Absence of Azole Antifungal Resistance in Aspergillus fumigatus Isolated from Root Vegetables Harvested from UK Arable and Horticultural Soils

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Environmental Sampling and Isolation of Aspergillus Species
2.2. Susceptibility Testing
2.3. Molecular Identification of Azole-Resistant Fungal Species
3. Results
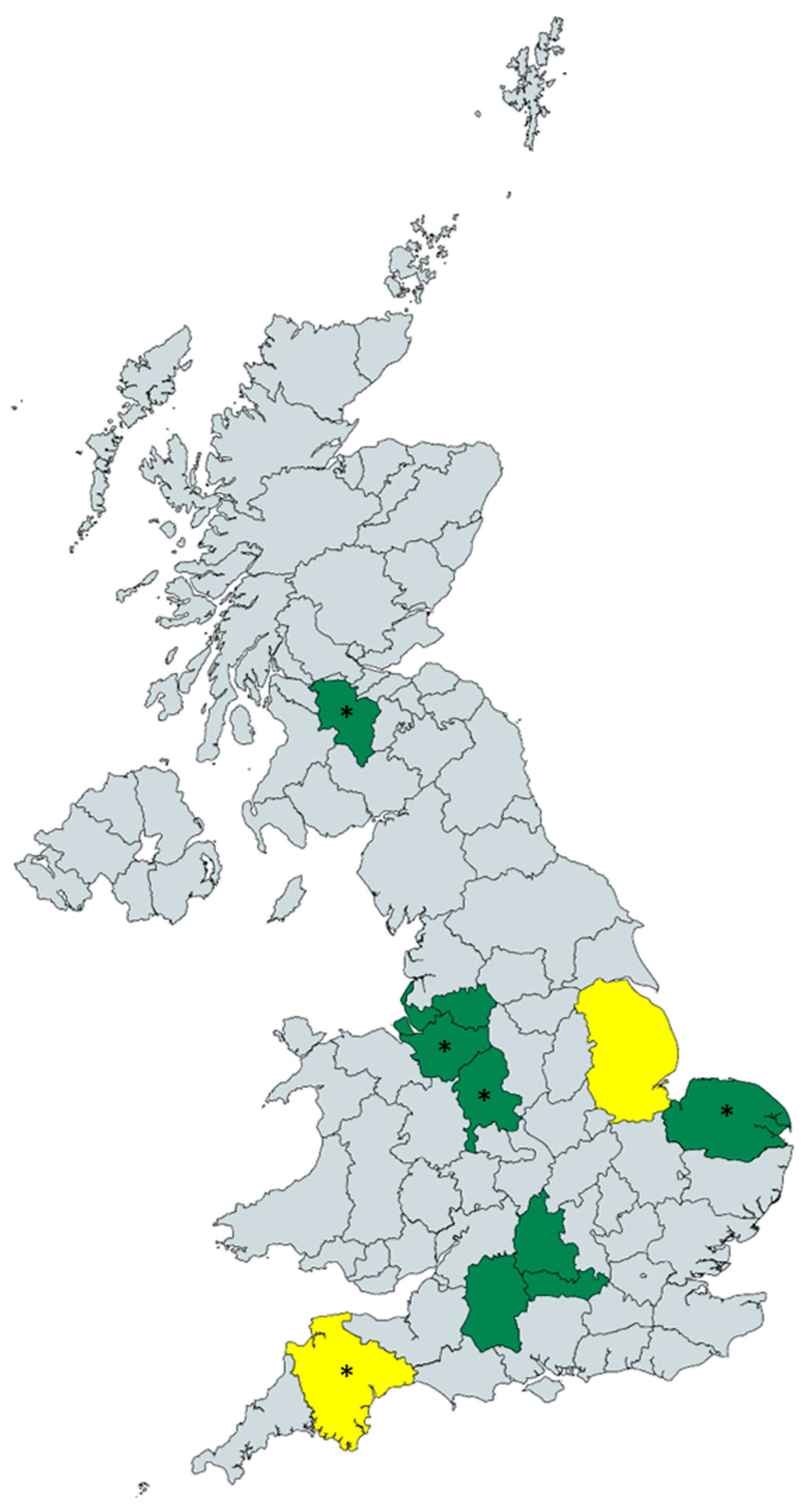
3.1. Environmental Sampling
3.2. Antifungal Susceptibility Testing of Aspergillus Species and Their Identification
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klich, M.A. Biogeography of Aspergillus species in soil and litter. Mycologia 2002, 94, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Susca, A.; Cozzi, G.; Ehrlich, K.; Varga, J.; Frisvad, J.C.; Meijer, M.; Noonim, P.; Mahakarnchanakul, W.; Samson, R.A. Biodiversity of Aspergillus species in some important agricultural products. Stud. Mycol. 2007, 59, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Seyedmousavi, S.; Guillot, J.; Arne, P.; de Hoog, G.S.; Mouton, J.W.; Melchers, W.J.; Verweij, P.E. Aspergillus and aspergilloses in wild and domestic animals: A global health concern with parallels to human disease. Med. Mycol. 2015, 53, 765–797. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33. [Google Scholar] [CrossRef]
- Barber, A.E.; Scheufen, S.; Walther, G.; Kurzai, O.; Schmidt, V. Low rate of azole resistance in cases of avian aspergillosis in Germany. Med. Mycol. 2020. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Sugui, J.A. Aspergillus fumigatus--what makes the species a ubiquitous human fungal pathogen? PLoS Path. 2013, 9, e1003743. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv113. [Google Scholar] [CrossRef]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24, e1–e38. [Google Scholar] [CrossRef]
- Snelders, E.; Camps, S.M.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.; van der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.; Verweij, P.E. Triazole fungicides can induce cross-resistance to medical triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef]
- Ren, J.; Jin, X.; Zhang, Q.; Zheng, Y.; Lin, D.; Yu, Y. Fungicides induced triazole-resistance in Aspergillus fumigatus associated with mutations of TR46/Y121F/T289A and its appearance in agricultural fields. J. Hazard. Mater. 2017, 326, 54–60. [Google Scholar] [CrossRef]
- Bowyer, P.; Moore, C.B.; Rautemaa, R.; Denning, D.W.; Richardson, M.D. Azole antifungal resistance today: Focus on Aspergillus. Current Infect. Dis. Rep. 2011, 13, 485–491. [Google Scholar] [CrossRef] [PubMed]
- van der Linden, J.W.; Snelders, E.; Kampinga, G.A.; Rijnders, B.J.; Mattsson, E.; Debets-Ossenkopp, Y.J.; Kuijper, E.J.; Van Tiel, F.H.; Melchers, W.J.; Verweij, P.E. Clinical implications of azole resistance in Aspergillus fumigatus, The Netherlands, 2007-2009. Emerg Infect. Dis. 2011, 17, 1846–1854. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-Resistant Aspergillosis: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S436–s444. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.E.; Riedel, J.; Sae-Ong, T.; Kang, K.; Brabetz, W.; Panagiotou, G.; Deising, H.B.; Kurzai, O. Azole use in agriculture reduces Aspergillus fumigatus abundance but does not alter its population structure. bioRxiv 2020. [Google Scholar] [CrossRef]
