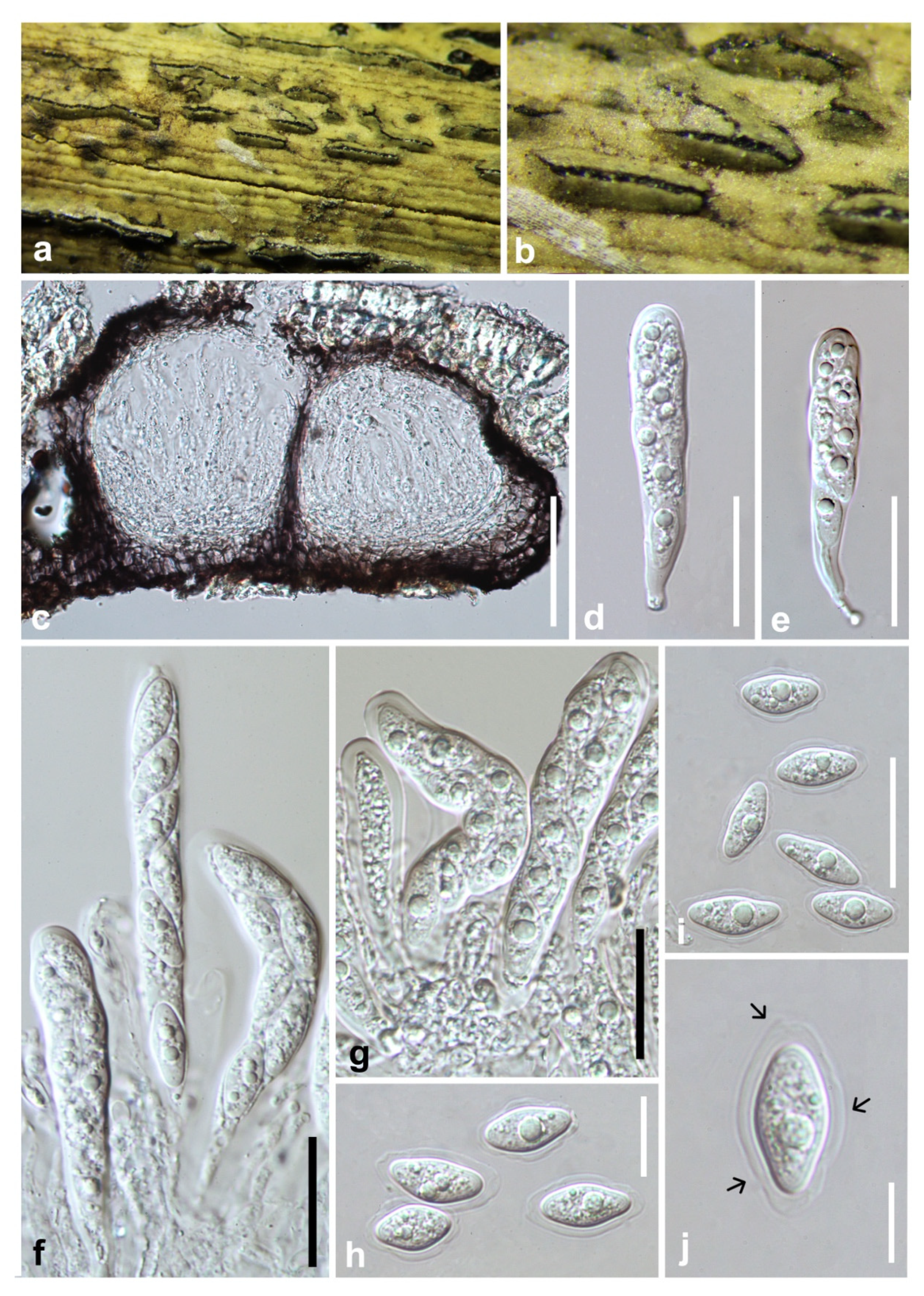
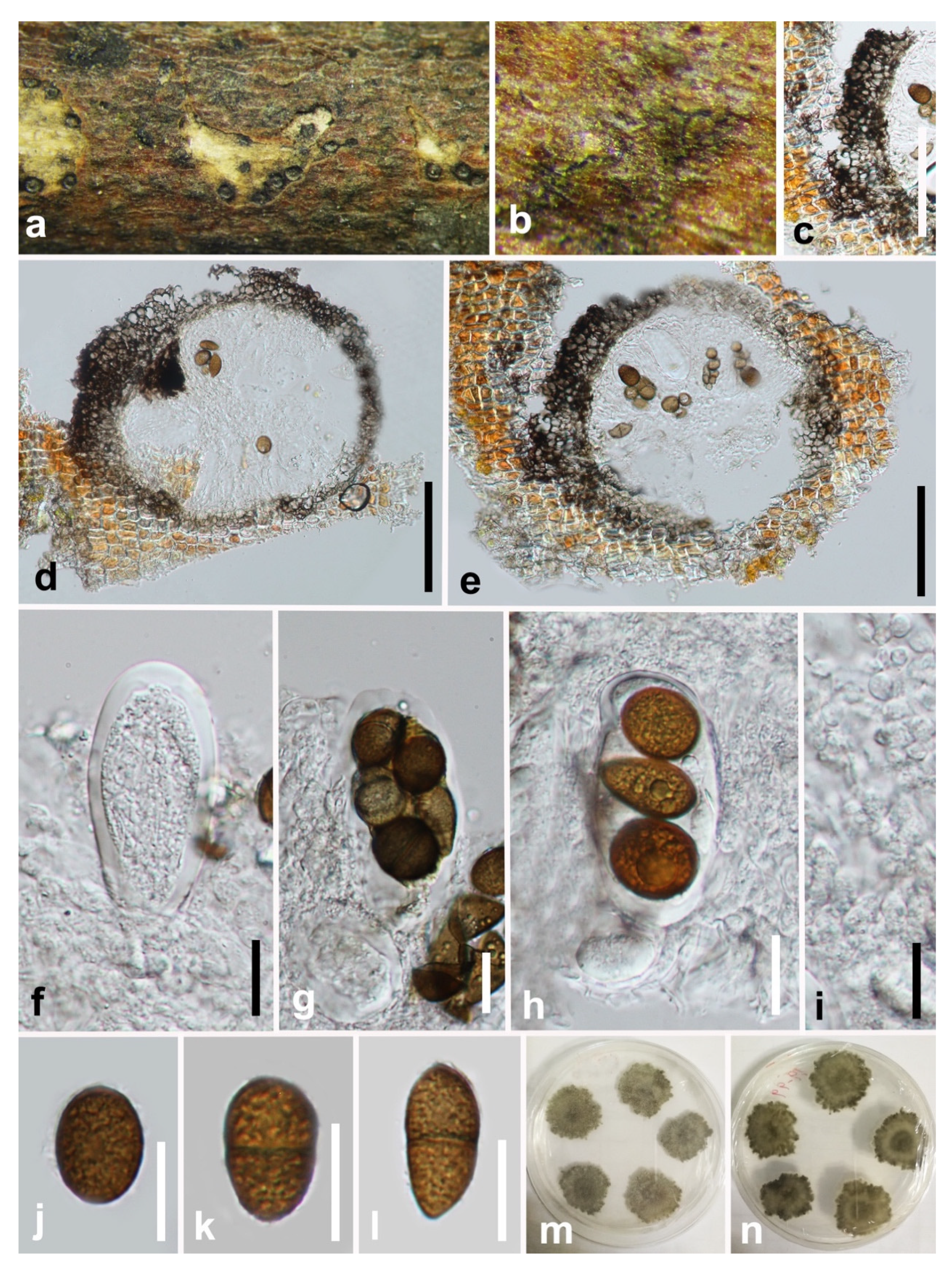
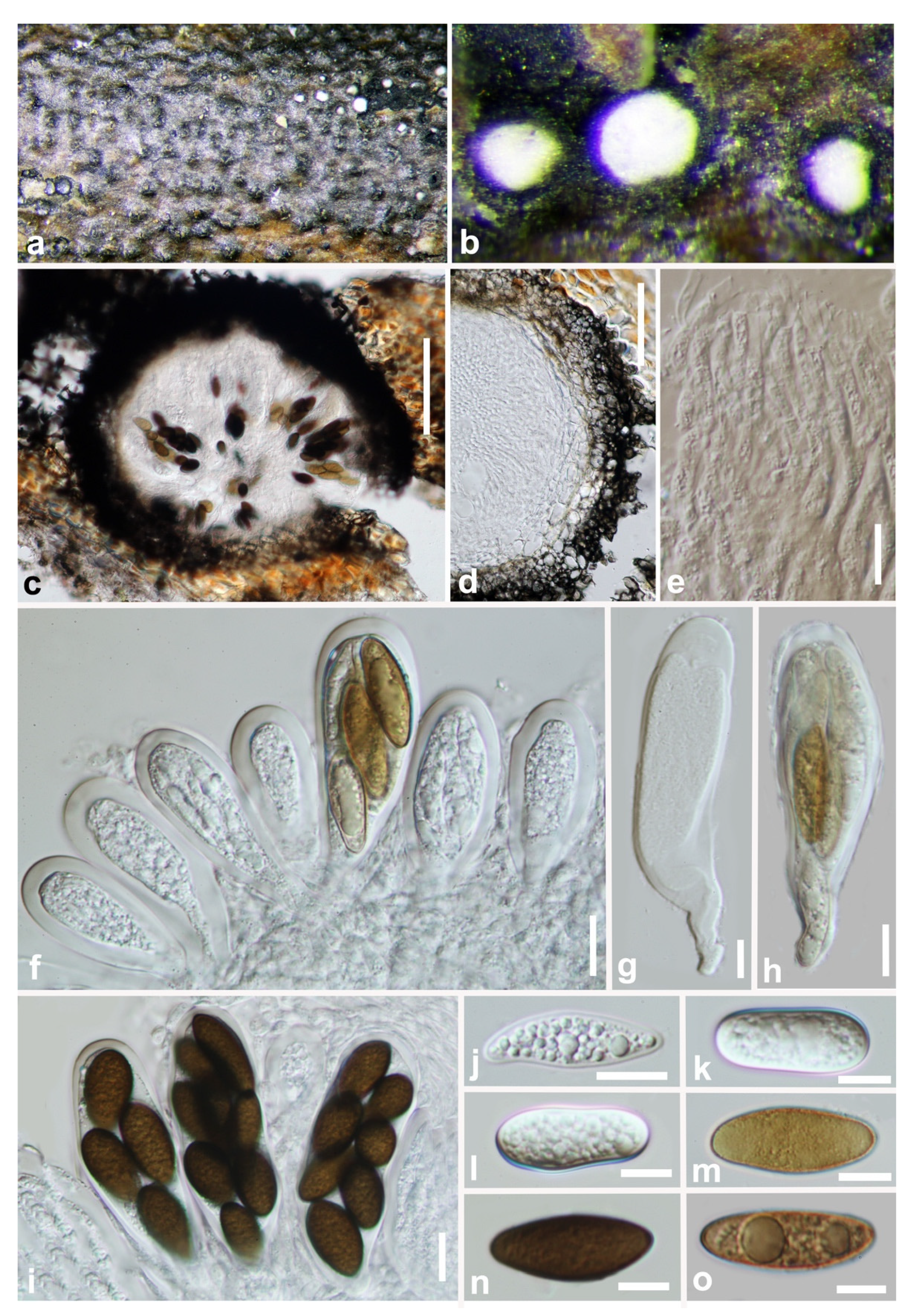
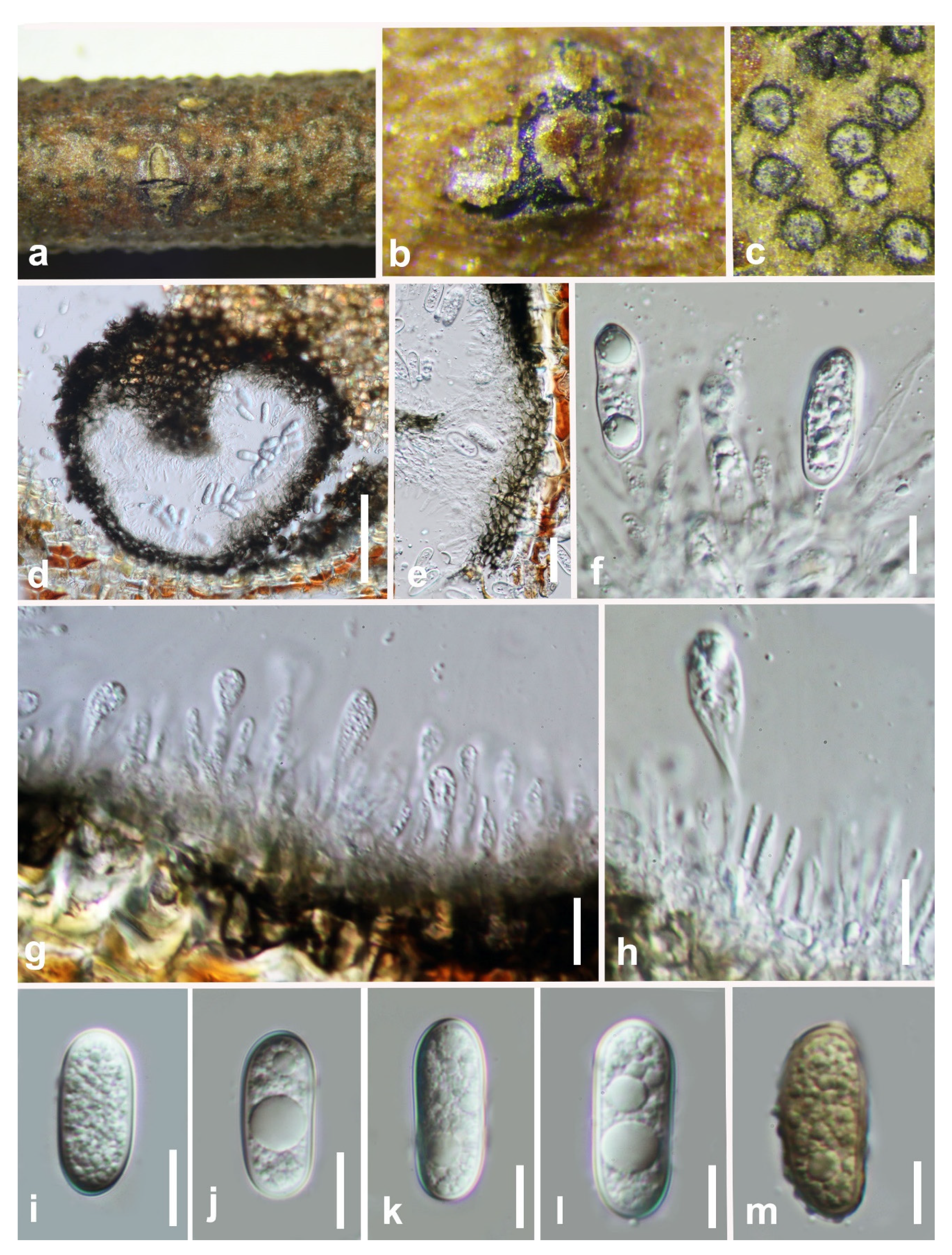
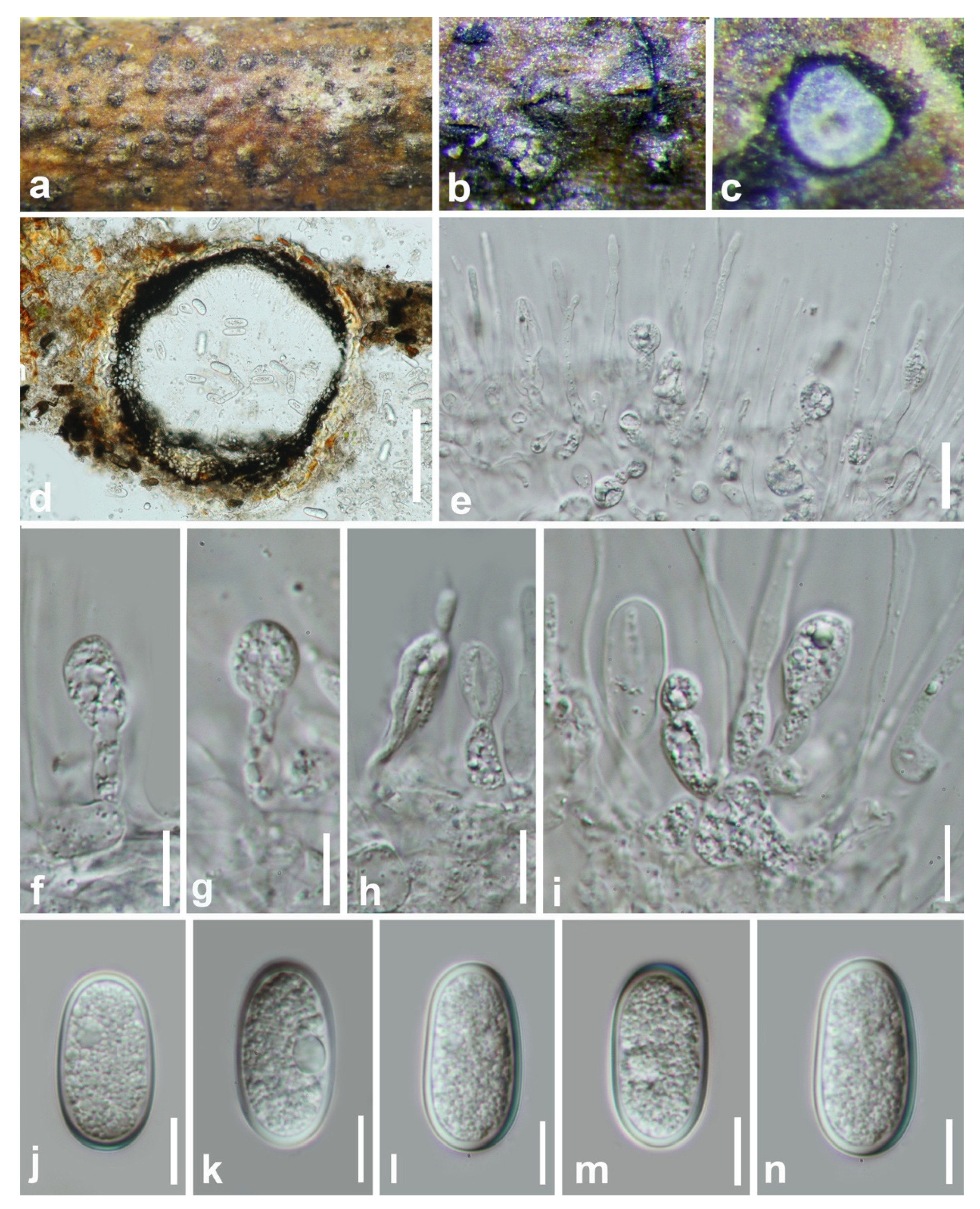
3.2. Taxonomy


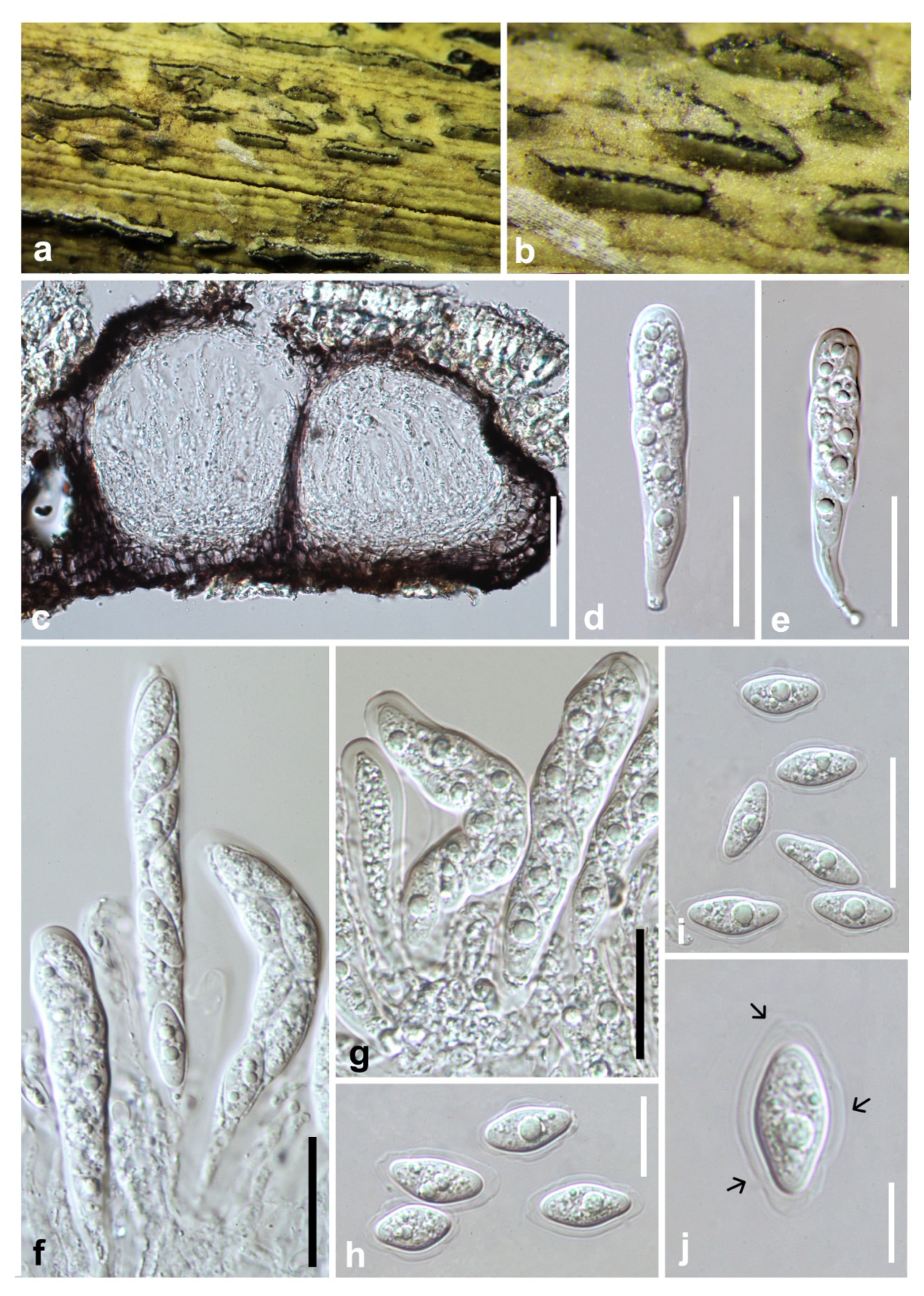
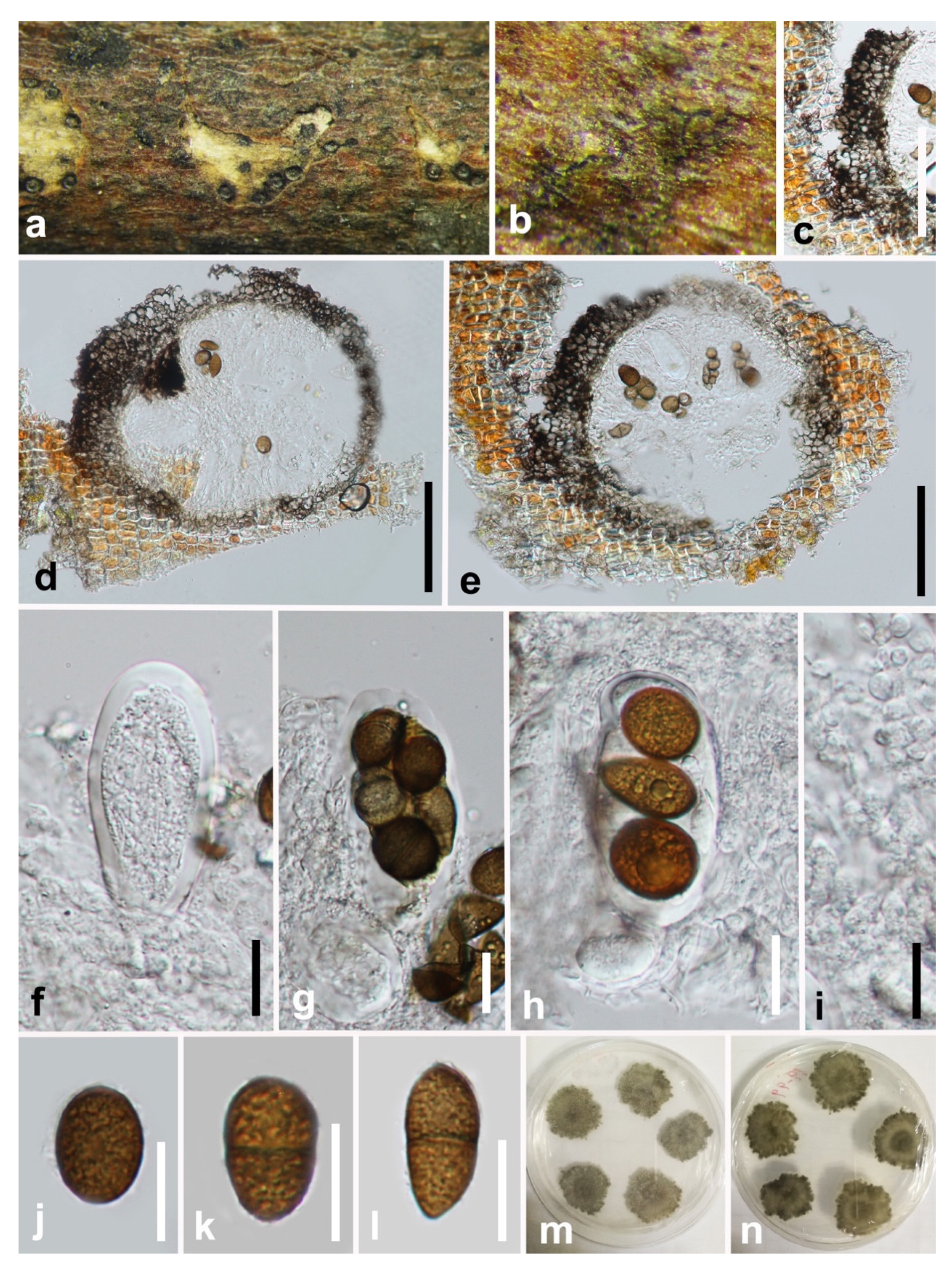







4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, J.K.; Phookamsak, R.; Doilom, M.; Wikee, S.; Li, Y.M. Towards a natural classification of Botryosphaeriales. Fungal Divers. 2012, 57, 149–210. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.L.; Hyde, K.D.; Alves, A.; Liu, J.K. Families in Botryosphaeriales: A phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1–22. [Google Scholar] [CrossRef]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L. Phylogenetic lineages in the Botryosphaeriales: A systematic and evolutionary framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Phillips, A.J.L.; Hyde, K.D.; Li, X.H. Botryosphaeriaceae: Current status of genera and species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, T.; Liu, J.K.; Maharachchikumbura, S.S.N. Outline of Ascomycota-2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar]
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef]
