Diversity of Linear Non-Ribosomal Peptide in Biocontrol Fungi



Abstract
1. Introduction
2. L-NRPs from Entomopathogenic Fungi
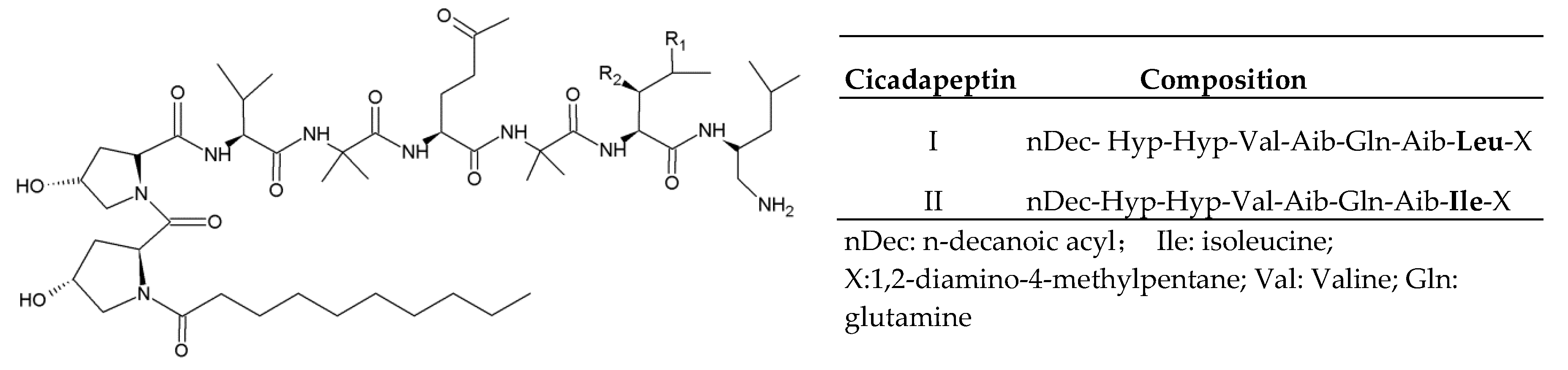
2.1. Cicadapeptins
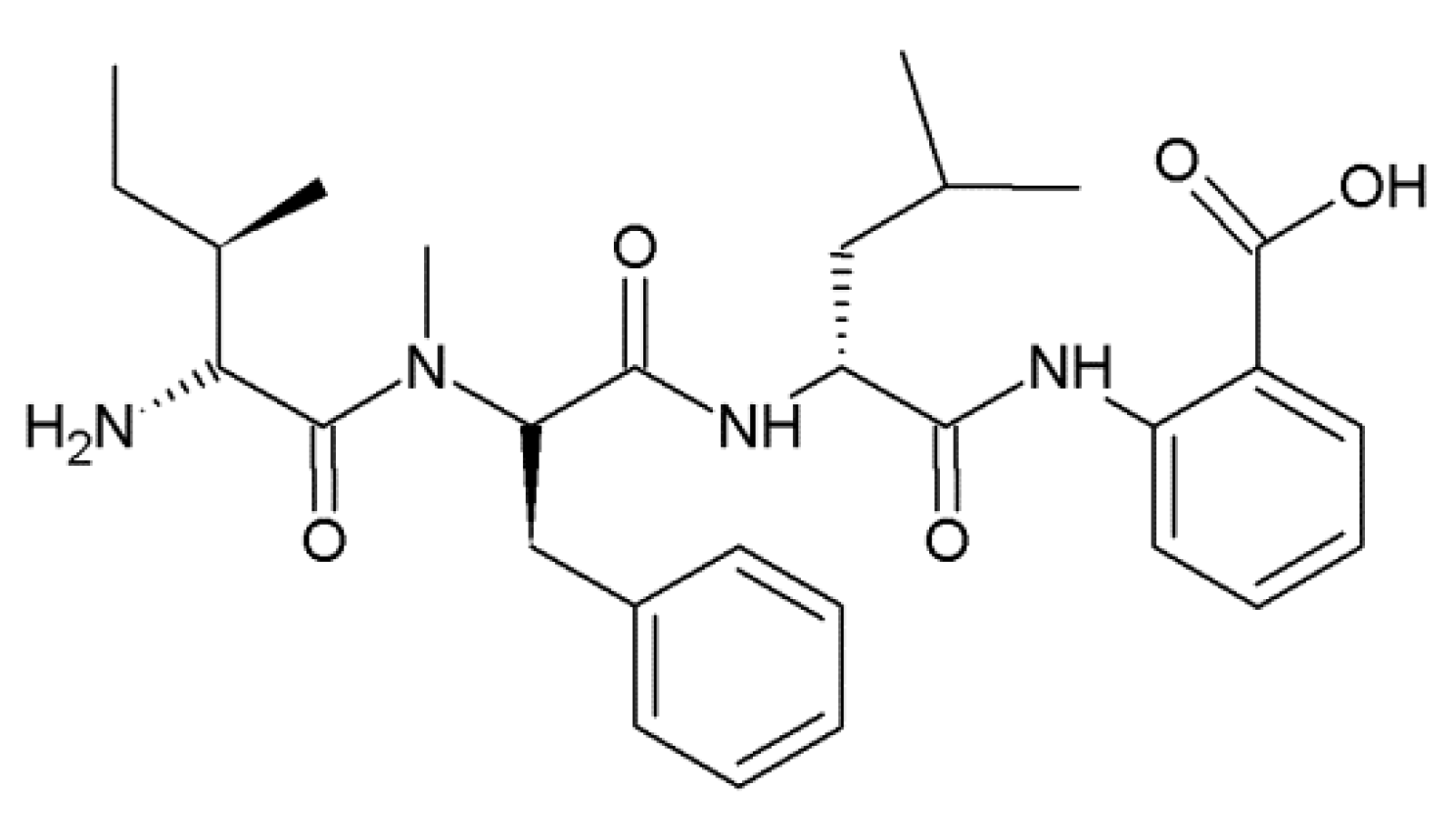
2.2. Hirsutellic Acid A
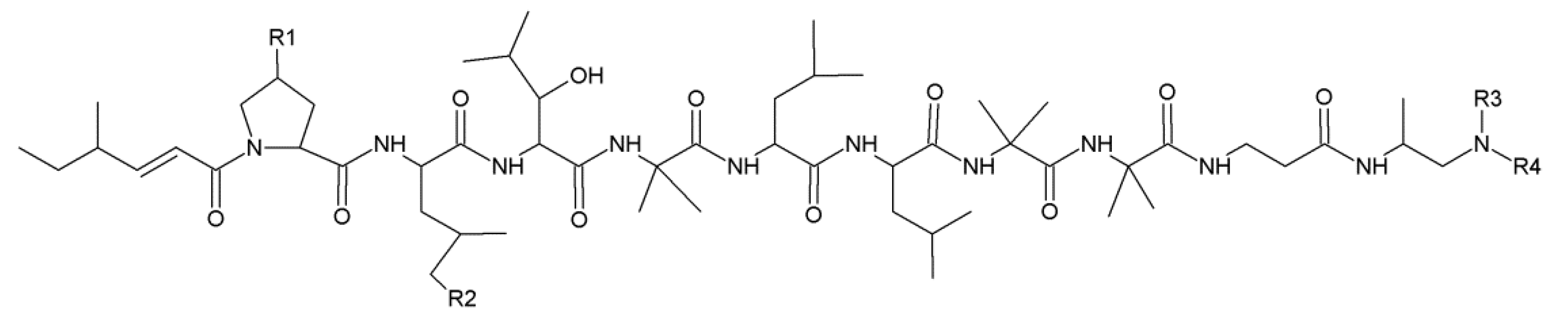
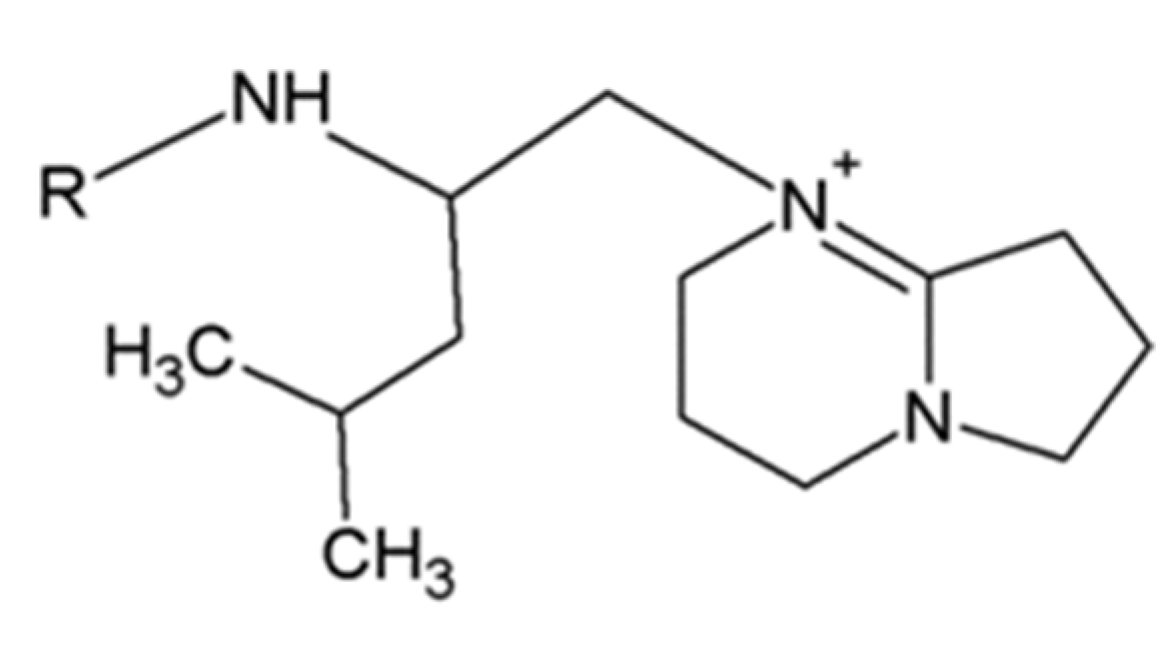
2.3. Leucinostatins
2.4. Efrapeptins
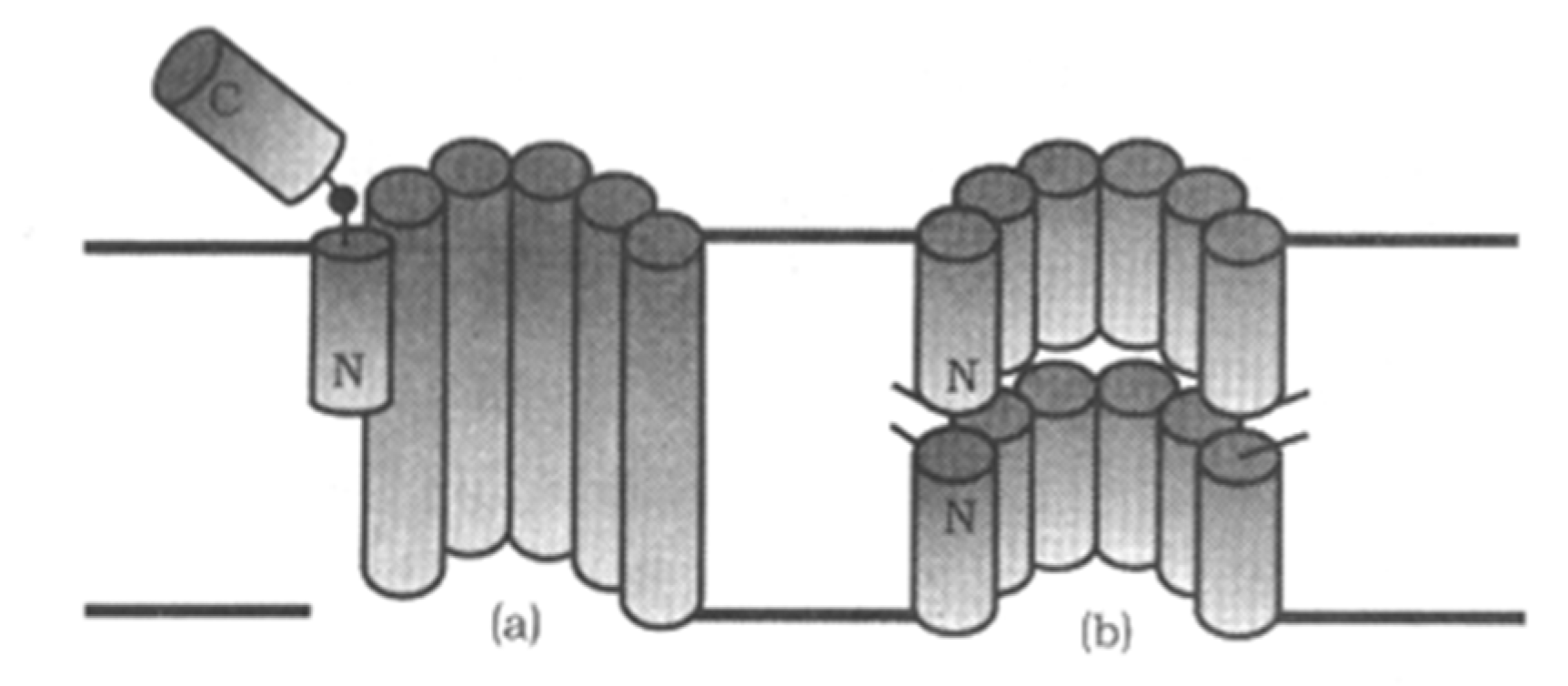
2.5. Peptaibol Compounds
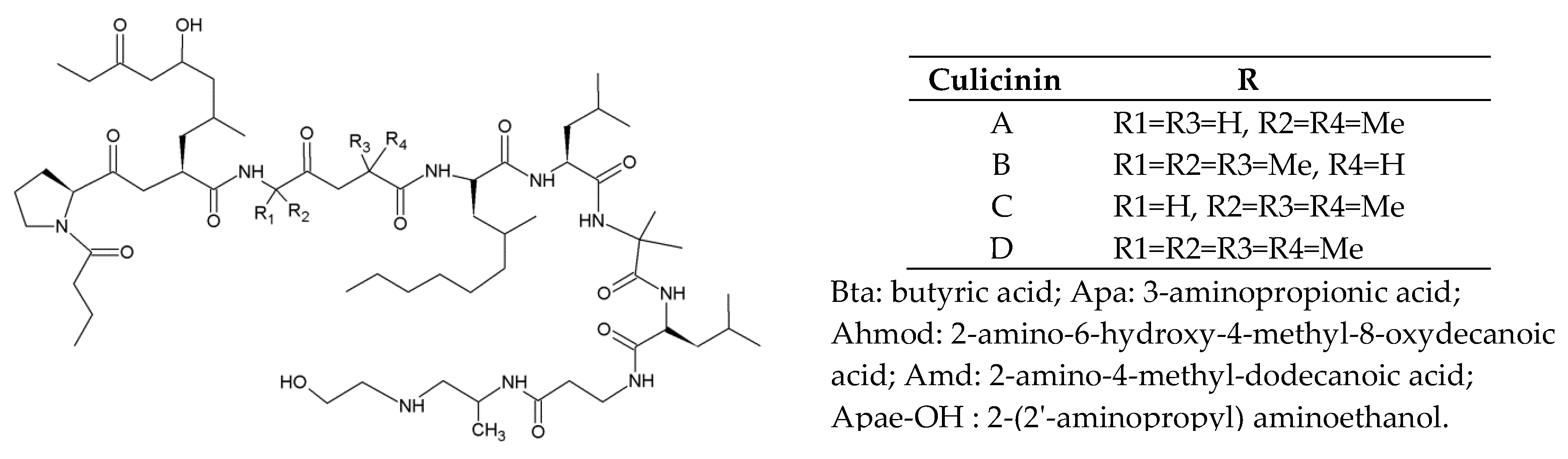
2.5.1. Culicinins
2.5.2. LP237
2.5.3. Metanicins
3. L-NRPs from Mycoparasitic Fungi
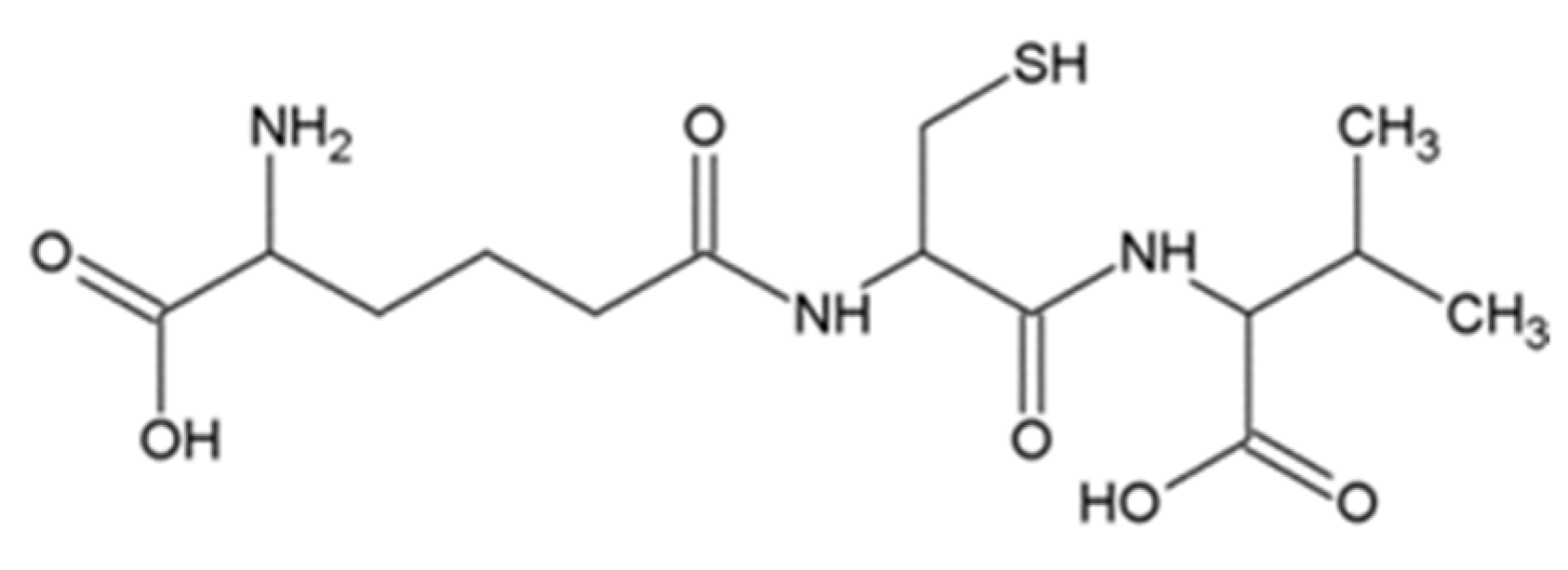
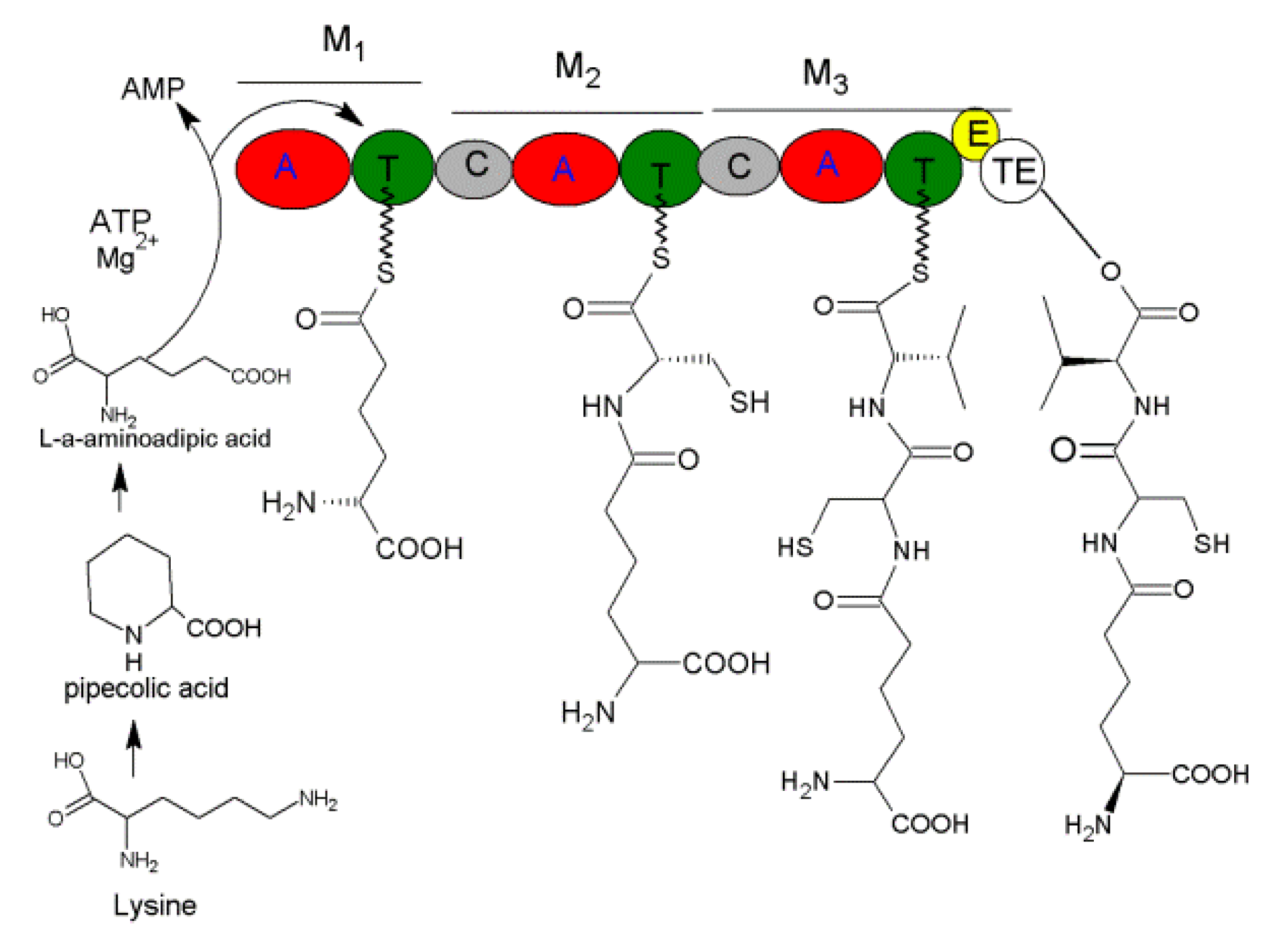
3.1. ACV
3.2. Harzianins
3.3. Trichorzins
3.4. Longibrachins
3.5. Trichobrachins
3.6. Trichogins
3.7. Trilongins
3.8. Trichokonins
3.9. Alamethicins
3.10. Trichotoxins
3.11. Suzukacillins
3.12. Hypomurocins
3.13. Atroviridins and Neoatroviridins
3.14. Peptaivirin
4. Biosynthesis of L-NRPs
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, X.F.; Hu, Q.B.; Weng, Q.F. Secondary metabolites (SMs) of Isaria cicadae and Isaria tenuipes. RSC Adv. 2019, 9, 172–184. [Google Scholar] [CrossRef]
- Weng, Q.F.; Zhang, X.F.; Chen, W.; Hu, Q.B. Secondary metabolites and the risks of Isaria fumosorosea and Isaria farinosa. Molecules 2019, 24, 664. [Google Scholar] [CrossRef]
- Marian, M.; Shimizu, M. Improving performance of microbial biocontrol agents against plant diseases. J. Gen. Plant Pathol. 2019, 85, 329–336. [Google Scholar] [CrossRef]
- Silva, R.N.; Monteiro, V.N.; Steindorff, A.S.; Gomes, E.V.; Noronha, E.F.; Ulhoa, C.J. Trichoderma/pathogen/plant interaction in pre-harvest food security. Fungal Biol. 2019, 123, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Süssmuth, R.D.; Mainz, A. Nonribosomal peptide synthesis-principles and prospects. Angew. Chem. Int. Ed. Engl. 2017, 56, 3770–3821. [Google Scholar] [CrossRef]
- Xue, G.; Stuart, W.H.; Brian, D.A.; Peng, W.; Linda, P.V.; Christopher, T.W.; Yi, T. Cyclization of fungal nonribosomal peptides by a terminal condensation-like domain. Nat. Chem. Biol. 2012, 8, 823–830. [Google Scholar]
- Matthias, S.; Alan, T.; Mohamed, A.M. Nonribosomal peptide synthetases: Structures and dynamics. Curr. Opin. Struct. Biol. 2010, 20, 234–240. [Google Scholar]
- Bradley, R.M.; Andrew, M.G. Structural biology of nonribosomal peptide synthetases. Methods Mol. Biol. 2016, 1401, 3–29. [Google Scholar]
- Hu, Q.; Dong, T. Chapter 8 Non-ribosomal Peptides from Entomogenous Fungi. In Biocontrol of Lepidoperan Pests—Use of Soil Microbes and their Metabolites; Sree, K.S., Varma, A., Eds.; Springer: Cham, Switzerland, 2015; Volume 43, pp. 169–206. [Google Scholar] [CrossRef]
- Krasnoff, S.B.; Reategui, R.F.; Wagenaar, M.M.; Gloer, J.B.; Gibson, D.M. Cicadapeptins I and II: New aib-containing peptides from the entomopathogenic fungus Cordyceps heteropoda. J. Nat. Prod. 2005, 68, 50–55. [Google Scholar] [CrossRef]
- Molnár, I.; Gibson, D.M.; Krasnoff, S.B. Secondary metabolites from entomopathogenic Hypocrealean fungi. Nat. Prod. Rep. 2010, 27, 1241–1275. [Google Scholar] [CrossRef]
- Thongtan, J.; Saenboonrueng, J.; Rachtawee, P.; Isaka, M. An antimalarial tetrapeptide from the entomopathogenic fungus Hirsutella sp. BCC 1528. J. Nat. Prod. 2006, 69, 713–714. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.F.C.; Moraes, L.A.B. Liquid chromatography-tandem mass spectrometry characterization of five new leucinostatins produced by Paecilomyces lilacinus CG—189. J. Antibiot. 2015, 68, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Radics, L.; Kajtarperedy, M.; Casinovi, C.G.; Rossi, C.; Ricci, M.; Tuttobello, L. Leucinostatins H and K, two novel peptide antibiotics with tertiary amine-oxide terminal group from Paecilomyces marquandii isolation, structure and biological activity. J. Antibiot. 1987, 40, 714–716. [Google Scholar] [PubMed]
- Strobel, G.A.; Torczynski, R.; Bollon, A. Acremonium sp.: A leucinostatin A producing endophyte of European yew (Taxus baccata). Plant Sci. 1997, 128, 97–108. [Google Scholar]
- Shima, A.; Fukushima, K.; Arai, T.; Terada, H. Dual inhibitory effects of the peptide antibiotics leucinostatins on oxidative phosphorylation in mitochondria. Cell Struct. Funct. 1990, 15, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Park, J.O.; Hargreaves, J.R.; Mcconville, E.J.; Stirling, G.R.; Ghisalberti, E.L.; Sivasithamparam, K. Production of leucinostatins and nematicidal activity of Australian isolates of Paecilomyces lilacinus (Thom) Samson. Lett. Appl. Microbiol. 2010, 38, 271–276. [Google Scholar] [CrossRef]
- Manabu, K.; Hiroyuki, I.; Shun-Ichi, O.; Tohru, M.; Isao, M.; Daishiro, I. Leucinostatin A inhibits prostate cancer growth through reduction of insulin-like growth factor-I expression in prostate stromal cells. Int. J. Cancer 2010, 126, 810–818. [Google Scholar]
- Krasnoff, S.B.; Gupta, S. Identification and directed biosynthesis of efrapeptins in the fungus Tolypocladium geodes gams (Deuteromycotina: Hyphomycetes). J. Chem. Ecol. 1991, 17, 1953–1962. [Google Scholar] [CrossRef]
- Krasnoff, S.B.; Gupta, S.; Leger, R.J.S.; Renwick, J.A.A.; Roberts, D.W. Antifungal and insecticidal properties of the efrapeptins: Metabolites of the fungus Tolypocladium niveum. J. Invertebr. Pathol. 1991, 58, 180–188. [Google Scholar] [CrossRef]
- Meyling, N.V.; Schmidt, N.M.; Eilenberg, J. Occurrence and diversity of fungal entomopathogens in soils of low and high Arctic Greenland. Polar Biol. 2012, 35, 1439–1445. [Google Scholar] [CrossRef]
- Boot, C.M.; Amagata, T.; Tenney, K.; Compton, J.E.; Pietraszkiewicz, H.; Valeriote, F.A.; Crews, P. Four classes of structurally unusual peptides from two marine-derived fungi: Structures and bioactivities. Tetrahedron 2007, 63, 9903–9914. [Google Scholar] [CrossRef] [PubMed]
