Target Activity of Isaria tenuipes (Hypocreales: Clavicipitaceae) Fungal Strains against Dengue Vector Aedes aegypti (Linn.) and Its Non-Target Activity Against Aquatic Predators


 ,
,  ,
, 
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methodology
2.1. Mosquito Culture
2.2. Isaria Tenuipes
2.3. Larvicidal Bioassay
2.4. Oviposition Deterrence Index
2.5. Fecundity Assay
2.6. Enzyme Assays
2.7. Gut Histological and External Physiological Assay
2.8. Non-Target Toxicity Assay
2.9. Data Analysis
3. Results
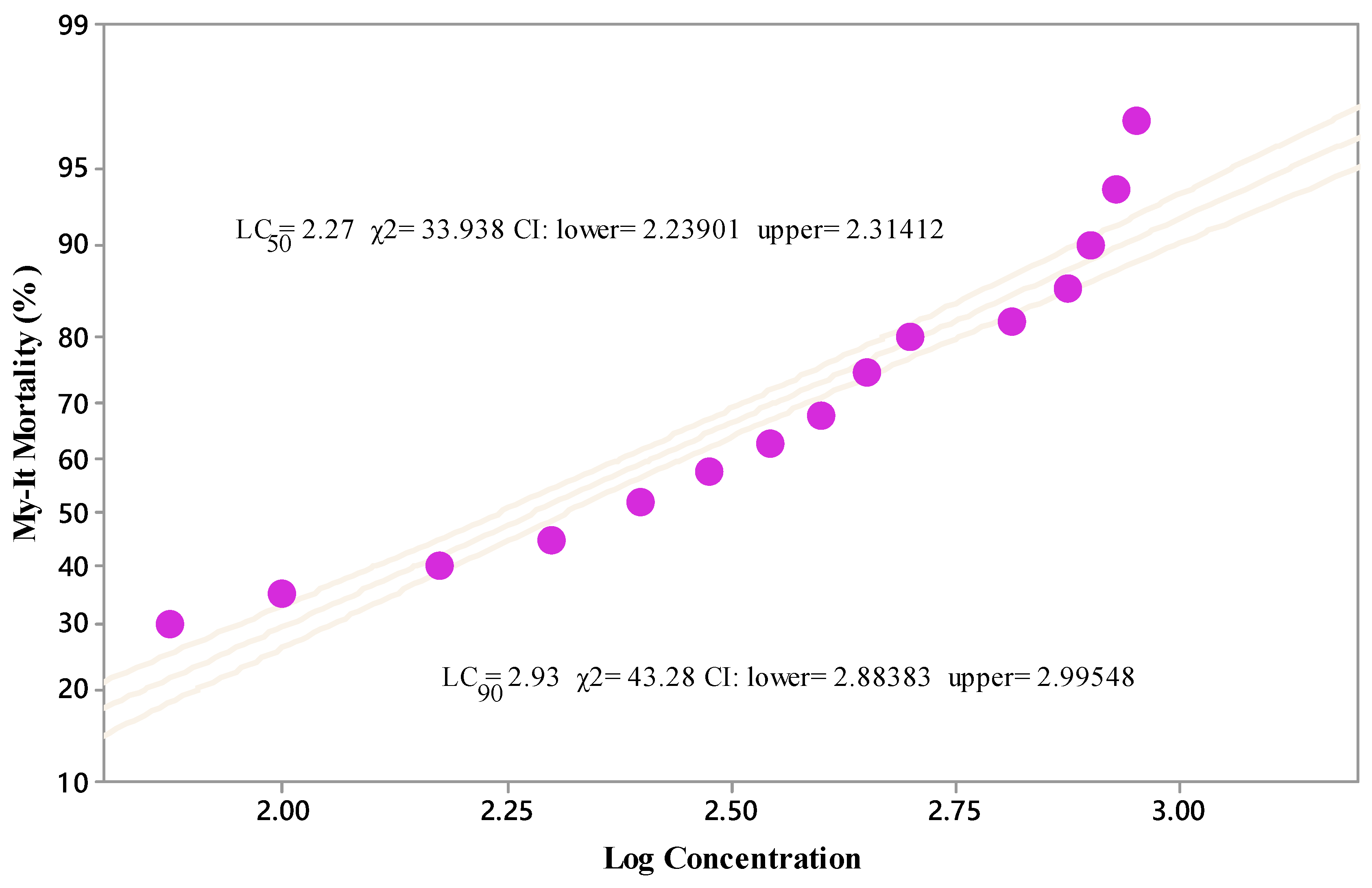
3.1. Effect of My-It on Mosquito Survival
3.2. Oviposition Deterrence Index
3.3. Fecundity of My-It
3.4. Enzyme Inhibition of My-It
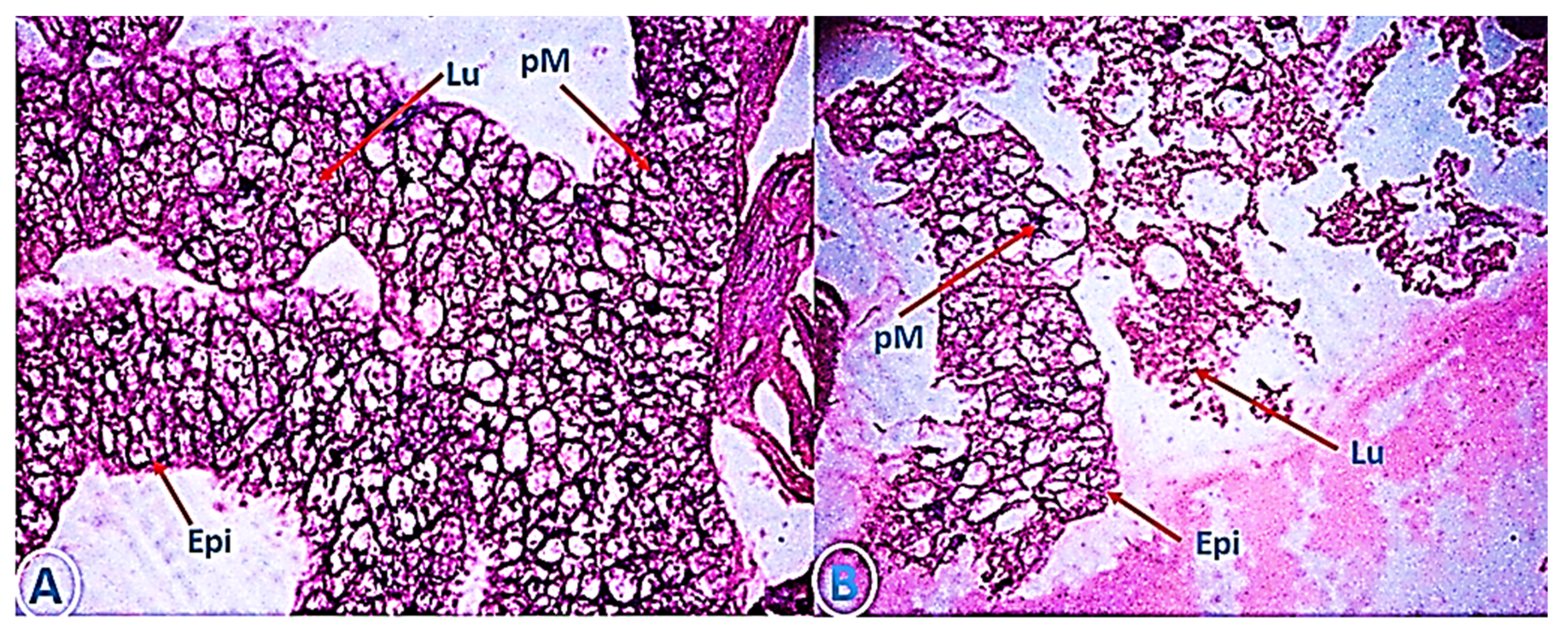
3.5. The Efficacy of of My-It on Gut-Histology
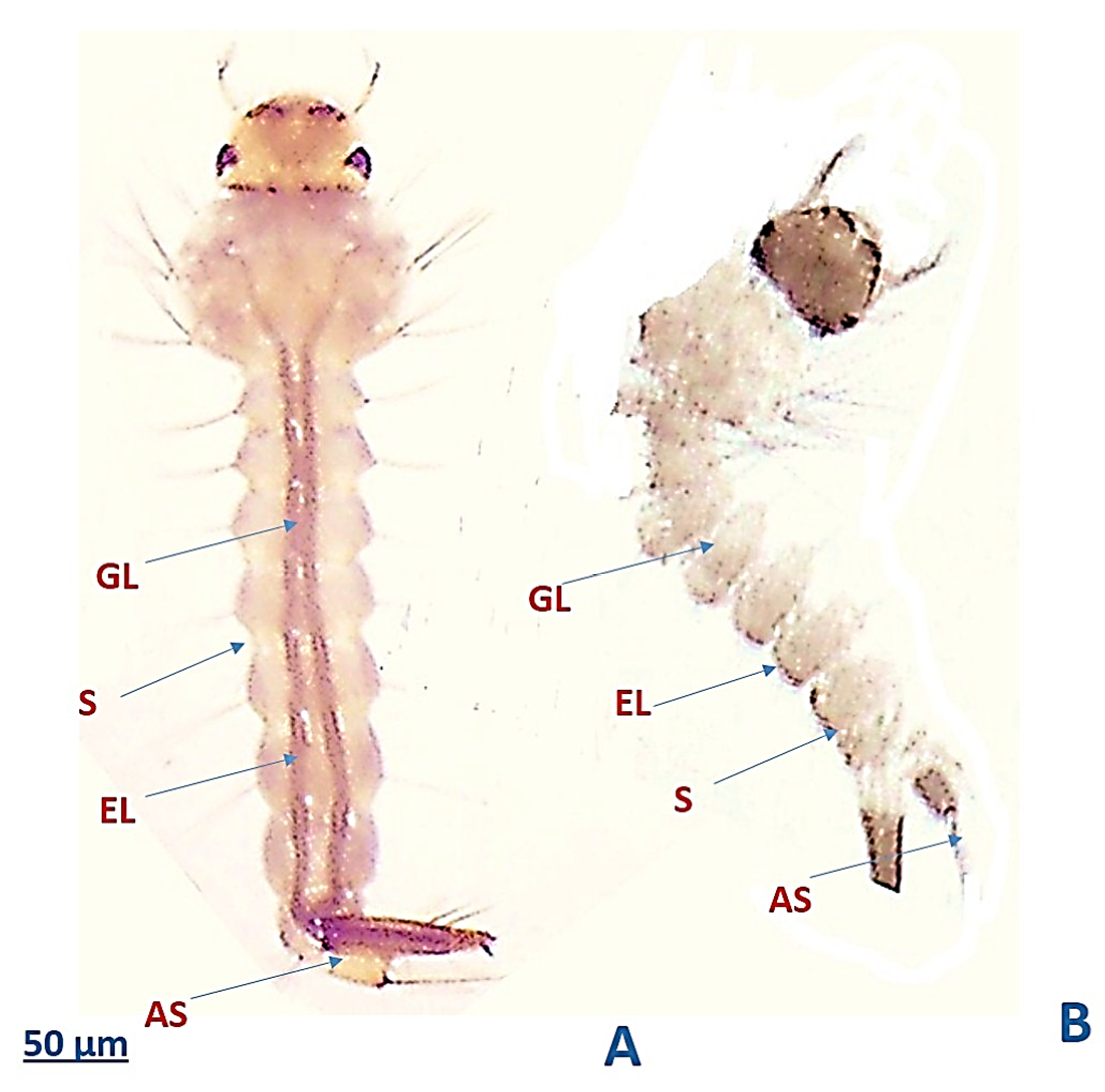
3.6. The Efficacy of My-It on the External Physiology of Ae. aegypti Larvae
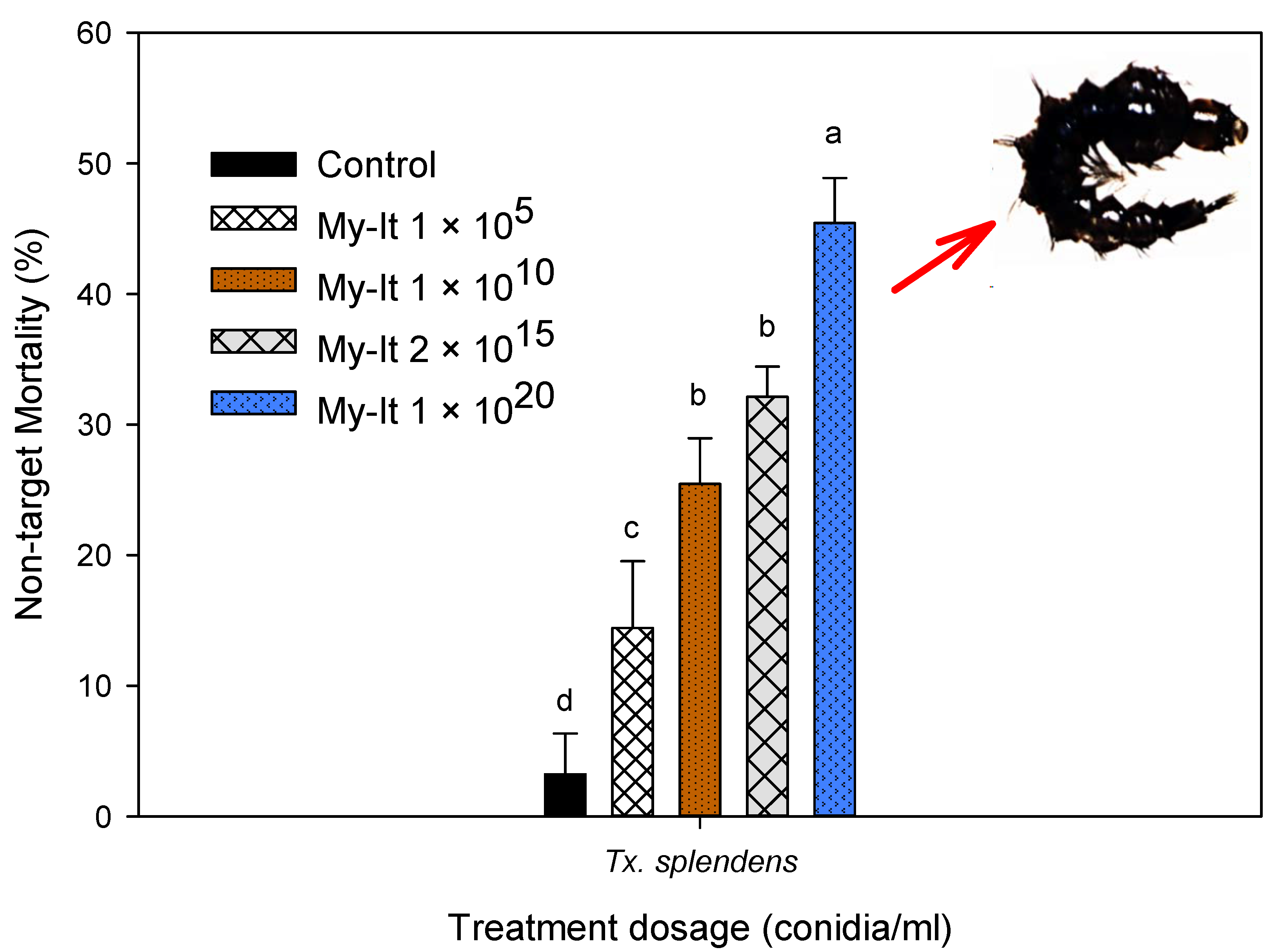
3.7. Non-Target Toxicity of My-It
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khater, H.F. Introductory Chapter: Back to the Future: Solutions for Parasitic Problems as Old as Pyramids. In Natural Remedies in the Fight against Parasites; InTech: Rijeka, Croatia, 2017. [Google Scholar]
- Benelli, G.; Mehlhorn, H. Declining malaria, rising of dengue and Zika virus: Insights for mosquito vector control. Parasitol. Res. 2016, 115, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G. Research in mosquito control: Current challenges for a brighter future. Parasitol. Res. 2015, 114, 2801–2805. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Nathan, S. A Review of Resistance Mechanisms of Synthetic Insecticides and Botanicals, Phytochemicals, and Essential Oils as Alternative Larvicidal Agents Against Mosquitoes. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue and Severe Dengue (Fact Sheet N°117). 2016. Available online: http://www.who.int/mediacentre/factsheets/fs117/en/ (accessed on 24 June 2016).
