
An Optimized and Efficient CRISPR/Cas9 System for the Endophytic Fungus Pestalotiopsis fici



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain, Media and Culture Conditions
2.2. Synthesis and Purification of Single Guide Sequence (sgRNA)
2.3. Gene Cloning, DNA Cassettes Construction and Genetic Manipulation
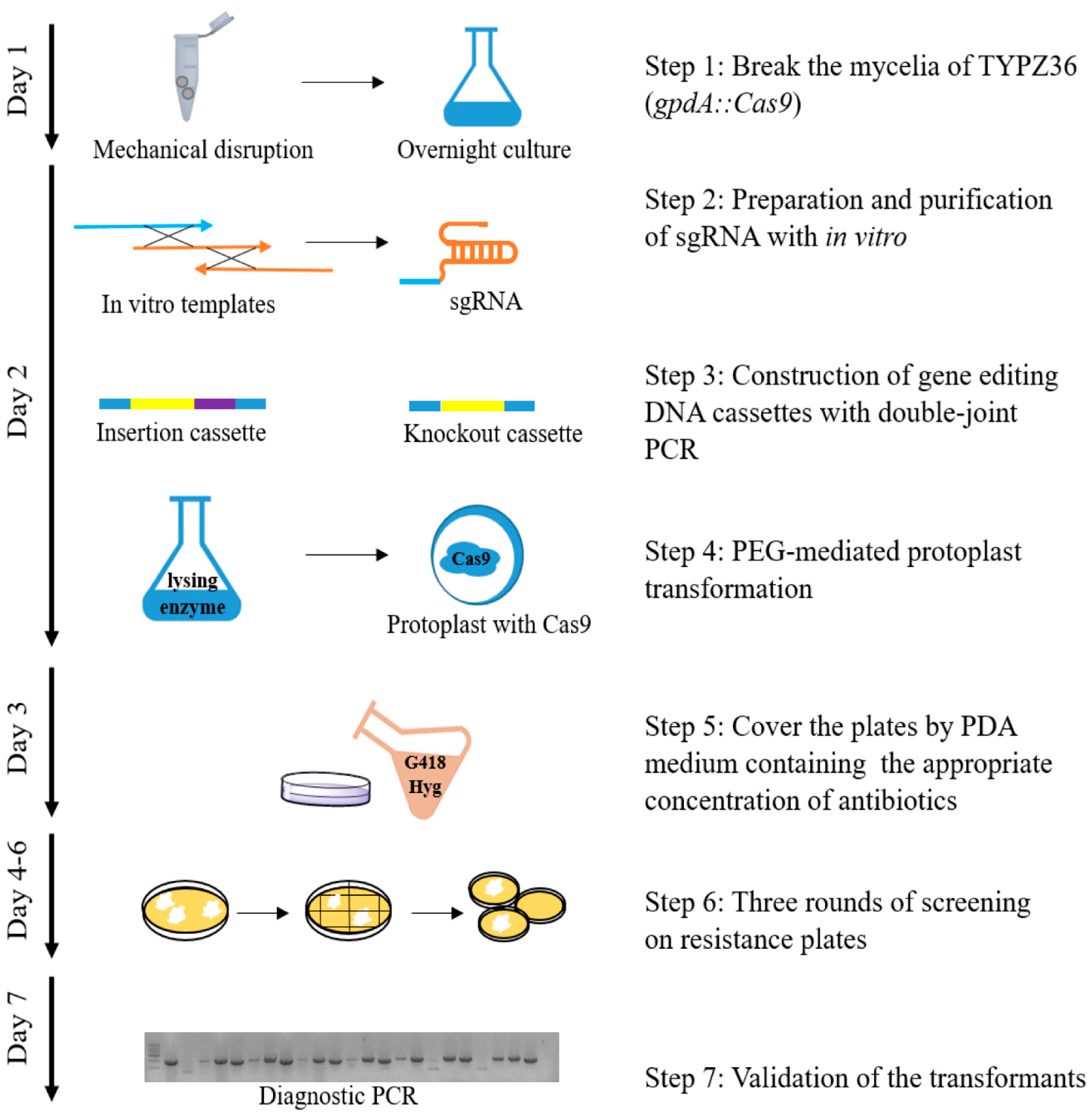
2.4. Transformations of P. fici
3. Results
3.1. Development of a Cas9-Dependent Genome Editing System for P. fici
3.2. DNA Fragment Insertion in CRISPR/Cas9-Dependent Genome Editing System in P. fici
3.3. DNA Deletion in CRISPR/Cas9-Dependent Genome Editing System in P. fici
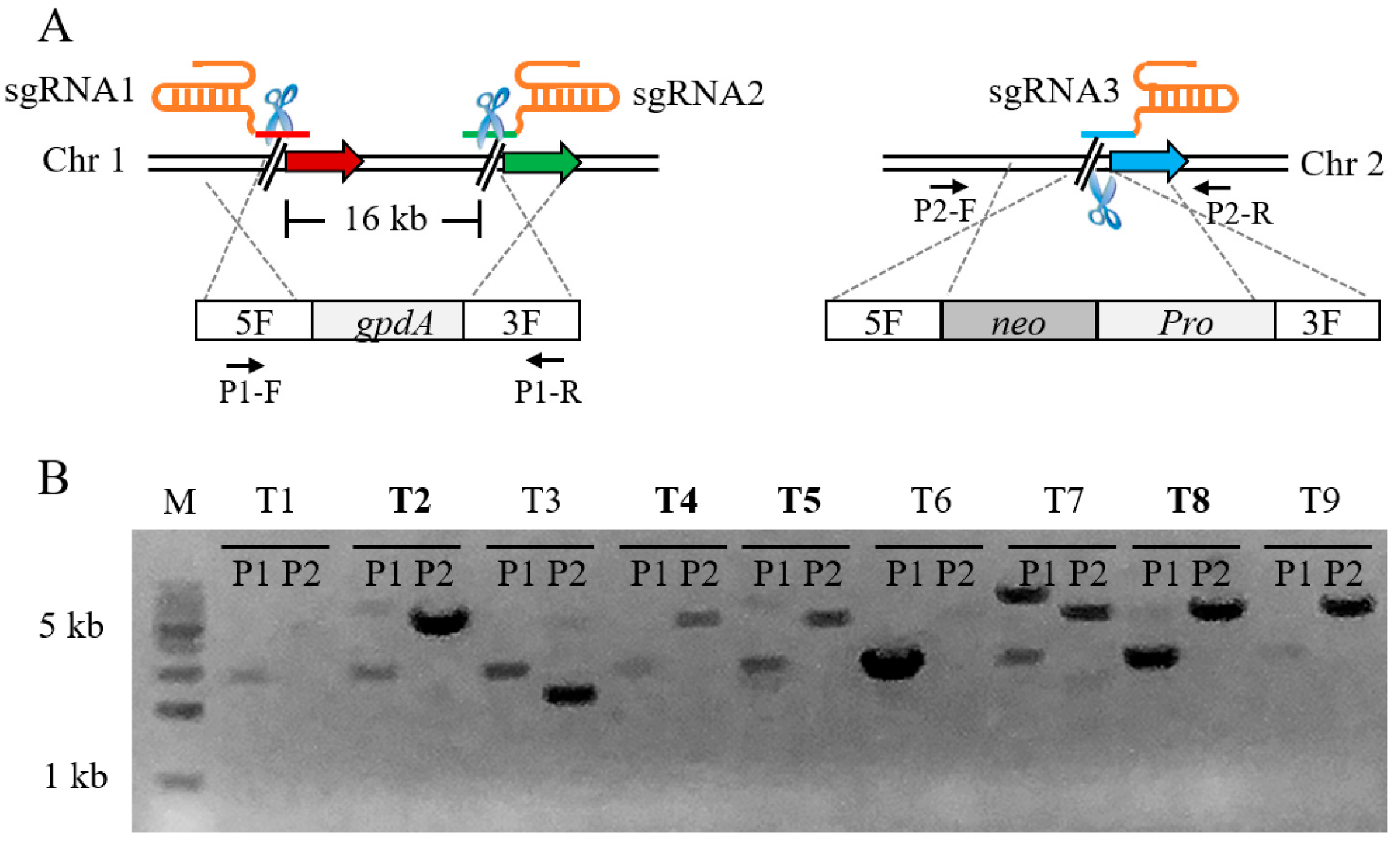
3.4. Dual-Locus Editing in CRISPR/Cas9-Dependent Genome Editing System in P. fici
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Dai, C.-C. Endophytes: A potential resource for biosynthesis, biotransformation, and biodegradation. Ann. Microbiol. 2010, 61, 207–215. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.-D.; Chukeatirote, E.; Bahkali, A.H.; Hyde, K.D. Pestalotiopsis—Morphology, phylogeny, biochemistry and diversity. Fungal Divers. 2011, 50, 167. [Google Scholar] [CrossRef]
- Wang, K.; Lei, J.; Wei, J.; Yao, N. Bioactive natural compounds from the plant endophytic fungi Pestalotiopsis Spp. Mini Rev. Med. Chem. 2012, 12, 1382–1393. [Google Scholar]
- Chen, L.; Zhang, Q.Y.; Jia, M.; Ming, Q.L.; Yue, W.; Rahman, K.; Qin, L.P.; Han, T. Endophytic fungi with antitumor activities: Their occurrence and anticancer compounds. Crit. Rev. Microbiol. 2016, 42, 454–473. [Google Scholar] [CrossRef]
- Sarasan, M.; Puthumana, J.; Job, N.; Han, J.; Lee, J.S.; Philip, R. Marine algicolous endophytic fungi—A promising drug resource of the era. J. Microbiol. Biotechnol. 2017, 27, 1039–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega, H.E.; Torres-Mendoza, D.; Caballero, E.Z.; Cubilla-Rios, L. Structurally uncommon secondary metabolites derived from endophytic fungi. J. Fungi 2021, 7, 570. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, G. Analysis of secondary metabolites from plant endophytic fungi. Methods Mol. Biol. 2018, 1848, 25–38. [Google Scholar]