- Papathanassiu, A.E.; MacDonald, N.J.; Emlet, D.R.; Hong, A.V. Antitumor activity of efrapeptins, alone or in combination with 2-deoxyglucose, in breast cancer in vitro and in vivo. Cell Stress Chaperon. 2011, 16, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, J.P.; Buchanan, S.K.; Raaij, M.J.V.; Fearnley, I.M.; Walker, J.E. The structure of bovine F1-ATPase complexed with the peptide antibiotic efrapeptin. Proc. Natl. Acad. Sci. USA 1996, 93, 9420–9424. [Google Scholar] [CrossRef] [PubMed]
- Papathanassiu, A.E.; MacDonald, N.J.; Bencsura, A.; Vu, H.A. F1F0-ATP synthase functions as a co-chaperone of HSP90—Substrate protein complexes. Biochem. Biophys. Res. Commun. 2006, 345, 420–429. [Google Scholar] [CrossRef]
- Laurence, H.P. Review: The HSP90 molecular chaperone—An enigmatic ATPase. Biopolymers 2016, 105, 594–607. [Google Scholar]
- Bandani, A.R.; Amiri, B.; Butt, T.M.; Gordon-Weeks, R. Effects of efrapeptin and destruxin, metabolites of entomogenous fungi, on the hydrolytic activity of a vacuolar type ATPase identified on the brush border membrane vesicles of Galleria mellonella midgut and on plant membrane bound hydrolytic enzymes. BBA Biomembranes 2001, 1510, 367–377. [Google Scholar] [CrossRef]
- Yoichi, H.; Yuki, H.; Takashi, K.; Kaneo, K.; Kyoko, A.; Yoshikazu, S.; Kazuo, S.Y. Efrapeptin J, a new down-regulator of the molecular chaperone GRP78 from a marine Tolypocladium sp. J. Antibiot. 2008, 61, 365–371. [Google Scholar]
- Szekeres, A.; Leiteb, B.; Kredics, L.; Antal, Z.; Hatvani, L.; Manczinger, L.; Vágvölgyi, C. Peptaibols and related peptaibiotics of Trichoderma. Acta Microbiol. Immunol. Hung. 2005, 52, 137–168. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, G.S. APD: The antimicrobial peptide database. Nucleic Acids Res. 2004, 32, 590–592. [Google Scholar] [CrossRef]
- Zhao, P.; Xue, Y.; Li, X.; Li, J.; Zhao, Z.; Quan, C.; Gao, W.; Zu, X.; Bai, X.; Feng, S. Fungi-derived lipopeptide antibiotics developed since 2000. Peptides 2019, 113, 52–65. [Google Scholar] [CrossRef]
- Higashimoto, Y.; Kodama, H.; Jelokhani-Niaraki, M.; Kato, F.; Kondo, M. Structure-function relationship of model Aib-containing peptides as ion transfer intermembrane templates. J. Biochem. 1999, 125, 705–712. [Google Scholar] [CrossRef]
- Miick, S.M.; Martinez, G.V.; Fiori, W.R.; Todd, A.P.; Millhauser, G.L. Short alanine-based peptides may form 310-helices and not α-helices in aqueous solution. Nature 1992, 359, 653–655. [Google Scholar] [CrossRef] [PubMed]
- Chugh, J.K.; Wallace, B.A. Peptaibols: Models for ion channels. Biochem. Soc. Trans. 2001, 29, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Poirier, L.; Quiniou, F.; Ruiz, N.; Montagu, M.; Amiard, J.C.; Pouchus, Y.F. Toxicity assessment of peptaibols and contaminated sediments on Crassostrea gigas embryos. Aquat. Toxicol. 2007, 83, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Peltola, J.; Ritieni, A.; Mikkola, R.; Grigoriev, P.A.; Pócsfalvi, G.; Andersson, M.A.; Salkinojasalonen, M.S. Biological Effects of Trichoderma harzianum Peptaibols on mammalian Cells. Appl. Environ. Microbiol. 2004, 70, 4996–5004. [Google Scholar] [CrossRef] [PubMed]
- Unkles, S.E.; Marriott, C.; Kinghorn, J.R.; Panter, C.; Blackwell, A. Efficacy of the entomopathogenic fungus, Culicinomyces clavisporus against larvae of the biting midge, Culicoides nubeculosus (Diptera: Ceratopogonidae). Biocontrol Sci. Technol. 2004, 14, 397–401. [Google Scholar] [CrossRef]
- Haiyin, H.; Janso, J.E.; Hui, Y.; Yang; Bernan, V.S.; Lin, S.L.; Ker, Y. Culicinin D, an antitumor peptaibol produced by the fungus Culicinomyces clavisporus, strain LL-12I252. J. Nat. Prod. 2006, 69, 736–741. [Google Scholar]
- Kebede, B.; Wrigley, S.K.; Prashar, A.; Rahlff, J.; Wolf, M.; Reinshagen, J.; Gribbon, P.; Imhoff, J.F.; Silber, J.; Labes, A. Establishing the secondary metabolite profile of the marine fungus:Tolypocladium geodes sp. MF458 and subsequent optimisation of bioactive secondary metabolite production. Mar. Drugs 2017, 15, 84. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S.; Pischos, S.; Sauriol, F. Structural assignment of the peptide antibiotic LP237-F8, a metabolite of\r, Tolypocladium geodes. J. Org. Chem. 1996, 61, 2118–2121. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S.; Pischos, S.; Sauriol, F.; Widden, P. Peptaibol metabolites of Tolypocladium geodes. Can. J. Chem. 1996, 74, 165–172. [Google Scholar] [CrossRef]
- Mario, R.; Alessandro, M.; Cristina, P.; Fernando, F.; Stefano, M.; Evaristo, P.; José Antonio, G.; Maria Dolores, D.D.V.; Carlos, C.; Claudio, T. Lipopeptaibol metabolites of tolypocladium geodes: Total synthesis, preferred conformation, and membrane activity. Chem. Eur. J. 2003, 9, 3567–3576. [Google Scholar]
- Kimonyo, A.; Brückner, H. Sequences of metanicins, 20-residue peptaibols from the ascomycetous fungus CBS 597.80. Chem. Biodivers. 2013, 10, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Tahlan, K.; Moore, M.A.; Jensen, S.E. δ-(L-α-aminoadipyl)-L-cysteinyl-D-valine synthetase (ACVS): Discovery and perspectives. J. Ind. Microbiol. Biotechnol. 2016, 44, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Barbara, T.; Andres, B.; Thomas, B.; Urs, G.; Sonia, J.; Christina, R.; Lucius, T. An aqueous extract of the dry mycelium of Penicillium chrysogenum induces resistance in several crops under controlled and field conditions. Eur. J. Plant Pathol. 2006, 114, 185–197. [Google Scholar]
- Konstantopoulou, M.A.; Mazomenos, B.E. Evaluation of Beauveria bassiana and B. brongniartii strains and four wild-type fungal species against adults of Bactrocera oleae and Ceratitis capitata. Biocontrol 2005, 50, 293–305. [Google Scholar] [CrossRef]