- Autran, E.; Neves, I.; DaSilva, C.; Santos, G.; Camara, C.; Navarro, D. Chemical composition, oviposition deterrent and larvicidal activities against Aedes aegypti of essential oils from Piper marginatum Jacq. (Piperaceae). Bioresour. Technol. 2009, 100, 2284–2288. [Google Scholar] [CrossRef] [PubMed]
- Khater, H.F. Bioactivity of essential oils as green biopesticides: Recent global scenario. Rec. Prog. Med. Plants 2013, 37, 151–218. [Google Scholar]
- Govindarajan, M.; Khater, H.F.; Panneerselvam, C.; Benelli, G. One-pot fabrication of silver nanocrystals using Nicandra physalodes: A novel route for mosquito vector control with moderate toxicity on non-target water bugs. Res. Veter Sci. 2016, 107, 95–101. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Kalaivani, K.; Murugan, K. Effects of neem limonoids on the malaria vector Anopheles stephensi Liston (Diptera: Culicidae). Acta Trop. 2005, 96, 47–55. [Google Scholar] [CrossRef]
- Khater, H.F. Prospects of botanical biopesticides in insect pest management. Pharmacologia 2012, 3, 641–656. [Google Scholar]
- Khater, H.F.; Selim, A.M.; Abouelella, G.A.; Abouelella, N.A.; Murugan, K.; Vaz, N.P.; Govindarajan, M. Commercial Mosquito Repellents and Their Safety Concerns. In Malaria [Working Title]; IntechOpen: London, UK, 2019. [Google Scholar]
- Khater, H.F.; Shalaby, A.A.-S. Potential of biologically active plant oils to control mosquito larvae (Culex pipiens, Diptera: Culicidae) from an egyptian locality. Rev. Inst. Med. Trop. São Paulo 2008, 50, 107–112. [Google Scholar] [CrossRef]
- Chellappandian, M.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Karthi, S.; Thanigaivel, A.; Ponsankar, A.; Kalaivani, K.; Hunter, W.B. Botanical essential oils and uses as mosquitocides and repellents against dengue. Environ. Int. 2018, 113, 214–230. [Google Scholar] [CrossRef]
- Govindarajan, M.; Jebanesan, A.; Reetha, D. Larvicidal effect of extracellular secondary metabolites of different fungi against the mosquito, Culex quinquefasciatus Say. Trop. Biomed. 2005, 22, 1–3. [Google Scholar]
- Vasantha-Srinivasan, P.; Karthi, S.; Chellappandian, M.; Ponsankar, A.; Thanigaivel, A.; Senthil-Nathan, S.; Chandramohan, D.; Ganesan, R. Aspergillus flavus (Link) toxins reduces the fitness of dengue vector Aedes aegypti (Linn.) and their non-target toxicity against aquatic predator. Microb. Pathog. 2019, 128, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Karthi, S.; Vaideki, K.; Shivakumar, M.S.; Ponsankar, A.; Thanigaivel, A.; Chellappandian, M.; Vasantha-Srinivasan, P.; Chanthini, K.M.; Hunter, W.B.; Senthil-Nathan, S. Effect of on the mortality of Aspergillus flavus and activity of antioxidant enzymes of Spodoptera litura Fab. (Lepidoptera: Noctuidae) larvae. Pestic. Biochem. Physiol. 2018, 149, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Kirubakaran, S.A.; Sathish-Narayanan, S.; Revathi, K.; Chandrasekaran, R.; Senthil-Nathan, S. Effect of oil-formulated Metarhizium anisopliae and Beauveria bassiana against the rice leaffolder Cnaphalocrocis medinalis Guenée (Lepidoptera: Pyralidae). Arch. Phytopathol. Plant Prot. 2013, 47, 977–992. [Google Scholar] [CrossRef]