- Liu, L.; Liu, S.; Jiang, L.; Chen, X.; Guo, L.; Che, Y. Chloropupukeananin, the first chlorinated pupukeanane derivative, and its precursors from Pestalotiopsis fici. Org. Lett. 2008, 10, 1397–1400. [Google Scholar] [CrossRef]
- Liu, S.; Liu, X.; Guo, L.; Che, Y.; Liu, L. 2H-Pyran-2-one and 2H-Furan-2-one derivatives from the plant endophytic fungus Pestalotiopsis fici. Chem. Biodivers. 2013, 10, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Liu, L. Bioactive metabolites from the plant endophyte Pestalotiopsis fici. Mycology 2011, 2, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Bruhn, T.; Guo, L.; Gotz, D.C.; Brun, R.; Stich, A.; Che, Y.; Bringmann, G. Chloropupukeanolides C-E: Cytotoxic pupukeanane chlorides with a spiroketal skeleton from Pestalotiopsis fici. Chemistry 2011, 17, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, S.; Chen, X.; Guo, L.; Che, Y. Pestalofones A-E, bioactive cyclohexanone derivatives from the plant endophytic fungus Pestalotiopsis fici. Bioorg. Med. Chem. 2009, 17, 606–613. [Google Scholar] [CrossRef]
- Liu, L.; Liu, S.; Niu, S.; Guo, L.; Chen, X.; Che, Y. Isoprenylated chromone derivatives from the plant endophytic fungus Pestalotiopsis fici. J. Nat. Prod. 2009, 72, 1482–1486. [Google Scholar] [CrossRef]
- Liu, L.; Niu, S.; Lu, X.; Chen, X.; Zhang, H.; Guo, L.; Che, Y. Unique metabolites of Pestalotiopsis fici suggest a biosynthetic hypothesis involving a Diels-Alder reaction and then mechanistic diversification. Chem. Commun. 2010, 46, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhang, C.; Gao, Q.; Hou, B.; Liu, L.; Yang, H.; Jiang, X. Chloropupukeananin and pestalofone C regulate autophagy through AMPK and glycolytic pathway. Chem. Biodivers. 