- Etchegaray, A.; Dieckmann, R.; Kennedy, J.; Turner, G.; Von, D.H. ACV synthetase: Expression of amino acid activating domains of the Penicillium chrysogenum enzyme in Aspergillus nidulans. Biochem. Biophys. Res. Commun. 1997, 237, 166–169. [Google Scholar] [CrossRef]
- Hoskins, J.A.; O’Callaghan, N.; Queener, S.W.; Cantwell, C.A.; Wood, J.S.; Chen, V.J.; Skatrud, P.L. Gene disruption of the pcb AB gene encoding ACV synthetase in Cephalosporium acremonium. Curr. Genet. 1990, 18, 523–530. [Google Scholar] [CrossRef]
- Jensen, S.E.; Westlake, D.W.S.; Bowers, R.J.; Ingold, C.F.; Jouany, M.; Lyubechansky, L.; Wolfe, S. Penicillin formation by cell-free extracts of Streptomyces clavuligerus. Behaviour of aminoadipyl-modified analogs of the natural peptide precursor δ-(L-α-aminoadipyl)-L-cysteinyl-D-valine (ACV). Can. J. Chem. 2011, 62, 2712–2720. [Google Scholar] [CrossRef]
- Sundaravadivelan, C.; Padmanabhan, M.N. Effect of mycosynthesized silver nanoparticles from filtrate of Trichoderma harzianum against larvae and pupa of dengue vector Aedes aegypti L. Environ. Sci. Pollut. Res. Int. 2014, 21, 4624–4633. [Google Scholar] [CrossRef]
- Lucaciu, M.; Rebuffat, S.; Goulard, C.; Duclohier, H.; Molle, G.; Bodo, B. Interaction of the 14-residue peptaibols, harzianins HC, with lipid bilayers: Permeability modifications and conductance properties. BBA Biomembranes 1997, 1323, 85–96. [Google Scholar] [CrossRef]
- Ségalas, I.; Prigent, Y.; Davoust, D.; Bodo, B.; Rebuffat, S. Characterization of a type of β-bend ribbon spiral generated by the repeating (Xaa–Yaa–Aib–Pro) motif: The solution structure of harzianin HC IX, a 14-residue peptaibol forming voltage-dependent ion channels. Biopolymers 1999, 50, 71–85. [Google Scholar] [CrossRef]
- Leclerc, G.; Goulard, C.; Bodo, B.; Rebuffat, S. Directed biosynthesis of peptaibol antibiotics in two Trichoderma strains I. fermentation and isolation. J. Antibiot. 1998, 51, 178–183. [Google Scholar] [CrossRef][Green Version]
- Augeven-Bour, I.; Rebuffat, S.; Auvin, C.; Goulard, C.; Bodo, B. Harzianin HB I, an 11-Residue peptaibol from Trichoderma harzianum: Isolation, sequence, solution synthesis and membrane activity. J. Chem. Soc. Perkin Trans. 1997, 1, 1587–1594. [Google Scholar] [CrossRef]
- Putzu, M.; Kara, S.; Afonin, S.; Grage, S.L.; Bordessa, A.; Chaume, G.; Brigaud, T.; Ulrich, A.S.; Kubaå, T. Structural behavior of the peptaibol harzianin HK VI in a DMPC bilayer: Insights from MD simulations. Biophys. J. 2017, 112, 2602–2614. [Google Scholar] [CrossRef] [PubMed]
- Rebuffat, S.; Duclohier, H.; Auvin-Guette, C.; Molle, G.; Spach, G.; Bodo, B. Membrane-modifying properties of the pore-forming peptaibols saturnisporin SA IV and harzianin HA V. FEMS Microbiol. Lett. 1992, 105, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Duval, D. Isolation and sequence analysis of the peptide antibiotics trichorzins PA from Trichoderma harzianum. J. Chem. Soc. Perkin Trans. 1997, 1, 2147–2154. [Google Scholar] [CrossRef]
- Béven, L.; Duval, D.; Rebuffat, S.; Riddell, F.G.; Bodo, B.; Wróblewski, H. Membrane permeabilisation and antimycoplasmic activity of the 18-residue peptaibols, trichorzins PA. BBA Biomembranes 1998, 1372, 78–90. [Google Scholar] [CrossRef]
- Aric, W.; Darlene, G.; Bi-Wen, X.; Christophe, G.; Sylvie, R.; Ebbole, D.J.; Bernard, B.; Charles, K. Identification of peptaibols from Trichoderma virens and cloning of a peptaibol synthetase. J. Biol. Chem. 2002, 277, 20862–20868. [Google Scholar]
- Duval, D.; Cosette, P.; Rebuffat, S.; Duclohier, H.; Bodo, B.; Molle, G. Alamethicin-like behaviour of new 18-residue peptaibols, trichorzins PA. role of the C-terminal amino-alcohol in the ion channel forming activity. BBA Biomembranes 1998, 1369, 309–319. [Google Scholar] [CrossRef][Green Version]
- Sreenivasaprasad, S.; Manibhushanrao, K. Antagonistic potential of Gliocladium virens and Trichoderma longibrachiatum to phytopathogenic fungi. Mycopathologia 1990, 109, 19–26. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Pal, S. Entomopathogenic potential of Trichoderma longibrachiatum and its comparative evaluation with malathion against the insect pest Leucinodes orbonalis. Environ. Monit. Assess. 2016, 188, 37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Gan, Y.T.; Xu, B.L. Efficacy of Trichoderma longibrachiatum in the control of Heterodera avenae. Biocontrol 2014, 59, 319–331. [Google Scholar] [CrossRef]
- Mohamed-Benkada, M.; Pouchus, Y.F.; Vérité, P.; Pagniez, F.; Caroff, N.; Ruiz, N. Identification and biological activities of long-chain peptaibols produced by a marine-derived strain of Trichoderma longibrachiatum. Chem. Biodivers. 2016, 13, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, G. Sequences and antimycoplasmic properties of longibrachins LGB II and LGB III, two novel 20-residue peptaibols from Trichoderma longibrachiatum. J. Nat. Prod. 2001, 64, 164–170. [Google Scholar] [CrossRef]
- Cosette, P.; Rebuffat, S.; Bodo, B.; Molle, G. The ion-channel activity of longibrachins LGA I and LGB II: Effects of pro-2/Ala and gln-18/Glu substitutions on the alamethicin voltage-gated membrane channels. BBA Biomembranes 1999, 1461, 113–122. [Google Scholar] [CrossRef][Green Version]
- Nicolas, R.; Karina, P.; Marieke, V.; Isabelle, K.; Joseph, B.; Zouher, A.; Jean-François, B.; Olivier, G.; Yves François, P. Enhancement of domoic acid neurotoxicity on Diptera larvae bioassay by marine fungal metabolites. Toxicon 2010, 55, 805–810. [Google Scholar]
- Krause, C.; Kirschbaum, J.; Brückner, H. Peptaibiomics: Microheterogeneity, dynamics, and sequences of trichobrachins, peptaibiotics from Trichoderma parceramosum Bissett (T. longibrachiatum Rifai). Chem. Biodivers. 2010, 38, 1083–1102. [Google Scholar] [CrossRef]