- Snelders, E.; Huis In ‘t Veld, R.A.; Rijs, A.J.; Kema, G.H.; Melchers, W.J.; Verweij, P.E. Possible environmental origin of resistance of Aspergillus fumigatus to medical triazoles. Appl. Environment. Microbiol. 2009, 75, 4053–4057. [Google Scholar] [CrossRef]
- Snelders, E.; van der Lee, H.A.; Kuijpers, J.; Rijs, A.J.; Varga, J.; Samson, R.A.; Mellado, E.; Donders, A.R.; Melchers, W.J.; Verweij, P.E. Emergence of azole resistance in Aspergillus fumigatus and spread of a single resistance mechanism. PLoS Med. 2008, 5, e219. [Google Scholar] [CrossRef]
- Vaezi, A.; Fakhim, H.; Javidnia, J.; Khodavaisy, S.; Abtahian, Z.; Vojoodi, M.; Nourbakhsh, F.; Badali, H. Pesticide behavior in paddy fields and development of azole-resistant Aspergillus fumigatus: Should we be concerned? J. Mycol. Med. 2018, 28, 59–64. [Google Scholar] [CrossRef]
- Prigitano, A.; Esposto, M.C.; Romanò, L.; Auxilia, F.; Tortorano, A.M. Azole-resistant Aspergillus fumigatus in the Italian environment. J. Glob. Antimicrob. Re. 2019, 16, 220–224. [Google Scholar] [CrossRef]
- Hagiwara, D. Current Status of Azole-resistant Aspergillus fumigatus Isolates in East Asia. Med. Mycol. J. 2018, 59, E71–E76. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Frade, J.P.; Etienne, K.A.; Pfaller, M.A.; Diekema, D.J.; Balajee, S.A. Azole resistance in Aspergillus fumigatus isolates from the ARTEMIS global surveillance study is primarily due to the TR/L98H mutation in the cyp51A gene. Antimicrob. Agents Ch. 2011, 55, 4465–4468. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Kathuria, S.; Hagen, F.; Meis, J.F. Azole-resistant Aspergillus fumigatus with the environmental TR46/Y121F/T289A mutation in India. J. Antimicrob. Ch. 2014, 69, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Le Pape, P.; Lavergne, R.A.; Morio, F.; Alvarez-Moreno, C. Multiple Fungicide-Driven Alterations in Azole-Resistant Aspergillus fumigatus, Colombia, 2015. Emerg. Infect. Dis. 2016, 22, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; van den Boom, M.; Yntema, J.B.; Hagen, F.; Verweij, P.E.; Meis, J.F. Multi-azole-resistant Aspergillus fumigatus in the environment in Tanzania. J. Antimicrob. Ch. 2014, 69, 2979–2983. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Dong, F.; Zhao, J.; Fan, H.; Qin, C.; Li, R.; Verweij, P.E.; Zheng, Y.; Han, L. High Azole Resistance in Aspergillus fumigatus Isolates from Strawberry Fields, China, 2018. J. Antimicrob. Ch. 2020, 26, 81–89. [Google Scholar] [CrossRef]
- Hurst, S.F.; Berkow, E.L.; Stevenson, K.L.; Litvintseva, A.P.; Lockhart, S.R. Isolation of azole-resistant Aspergillus fumigatus from the environment in the south-eastern USA. J. Antimicrob. Ch. 2017, 72, 2443–2446. [Google Scholar] [CrossRef]
- Bader, O.; Tünnermann, J.; Dudakova, A.; Tangwattanachuleeporn, M.; Weig, M.; Groß, U. Environmental isolates of azole-resistant Aspergillus fumigatus in Germany. Antimicrob. Agents Ch. 2015, 59, 4356–4359. [Google Scholar] [CrossRef]
- Godeau, C.; Reboux, G.; Scherer, E.; Laboissiere, A.; Lechenault-Bergerot, C.; Millon, L.; Rocchi, S. Azole-resistant Aspergillus fumigatus in the hospital: Surveillance from flower beds to corridors. Am. J. Infect. Control 2020, 48, 702–704. [Google Scholar] [CrossRef]