2020, 17, e1900583. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, X.; Liu, L.; Xiang, M.; Wang, W.; Sun, X.; Che, Y.; Guo, L.; Liu, G.; Guo, L.; et al. Genomic and transcriptomic analysis of the endophytic fungus Pestalotiopsis fici reveals its lifestyle and high potential for synthesis of natural products. BMC Genomics 2015, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Zhou, H.; Zhang, P.; Wang, X.; Li, W.; Zhang, W.; Liu, X.; Liu, H.W.; Keller, N.P.; An, Z.; et al. Polyketide production of pestaloficiols and macrodiolide ficiolides revealed by manipulations of epigenetic regulators in an endophytic fungus. Org. Lett. 2016, 18, 1832–1835. [Google Scholar] [CrossRef]
- Xu, X.; Liu, L.; Zhang, F.; Wang, W.; Li, J.; Guo, L.; Che, Y.; Liu, G. Identification of the first diphenyl ether gene cluster for pestheic acid biosynthesis in plant endophyte Pestalotiopsis Fici. Chembiochem 2014, 15, 284–292. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, L.; Guan, F.; Li, E.; Jin, J.; Li, J.; Che, Y.; Liu, G. Characterization of a prenyltransferase for Iso-A82775C biosynthesis and generation of new congeners of chloropestolides. ACS Chem. Biol. 2018, 13, 703–711. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, X.; Fan, A.; Zheng, Y.; Liu, X.; Wang, S.; Zou, H.; Oakley, B.R.; Keller, N.P.; Yin, W.B. A cryptic pigment biosynthetic pathway uncovered by heterologous expression is essential for conidial development in Pestalotiopsis fici. Mol. Microbiol. 2017, 105, 469–483. [Google Scholar] [CrossRef] [Green Version]
- Fan, A.; Mi, W.; Liu, Z.; Zeng, G.; Zhang, P.; Hu, Y.; Fang, W.; Yin, W.B. Deletion of a histone acetyltransferase leads to the pleiotropic activation of natural products in Metarhizium robertsii. Org. Lett. 2017, 19, 1686–1689. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, P.; Zhou, H.; Liu, X.; Li, S.M.; Guo, L.; Li, K.; Yin, W.B. A new regulator RsdA mediating fungal secondary metabolism has a detrimental impact on asexual development in Pestalotiopsis fici. Environ. Microbiol. 2019, 21, 416–426. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Zhou, S.; Wang, G.; An, Z.; Liu, X.; Li, K.; Yin, W.B. Two transcription factors cooperatively regulate DHN melanin biosynthesis and development in Pestalotiopsis fici. Mol. Microbiol. 2019, 112, 649–666. [Google Scholar] [CrossRef]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 2015, 1, 15007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohl, C.; Kiel, J.A.; Driessen, A.J.; Bovenberg, R.A.; Nygard, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synth. Biol. 2016, 5, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Meng, X.; Wei, X.; Lu, L. Highly efficient CRISPR mutagenesis by microhomology-mediated end joining in Aspergillus fumigatus. Fungal Genet. Biol. 2016, 86, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Sarkari, P.; Marx, H.; Blumhoff, M.L.; Mattanovich, D.; Sauer, M.; Steiger, M.G. An efficient tool for metabolic pathway construction and gene integration for Aspergillus niger. Bioresour Technol. 2017, 245, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.Y.; Wu, Y.J.; Xie, Q.P.; Tang, J.W.; Yu, Z.T.; Yang, S.B.; Chen, S.X. CRISPR/Cas9-based genome editing in the filamentous fungus Glarea lozoyensis and its application in manipulating gloF. ACS Synth. Biol. 2020, 9, 1968–1977. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat. Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Feng, J.; Zhang, P.; Fan, J.; Yin, W.B. A CRISPR/Cas9 cleavage system for capturing fungal secondary metabolite gene clusters. J. Microbiol. Biotechnol. 2021, 31, 8–15. [Google Scholar] [CrossRef] [PubMed]
- E-CRISP Design. Available online: http://www.e-crisp.org/E-CRISP/designcrispr.html (accessed on 20 September 2021).