- Mustapha, M.B.; Monique, M.; Jean-François, B.; Florence, M.; Philippe, V.; Michèle, D.; John, B.; Yves François, P. New short peptaibols from a marine Trichoderma strain. Rapid Commun. Mass Spectrom. 2006, 20, 1176–1180. [Google Scholar]
- Auvin-Guette, C.; Rebuffat, S.; Prigent, Y.; Bodo, B. Trichogin A IV, an 11-residue lipopeptaibol from Trichoderma longibrachiatum. J. Am. Chem. Soc. 1992, 114, 2170–2174. [Google Scholar] [CrossRef]
- Salnikov, E.S.; Zotti, M.D.; Bobone, S.; Mazzuca, C.; Bechinger, B. Trichogin GA IV alignment and oligomerization in phospholipid bilayers. ChemBioChem 2019, 20, 2141–2150. [Google Scholar] [CrossRef]
- Toniolo, C.; Crisma, M.; Formaggio, F.; Peggion, C.; Epand, R.F.; Epand, R.M. Lipopeptaibols, a novel family of membrane active, antimicrobial peptides. Cell. Mol. Life Sci. 2001, 58, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- De, Z.M.; Biondi, B.; Peggion, C.; Formaggio, F.; Park, Y.; Hahm, K.S.; Toniolo, C. Trichogin GA IV: A versatile template for the synthesis of novel peptaibiotics. Org. Biomol. Chem. 2012, 10, 1285–1299. [Google Scholar]
- Marta, D.Z.; Barbara, B.; Fernando, F.; Claudio, T.; Lorenzo, S.; Yoonkyung, P.; Kyung-Soo, H. Trichogin GA IV: An antibacterial and protease-resistant peptide. J. Pept. Sci. 2010, 15, 615–619. [Google Scholar]
- Regina, T.; Giulia, M.; Marta, D.Z.; Cristina, P.; Barbara, B.; Fernando, F.; Emanuele, P. The peculiar N- and C-termini of trichogin GA IV are needed for membrane interaction and human cell death induction at doses lacking antibiotic activity. BBA Biomembranes 2015, 1848, 134–144. [Google Scholar]
- Röhrich, C.R.; Iversen, A.; Jaklitsch, W.M.; Voglmayr, H.; Vilcinskas, A.; Nielsen, K.F.; Thrane, U.; Von, D.H.; Brückner, H.; Degenkolb, T. Screening the biosphere: The fungicolous fungus Trichoderma phellinicola, a prolific source of hypophellins, new 17-, 18-, 19-, and 20-residue peptaibiotics. Chem. Biodivers. 2013, 10, 787–812. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, R.; Andersson, M.A.; Kredics, L.; Grigoriev, P.A.; Sundell, N.; Salkinoja-Salonen, M.S. 20-Residue and 11-residue peptaibols from the fungus Trichoderma longibrachiatum are synergistic in forming Na+/K+ -permeable channels and adverse action towards mammalian cells. FEBS J. 2012, 279, 4172–4190. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, D.A.; Wilson, C.R. The process of antagonism of Sclerotium cepivorum in white rot affected onion roots by Trichoderma koningii. Plant Pathol. 2001, 50, 249–257. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Sherkhane, P.D.; Murthy, N.B. Induction of stable benomyl-tolerant phenotypic mutants of Trichoderma pseudokoningii MTCC 3011, and their evaluation for antagonistic and biocontrol potential. Indian J. Exp. Biol. 1999, 37, 710–712. [Google Scholar]
- Luo, Y.; Zhang, D.D.; Dong, X.W.; Zhao, P.B.; Chen, L.L.; Song, X.Y.; Wang, X.J.; Chen, X.L.; Shi, M.; Zhang, Y.Z. Antimicrobial peptaibols induce defense responses and systemic resistance in tobacco against tobacco mosaic virus. FEMS Microbiol. Lett. 2010, 313, 120–126. [Google Scholar] [CrossRef]
- Huang, Q.; Tezuka, Y.; Hatanaka, Y.; Kikuchi, T.; Nishi, A.; Tubaki, K. Studies on metabolites of mycoparasitic fungi. IV. Minor peptaibols of Trichoderma koningii. Chem. Pharm. Bull. 1995, 43, 1663–1667. [Google Scholar] [CrossRef][Green Version]
- Song, X.Y.; Shen, Q.T.; Xie, S.T.; Chen, X.L.; Sun, C.Y.; Zhang, Y.Z. Broad-spectrum antimicrobial activity and high stability of trichokonins from Trichoderma koningii SMF2 against plant pathogens. FEMS Microbiol. Lett. 2006, 260, 119–125. [Google Scholar]
- Huang, Q.; Tezuka, Y.; Kikuchi, T.; Momose, Y. Trichokonin VI, a new Ca2+ channel agonist in bullfrog cardiac myocytes. Eur. J. Pharmacol. 1994, 271, 5–6. [Google Scholar] [CrossRef]
- Ubalua, A.O.; Oti, E. Antagonistic properties of Trichoderma viride on post harvest cassava root rot pathogens. Afr. J. Biotechnol. 2007, 6, 2447–2450. [Google Scholar] [CrossRef]
- Naglot, A.; Goswami, S.; Rahman, I.; Shrimali, D.D.; Yadav, K.K.; Gupta, V.K.; Rabha, A.J.; Gogoi, H.K.; Veer, V. Antagonistic potential of native Trichoderma viride Strain against potent tea fungal pathogens in north east India. Plant Pathol. J. 2015, 31, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Psurek, A.; Neususs, C.; Pelzing, M.; Scriba, G.K.E. Analysis of the lipophilic peptaibol alamethicin by nonaqueous capillary electrophoresis-electrospray ionization-mass spectrometry. Electrophoresis 2005, 26, 4368–4378. [Google Scholar] [CrossRef]
- Cafiso, D.S. Alamethicin: A peptide model for voltage gating and protein-membrane interactions. Ann. Rev. Biophys. 1994, 23, 141–165. [Google Scholar]
- Tieleman, D.P.; Berendsen, H.J.C.; Mark, S.P.S. An alamethicin channel in a lipid bilayer: Molecular dynamics simulations. Biophys. J. 1999, 76, 1757–1769. [Google Scholar] [CrossRef]
- Matsue, T.; Shiku, H.; Yamada, H.; Uchida, I. Permselectivity of voltage-gated alamethicin ion channel studied by microamperometry. J. Phys. Chem. C 2002, 98, 394–402. [Google Scholar] [CrossRef]
- Chugh, J.K.; Brückner, H.; Wallace, B.A. Model for a helical bundle channel based on the high-resolution crystal structure of trichotoxinA 50E. Biochemistry 2002, 41, 12934–12941. [Google Scholar] [CrossRef]
- Chutrakul, C.; Alcocer, M.; Bailey, K.; Peberdy, J.F. The production and characterisation of trichotoxin peptaibols, by Trichoderma asperellum. Chem. Biodivers. 2008, 5, 1694–1706. [Google Scholar] [CrossRef]
- Schock, T.B.; Huncik, K.; Beauchesne, K.R.; Villareal, T.A.; Moeller, P.D. Identification of trichotoxin, a novel chlorinated compound associated with the bloom forming Cyanobacterium, Trichodesmium thiebautii. Environ. Sci. Technol. 2011, 45, 7503–7509. [Google Scholar] [CrossRef] [PubMed]