- Wikee, S.; Lombard, L.; Nakashima, C.; Motohashi, K.; Chukeatirote, E. A phylogenetic re-evaluation of Phyllosticta (Botryosphaeriales). Stud. Mycol. 2013, 76, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slippers, B.; Crous, P.W.; Jami, F.; Groenewald, J.Z.; Wingfield, M.J. Diversity in the Botryosphaeriales: Looking back, looking forward. Fungal Biol. 2017, 121, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.Y.; Xie, Y.; Zhang, W.; Wang, Y.; Liu, J.K. Species of Botryosphaeriaceae involved in grapevine dieback in China. Fungal Divers. 2013, 61, 221–236. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Zhang, W.; Liu, M.; Chukeatirote, E.; Yan, J.Y. Lasiodiplodia pseudotheobromae causes pedicel and peduncle discolouration of grapes in China. Australas. Plant Dis. 2015, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Zhang, W.; Li, X.H.; Zhou, Y.; Hyde, K.D. First report of Neofusicoccum mangiferae associated with grapevine dieback in China. Phytopathol. Mediterr. 2015, 54, 414–419. [Google Scholar]
- Chen, Y.Y.; Dissanayake, A.J.; Liu, Z.Y.; Liu, J.K. Additions to Karst Fungi 4: Botryosphaeria spp. associated with woody hosts in Guizhou province, China including B. guttulata sp. nov. Phytotaxa 2020, 454, 186–202. [Google Scholar] [CrossRef]
- Dou, Z.P.; He, W.; Zhang, Y. Lasiodiplodia chinensis, a new holomorphic species from China. Mycosphere 2017, 8, 521–532. [Google Scholar] [CrossRef]
- Liang, L.; Li, H.; Zhou, L.; Chen, F. Lasiodiplodia pseudotheobromae causes stem canker of Chinese hackberry in China. J. For. Res. 2020, 31, 2571–2580. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Impaprasert, R.; Zhao, J.R.; Xu, S.G.; Wu, M. Stem die-back of highbush blueberries caused by Neofusicoccum parvum in China. New Dis. Rep. 2013, 27, 3. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Zhang, H.; Zhou, Z.; Hu, T.; Wang, S. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752. [Google Scholar] [CrossRef]
- Pan, Y.; Ye, H.; Lu, J. Isolation and identification of Sydowia polyspora and its pathogenicity on Pinus yunnanensis in southwestern China. J. Phytopathol. 2019, 166, 386–395. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Tian, C.M.; Fan, X.L. Studies of botryosphaerialean fungi associated with canker and dieback of tree hosts in Dongling Mountain of China. Phytotaxa 2018, 348, 63–76. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R.; Chukeatirote, E. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute & British Mycological Society: Kew, UK, 1970. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Hall, T. Bioedit 7.5.0.3. Department of Microbiology, North Carolina State University. 2006. Available online: http://www.mbio.ncsu.edu/BioEdit/Bioedit.html (accessed on 11 August 2020).