- Yu, J.H.; Hamari, Z.; Han, K.H.; Seo, J.A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.B.; Amaike, S.; Wohlbach, D.J.; Gasch, A.P.; Chiang, Y.M.; Wang, C.C.; Bok, J.W.; Rohlfs, M.; Keller, N.P. An Aspergillus nidulans bZIP response pathway hardwired for defensive secondary metabolism operates through aflR. Mol. Microbiol. 2012, 83, 1024–1034. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, G.; Li, W.; Liu, X.; Li, E.; Yin, W.B. A highly efficient genetic system for the identification of a harzianum B biosynthetic gene cluster in Trichoderma hypoxylon. Microbiology 2018, 164, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Gorucu Yilmaz, S. Genome editing technologies: CRISPR, LEAPER, RESTORE, ARCUT, SATI, and RESCUE. EXCLI J. 2021, 20, 19–45. [Google Scholar]
- Jiang, C.; Lv, G.; Tu, Y.; Cheng, X.; Duan, Y.; Zeng, B.; He, B. Applications of CRISPR/Cas9 in the synthesis of secondary metabolites in filamentous fungi. Front. Microbiol. 2021, 12, 638096. [Google Scholar] [CrossRef]
- Meliawati, M.; Schilling, C.; Schmid, J. Recent advances of Cas12a applications in bacteria. Appl. Microbiol. Biotechnol. 2021, 105, 2981–2990. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR-Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef]
- Zhang, R.; Xu, W.; Shao, S.; Wang, Q. Gene Silencing Through CRISPR interference in bacteria: Current advances and future prospects. Front. Microbiol. 2021, 12, 635227. [Google Scholar] [CrossRef]
- Shihong Gao, D.; Zhu, X.; Lu, B. Development and application of sensitive, specific, and rapid CRISPR-Cas13-based diagnosis. J. Med. Virol. 2021, 93, 4198–4204. [Google Scholar]
- Liu, Q.; Zhang, Y.; Li, F.; Li, J.; Sun, W.; Tian, C. Upgrading of efficient and scalable CRISPR-Cas-mediated technology for genetic engineering in thermophilic fungus Myceliophthora thermophila. Biotechnol. Biofuels 2019, 12, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Dong, H.; Zheng, J.; Wang, B.; Pan, L. Highly efficient single base editing in Aspergillus niger with CRISPR/Cas9 cytidine deaminase fusion. Microbiol. Res. 2019, 223, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, F.; Liu, L.; Liu, X.; Che, Y.; Keller, N.P.; Guo, L.; Yin, W.B. The bZIP transcription factor PfZipA regulates secondary metabolism and oxidative stress response in the plant endophytic fungus Pestalotiopsis fici. Fungal Genet. Biol. 2015, 81, 221–228. [Google Scholar] [CrossRef]
- Lin, H.; Lyu, H.; Zhou, S.; Yu, J.; Keller, N.P.; Chen, L.; Yin, W.B. Deletion of a global regulator LaeB leads to the discovery of novel polyketides in Aspergillus nidulans. Org. Biomol. Chem. 2018, 16, 4973–4976. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Keller, N.; Oakley, B.R.; Wang, C.C. A gene cluster containing two fungal polyketide synthases encodes the biosynthetic pathway for a polyketide, asperfuranone, in Aspergillus nidulans. J. Am. Chem. Soc. 2009, 131, 2965–2970. [Google Scholar] [CrossRef] [Green Version]
- Zabala, A.O.; Xu, W.; Chooi, Y.H.; Tang, Y. Characterization of a silent azaphilone gene cluster from Aspergillus niger ATCC 1015 reveals a hydroxylation-mediated pyran-ring formation. Chem. Biol. 2012, 19, 1049–1059. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.W.; Lee, S.H.; Kim, L.H.; Ryu, J.G.; Lee, S.; Seo, Y.; Kim, Y.H.; Busman, M.; Yun, S.H.; Proctor, R.H.; et al. Identification of a 12-gene fusaric acid biosynthetic gene cluster in fusarium species through comparative and functional genomics. Mol. Plant-Microbe Interact. 2015, 28, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Awakawa, T.; Yang, X.L.; Wakimoto, T.; Abe, I. Pyranonigrin E: A PKS-NRPS hybrid metabolite from Aspergillus niger identified by genome mining. Chembiochem 2013, 14, 2095–2099. [Google Scholar] [CrossRef]