- Duclohier, H.; Alder, G.M.; Bashford, C.L.; Brückner, H.; Chugh, J.K.; Wallace, B.A. Conductance Studies on Trichotoxin_A50E and Implications for Channel Structure. Biophys. J. 2004, 87, 1705–1710. [Google Scholar] [CrossRef][Green Version]
- Krause, C.; Kirschbaum, J.; Jung, G.; Brückner, H. Sequence diversity of the peptaibol antibiotic suzukacillin-A from the mold Trichoderma viride. J. Pept. Sci. 2005, 12, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Röhrich, C.R.; Vilcinskas, A.; Brückner, H.; Degenkolb, T. The sequences of the eleven-residue peptaibiotics: Suzukacillins-B. Chem. Biodivers. 2013, 10, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Kiess, M.; Brückner, H. Structures of peptaibol antibiotics hypomurocin A and B from the Ascomycetous fungus Hypocrea muroiana hino et katsumoto. Eur. J. Org. Chem. 1997, 1997, 767–772. [Google Scholar] [CrossRef]
- Röhrich, C.R.; Jaklitsch, W.M.; Voglmayr, H.; Iversen, A.; Vilcinskas, A.; Nielsen, K.F.; Thrane, U.; von Döhren, H.; Brückner, H.; Degenkolb, T. Front line defenders of the ecological niche: Screening the structural diversity of peptaibiotics from saprotrophic and fungicolous Trichoderma/Hypocreaspecies. Fungal Divers. 2014, 69, 117–146. [Google Scholar] [CrossRef]
- Pradeille, N.; Zerbe, O.; Möhle, K.; Linden, A.; Heimgartner, H. The first total synthesis of the peptaibol hypomurocin A1 and its conformation analysis: An application of the ‘Azirine/Oxazolone Method’. Chem. Biodivers. 2005, 2, 1127–1152. [Google Scholar] [CrossRef]
- Pradeille, N.; Tzouros, M.; Möhle, K.; Linden, A.; Heimgartner, H. Total synthesis of the peptaibols hypomurocin A3 and hypomurocin A5, and their conformation analysis. Chem. Biodivers. 2012, 9, 2528–2558. [Google Scholar] [CrossRef]
- Zoltán, N.; János, H.; Balázs, L. In silico conformational analysis of the short-sequence hypomurocin A peptides. Int. J. Pept. 2015, 2015, 281065. [Google Scholar] [CrossRef]
- Horváth, J.; Násztor, Z.; Bartha, F.; Bogár, F.; Leitgeb, B. Characterizing the structural and folding properties of long-sequence hypomurocin B peptides and their analogs. Biopolymers 2016, 106, 645–657. [Google Scholar] [CrossRef]
- Seung-Uk, O.H.; Yun, B.S.; Sang-Jun, L.; Jung-Han, K.; Ick-Han, Y. Atroviridins A~C and neoatroviridins A~D, novel peptaibol antibiotics produced by Trichoderma atroviride F80317 I. taxonomy, fermentation, isolation and biological activities. J. Antibiot. 2002, 55, 211. [Google Scholar]
- Oh, S.U.; Lee, S.J.; Kim, J.H.; Yoo, I.-D. Structural elucidation of new antibiotic peptides, atroviridins A, B and C from Trichoderma atroviride. Tetrahedron Lett. 2010, 41, 61–64. [Google Scholar] [CrossRef]
- Carver, C.E.; Pitt, D.; Rhodes, D.J. Aetiology and biological control of Fusarium wilt of pinks (Dianthus caryophyllus) using Trichoderma aureoviride. Plant Pathol. 1996, 45, 618–630. [Google Scholar] [CrossRef]
- Kandasamy, S.K.; Myeong-Hyeon, W. Isolation and molecular identification of Trichoderma species from wetland soil and their antagonistic activity against phytopathogens. Physiol. Mol. Plant Pathol. 2020, 109, 101458. [Google Scholar] [CrossRef]
- Yun, B.S.; Yoo, I.D.; Kim, Y.H.; Kim, Y.S.; Lee, S.J.; Kim, K.S.; Yeo, W.H. Peptaivirins A and B, two new antiviral peptaibols against TMV infection. Tetrahedron Lett. 2000, 41, 1429–1431. [Google Scholar] [CrossRef]
- Walsh, C.T.; Chen, H.; Keating, T.A.; Hubbard, B.K.; Losey, H.C.; Luo, L.; Marshall, C.G.; Miller, D.A.; Patel, H.M. Tailoring enzymes that modify nonribosomal peptides during and after chain elongation on NRPS assembly lines. Curr. Opin. Chem. Biol. 2001, 5, 525–534. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, N.; Cacho, R.A.; Gong, Z.; Liu, Z.; Qin, W.; Tang, C.; Tang, Y.; Zhou, J. Structural basis of nonribosomal peptide macrocyclization in fungi. Nat. Chem. Biol. 2016, 12, 1001–1003. [Google Scholar] [CrossRef]
- Mootz, H.D.; Schwarzer, D.; Marahiel, M.A. Ways of assembling complex natural products on modular nonribosomal peptide synthetases. ChemBioChem 2002, 3, 490–504. [Google Scholar] [CrossRef]
- Axel, A.B.; Petra, S.; Qusai, A.A.; Alexander, G.; Hans, P.; André, T. Regulation of penicillin biosynthesis in filamentous fungi. Adv. Biochem. Eng. Biot. 2004, 88, 45–90. [Google Scholar]
- Gutierrez, S.; Diez, B.; Montenegro, E.; Martin, J.F. Characterization of the Cephalosporium acremonium pcbAB gene encoding alpha-aminoadipyl-cysteinyl-valine synthetase, a large multidomain peptide synthetase: Linkage to the pcbC gene as a cluster of early cephalosporin biosynthetic genes and evidence of multiple functional domains. J. Bacteriol. 1991, 173, 2354–2365. [Google Scholar]
- MacCabe, A.P.; Liempt, H.V.; Palissa, H.; Unkles, S.E.; Riach, M.B.; Pfeifer, E.; Dohren, H.V.; Kinghorn, J.R. Delta-(L-alpha-aminoadipyl)-L-cysteinyl-D-valine synthetase from Aspergillus nidulans. molecular characterization of the acvA gene encoding the first enzyme of the penicillin biosynthetic pathway. J. Biol. Chem. 1991, 266, 12646–12654. [Google Scholar] [PubMed]
- Wu, X.; García-Estrada, C.; Vaca, I.; Martín, J.F. Motifs in the C-terminal region of the Penicillium chrysogenum ACV synthetase are essential for valine epimerization and processivity of tripeptide formation. Biochimie 2012, 94, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Zompra, A.A.; Galanis, A.S.; Werbitzky, O.; Albericio, F. Manufacturing peptides as active pharmaceutical ingredients. Future Med. Chem. 2009, 1, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leucinostatin | Strain | R1 | R2 | R3 | R4 | Ref. |
|---|---|---|---|---|---|---|