- Khater, H.F. Biocontrol of Some Insects. Ph.D. Thesis, Zagazig University, Benha Branch, Egypt, 2003. [Google Scholar]
- Lu, Y.; Luo, F.; Cen, K.; Yin, Y.; Zhan, S.; Wang, C.; Zhang, H.; Xiao, G.; Li, C.; Li, Z. Omics data reveal the unusual asexual-fruiting nature and secondary metabolic potentials of the medicinal fungus Cordyceps cicadae. BMC Genom. 2017, 18, 668. [Google Scholar] [CrossRef]
- Weng, S.-C.; Chou, C.-J.; Lin, L.-C.; Tsai, W.-J.; Kuo, Y.-C. Immunomodulatory functions of extracts from the Chinese medicinal fungus Cordyceps cicadae. J. Ethnopharmacol. 2002, 83, 79–85. [Google Scholar] [CrossRef]
- Hsu, J.H.; Jhou, B.Y.; Yeh, S.H.; Chen, Y.I.; Chen, C.C. Healthcare Functions of Cordyceps cicadae. J. Nutr. Food Sci. 2015, 5. [Google Scholar] [CrossRef]
- Olatunji, O.J.; Feng, Y.; Tang, J.; Olatunji, O.; Ouyang, Z.; Su, Z. Cordycepin protects PC12 cells against 6-hydroxydopamine induced neurotoxicity via its antioxidant properties. Biomed Pharmacother. 2016, 81, 7–14. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Hwang, Y.-S.; Schultz, G.W.; Axelrod, H.; Kramer, W.L.; Mulla, M.S. Ovipositional Repellency of Fatty Acids and Their Derivatives Against Culex1 and Aedes1 Mosquitoes. Environ. Entomol. 1982, 11, 223–226. [Google Scholar] [CrossRef]
- Napoleão, T.H.; Pontual, E.V.; Lima, T.D.A.; Santos, N.D.D.L.; Sá, R.A.; Coelho, L.C.B.B.; Navarro, D.M.D.A.F.; Paiva, P. Effect of Myracrodruon urundeuva leaf lectin on survival and digestive enzymes of Aedes aegypti larvae. Parasitol. Res. 2011, 110, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Thanigaivel, A.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Edwin, E.; Ponsankar, A.; Selin-Rani, S.; Pradeepa, V.; Chellappandian, M.; Kalaivani, K.; Abdel-Megeed, A.; et al. Chemicals isolated from Justicia adhatoda Linn reduce fitness of the mosquito, Aedes aegypti L. Arch. Insect Biochem. Physiol. 2017, 94, e21384. [Google Scholar] [CrossRef] [PubMed]
- Coelho, J.S.; Santos, N.D.; Napoleão, T.H.; Gomes, F.S.; Ferreira, R.S.; Zingali, R.B.; Coelho, L.; Leite, S.P.; Navarro, D.M.; Paiva, P. Effect of Moringa oleifera lectin on development and mortality of Aedes aegypti larvae. Chemosphere 2009, 77, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Allison, P.D. Survival Analysis Using the SAS System: A Practical Guide; SAS Institute: Cary, NC, USA, 1995. [Google Scholar]
- Rodriguez, M.M.; Bisset, J.A.; Fernández, D. Levels of insecticide resistance and resistance mechanisms in aedes aegypti from some latin american countries. J. Am. Mosq. Control. Assoc. 2007, 23, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crop. Prod. 2015, 76, 174–187. [Google Scholar] [CrossRef]
- Vasantha-Srinivasan, P.; Thanigaivel, A.; Edwin, E.-S.; Ponsankar, A.; Senthil-Nathan, S.; Selin-Rani, S.; Kalaivani, K.; Hunter, W.B.; Duraipandiyan, V.; Al-Dhabi, N.A. Toxicological effects of chemical constituents from Piper against the environmental burden Aedes aegypti Liston and their impact on non-target toxicity evaluation against biomonitoring aquatic insects. Environ. Sci. Pollut. Res. 2017, 25, 10434–10446. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Choi, M.-Y.; Paik, C.-H.; Seo, H.