- Ma, K.; Zhang, P.; Tao, Q.; Keller, N.P.; Yang, Y.; Yin, W.B.; Liu, H. Characterization and biosynthesis of a rare fungal hopane-type triterpenoid glycoside involved in the antistress property of Aspergillus fumigatus. Org. Lett. 2019, 21, 3252–3256. [Google Scholar] [CrossRef]
- Chen, L.; Wu, H.; Liu, H.; Li, E.; Ren, J.; Wang, W.; Wang, S.; Yin, W.B. Genetic dereplication of Trichoderma hypoxylon reveals two novel polycyclic lactones. Bioorg. Chem. 2019, 91, 103185. [Google Scholar] [CrossRef]
- Fan, J.; Liao, G.; Ludwig-Radtke, L.; Yin, W.B.; Li, S.M. Formation of terrestric acid in Penicillium crustosum requires redox-assisted decarboxylation and stereoisomerization. Org. Lett. 2020, 22, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Pu, Y.H.; Ren, J.W.; Li, E.W.; Guo, L.X.; Yin, W.B. Genetic dereplication driven discovery of a tricinoloniol acid biosynthetic pathway in Trichoderma hypoxylon. Org. Biomol. Chem. 2020, 18, 5344–5348. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, W.; Zhang, P.; Fan, J.; Zhang, F.; Wang, C.; Li, S.; Sun, Y.; Chen, S.; Yin, W. Tricarbocyclic core formation of tyrosine-decahydrofluorenes implies a three-enzyme cascade with XenF-mediated sigmatropic rearrangement as a prerequisite. Acta Pharm Sin. B 2021, in press. [Google Scholar] [CrossRef]
- Zhang, J.; Qi, L.; Chen, G.; Yin, W.B. Discovery and genetic identification of amphiphilic coprogen siderophores from Trichoderm hypoxylon. Appl. Microbiol. Biotechnol. 2021, 105, 2831–2839. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fan, J.; Zhang, P.; Hu, Y.; Liu, X.; Li, S.M.; Yin, W.B. New insights into the disulfide bond formation enzymes in epidithiodiketopiperazine alkaloids. Chem. Sci. 2021, 12, 4132–4138. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Name | Length (bp) | Amino Acid (aa) | Transformants | Verified Transformants | Efficiency |
|---|---|---|---|---|---|
| gd1 | 5458 | 1796 | 12 | 3 | 25.0% |
| gd2 | 5802 | 1933 | 16 | 4 | 25.0% |
| gd3 | 7110 | 2209 | 15 | 6 | 40.0% |
| gd4 | 7134 | 2313 | 22 | 3 | 13.6% |
| gd5 | 7250 | 2269 | 32 | 6 | 18.7% |
| gd6 | 8031 | 2568 | 15 | 5 | 33.3% |
| gd7 | 8085 | 2536 | 26 | 7 | 26.9% |
| gd8 | 8139 | 2600 | 15 | 6 | 40.0% |
| gd9 | 8512 | 2583 | 24 | 9 | 37.5% |
| gd10 | 9133 | 2969 | 33 | 4 | 12.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Huang, R.; Yin, W.-B. An Optimized and Efficient CRISPR/Cas9 System for the Endophytic Fungus Pestalotiopsis fici. J. Fungi 2021, 7, 809. https://doi.org/10.3390/jof7100809
Xu X, Huang R, Yin W-B. An Optimized and Efficient CRISPR/Cas9 System for the Endophytic Fungus Pestalotiopsis fici. Journal of Fungi. 2021; 7(10):809. https://doi.org/10.3390/jof7100809
Chicago/Turabian StyleXu, Xinran, Runye Huang, and Wen-Bing Yin. 2021. "An Optimized and Efficient CRISPR/Cas9 System for the Endophytic Fungus Pestalotiopsis fici" Journal of Fungi 7, no. 10: 809. https://doi.org/10.3390/jof7100809
APA StyleXu, X., Huang, R., & Yin, W.-B. (2021). An Optimized and Efficient CRISPR/Cas9 System for the Endophytic Fungus Pestalotiopsis fici. Journal of Fungi, 7(10), 809. https://doi.org/10.3390/jof7100809