- Katoh, K.; Standley, K. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2653–2677. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Spegazzini, C. Fungi argentini additis nonnullis brasiliensibus montevideensibusque. Pugillus quartus (Continuacion). Anal. Soc. Cient. Argent. 1882, 12, 97–117. [Google Scholar]
- Doilom, M.; Shuttleworth, L.; Roux, J.; Chukeatirote, E.; Hyde, K.D. Barriopsis tectonae sp. nov. a new species of Botryosphaeriaceae from Tectona grandis (teak) in Thailand. Phytotaxa 2014, 176, 081–091. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T. Fungal diversity notes 603–708: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Dissanayake, A.J.; Liu, J.K. Additions to Karst Fungi 5: Sardiniella guizhouensis sp. nov. (Botryosphaeriaceae) associated with woody hosts in Guizhou province, China. Phytotaxa 2021, 508, 187–196. [Google Scholar] [CrossRef]
- Jami, F.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Botryosphaeriaceae species overlap on four unrelated, native South African hosts. Fungal Biol. 2014, 118, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Hyde, K.D.; Liu, J.K.; Liang, Y.M.; Tian, C.M. Multigene phylogeny and morphology reveal Phaeobotryon rhois sp. nov (Botryosphaeriales, Ascomycota). Phytotaxa 2015, 205, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Ekanayaka, A.H.; Dissanayake, A.J.; Jayasiri, S.C.; To-anun, C.; Jones, E.B.G. Aplosporella thailandica; a novel species revealing the sexual-asexual connection in Aplosporellaceae (Botryosphaeriales). Mycosphere 2016, 7, 440–447. [Google Scholar] [CrossRef]
- Du, Z.; Fan, X.L.; Yang, Q.; Hyde, K.D.; Tian, C.M. Aplosporella ginkgonis (Aplosporellaceae, Botryosphaeriales), a new species isolated from Ginkgo biloba in China. Mycosphere 2017, 8, 1246–1252. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Pennycook, S.R.; Johnston, P.R.; Ramaley, A. Resolving the phylogenetic and taxonomic status of dark-spored teleomorph genera in the Botryosphaeriaceae. Persoonia 2008, 21, 29–55. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Mohammadi, G.E.; Javadi, A.; Shams-Bakhsh, M.; Zare, R. Barriopsis iraniana and Phaeobotryon cupressi: Two new species of the Botryosphaeriaceae from trees in Iran. Persoonia 2009, 23, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Konta, S.; Phillips, A.J.L.; Bahkali, A.H.; Jones, E.B.G.; Eungwanichayapant, D.P. Botryosphaeriaceae from palms in Thailand—Barriopsis archontophoenicis sp. nov, from Archontophoenix alexandrae. Mycosphere 2016, 7, 921–932. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Alves, A.; Phillips, A.J.L. Sardiniella urbana gen. et sp. nov., a new member of the Botryosphaeriaceae isolated from declining Celtis australis trees in Sardinian streetscapes. Mycosphere 2016, 7, 893–905. [Google Scholar] [CrossRef]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Morphology, phylogeny and pathogenicity of Botryosphaeria and Neofusicoccum species associated with drupe rot of olives in southern Italy. Plant Pathol. 2008, 57, 948–956. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Scanu, B.; Maddau, L.; Franceschini, A. Diplodia corticola and Phytophthora cinnamomi: The main pathogens involved in holm oak decline on Caprera island (Italy). For. Pathol. 2014, 44, 191–200. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Giambra, S.; Piazza, G.; Alves, A.; Mondello, V.; Berbegal, M. Botryosphaeriaceae species associated with diseased loquat trees in Italy and description of Diplodia rosacearum sp. nov. Mycosphere 2016, 7, 978–989. [Google Scholar] [CrossRef]
- Marsberg, A.; Kemlera, M.; Jami, F.; Nagela, J.H. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Isolate Number | Location | Date | ITS | LSU | tef |
|---|---|---|---|---|---|---|
| Aplosporella hesperidica | GZCC 19-0095 | Fanjing mountain, Tongren District | July 2018 | MZ781423 | MZ781490 | MZ852496 |
| Barriopsis tectonae | GZCC 19-0266 | Maolan natural reserve, Libo District | July 2017 | MZ781424 | MZ781491 | MZ852497 |
| Botryobambusa guizhouensis | CGMCC 3. 20348 | Forest Park, Chishui District | July 2019 | MZ781425 | MZ781492 | MZ852498 |
| GZCC 19-0734 | Forest Park, Chishui District | July 2019 | MZ781426 | MZ781493 | MZ852499 | |
| Botryosphaeria dothidea | GZCC 20-0524 | Huaxi wetland park, Guiyang District | April 2017 | MZ781435 | N/A | MZ852508 |
| GZCC 20-0525 | Xingyi Wanfenglin | June 2019 | MZ781436 | N/A | MZ852509 | |
| GZCC 20-0526 | Xingyi Wanfenglin | June 2019 | MZ781437 | N/A | MZ852510 | |
| GZCC 20-0527 | Suiyang broad water nature reserve | April 2018 | MZ781438 | N/A | MZ852511 | |
| GZCC 20-0528 | Forest Park, Chishui District | July 2019 | MZ781439 | N/A | MZ852512 | |
| GZCC 20-0529 | Suiyang broad water nature reserve | April 2018 | MZ781440 | N/A | MZ852513 | |
| GZCC 20-0530 | Suiyang broad water nature reserve | April 2018 | MZ781441 | N/A | MZ852514 | |
| GZCC 20-0531 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781442 | N/A | MZ852515 | |
| GZCC 20-0532 | Forest Park, Chishui District | July 2019 | MZ781443 | N/A | MZ852516 | |
| GZCC 20-0533 | Huaxi wetland park, Guiyang District | April 2017 | MZ781444 | N/A | MZ852517 | |
| GZCC 20-0534 | Suiyang broad water nature reserve | April 2018 | MZ781445 | N/A | MZ852518 | |
| GZCC 20-0535 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781446 | N/A | MZ852519 | |
| GZCC 20-0536 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781447 | N/A | MZ852520 | |
| GZCC 20-0537 | Huaxi wetland park, Guiyang District | April 2017 | MZ781448 | N/A | MZ852521 | |
| GZCC 20-0538 | Forest Park, Chishui District | July 2019 | MZ781449 | N/A | MZ852522 | |
| GZCC 20-0539 | Forest Park, Chishui District | July 2019 | MZ781450 | N/A | MZ852523 | |
| GZCC 20-0540 | Huaxi wetland park, Guiyang District | April 2017 | MZ781451 | N/A | MZ852524 | |
| GZCC 20-0541 | Forest Park, Chishui District | July 2019 | MZ781452 | N/A | MZ852525 | |
| GZCC 20-0542 | Forest Park, Chishui District | July 2019 | MZ781453 | N/A | MZ852526 | |
| GZCC 20-0543 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781454 | N/A | MZ852527 | |
| GZCC 20-0544 | Suiyang broad water nature reserve | April 2018 | MZ781455 | N/A | MZ852528 | |
| GZCC 20-0545 | Xingyi Wanfenglin | June 2019 | MZ781456 | N/A | MZ852529 | |
| GZCC 20-0546 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781457 | N/A | MZ852530 | |
| Diplodia mutila | GZCC 20-0547 | Xingyi Wanfenglin | June 2019 | MZ781459 | N/A | MZ852531 |
| GZCC 20-0548 | Huaxi wetland park, Guiyang District | April 2017 | MZ781460 | N/A | MZ852532 | |
| Di. neojuniperi | GZCC 19-0191 | Maolan natural reserve, Libo District | July 2017 | MZ781463 | N/A | MZ852533 |
| Di. pseudoseriata | GZCC 19-0072 | Xingyi Wanfenglin | June 2019 | MZ781461 | N/A | MZ852534 |
| Di. sapinea | GZCC 19-0075 | Suiyang broad water nature reserve | April 2018 | MZ781462 | N/A | MZ852535 |
| Di. seriata | GZCC 19-0548 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781458 | N/A | MZ852536 |
| Dothiorella sarmentorum | GZCC 19-0092 | Xingyi Wanfenglin | June 2019 | MZ781464 | N/A | MZ852537 |
| Do. yunnana | GZCC 19-0175 | Suiyang broad water nature reserve | April 2018 | MZ781465 | N/A | MZ852538 |
| Lasiodiplodia pseudotheobromae | GZCC 19-0162 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781466 | N/A | MZ852539 |
| GZCC 19-0184 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781467 | N/A | MZ852540 | |