| C | P. lilacinus CG-189 | -CH3 | -H | -H | -H | [13] |
| T | P. lilacinus CG-189 | -H | -H | -H | -CH3 | [13] |
| F | P. lilacinus CG-189 | -CH3 | -H | -H | -CH3 | [13] |
| D | P. lilacinus CG-189 | -CH3 | -H | -CH3 | -CH3 | [13] |
| N | P. lilacinus CG-189 | -CH3 | -OH | -H | -CH3 | [13] |
| H * | P. lilacinu CG-189 P. marquandii | -CH3 | -H | \ | \ | [13,14] |
| B2 | P. lilacinus CG-189 | -CH3 | -CH=CHCOCH2CH3 | -H | -CH3 | [13] |
| V | P. lilacinus CG-189 | -CH3 | -CH2CH2COCH2CH3 | -H | -CH3 | [13] |
| L | P. lilacinus CG-189 | -H | -CH(OH)CH2COCH2CH3 | -H | -CH3 | [13] |
| A2 | P. lilacinus CG-189 | -CH3 | -CH=CHCOCH2CH3 | -CH3 | -CH3 | [13] |
| R | P. lilacinus CG-189 | -CH3 | -CH2CH2COCH2CH3 | -CH3 | -CH3 | [13] |
| B | P. lilacinus CG-189 | -CH3 | -CH(OH)CH2COCH2CH3 | -H | -CH3 | [13] |
| S | P. lilacinus CG-189 | -CH3 | -CH2CH2(OH)CHCH2CH3 | -CH3 | -CH3 | [13] |
| A | P. lilacinus CG-189 | -CH3 | -CH(OH)CH2COCH2CH3 | -CH3 | -CH3 | [13] |
| U | P. lilacinus CG-189 | -CH3 | -CH(OCH3)CH2COCH2CH3 | -H | -CH3 | [13] |
| K | P. lilacinus CG-189, P. marquandii | -CH3 | -CH(OH)CH2COCH2CH3 | \ | \ | [13,14] |
| W | P. lilacinus CG-189 | -CH3 | -(C10H15O3) | -H | -CH3 | [13] |
| Q | P. lilacinus CG-189 | -CH3 | -(C11H17O3) | -H | -CH3 | [13] |
| O | P. lilacinus CG-189 | -CH3 | -(C11H17O3) | -CH3 | -CH3 | [13] |
| Ⅰ | P. lilacinus CG-189 | -H | -CH3 | -CH3 | -CH3 | [13] |
| Ⅱ | P. lilacinus CG-189 | -H | -H | -H | -CH3 | [13] |
| Ⅲ | P. lilacinus CG-189 | -H | -CH2CH2(OH)CHCH2CH3 | -CH3 | -CH3 | [13] |
| Ⅳ | P. lilacinus CG-189 | -CH3 | -CH2CH2(OH)CHCH2CH3 | -H | -CH3 | [13] |
| V | P. lilacinus CG-189 | -H | -CH(OH)CH2COCH2CH3 | -CH3 | -CH3 | [13] |
| Efrapeptin | Strain | R | Ref. |
|---|---|---|---|
| A | T. niveum ARSEF NO.616 | Ac-Aib-Gly-Leu-Iva- | [20] |
| B | T. niveum ARSEF NO.616 | Ac-Leu-Iva- | [20] |
| C | T. niveum ARSEF NO.616, T. geodes ARSEF 2684 | Ac-Pip-Aib-Pip-Aib-Aib-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Gly-Leu-Aib- | [19,20] |
| D | T. niveum ARSEF 616, T. geodes ARSEF 2684, M. anisopliae ME1 | Ac-Pip-Aib-Pip-Aib-Aib-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Gly-Leu-Iva- | [19,20] |
| E | T. niveum ARSEF 616, T. geodes ARSEF 2684 | Ac-Pip-Aib-Pip-Iva-Aib-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Gly-Leu-Iva- | [19,20] |
| Eα | Acremonium sp. 021172c | Ac-Pip-Aib-Pip-Iva-Aib-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Ala-Leu-Aib- | [22] |
| F | T. niveum ARSEF 616, T. geodes ARSEF 2684, M. anisopliae ME1 | Ac-Pip-Aib-Pip-Aib-Aib-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Ala-Leu-Iva- | [19,20] |
| G | T. niveum ARSEF 616, T. geodes ARSEF 2684 | Ac-Pip-Aib-Pip-Iva-Aib-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Ala-Leu-Iva- | [19,20] |
| H | T. geodes ARSEF 2684, Acremonium sp. 021172c | Ac-Pip-Aib-Pip-Iva-Iva-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Ala-Leu-Iva- | [19,22] |
| J | Tolypocladium sp. AMB18 | Ac-Pip-Aib-Pip-Aib-Aib-Leu-βAla-Gly-Aib-Aib-Pip-Aib-Ala-Leu-Aib- | [28] |
| LP237 | Composition |
|---|---|
| F8 | nOca-Aib-Pro-Phe-Aib-Gln-Gln-Aib-Et-Nva-Gln-Ala-Leuol |
| F5 | nOca-Aib-Pro-Tyr-Aib-Gln-Gln-Aib-Et-Nva-Gln-Ala-Leuol |
| F7 | nDec-Aib-Pro-Phe-Aib-Gln-Gln-Aib-Aib-Gln-Ala-Leuol |
| Metanicins | Composition |
|---|---|
| A | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln-Gln-Pheol |
| B | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-D-Iva-Gln-Gln- Pheol |
| C | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln-Gln-Pheol |
| D | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-D-Iva-Gln-Gln-Pheol |
| Harzianin | Residue | Analogs | Strain | Composition | Ref. |
|---|---|---|---|---|---|
| HC | 14 | 11 | T. harzianum M-903603 | Ac-Aib-Gln/Asn-Leu-Aib-Pro-Ala/Ser-Ile/Val-Aib-Pro-Iva/Aib-Leu-Aib-Pro-Leuol | [51] |
| HB I | 11 | 1 | T. harzianum M-903603 | Ac-Aib-Asn-Leu-Ile-Aib-Pro-Iva-Leu-Aib-Pro-Leuol | [54] |
| HK VI | 11 | 1 | Trichoderma sp. | Ac-Aib-Asn-Ile-Ile-Aib-Pro-Leu-Leu-Aib-Pro-Leuol | [55] |
| HA V | 18 | 1 | T. harzianum M-903603 | Ac-Aib-Gly-Ala-Aib-Iva-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Iva-Gln-Leuol | [56] |
| PCU4 | 14 | 1 | T. harzianum | Ac-Aib-Asn-Leu-Aib-Pro-Ser-Ile-Aib-Pro-Aib-Leu-Aib-Pro-Valinol | [53] |
| Trichorzin | Strain | Composition | Ref. |
|---|---|---|---|
| PA II | T. harzianum M-902608 | Ac-Aib-Ser-Ala-Aib-Iva-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Trpol | [57] |
| PA IV | T. harzianum M-902608 | Ac-Aib-Ser-Ala-Aib-Iva-Gln-Iva-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Trpol | [57] |
| PA V | T. harzianum M-902608 | Ac-Aib-Ser-Ala-Iva-Iva-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Trpol | [57] |
| PA VI | T. harzianum M-902608 | Ac-Aib-Ser-Ala-Aib-Iva-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Pheol | [57] |
| PA VIII | T. Harzianum M-902608) | Ac-Aib-Ser-Ala-Aib-Iva-Gln-Iva-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Pheol | [57] |
| PA IX | T. harzianum M-902608 | Ac-Aib-Ser-Ala-Iva-Iva-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Pheol | [57] |
| PAU4 | T. harzianum M-902608 | Ac-Aib-Ser-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Trpol | [57] |
| TVB I | T. virens TV29–8 | Ac-Aib-Gly-Ala-Val-Aib-Gln-Aib-Ala-Aib-Ser-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [59] |
| TVB II | T. virens TV29–8 | Ac-Aib-Gly-Ala-Leu-Aib-Gln-Aib-Ala-Aib-Ser-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [59] |
| TVB IV | T. virens TV29–8 | Ac-Aib-Gly-Ala-Leu-Aib-Gln-Iva-Ala-Aib-Ser-Leu-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [59] |
| Longibrachin | Strain | Composition | Ref. |
|---|---|---|---|
| LG A I | T. longbrachitum MMS151 | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln18-Gln-Pheol | [64] |