- Mortensen, K.L.; Mellado, E.; Lass-Flörl, C.; Rodriguez-Tudela, J.L.; Johansen, H.K.; Arendrup, M.C. Environmental study of azole-resistant Aspergillus fumigatus and other aspergilli in Austria, Denmark, and Spain. Antimicrob. Agents Ch. 2010, 54, 4545–4549. [Google Scholar] [CrossRef]
- Dauchy, C.; Bautin, N.; Nseir, S.; Reboux, G.; Wintjens, R.; Le Rouzic, O.; Sendid, B.; Viscogliosi, E.; Le Pape, P.; Arendrup, M.C.; et al. Emergence of Aspergillus fumigatus azole resistance in azole-naïve patients with chronic obstructive pulmonary disease and their homes. Indoor Air 2018, 28, 298–306. [Google Scholar] [CrossRef]
- van der Linden, J.W.; Camps, S.M.; Kampinga, G.A.; Arends, J.P.; Debets-Ossenkopp, Y.J.; Haas, P.J.; Rijnders, B.J.; Kuijper, E.J.; van Tiel, F.H.; Varga, J.; et al. Aspergillosis due to voriconazole highly resistant Aspergillus fumigatus and recovery of genetically related resistant isolates from domiciles. Clin. Infect. Dis. 2013, 57, 513–520. [Google Scholar] [CrossRef]
- Lavergne, R.A.; Chouaki, T.; Hagen, F.; Toublanc, B.; Dupont, H.; Jounieaux, V.; Meis, J.F.; Morio, F.; Le Pape, P. Home Environment as a Source of Life-Threatening Azole-Resistant Aspergillus fumigatus in Immunocompromised Patients. Clin. Infect. Dis. 2017, 64, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, P.; Kleinkauf, N.; Verweij, P.E.; Arendrup, M.C.; Donnelly, P.J.; Cuenca-Estrella, M.; Fraaije, B.; Melchers, W.J.G.; Adriaenssens, N.; Kema, G.H.J.; et al. Risk assessment on the impact of environmental usage of triazoles on the development and spread of resistance to medical triazoles in Aspergillus species. Stockholm: ECDC 2013. [Google Scholar] [CrossRef]
- Bromley, M.J.; van Muijlwijk, G.; Fraczek, M.G.; Robson, G.; Verweij, P.E.; Denning, D.W.; Bowyer, P. Occurrence of azole-resistant species of Aspergillus in the UK environment. J. Global Antimicrob. Res. 2014, 2, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Tsitsopoulou, A.; Posso, R.; Vale, L.; Bebb, S.; Johnson, E.; White, P.L. Determination of the Prevalence of Triazole Resistance in Environmental Aspergillus fumigatus Strains Isolated in South Wales, UK. Front. Microbiol. 2018, 9, 1395. [Google Scholar] [CrossRef]
- Sewell, T.R.; Zhang, Y.; Brackin, A.P.; Shelton, J.M.G.; Rhodes, J.; Fisher, M.C. Elevated Prevalence of Azole-Resistant Aspergillus fumigatus in Urban versus Rural Environments in the United Kingdom. Antimicrob. Agents Chem. 2019, 63. [Google Scholar] [CrossRef]
- Rodriquez-Tudela, J.L.; Donnelly, J.P.; Arendrup, M.C.; Arikan, S.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Dannaoui, E.; Denning, D.; et al. EUCAST Technical Note on the method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia-forming moulds. Clin. Microbiol. Infect. 2008, 14, 982–984. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proceed. Nat. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Peterson, S.W. Phylogenetic analysis of Aspergillus species using DNA sequences from four loci. Mycologia 2008, 100, 205–226. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38 - Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environment. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Hong, S.B.; Go, S.J.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Aspergillus fumigatus and related species. Mycologia 2005, 97, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Hong, S.; Peterson, S.W.; Frisvad, J.C.; Varga, J. Polyphasic taxonomy of Aspergillus section Fumigati and its teleomorph Neosartorya. Stud. Mycol. 2007, 59, 147–203. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Weig, M.; Gross, U.; Meijer, M.; Bader, O. Aspergillus oerlinghausenensis, a new mould species closely related to A. fumigatus. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Krekels, E.H.J.; Verweij, P.E.; Buil, J.B.; Knibbe, C.A.J.; Brüggemann, R.J.M. Pharmacokinetics and Pharmacodynamics of Posaconazole. Drugs 2020, 80, 671–695. [Google Scholar] [CrossRef]
- Astvad, K.M.; Jensen, R.H.; Hassan, T.M.; Mathiasen, E.G.; Thomsen, G.M.; Pedersen, U.G.; Christensen, M.; Hilberg, O.; Arendrup, M.C. First detection of TR46/Y121F/T289A and TR34/L98H alterations in Aspergillus fumigatus isolates from azole-naive patients in Denmark despite negative findings in the environment. Antimicrob. Agents Chem. 2014, 58, 5096–5101. [Google Scholar] [CrossRef]
- Hong, S.B.; Kim, D.H.; Park, I.C.; Samson, R.A.; Shin, H.D. Isolation and identification of Aspergillus section fumigati strains from arable soil in Korea. Mycobiology 2010, 38, 1–6. [Google Scholar] [CrossRef]
- Ugwuanyi, J.O.; Obeta, J.A. Incidence of heat-resistant fungi in Nsukka, southern Nigeria. Int. J. Food Microbiol. 1991, 13, 157–164. [Google Scholar] [CrossRef]
- Santos, J.; Samapundo, S.; Biyikli, A.; Van Impe, J.; Akkermans, S.; Höfte, M.; Abatih, E.N.; Sant’Ana, A.S.; Devlieghere, F. Occurrence, distribution and contamination levels of heat-resistant moulds throughout the processing of pasteurized high-acid fruit products. Int. J. Food Microbiol. 2018, 281, 72–81. [Google Scholar] [CrossRef]
- Jeanvoine, A.; Rocchi, S.; Reboux, G.; Crini, N.; Crini, G.; Millon, L. Azole-resistant Aspergillus fumigatus in sawmills of Eastern France. J. Appl. Microbiol. 2017, 123, 172–184. [Google Scholar] [CrossRef]
- Van Der Linden, J.W.; Warris, A.; Verweij, P.E. Aspergillus species intrinsically resistant to antifungal agents. Med. Mycol. 2011, 49, S82–S89. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Verweij, P.E.; Mouton, J.W.; Lagrou, K.; Meletiadis, J. Multicentre validation of 4-well azole agar plates as a screening method for detection of clinically relevant azole-resistant Aspergillus fumigatus. J. Antimicrob. Ch. 2017, 72, 3325–3333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Snelders, E.; Zwaan, B.J.; Schoustra, S.E.; Meis, J.F.; van Dijk, K.; Hagen, F.; van der Beek, M.T.; Kampinga, G.A.; Zoll, J.; et al. A Novel Environmental Azole Resistance Mutation in Aspergillus fumigatus and a Possible Role of Sexual Reproduction in Its Emergence. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Pung, H.; Hall, B.H. Managing leaf diseases of carrots with traditional and alternative fungicides including baseline sensitivity studies. Crop. Protect. 2017, 93, 52–59. [Google Scholar] [CrossRef]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest. Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Alshareef, F.; Robson, G.D. Prevalence, persistence, and phenotypic variation of Aspergillus fumigatus in the outdoor environment in Manchester, UK, over a 2-year period. Med. Mycol. 2014, 52, 367–375. [Google Scholar] [CrossRef]