| GZCC 19-0227 | Huaxi wetland park, Guiyang District | April 2017 | MZ781468 | N/A | MZ852541 | |
| GZCC 20-1575 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781469 | N/A | MZ852542 | |
| GZCC 19-0063 | Suiyang broad water nature reserve | April 2018 | MZ781470 | N/A | MZ852543 | |
| Neofusicoccum parvum | GZCC 19-0123 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781471 | N/A | MZ852544 |
| GZCC 19-0198 | Xingyi Wanfenglin | June 2019 | MZ781472 | N/A | MZ852545 | |
| GZCC 19-0218 | Xingyi Wanfenglin | June 2019 | MZ781473 | N/A | MZ852546 | |
| GZCC 19-2013 | Maolan natural reserve, Libo District | July 2017 | MZ781474 | N/A | MZ852547 | |
| GZCC 20-0549 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781475 | N/A | MZ852548 | |
| GZCC 20-0550 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781476 | N/A | MZ852549 | |
| GZCC 20-0551 | Xingyi Wanfenglin | June 2019 | MZ781477 | N/A | MZ852550 | |
| GZCC 20-0552 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781478 | N/A | MZ852551 | |
| GZCC 20-0553 | Huaxi wetland park, Guiyang District | April 2017 | MZ781479 | N/A | MZ852552 | |
| GZCC 20-0554 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781480 | N/A | MZ852553 | |
| GZCC 20-0555 | Suiyang broad water nature reserve | April 2018 | MZ781481 | N/A | MZ852554 | |
| GZCC 20-0556 | Xiaochehe wetland park, Guiyang District | May 2017 | MZ781482 | N/A | MZ852555 | |
| Sardiniella celtidis | GZCC 19-0248 | Xingyi Wanfenglin | June 2019 | MZ781427 | MZ781494 | MZ852500 |
| Sa. elliptica | CGMCC 3.20349 | Huaxi wetland park, Guiyang District | April 2017 | MZ781429 | MZ781496 | MZ852502 |
| GZCC 19-0245 | Xingyi Wanfenglin | June 2019 | MZ781431 | MZ781498 | MZ852504 | |
| GZCC 19-0262 | Maolan natural reserve, Libo District | July 2017 | MZ781430 | MZ781497 | MZ852503 | |
| Sa. guizhouensis | GZCC 19-0229 | Maolan natural reserve, Libo District | July 2017 | MZ781428 | MZ781495 | MZ852501 |
| Sphaeropsis citrigena | GZCC 20-0557 | Xingyi Wanfenglin | June 2019 | MZ781432 | MZ781499 | MZ852505 |
| Sp. guizhouensis | CGMCC 3.20352 | Maolan natural reserve, Libo District | July 2017 | MZ781433 | MZ781500 | MZ852506 |
| GZCC 19-0273 | Xingyi Wanfenglin | June 2019 | MZ781434 | MZ781501 | MZ852507 |
| Gene Region | Primers | Sequence 5′-3′ | Optimized PCR Protocols | References |
|---|---|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGG | 94 °C: 5 min, (94 °C: 30 s, 55 °C: 50 s, 72 °C: 1 min) × 34 cycles 72 °C: 10 min | White et al. [26] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| LSU | LR0R | ACCCGCTGAACTTAAGC | 94 °C: 5 min, (94 °C: 30 s, 54 °C: 50 s, 72 °C: 1 min) × 34 cycles 72 °C: 10 min | Vilgalys and Hester [27] |
| LR5 | TCCTGAGGGAAACTTCG | |||
| tef | EF1-728F | CATCGAGAAGTTCGAGAAGG | 95 °C: 5 min, (95 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 34 cycles 72 °C: 10 min | Carbone and Kohn [28] |
| EF1-986R | TACTTGAAGGAACCCTTACC |
| Species Name | Isolate Number | ITS | LSU | tef |
|---|---|---|---|---|
| Aplosporellaceae | ||||
| Alanomyces indica | CBS 134264 | HF563622 | HF563623 | AB872219 |
| Aplosporella africana | CBS 121777 | KF766196 | EU101380 | EU101360 |
| A. africana | CBS 121778 | EU101316 | EU101381 | EU101361 |
| A. artocarpi | CPC 22791 | KM006450 | N/A | KM006481 |
| A. ginkgonis | CFCC 52442 | MH133916 | MH133933 | MH133950 |
| A. ginkgonis | CFCC 89661 | KM030583 | KM030590 | KM030597 |
| A. hesperidica | CBS 732.79 | KX464083 | KX464239 | N/A |
| A. hesperidica | CBS 208.37 | JX681069 | MH867398 | N/A |
| A. javeedii | CFCC 50054 | KP208840 | KP208843 | KP208846 |
| A. javeedii | CFCC 50052 | KP208838 | KP208841 | KP208844 |
| A. macropycnidia | CGMCC 3.17725 | KT343648 | N/A | KX011176 |
| A. macropycnidia | CGMCC 3.17726 | KT343649 | N/A | KX011177 |
| A. papillata | CBS 121780 | EU101328 | EU101383 | EU101373 |
| A. papillata | CBS 121781 | EU101329 | EU101384 | EU101374 |
| A. prunicola | CBS 121167 | KF766147 | KF766315 | N/A |
| A. prunicola | STE-U 6326 | EF564375 | EF564377 | N/A |
| A. sophorae | CPC 29688 | KY173388 | KY173482 | N/A |
| A. thailandica | MFLU 16-0615 | KX423536 | N/A | KX423537 |
| A. yalgorensis | MUCC511 | EF591926 | EF591943 | EF591977 |
| A. yalgorensis | MUCC512 | EF591927 | EF591944 | EF591978 |
| Botryosphaeriaceae | ||||
| Alanphillipsia aloeicola | CBS 138896 | KP004444 | KP004472 | N/A |
| A. euphorbiae | CPC 21628 | KF777140 | KF777196 | N/A |
| Barriopsis fusca | CBS 174.26 | EU673330 | DQ377857 | EU673296 |
| Ba. iraniana | IRAN1448C | FJ919663 | KF766318 | FJ919652 |
| Ba. iraniana | IRAN1449C | FJ919665 | N/A | FJ919654 |
| Ba. tectonae | MFLUCC 12-0381 | KJ556515 | N/A | KJ556516 |
| Botryobambusa fusicoccum | MFLUCC 11-0143 | JX646792 | JX646809 | JX646857 |
| Bo. fusicoccum | MFLUCC 11-0657 | JX646793 | JX646810 | JX646858 |
| Botryosphaeria agaves | MFLUCC 11-0125 | JX646791 | JX646808 | JX646856 |
| B. agaves | MFLUCC 10-0051 | JX646790 | JX646807 | JX646855 |
| B. corticis | CBS 119047 | DQ299245 | EU673244 | EU017539 |
| B. corticis | ATCC 22927 | DQ299247 | EU673245 | EU673291 |
| B. dothidea | CMW8000 | AY236949 | DQ377852 | AY236898 |
| B. dothidea | CBS 110302 | AY259092 | EU673243 | AY573218 |
| B. dothidea | CMW 25413 | KF766167 | KF766332 | EU101348 |
| B. dothidea | GZCC 16-0013 | KX447675 | N/A | KX447678 |
| B. dothidea | GZCC 16-0014 | KX447676 | N/A | KX447679 |
| B. dothidea | CGMCC 3.17722 | KT343254 | N/A | KU221233 |
| B. dothidea | CGMCC 3.17724 | KT343256 | N/A | KU221234 |
| B. dothidea | CGMCC 3.18744 | KX278002 | N/A | KX278107 |
| B. dothidea | CGMCC 3.18745 | KX278003 | N/A | KX278108 |
| B. dothidea | CGMCC3.20094 | MT327839 | N/A | MT331606 |
| B. dothidea | GZCC 19-0186 | MT327832 | N/A | MT331600 |
| B. dothidea | GZCC 19-0188 | MT327833 | N/A | MT331601 |
| B. fabicerciana | CMW27094 | HQ332197 | N/A | HQ332213 |
| B. fabicerciana | CMW27108 | HQ332200 | N/A | HQ332216 |
| B. fabicerciana | MFLUCC 10-0098 | JX646789 | JX646806 | JX646854 |
| B. fabicerciana | MFLUCC 11-0507 | JX646788 | JX646805 | JX646853 |
| B. kuwatsukai | CBS 135219 | KJ433388 | N/A | KJ433410 |
| B. kuwatsukai | LSP5 | KJ433395 | N/A | KJ433417 |
| B. kuwatsukai | CGMCC 3.18007 | KX197074 | N/A | KX197094 |
| B. kuwatsukai | CGMCC 3.18008 | KX197075 | N/A | KX197095 |
| B. qingyuanensis | CGMCC 3.18742 | KX278000 | N/A | KX278105 |
| B. qingyuanensis | CGMCC 3.18743 | KX278001 | N/A | KX278106 |
| B. ramosa | CBS 122069 | EU144055 | N/A | EU144070 |
| B. ramosa | CGMCC 3.18739 | KX277988 | N/A | KX278093 |
| B. ramosa | CGMCC 3.18740 | KX277989 | N/A | KX278094 |
| B. scharifii | IRAN1529C | JQ772020 | N/A | JQ772057 |
| B. scharifii | IRAN1543C | JQ772019 | N/A | JQ772056 |
| Cophinforma eucalypti | MFLUCC 11-0655 | JX646801 | JX646818 | JX646866 |
| C. mamane | CBS 117444 | KF531822 | DQ377855 | KF531801 |
| Diplodia africana | CBS 120835 | EF445343 | N/A | EF445382 |
| Di. africana | CBS 121104 | EF445344 | N/A | EF445383 |
| Di. agrifolia | CBS 132777 | JN693507 | N/A | JQ517317 |
| Di. agrifolia | UCROK1429 | JQ411412 | N/A | JQ512121 |
| Di. allocellula | CBS 130408 | JQ239397 | JQ239410 | JQ239384 |
| Di. allocellula | CBS 130410 | JQ239399 | JQ239412 | JQ239386 |
| Di. arengae | MFLU 17-2769 | MG762771 | N/A | MG762774 |
| Di. bulgarica | CBS 124135 | GQ923853 | N/A | GQ923821 |
| Di. bulgarica | CBS 124254 | GQ923852 | N/A | GQ923820 |
| Di. corticola | CBS 112546 | AY259100 | AY928051 | AY573227 |
| Di. corticola | CBS 112549 | AY259110 | EU673262 | DQ458872 |