| LG A Ⅱ | T. longbrachitum MMS151 | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Iva-Gln18-Gln-Pheol | [64] |
| LG A Ⅲ | T. longbrachitum MMS151 | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln18-Gln-Pheol | [64] |
| LG A Ⅳ | T. longbrachitum MMS151 | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Iva-Gln18-Gln-Pheol | [64] |
| LG B Ⅱ | T. longbrachitum LCP-853431 | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Glu18-Gln-Pheol | [65] |
| LG B Ⅲ | T. longbrachitum LCP-853431 | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Iva-Glu18-Gln-Pheol | [65] |
| Trichobrachin | Strain | Composition | Ref. |
|---|---|---|---|
| A I | T. longibrachiatum | Ac-Aib-Asn-Leu-Leu-Aib-Pro-Leu-Aib-Aib-Pro-Leuol | [69] |
| A II | T. longibrachiatum | Ac-Aib-Asn-Leu-Leu-Aib-Pro-Val-Leu-Aib-Pro-Valol | [69] |
| A III | T. longibrachiatum | Ac-Aib-Asn-Val-Leu-Aib-Pro-Leu-Leu-Aib-Pro-Valol | [69] |
| A IV | T. longibrachiatum | Ac-Aib-Asn-Leu-Val-Aib-Pro-Leu-Leu-Aib-Pro-Valol | [69] |
| B I | T. longibrachiatum | Ac-Aib-Asn-Leu-Leu-Aib-Pro-Val-Aib-Val-Pro-Leuol | [69] |
| B II | T. longibrachiatum | Ac-Aib-Asn-Val-Leu-Aib-Pro-Leu-Aib-Val-Pro-Leuol | [69] |
| B III | T. longibrachiatum | Ac-Aib-Asn-Leu-Val-Aib-Pro-Leu-Aib-Val-Pro-Leuol | [69] |
| B IV | T. longibrachiatum | Ac-Aib-Asn-Leu-Leu-Aib-Pro-Leu-Aib-Val-Pro-Valol | [69] |
| I | T. parceramosum CBS 936.69 | Ac-Aib-Ala-Ala/Aib-Ala-Aib-Ala/Aib-Gln-Aib-Vxx-Aib-Gly-Leu-Aib-Pro-Vxx-Aib-Aib/Vxx-Gln-Gln | [68] |
| Ⅱ | T. parceramosum CBS 936.69 | Ac-Aib-Ala-Ala/Aib-Ala-Aib-Ala/Aib-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib/Vxx/Ala-Gln Ac-Aib-Ala-Ala/Aib-Ala-Aib-Ala/Aib-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib/Vxx/Ala-Gln-Gln-Pheol | [68] |
| III | T. parceramosum CBS 936.69 | Ac-Aib-Asn/Gln-Vxx/Lxx-Vxx/Lxx-Aib-Pro-Lxx-Vxx/Lxx-Aib-Pro-Lxxol/Valol | [68] |
| Trilongin | Strain | Composition | Ref. |
|---|---|---|---|
| AIV a | T. longibrachiatum | Ac-Aib-Asn-Vxx-Vxx-Aib-Pro-Vxx-Lxx-Aib-Pro-Lxxol | [77] |
| AIV b | T. longibrachiatum | Ac-Aib-Asn-Vxx-Vxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [77] |
| AIV c | T. longibrachiatum | Ac-Aib-Asn-Vxx-Vxx-Aib-Pro-Lxx-Vxx-Aib-Pro-Lxxol | [77] |
| AIII a | T. longibrachiatum | Ac-Aib-Asn-Lxx-Vxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [77] |
| AIII b | T. longibrachiatum | Ac-Aib-Asn-Lxx-Vxx-Aib-Pro-Vxx-Lxx-Aib-Pro-Lxxol | [77] |
| AIII c | T. longibrachiatum | Ac-Aib-Asn-Vxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [77] |
| AIII d | T. longibrachiatum | Ac-Aib-Asn-Vxx-Lxx-Aib-Pro-Vxx-Lxx-Aib-Pro-Lxxol | [77] |
| AII a | T. longibrachiatum | Ac-Aib-Asn-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [77] |
| AII b | T. longibrachiatum | Ac-Aib-Asn-Lxx-Lxx-Aib-Pro-Lxx-Vxx-Aib-Pro-Lxxol | [77] |
| AII c | T. longibrachiatum | Ac-Aib-Asn-Lxx-Lxx-Aib-Pro-Vxx-Lxx-Aib-Pro-Lxxol | [77] |
| AII d | T. longibrachiatum | Ac-Aib-Asn-Lxx-Vxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [77] |
| AII e | T. longibrachiatum | Ac-Aib-Asn-Vxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [77] |
| AI | T. longibrachiatum | Ac-Aib-Asn-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [77] |
| A0 | T. longibrachiatum | Ac-Aib-Gln-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [77] |
| BI | T. longibrachiatum | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [77] |
| BII | T. longibrachiatum | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [77] |
| BIII | T. longibrachiatum | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [77] |
| BIV | T. longibrachiatum | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [77] |
| CI | T. atroviride H1/226 | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib-Glu-Gln-Pheol | [76] |
| CII | T. atroviride H1/226 | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Glu-Gln-Pheol | [76] |
| CIII | T. atroviride H1/226 | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib-Glu-Gln-Pheol | [76] |
| CIV | T. atroviride H1/226 | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Glu-Gln-Pheol | [76] |
| Trichokonin | Strain | Composition | Ref. |
|---|---|---|---|
| VI | T. koningii OUDEMANS, T. pseudokoningii SMF2 | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln-Gln-Pheol | [81,82] |
| IX | T. koningii OUDEMANS | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Iva-Gin-Gln-Pheol | [81] |
| Ia | T. koningii OUDEMANS | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Ala- Pro-Val-Aib-Aib-Gln-Gln-Pheol | [81] |
| Ib | T. koningii OUDEMANS | Ac-Aib-Gly-Aib-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln-Gln-Pheol | [81,84] |
| Alamethicin | Composition |
|---|---|
| F30 | Ac-Aib-Pro-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Glu-Gln-Pheol |
| F50 | Ac-Aib-Pro-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln-Gln-Pheol |
| Trichotoxin | Strain | Composition | Ref. |
|---|---|---|---|
| A-40 | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Glu-Aib-Aib-Aib-Ala-Aib-Aib-Pro-Leu-Aib-D-Iva-Gln-Valol | [90] |
| A-40 I | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Ala-Ala-Aib-Aib-Pro-Leu-Aib-Aib-Glu-Valol | [90] |
| A-40 II | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Ala-Aib-Pro-Leu-Aib-Aib-Glu-Valol | [90] |
| A-40 III | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Ala-Ala-Aib-Aib-Pro-Leu-Aib-D-Iva-Glu-Valol | [90] |
| A-40 IV | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Aib-Aib-Pro-Leu-Aib-Aib-Glu-Valol | [90] |
| A-40 V | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Aib-Aib-Pro-Leu-Aib-D-Iva-Glu-Valol | [90] |
| A-40 Va | T. viride NRRL 5242 | Ac-Aib-Ala-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Aib-Aib-Pro-Leu-Aib-Aib-Glu-Valol | [90] |