-Y. Food consumption, utilization, and detoxification enzyme activity of the rice leaffolder larvae after treatment with Dysoxylum triterpenes. Pestic. Biochem. Physiol. 2007, 88, 260–267. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. Physiological and biochemical effect of neem and other Meliaceae plants secondary metabolites against Lepidopteran insects. Front. Physiol. 2013, 4, 1–17. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A review of bio pesticides and their mode of action against insect pests. In Environmental Sustainability-Role of Green Technologies; Springer: Berlin, Germany, 2015; pp. 49–63. [Google Scholar]
- Mishra, P.K.; Shukla, R.; Singh, P.; Prakash, B.; Dubey, N.K. Antifungal and antiaflatoxigenic efficacy of Caesulia axillaris Roxb. essential oil against fungi deteriorating some herbal raw materials, and its antioxidant activity. Ind. Crop. Prod. 2012, 36, 74–80. [Google Scholar] [CrossRef]
- Holighaus, G.; Rohlfs, M. Fungal allelochemicals in insect pest management. Appl. Microbiol. Biotechnol. 2016, 100, 5681–5689. [Google Scholar] [CrossRef]
- Luangsa-Ard, J.J.; Hywel-Jones, N.L.; Manoch, L.; Samson, R.A. On the relationships of Paecilomyces sect. Isarioidea species. Mycol. Res. 2005, 109, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Guo, S.; Wang, W.; Liu, X. Cordyceps industry in China. Mycological 2015, 6, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Vega-Aquino, P.; Sánchez-Peña, S.; Blanco, C.A. Activity of oil-formulated conidia of the fungal entomopathogens Nomuraea rileyi and Isaria tenuipes against lepidopterous larvae. J. Invertebr. Pathol. 2010, 103, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Butt, T.; Greenfield, B.P.J.; Greig, C.; Maffeis, T.G.G.; Taylor, J.; Piasecka, J.; Dudley, E.; Abdulla, A.; Dubovskiy, I.M.; Jurado, I.G.; et al. Metarhizium anisopliae Pathogenesis of Mosquito Larvae: A Verdict of Accidental Death. PLoS ONE 2013, 8, e81686. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Muturi, E.J.; Dunlap, C.; Rooney, A.P. Strain-specific pathogenicity and subversion of phenoloxidase activity in the mosquito Aedes aegypti by members of the fungal entomopathogenic genus Isaria. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Soonwera, M. Efficacy of essential oil from Cananga odorata (Lamk.) Hook.f. & Thomson (Annonaceae) against three mosquito species Aedes aegypti (L.), Anopheles dirus (Peyton and Harrison), and Culex quinquefasciatus (Say). Parasitol. Res. 2015, 114, 4531–4543. [Google Scholar] [CrossRef]
- Tasin, M.; Knudsen, G.K.; Pertot, I. Smelling a diseased host: Grapevine moth responses to healthy and fungus-infected grapes. Anim. Behav. 2012, 83, 555–562. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; Jermy, T.; Van Loon, J.J.A. Insect-Plant Biology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Scholte, E.-J.; Knols, B.G.; Takken, W. Infection of the malaria mosquito Anopheles gambiae with the entomopathogenic fungus Metarhizium anisopliae reduces blood feeding and fecundity. J. Invertebr. Pathol. 2006, 91, 43–49. [Google Scholar] [CrossRef]