| Di. crataegicola | MFLU 15-1311 | KT290244 | N/A | KT290248 |
| Di. cupressi | CBS 168.87 | DQ458893 | EU673263 | DQ458878 |
| Di. cupressi | CBS 261.85 | DQ458894 | EU673264 | DQ458879 |
| Di. eriobotryicola | BN 21 | MT587342 | N/A | MT592047 |
| Di. estuarina | CMW41231 | KP860831 | N/A | KP860676 |
| Di. estuarina | CMW41230 | KP860830 | N/A | KP860675 |
| Di. fraxini | CBS 136010 | KF307700 | N/A | KF318747 |
| Di. galiicola | MFLU 15-1310 | KT290245 | N/A | KT290249 |
| Di. gallae | CBS 213.25 | KX464092 | N/A | KX464566 |
| Di. malorum | CBS 124130 | GQ923865 | N/A | GQ923833 |
| Di. malorum | CBS 112554 | AY259095 | N/A | DQ458870 |
| Di. mutila | CBS 112553 | AY259093 | AY928049 | AY573219 |
| Di. mutila | CBS 230.30 | DQ458886 | AY928049 | DQ458869 |
| Di. mutila | CBS 121862 | KX464093 | N/A | KX464567 |
| Di. neojuniperi | CPC22753 | KM006431 | N/A | KM006462 |
| Di. olivarum | CBS 121887 | EU392302 | N/A | EU392279 |
| Di. olivarum | CBS 121886 | EU392297 | N/A | EU392274 |
| Di. pseudoseriata | CBS 124906 | EU080927 | MH874931 | EU863181 |
| Di. pseudoseriata | CBS 124907 | EU080922 | N/A | EU863179 |
| Di. pseudoseriata | CBS 124931 | FJ888460 | MH874935 | FJ888444 |
| Di. pseudoseriata | CBS 124933 | FJ888478 | N/A | FJ888446 |
| Di. pseudoseriata | CBS 140350 | KX833072 | N/A | KX833073 |
| Di. quercivora | CBS 133852 | JX894205 | N/A | JX894229 |
| Di. rosulata | CBS 116470 | EU430265 | DQ377896 | EU430267 |
| Di. rosulata | CBS 116472 | EU430266 | DQ377897 | EU430268 |
| Di. sapinea | CBS 393.84 | DQ458895 | DQ377893 | DQ458880 |
| Di. sapinea | CBS 109725 | DQ458896 | EU673270 | DQ458881 |
| Di. sapinea | CBS 124462 | GQ923858 | MH874896 | GQ923826 |
| Di. sapinea | CBS 124134 | HM036528 | N/A | GQ923851 |
| Di. sapinea | CBS 141915 | KT956270 | N/A | KU378605 |
| Di. scrobiculata | CBS 118110 | AY253292 | KF766326 | AY624253 |
| Di. scrobiculata | CBS 109944 | DQ458899 | EU673268 | DQ458884 |
| Di. scrobiculata | CBS 113423 | DQ458900 | EU673267 | DQ458885 |
| Di. seriata | CBS 112555 | AY259094 | AY928050 | AY573220 |
| Di. seriata | CBS 119049 | DQ458889 | EU673266 | DQ458874 |
| Di. subglobosa | CBS 124133 | GQ923856 | N/A | GQ923824 |
| Di. tsugae | CBS 418.64 | DQ458888 | DQ377867 | DQ458873 |
| Dothiorella acacicola | CPC26349 | KX228269 | KX228320 | KX228376 |
| Do. acericola | KUMCC 18-0137 | KY385661 | N/A | KY393212 |
| Do. alpina | CGMCC 3.18001 | KX499645 | N/A | KX499651 |
| Do. brevicollis | CBS 130411 | JQ239403 | JQ239416 | JQ239390 |
| Do. capri-amissi | CBS 121763 | EU101323 | N/A | EU101368 |
| Do. capri-amissi | CMW25404 | EU101324 | N/A | EU101369 |
| Do. casuarinae | CBS 120688 | DQ846773 | N/A | DQ875331 |
| Do. casuarinae | CBS 120690 | DQ846774 | N/A | DQ875333 |
| Do. citricola | ICMP 16828 | EU673323 | N/A | EU673290 |
| Do. citricola | ICMP 16827 | EU673322 | N/A | EU673289 |
| Do. dulcispinae | CBS 130413 | JQ239400 | JQ239413 | JQ239387 |
| Do. dulcispinae | CBS 121765 | EU101300 | KX464317 | EU101345 |
| Do. iranica | IRAN1587C | KC898231 | N/A | KC898214 |
| Do. juglandis | CBS 188.87 | EU673316 | DQ377891 | EU673283 |
| Do. lampangensis | MFLUCC 18-0232 | MK347758 | N/A | MK340869 |
| Do. longicollis | CBS 122068 | EU144054 | MH874718 | EU144069 |
| Do. longicollis | CBS 122066 | EU144052 | KX464311 | EU144067 |
| Do. magnoliae | CFCC 51563 | KY111248 | N/A | KY213687 |
| Do. mangifericola | IRAN1584C | MT587407 | N/A | MT592119 |
| Do. mangifericola | IRAN1545C | KC898221 | N/A | KX464614 |
| Do. mangifericola | CBS 121760 | KF766227 | N/A | EU101335 |
| Do. mangifericola | CBS 121761 | EU101293 | N/A | EU101338 |
| Do. moneti | MUCC 505 | EF591920 | EF591937 | EF591971 |
| Do. moneti | MUCC 507 | EF591922 | EF591939 | EF591973 |
| Do. plurivora | IRAN1557C | KC898225 | N/A | KC898208 |
| Do. plurivora | IRAN1537C | KC898226 | N/A | KC898209 |
| Do. pretoriensis | CBS 130404 | JQ239405 | JQ239418 | JQ239392 |
| Do. pretoriensis | CBS 130403 | JQ239406 | JQ239419 | JQ239393 |
| Do. prunicola | IRAN1541 | EU673313 | EU673232 | EU673280 |
| Do. rhamni | MFLUCC 14-0902 | KU246381 | KU246382 | N/A |
| Do. rhamni | BN 81 | MT587399 | N/A | MT592111 |
| Do. santali | MUCC 509 | EF591924 | EF591941 | EF591975 |
| Do. santali | MUCC 508 | EF591923 | EF591940 | EF591974 |
| Do. sarmentorum | IMI63581b | AY573212 | AY928052 | AY573235 |
| Do. sarmentorum | CBS 115038 | AY573206 | DQ377860 | AY573223 |
| Do. sarmentorum | CBS 128309 | HQ288218 | MH876298 | HQ288262 |
| Do. sarmentorum | CBS 128310 | HQ288219 | MH876299 | HQ288263 |
| Do. sarmentorum | CBS 141587 | KX357188 | N/A | KX357211 |
| Do. sarmentorum | MFLUCC 17-0242 | KY797637 | KY815014 | KY815020 |
| Do. sarmentorum | CBS 115045 | AY573202 | AY928053 | AY573222 |
| Do. sarmentorum | CBS 113188 | AY573198 | EU673230 | EU673278 |
| Do. sarmentorum | MFLUCC 17-0951 | MF398891 | N/A | MF398943 |
| Do. sarmentorum | CBS 140349 | KP205497 | N/A | KP205470 |
| Do. sarmentorum | CBS 188.87 | EU673316 | DQ377891 | EU673283 |
| Do. sarmentorum | IRAN1579C | KC898234 | N/A | KC898217 |
| Do. sarmentorum | IRAN1585C | KC898235 | N/A | KC898218 |
| Do. sarmentorum | IRAN1583C | KC898236 | N/A | KC898219 |
| Do. sarmentorum | IRAN1581C | KC898237 | N/A | KC898220 |
| Do. sarmentorum | MFLUCC 13-0497 | KJ742378 | N/A | KJ742381 |
| Do. sarmentorum | MFLUCC 13-0196 | KU234782 | N/A | KU234796 |
| Do. sarmentorum | DAR78992 | EU768874 | N/A | EU768881 |
| Do. sarmentorum | DAR78993 | EU768876 | N/A | EU768882 |
| Do. striata | ICMP 16824 | EU673320 | EU673240 | EU673287 |
| Do. striata | ICMP 16819 | EU673320 | EU673239 | EU673287 |
| Do. striata | DAR 80992 | KJ573643 | N/A | KJ573640 |
| Do. styphnolobii | JZB3150013 | MH880849 | N/A | MK069594 |
| Do. tectonae | MFLUCC 12-0382 | KM396899 | N/A | KM409637 |
| Do. thailandica | MFLUCC 11-0438 | JX646796 | JX646813 | JX646861 |
| Do. thripsita | BRIP 51876 | FJ824738 | N/A | KJ573639 |
| Do. ulmacea | CBS 138855 | KR611881 | KR611899 | KR611910 |
| Do. uruguayensis | CBS 124908 | EU080923 | MH874932 | EU863180 |
| Do. vinea gemmae | DAR81012 | KJ573644 | N/A | KJ573641 |
| Do. viticola | CBS 117009 | AY905554 | MH874565 | AY905559 |
| Do. viticola | CBS 117006 | AY905555 | EU673236 | AY905562 |
| Do. viticola | DAR80529 | HM009376 | N/A | HM800511 |
| Do. viticola | DAR80530 | HM009378 | N/A | HM800513 |
| Do. yunnana | CGMCC 3.17999 | KX499643 | N/A | KX499649 |
| Endomelanconiopsis endophytica | CBS 120397 | EU683656 | EU683629 | EU683637 |
| E. microspora | CBS 353.97 | EU683655 | KF766330 | EU683636 |
| Lasiodiplodia americana | CERC1960 | KP217058 | N/A | KP217066 |
| L. americana | CERC1961 | KP217059 | N/A | KP217067 |
| L. avicenniae | CMW41467 | KP860835 | N/A | KP860680 |
| L. avicenniarum | MFLUCC 17-2591 | MK347777 | MK347994 | MK340867 |
| L. brasiliense | CMM4015 | JX464063 | N/A | JX464049 |
| L. brasiliense | CMM2185 | KC484800 | N/A | KC481530 |
| L. bruguierae | CMW42480 | KP860832 | N/A | KP860677 |
| L. chonburiensis | MFLUCC 16-0376 | N/A | ||
| L. cinnamomi | CFCC 51997 | MG866028 | N/A | MH236799 |
| L. citricola | CBS 124707 | GU945354 | N/A | GU945340 |
| L. citricola | IRAN1521C | GU945353 | N/A | GU945339 |
| L. crassispora | CBS 118741 | DQ103550 | DQ377901 | EU673303 |
| L. crassispora | WAC12534 | DQ103551 | N/A | DQ103558 |
| L. crassispora | CBS 110492 | EF622086 | EU673251 | EF622066 |
| L. crassispora | CBS 121770 | EU101307 | N/A | EU101352 |
| L. crassispora | CBS 121771 | EU101308 | N/A | EU101353 |