- Shelton, J.M.G.; Fisher, M.C.; Singer, A.C. Campaign-Based Citizen Science for Environmental Mycology: The “Science Solstice” and “Summer Soil-stice” Projects to Assess Drug Resistance in Air and Soilborne Aspergillus fumigatus. bioRxiv 2020. [Google Scholar] [CrossRef]


{kind=link}
| Sampling Site | Number of Samples Collected | Number of Samples with A. fumigatus Growth (%) | Total A. fumigatus Isolates Recovered |
|---|---|---|---|
| Berkshire | 9 | 5 (56) | 9 |
| Cheshire | 28 | 15 (54) | 34 |
| Devon | 3 | 0 (0) | 0 |
| Greater Manchester | 3 | 1 (33) | 2 |
| Lincolnshire | 3 | 0 (0) | 0 |
| Merseyside | 3 | 3 (100) | 15 |
| Norfolk | 6 | 3 (50) | 6 |
| Oxfordshire | 6 | 3 (50) | 5 |
| South Lanarkshire | 1 | 1 (100) | 10 |
| Staffordshire | 9 | 8 (89) | 60 |
| Wiltshire | 3 | 2 (67) | 5 |
| Total | 74 | 41 (55) | 146 |
| Isolate | Sampling Site | Sample Source | Azole Resistance Primary | MIC (mg/L) 2 | Species | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Screening Method Results 1 | ||||||||||
| Control | ITC | VRC | POS | ITC | VRC | POS | ||||
| 4 | Norfolk | Beetroot | + | - | + | - | >8 | 4 | 0.5 | Neosartorya fischeri |
| 13 | Staffordshire | Carrot | + | - | + | - | >8 | 2 | 0.5 | Neosartorya fischeri |
| 99 | South Lanarkshire | Potato | + | + | + | - | >8 | 4 | 0.5 | Aspergillus oerlinghausenensis |
| 142 | Devon | Potato | + | + | + | + | >8 | 8 | 2 | Neosartorya spinosa |
| 146 | Cheshire | Potato | + | + | + | - | >8 | 4 | 0.5 | Aspergillus oerlinghausenensis |
| 148 | Cheshire | Potato | + | + | + | - | >8 | 4 | 0.5 | Neosartorya fischeri |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Torre, M.H.; Whitby, C.; Eades, C.P.; Moore, C.B.; Novak-Frazer, L.; Richardson, M.D.; Rautemaa-Richardson, R. Absence of Azole Antifungal Resistance in Aspergillus fumigatus Isolated from Root Vegetables Harvested from UK Arable and Horticultural Soils. J. Fungi 2020, 6, 208. https://doi.org/10.3390/jof6040208
van der Torre MH, Whitby C, Eades CP, Moore CB, Novak-Frazer L, Richardson MD, Rautemaa-Richardson R. Absence of Azole Antifungal Resistance in Aspergillus fumigatus Isolated from Root Vegetables Harvested from UK Arable and Horticultural Soils. Journal of Fungi. 2020; 6(4):208. https://doi.org/10.3390/jof6040208
Chicago/Turabian Stylevan der Torre, Mireille H., Cheryl Whitby, Christopher P. Eades, Caroline B. Moore, Lilyann Novak-Frazer, Malcolm D. Richardson, and Riina Rautemaa-Richardson. 2020. "Absence of Azole Antifungal Resistance in Aspergillus fumigatus Isolated from Root Vegetables Harvested from UK Arable and Horticultural Soils" Journal of Fungi 6, no. 4: 208. https://doi.org/10.3390/jof6040208
APA Stylevan der Torre, M. H., Whitby, C., Eades, C. P., Moore, C. B., Novak-Frazer, L., Richardson, M. D., & Rautemaa-Richardson, R. (2020). Absence of Azole Antifungal Resistance in Aspergillus fumigatus Isolated from Root Vegetables Harvested from UK Arable and Horticultural Soils. Journal of Fungi, 6(4), 208. https://doi.org/10.3390/jof6040208