- Ondiaka, S.; Bukhari, T.; Farenhorst, M.; Takken, W.; Knols, G.J. Effects of fungal infection on the host-seeking behaviour and fecundity of the malaria mosquito Anopheles gambiae Giles. Proc. Neth. Entomol. Soc. Meet. 2008, 19, 121–128. [Google Scholar]
- Senthil-Nathan, S.; Choi, M.Y.; Paik, C.H.; Seo, H.Y.; Kalivani, K.; Kim, J.D. Effect of azadirachtin on acetylcholinesterase (AChE) activity and histology of the brown plant hopper Nilaparvata lugens (Stal). Ecotoxicol. Environ. Saf. 2008, 70, 244–250. [Google Scholar] [CrossRef]
- Ross, M.K.; Streit, T.M.; Herring, K.L.; Xie, S. Carboxylesterases: Dual roles in lipid and pesticide metabolism. J. Pestic. Sci. 2010, 35, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Ranson, H. Insecticide Resistance in Insect Vectors of Human Disease. Ann. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef]
- Rudin, W.; Hecker, H. Lectin-binding sites in the midgut of the mosquitoes Anopheles stephensi Liston and Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res. 1989, 75, 268–279. [Google Scholar]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2014, 22, 68–102. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Vasantha-Srinivasan, P.; Edwin, E.-S.; Ponsankar, A.; Selin-Rani, S.; Chellappandian, M.; Kalaivani, K.; Senthil-Nathan, S.; Benelli, G. Development of an eco-friendly mosquitocidal agent from Alangium salvifolium against the dengue vector Aedes aegypti and its biosafety on the aquatic predator. Environ. Sci. Pollut. Res. 2017, 25, 10340–10352. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karthi, S.; Vasantha-Srinivasan, P.; Ganesan, R.; Ramasamy, V.; Senthil-Nathan, S.; Khater, H.F.; Radhakrishnan, N.; Amala, K.; Kim, T.-J.; El-Sheikh, M.A.; et al. Target Activity of Isaria tenuipes (Hypocreales: Clavicipitaceae) Fungal Strains against Dengue Vector Aedes aegypti (Linn.) and Its Non-Target Activity Against Aquatic Predators. J. Fungi 2020, 6, 196. https://doi.org/10.3390/jof6040196
Karthi S, Vasantha-Srinivasan P, Ganesan R, Ramasamy V, Senthil-Nathan S, Khater HF, Radhakrishnan N, Amala K, Kim T-J, El-Sheikh MA, et al. Target Activity of Isaria tenuipes (Hypocreales: Clavicipitaceae) Fungal Strains against Dengue Vector Aedes aegypti (Linn.) and Its Non-Target Activity Against Aquatic Predators. Journal of Fungi. 2020; 6(4):196. https://doi.org/10.3390/jof6040196
Chicago/Turabian StyleKarthi, Sengodan, Prabhakaran Vasantha-Srinivasan, Raja Ganesan, Venkatachalam Ramasamy, Sengottayan Senthil-Nathan, Hanem F. Khater, Narayanaswamy Radhakrishnan, Kesavan Amala, Tae-Jin Kim, Mohamed A. El-Sheikh, and et al. 2020. "Target Activity of Isaria tenuipes (Hypocreales: Clavicipitaceae) Fungal Strains against Dengue Vector Aedes aegypti (Linn.) and Its Non-Target Activity Against Aquatic Predators" Journal of Fungi 6, no. 4: 196. https://doi.org/10.3390/jof6040196
APA StyleKarthi, S., Vasantha-Srinivasan, P., Ganesan, R., Ramasamy, V., Senthil-Nathan, S., Khater, H. F., Radhakrishnan, N., Amala, K., Kim, T.-J., El-Sheikh, M. A., & Krutmuang, P. (2020). Target Activity of Isaria tenuipes (Hypocreales: Clavicipitaceae) Fungal Strains against Dengue Vector Aedes aegypti (Linn.) and Its Non-Target Activity Against Aquatic Predators. Journal of Fungi, 6(4), 196. https://doi.org/10.3390/jof6040196