| L. euphorbicola | CMM3609 | KF234543 | N/A | KF226689 |
| L. euphorbicola | IBL329 | KT247490 | N/A | KT247492 |
| L. gilanensis | CBS 124704 | GU945351 | N/A | GU945342 |
| L. gilanensis | IRAN1501C | GU945352 | N/A | GU945341 |
| L. gilanensis | CBS 128311 | HQ288225 | N/A | HQ288267 |
| L. gilanensis | UCD2199MO | HQ288226 | N/A | HQ288268 |
| L. gonubiensis | CBS 115812 | AY639595 | DQ377902 | DQ103566 |
| L. gonubiensis | CBS 116355 | AY639594 | EU673252 | DQ103567 |
| L. gravistriata | CMM4564 | KT250949 | N/A | KT250950 |
| L. hormozganensis | CBS 124709 | GU945355 | N/A | GU945343 |
| L. hormozganensis | CBS 124708 | GU945356 | N/A | GU945344 |
| L. iraniensis | CBS 124710 | GU945346 | MH874918 | GU945334 |
| L. iraniensis | CBS 124711 | GU945347 | N/A | GU945335 |
| L. iraniensis | CMM3610 | MT587430 | N/A | MT592142 |
| L. iraniensis | CMM0247 | MT587431 | N/A | MT592143 |
| L. laeliocattleyae | CBS 167.28 | KU507487 | N/A | KU507454 |
| L. laeliocattleyae | CBS 130992 | JN814397 | N/A | JN814424 |
| L. laeliocattleyae | CMM3611 | JN814401 | N/A | JN814428 |
| L. lignicola | CBS 134112 | JX646797 | N/A | KU887003 |
| L. lignicola | MFLUCC 11-0656 | JX646797 | JX646815 | KU887003 |
| L. lignicola | CGMCC 3.18061 | KX499889 | N/A | KX499927 |
| L. lignicola | CBS 342.78 | KX464140 | N/A | KX464634 |
| L. macrospora | CMM3833 | KF234557 | N/A | KF226718 |
| L. mahajangana | CBS 124927 | FJ900597 | N/A | FJ900643 |
| L. mahajangana | CBS 124925 | FJ900595 | N/A | FJ900641 |
| L. mahajangana | IBL352 | KT154759 | N/A | KT154753 |
| L. mahajangana | CMM1325 | KT154760 | N/A | KT008006 |
| L. mahajangana | CBS 137785 | KJ638317 | N/A | KJ638336 |
| L. mahajangana | BL184 | KJ638318 | N/A | KJ638337 |
| L. mahajangana | MFLUCC 16-0265 | MH275068 | MH260301 | MH412774 |
| L. margaritacea | CBS 122519 | EU144050 | N/A | EU144065 |
| L. margaritacea | CBS 122065 | EU144051 | N/A | EU144066 |
| L. marypalme | CMM2275 | KC484843 | N/A | KC481567 |
| L. marypalme | CMM2271 | KC484844 | N/A | KC481568 |
| L. mediterranea | CBS 137783 | KJ638312 | N/A | KJ638331 |
| L. mediterranea | ALG36 | KJ638314 | N/A | KJ638333 |
| L. parva | CBS 456.78 | EF622083 | KF766362 | EF622063 |
| L. parva | CBS 494.78 | EF622084 | EU673258 | EF622064 |
| L. parva | CBS 356.59 | AY343482 | N/A | EF445396 |
| L. plurivora | CBS 120832 | EF445362 | KX464356 | EF445395 |
| L. plurivora | CBS 121103 | AY343482 | KX464357 | EF445396 |
| L. pontae | CMM1277 | KT151794 | N/A | KT151791 |
| L. pontae | IBL14 | KT151794 | N/A | KT151791 |
| L. pseudotheobromae | CBS 116459 | EF622077 | EU673256 | EF622057 |
| L. pseudotheobromae | CBS 447.62 | EF622081 | MH869806 | EF622060 |
| L. rubropurpurea | CBS 118740 | DQ103553 | DQ377903 | EU673304 |
| L. rubropurpurea | WAC12536 | DQ103554 | N/A | DQ103572 |
| L. subglobosa | CMM3872 | KF234558 | N/A | KF226721 |
| L. subglobosa | CMM4046 | KF234560 | N/A | KF226723 |
| L. thailandica | CPC22795 | KJ193637 | N/A | KJ193681 |
| L. thailandica | CGMCC 3.17975 | KX499879 | MG321677 | KX499917 |
| L. thailandica | MFLUCC 18-0244 | MK347789 | N/A | MK340870 |
| L. theobromae | CBS 164.96 | AY640255 | EU673253 | AY640258 |
| L. theobromae | CBS 124.13 | DQ458890 | AY928054 | DQ458875 |
| L. theobromae | CBS 111530 | EF622074 | N/A | EF622054 |
| L. theobromae | CAA006 | DQ458891 | EU673254 | DQ458876 |
| L. theobromae | CBS 164.96 | AY640255 | EU673253 | AY640258 |
| L. venezuelensis | CBS 118739 | DQ103547 | DQ377904 | EU673305 |
| L. venezuelensis | WAC12540 | DQ103548 | N/A | DQ103569 |
| L. viticola | CBS 128313 | HQ288227 | KX098286 | HQ288269 |
| L. viticola | CBS 128315 | HQ288228 | N/A | HQ288270 |
| L. vitis | CBS 124060 | KX464148 | N/A | MN938928 |
| Lecanosticta acicola | LNPV252 | JX901755 | JX901844 | JX901639 |
| Macrophomina phaseolina | CBS 227.33 | KF951627 | DQ377906 | KF952000 |
| Neodeightonia palmicola | MFLUCC 10-0822 | HQ199221 | HQ199222 | N/A |
| N. phoenicum | CBS 122528 | EU673340 | EU673261 | EU673309 |
| N. subglobosa | CBS 448.91 | EU673337 | DQ377866 | EU673306 |
| Neofusicoccum arbuti | CBS 116131 | AY819720 | DQ377915 | KF531792 |
| N. arbuti | UW13 | AY819724 | N/A | KF531791 |
| N. arbuti | CBS 117453 | AY693976 | DQ377914 | AY693977 |
| N. arbuti | CMW13446 | DQ306263 | N/A | DQ306264 |
| N. australe | CMW6837 | AY339262 | KF766367 | AY339270 |
| N. australe | CMW6853 | AY339263 | N/A | AY339271 |
| N. brasiliense | CMM1338 | JX513630 | N/A | JX513610 |
| N. buxi | CBS 116.75 | KX464164 | KX464406 | KX464677 |
| N. cordaticola | CBS 123634 | EU821898 | MH874849 | EU821868 |
| N. cordaticola | CBS 123635 | EU821903 | KX464410 | EU821873 |
| N. cryptoaustrale | CMW23785 | FJ752742 | N/A | FJ752713 |
| N. eucalypticola | CBS 115679 | AY615141 | N/A | AY615133 |
| N. eucalypticola | CBS 115766 | AY615143 | N/A | AY615135 |
| N. eucalyptorum | CBS 115791 | AF283686 | N/A | AY236891 |
| N. eucalyptorum | CMW10126 | AF283687 | N/A | AY236892 |
| N. grevilleae | CBS 129518 | JF951137 | N/A | N/A |
| N. hellenicum | CERC1947 | KP217053 | N/A | KP217061 |
| N. hongkongensis | CERC 2973 | KX278052 | MF410096 | KX278157 |
| N. illicii | CGMCC 3.18311 | KY350150 | N/A | KY817756 |
| N. kwambonambiense | EU821900 | EU821900 | N/A | EU821870 |
| N. kwambonambiense | CMW14140 | EU821919 | N/A | EU821889 |
| N. lumnitzerae | CMW41469 | KP860881 | N/A | KP860724 |
| N. luteum | CBS 110299 | AY259091 | AY928043 | AY573217 |
| N. luteum | CBS 110497 | EU673311 | N/A | EU673277 |
| N. luteum | CMW41365 | KP860859 | N/A | KP860702 |
| N. macroclavatum | CBS 118223 | DQ093196 | N/A | DQ093217 |
| N. macroclavatum | WAC12446 | DQ093197 | N/A | DQ093218 |
| N. mangiferae | CBS 118532 | AY615185 | DQ377921 | DQ093221 |
| N. mangiferae | CMW7797 | AY615186 | N/A | DQ093220 |
| N. mediterraneum | CBS 121718 | GU251176 | MH874696 | GU251308 |
| N. mediterraneum | CBS 121558 | GU799463 | N/A | GU799462 |
| N. mediterraneum | CBS 113083 | KX464186 | KX464465 | KX464712 |
| N. mediterraneum | CBS 113089 | KX464199 | N/A | KX464727 |
| N. microconidium | CERC 3497 | KX278053 | MF410097 | KX278158 |
| N. nonquaesitum | CBS 126655 | GU251163 | MH875645 | GU251295 |
| N. nonquaesitum | PD301 | GU251164 | N/A | GU251296 |
| N. occulatum | CBS 128008 | EU301030 | MH876179 | EU339509 |
| N. occulatum | MUCC286 | EU736947 | N/A | EU339511 |
| N. parvum | CMW9081 | AY236943 | AY928045 | AY236888 |
| N. parvum | CBS 110301 | AY259098 | AY928046 | AY573221 |
| N. parvum | CBS 137504 | KJ657702 | N/A | KJ657715 |
| N. parvum | ALG9 | KJ657704 | N/A | KJ657721 |
| N. parvum | MFLUCC 15-0900 | KY856755 | N/A | KY856754 |
| N. pennatisporum | WAC13153 | EF591925 | EF591942 | EF591976 |
| N. pistaciae | CBS 595.76 | KX464163 | KX464404 | KX464676 |
| N. protearum | STE-U 4361 | AF196295 | N/A | N/A |
| N. protearum | STE-U 1775 | AF452539 | N/A | N/A |
| N. ribis | CMW7772 | AY236935 | N/A | AY236877 |
| N. ribis | CBS 121.26 | AF241177 | N/A | AY236879 |
| N. ribis | CBS 124923 | FJ900607 | N/A | FJ900653 |
| N. ribis | CBS 124924 | FJ900608 | N/A | FJ900654 |
| N. ribis | CMW14058 | EU821904 | N/A | EU821874 |
| N. ribis | CMW14060 | EU821905 | N/A | EU821875 |
| N. sinense | CGMCC 3.18315 | KY350148 | N/A | KY817755 |
| N. sinoeucalypti | CERC 2005 | KX278061 | MF410105 | KX278166 |
| N. stellenboschiana | CBS 110864 | AY343407 | KX464513 | AY343348 |
| N. terminaliae | CBS 125264 | GQ471802 | N/A | GQ471780 |
| N. ursorum | CBS 122811 | FJ752746 | MH874765 | FJ752709 |
| N. viticlavatum | CBS 112878 | AY343381 | MH874474 | AY343342 |
| N. viticlavatum | CBS 112977 | AY343380 | KX464528 | AY343341 |
| N. vitifusiforme | CBS 110887 | AY343383 | MH874455 | AY343343 |
| N. vitifusiforme | CBS 110880 | AY343382 | KX464475 | AY343344 |
| N. vitifusiforme | CBS 120081 | DQ923533 | MN162190 | KX464682 |
| N. vitifusiforme | CBS 121112 | EF445349 | N/A | EF445391 |
| Oblongocollomyces variabile | CBS 121774 | NR136994 | KX464536 | EU101357 |
| O. variabile | CMW25420 | EU101313 | N/A | EU101358 |
| O. variabile | CMW25421 | EU101314 | N/A | EU101359 |
| Phaeobotryon mamane | CPC 12264 | EU673331 | DQ377898 | EU673297 |
| P. mamane | CBS 122980 | EU673332 | EU673248 | EU673298 |
| Sardiniella celtidis | MFLUCC 17-981 | MF443249 | N/A | MF443248 |
| Sa. urbana | BL180 | KX379677 | KX379679 | KX379678 |
| Sa. urbana | BL181 | KX379680 | KX379682 | KX379681 |
| Sa. urbana | CBS 141580 | KX379674 | KX379676 | KX379675 |
| Sphaeropsis citrigena | ICMP16812 | EU673328 | EU673246 | EU673294 |
| Sp. citrigena | ICMP16818 | EU673329 | EU673247 | EU673295 |
| Sp. eucalypticola | MFLUCC 11-0579 | JX646802 | JX646819 | JX646867 |
| Sp. eucalypticola | MFLUCC 11-0654 | JX646803 | JX646820 | JX646868 |
| Sp. porosa | CBS 110496 | AY343379 | DQ377894 | AY343340 |
| Sp. porosa | CBS 110574 | AY343378 | DQ377895 | AY343339 |
| Sp. visci | CBS 100163 | EU673324 | N/A | EU673292 |
| Sp. visci | CBS 186.97 | EU673325 | DQ377868 | EU673293 |
| Tiarosporella graminis-karoo | CBS 118718 | KF531828 | DQ377939 | KF531807 |
| Melanopsaceae | ||||
| Melanops sp. | CBS 118.39 | FJ824771 | DQ377856 | FJ824776 |
| M. tulasnei | CBS 116805 | FJ824769 | KF766365 | FJ824774 |
| M. tulasnei | CBS 116806 | FJ824770 | FJ824765 | KX464644 |
| Phyllostictaceae | ||||
| Phyllosticta citricarpa | CBS 102374 | FJ538313 | DQ377877 | FJ538376 |
| P. minima | CBS 111635 | KF766215 | EU754194 | KF766433 |
| P. parthenocissi | CBS 111645 | EU683672 | DQ377876 | EU683653 |
| P. podocarpi | CBS 111647 | KF766217 | KF766383 | KF766434 |
| Pseudofusicoccum adansoniae | CMW 26147 | KF766220 | KF766386 | N/A |
| P. ardesiacum | CMW 26159 | EU144060 | KF766387 | N/A |
| P. kimberleyensis | CMW 26156 | EU144057 | KF766388 | N/A |
| Planistromellaceae | ||||
| Kellermania agaves | CPC 21713 | KF777164 | KF777217 | N/A |
| K. confusa | CBS 131723 | KF766174 | KF766344 | KF766405 |
| K. micranthae | CBS 131724 | KF766179 | NG042706 | KF766410 |
| K. plurilocularis | CBS 131719 | KF766181 | KF766351 | KF766412 |
| K. yuccifoliorum | CBS 131726 | KF766185 | KF766355 | KF766416 |
| Umthunziomyces hagahagensis | CPC 29917 | KY173472 | KY173561 | N/A |
| Saccharataceae | ||||
| Neoseptorioides eucalypti | CBS 140665 | KT950857 | KT950871 | KT950882 |
| Saccharata banksiae | CPC 27698 | KY173449 | KY173539 | KY173596 |
| S. daviesiae | CPC 29174 | KY173450 | KY173540 | N/A |
| S. proteae | CBS 115206 | KF766226 | DQ377882 | GU349030 |
| Septorioides pini-thunbergi | CBS 473.91 | NR145234 | KF251746 | N/A |
| S. strobi | CBS 141443 | KT884699 | KT884685 | KT884713 |
| Character | Botryosphaeriales (Figure 1) | Botryosphaeria (Figure 10) | Diplodia (Figure 11) | Dothiorella (Figure 12) | Lasiodiplodia (Figure 13) | Neofusicoccum (Figure 14) | ||
|---|---|---|---|---|---|---|---|---|
| Number of base pairs in each gene region (including the gaps after alignment) | ITS (680), LSU (803), tef (374) | ITS (565), tef (265) | ITS (551), tef (331) | ITS (504), tef (343) | ITS (490), tef (344) | ITS (549), tef (253) | ||
| Number of isolates obtained in this study | 12 | 23 | 6 | 2 | 5 | 12 | ||
| Number of taxa originated from GenBank | 95 | 32 | 49 | 68 | 78 | 65 | ||
| Outgroup taxon | Lecanosticta acicula | Macrophomina phaseolina | Lasiodiplodia theobromae | Neofusicoccum parvum | Diplodia mutila | Dothiorella viticola | ||
| MP | Total number of characters | 1857 | 830 | 882 | 847 | 834 | 802 | |
| Constant characters | 954 | 690 | 688 | 578 | 592 | 565 | ||
| Variable/parsimony uninformative characters | 204 | 75 | 52 | 77 | 54 | 107 | ||
| Parsimony informative characters | 704 | 70 | 146 | 202 | 192 | 134 | ||
| Number of parsimonious trees obtained | 77 | 5 | 10 | 10 | 10 | 10 | ||
| Tree length (TL) | 4199 | 180 | 385 | 725 | 559 | 463 | ||
| (CI) | 0.393 | 0.911 | 0.644 | 0.568 | 0.612 | 0.646 | ||
| (RI) | 0.762 | 0.938 | 0.865 | 0.835 | 0.857 | 0.865 | ||
| (RC) | 0.300 | 0.854 | 0.557 | 0.475 | 0.524 | 0.559 | ||
| (HI) | 0.607 | 0.089 | 0.356 | 0.432 | 0.388 | 0.354 | ||
| ML | Final likelihood value | −22,205.648157 | −2130.358718 | −3417.346418 | −4843.109271 | −4183.640855 | −3665.812286 | |
| Number of distinct alignment patterns | 1096 | 212 | 297 | 390 | 293 | 310 | ||
| Percentage of undetermined characters or gaps | 26.52% | 7.31% | 11.38% | 21.84% | 8.52% | 9.63% | ||
| Base frequencies | A | 0.223450 | 0.206023 | 0.207760 | 0.210373 | 0.213076 | 0.201476 | |
| C | 0.259348 | 0.298959 | 0.297528 | 0.293067 | 0.286249 | 0.297204 | ||
| G | 0.288244 | 0.259350 | 0.261525 | 0.252521 | 0.258157 | 0.273045 | ||
| T | 0.228958 | 0.235668 | 0.233187 | 0.244039 | 0.242519 | 0.228274 | ||
| Substitution rates | AC | 1.251462 | 0.503912 | 0.982007 | 1.364862 | 0.729024 | 1.047005 | |
| AG | 2.644446 | 1.747452 | 3.644983 | 2.268590 | 2.724108 | 4.839820 | ||
| AT | 1.415897 | 1.056533 | 0.863841 | 1.240011 | 1.053880 | 1.265619 | ||
| CG | 1.353737 | 0.612519 | 1.681164 | 1.176948 | 0.817807 | 0.870409 | ||
| CT | 4.677551 | 3.387698 | 4.528302 | 4.552526 | 4.062263 | 8.746955 | ||
| GT | 1.000000 | 1.000000 | 1.000000 | 1.000000 | 1.000000 | 1.000000 | ||
| Gamma distribution rate parameter (alpha) | 0.286904 | 0.231276 | 0.174245 | 0.234867 | 0.223197 | 0.268848 | ||
| BI (model of each gene region) | ITS | SYM+I+G | GTR+I | GTR+I+G | HKY+I+G | SYM+I+G | GTR+I+G | |
| tef | HKY+G | GTR+G | HKY+I+G | GTR+G | HKY+I+G | HKY+G | ||
| Tree base ID | 28690 | 28685 | 28686 | 28687 | 28688 | 28689 | ||
| Reviewer access URL | http://purl.org/phylo/treebase/phylows/study/TB2:S28690?x-access-code=fe183dad30514d2dfbbfb8087dbbe53a&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28685?x-access-code=ac877963f1b00fed7de4920660ebc78f&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28686?x-access-code=9df25bff3d5c7c2faeb0029110c6675d&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28687?x-access-code=382c7e58f8f7e3b815b9ae27dbe6f639&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28688?x-access-code=e58fc08c292227a8965dd978203b449a&format=html | http://purl.org/phylo/treebase/phylows/study/TB2:S28689?x-access-code=7152a6819a1dd4ef1e743d4db1e0240c&format=html | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dissanayake, A.J.; Chen, Y.-Y.; Cheewangkoon, R.; Liu, J.-K. Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China. J. Fungi 2021, 7, 893. https://doi.org/10.3390/jof7110893
Dissanayake AJ, Chen Y-Y, Cheewangkoon R, Liu J-K. Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China. Journal of Fungi. 2021; 7(11):893. https://doi.org/10.3390/jof7110893
Chicago/Turabian StyleDissanayake, Asha J., Ya-Ya Chen, Ratchadawan Cheewangkoon, and Jian-Kui Liu. 2021. "Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China" Journal of Fungi 7, no. 11: 893. https://doi.org/10.3390/jof7110893
APA StyleDissanayake, A. J., Chen, Y.-Y., Cheewangkoon, R., & Liu, J.-K. (2021). Occurrence and Morpho-Molecular Identification of Botryosphaeriales Species from Guizhou Province, China. Journal of Fungi, 7(11), 893. https://doi.org/10.3390/jof7110893