| A-50 E | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Ala-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [90] |
| A-50 F | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Ala-Ala-Ala-Aib-Pro-Leu-Aib-D-Iva-Gln-Valol | [90] |
| A-50 G | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Ala-Aib-Pro-Leu-Aib-D-Iva-Gln-Valol | [90] |
| A-50 H | T. viride NRRL 5242 | Ac-Aib-Ala-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Ala-Aib-Pro-Leu-Aib-D-Iva-Gln-Valol | [90] |
| A-50 I | T. viride NRRL 5242 | Ac-Aib-Gly-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Aib-Aib-Pro-Leu-Aib-D-Iva-Gln-Valol | [90] |
| A-50 J | T. viride NRRL 5242 | Ac-Aib-Ala-Aib-Leu-Aib-Gln-Aib-Aib-Aib-Ala-Aib-Aib-Pro-Leu-Aib-D-Iva-Gln-Valol | [90] |
| Suzukacillin | Strain | Composition | Ref. |
|---|---|---|---|
| SZ-A1 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Vxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [94] |
| SZ-A2 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [94] |
| SZ-A3 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Vxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A4 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A5 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Lxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [94] |
| SZ-A6 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [94] |
| SZ-A7 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Lxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A8 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Aib-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A9 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [94] |
| SZ-A10a | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Vxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A10b | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A11a | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Aib-Gln-Gln-Pheol | [94] |
| SZ-A11b | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A12 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Aib-Gln-Aib-Lxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-A13 | T. viride 63C-I | Ac-Aib-Ala-Aib-Ala-Aib-Ala-Gln-Aib-Lxx-Aib-Gly-Lxx-Aib-Pro-Vxx-Aib-Vxx-Gln-Gln-Pheol | [94] |
| SZ-B1 | T. viride 63C-I | Ac-Aib-Gln-Vxx-Lxx-Aib-Pro-Lxx-Lxx-Ala-Pro | [95] |
| SZ-B2 | T. viride 63C-I | Ac-Aib-Gln-Lxx-Vxx-Ala-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [95] |
| SZ-B3 | T. viride 63C-I | Ac-Aib-Asn-Vxx-Vxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B4 | T. viride 63C-I | Ac-Ala-Gln-Vxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B5 | T. viride 63C-I | Ac-Ala-Gln-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [95] |
| SZ-B6 | T. viride 63C-I | Ac-Ala-Gln-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol - | [95] |
| SZ-B7 | T. viride 63C-I | Ac-Ala-Gln-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [95] |
| SZ-B8 | T. viride 63C-I | Ac-Aib-Asn-Vxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B9 | T. viride 63C-I | Ac-Aib-Gln-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Vxxol | [95] |
| SZ-B10 | T. viride 63C-I | Ac-Aib-Gln-Lxx-Vxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B11 | T. viride 63C-I | Ac-Aib-Asn-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B12 | T. viride 63C-I | Ac-Aib-Gln-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B13 | T. viride 63C-I | Ac-Vxx-Gln-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B14 | T. viride 63C-I | Ac-Aib-Ala-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| SZ-B15 | T. viride 63C-I | Ac-Vxx-Glu-Lxx-Lxx-Aib-Pro-Lxx-Lxx-Aib-Pro-Lxxol | [95] |
| Hypomurocin | Strain | Composition | Ref. |
|---|---|---|---|
| A-1 | H. muroiana IFO31288 | Ac-Aib-Gln-Val-Val-Aib-Pro-Leu-Leu-Aib-Pro-Leuol | [96] |
| A-2 | H. muroiana IFO31288 | Ac-D-Iva-Gln-Val-Val-Aib-Pro-Leu-Leu-Aib-Pro-Leuol | [96] |
| A-3 | H. muroiana IFO31288 | Ac-Aib-Gln-Val-Leu-Aib-Pro-Leu-Ile-Aib-Pro-Leuol | [96] |
| A-4 | H. muroiana IFO31288 | Ac-Aib-Gln-Ile-Val-Aib-Pro-Leu-Leu-Aib-Pro-Leuol | [96] |
| A-5 | H. muroiana IFO31288 | Ac-Aib-Gln-Ile-Ile-Aib-Pro-Leu-Leu-Aib-Pro-Leuol | [96] |
| A-5a | H. muroiana IFO31288 | Ac-Aib-Gln-Ile-Leu-Aib-Pro-Leu-Ile-Aib-Pro-Leuol | [96] |
| B-1 | H. muroiana IFO31288 | Ac-Aib-Ser-Ala-Leu-Aib-Gln-Aib-Val-Aib-Gly-Aib-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [96] |
| B-2 | H. muroiana IFO31288 | Ac-Aib-Ser-Ala-Leu-Aib-Gln-Aib-Val-Aib-Gly-Aib-Aib-Pro-Leu-Aib-Aib-Gln-Leuol | [96] |
| B-3a | H. muroiana IFO31288 | Ac-Aib-Ala-Ala-Leu-Aib-Gln-Aib-Val-Aib-Gly-Aib-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [96] |
| B-3b | H. muroiana IFO31288 | Ac-Aib-Ser-Ala-Leu-Aib-Gln-Iva-Val-Aib-Gly-Aib-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [96] |
| B-4 | H. muroiana IFO31288 | Ac-Aib-Ser-Ala-Leu-Aib-Gln-Aib-Val-Aib-Gly-Iva-Aib-Pro-Leu-Aib-Aib-Gln-Valol | [96] |
| B-5 | H. muroiana IFO31288 | Ac-Aib-Ser-Ala-Leu-Aib-Gln-Aib-Val-Aib-Gly-Iva-Aib-Pro-Leu-Aib-Aib-Gln-Leuol | [96] |
| B | T. harzianum | Ac-Aib-Ser-Ala-Leu-Ala-Gln-Aib-Val-Aib-Gly-Aib-Aib-Pro-Leu-Aib- Aib-Gln-Valol | [59] |
| Atroviridin | Composition |
|---|---|
| A | Ac-Aib-Pro-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Aib-Gln-Gln-Pheol |
| B | Ac-Aib-Pro-Aib-Ala-Aib-Ala-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Iva-Gln-Gln-Pheol |
| C | Ac-Aib-Pro-Aib-Ala-Aib-Aib-Gln-Aib-Val-Aib-Gly-Leu-Aib-Pro-Val-Aib-Iva-Gln-Gln-Pheol |
| Neoatroviridin | Composition |
|---|---|
| A | Ac-Aib-Gly-Ala-Leu-Aib-Gln-Aib-Leu-Aib-Gly-Iva-Aib-Pro-Leu-Aib-Aib-Gln-Leuol |
| B | Ac-Aib-Gly-Ala-Leu-Iva-Gln-Aib-Leu-Aib-Gly-Iva-Aib-Pro-Leu-Aib-Aib-Gln-Leuol |
| C | Ac-Aib-Gly-Ala-Leu-Aib-Gln-Iva-Leu-Aib-Gly-Iva-Aib-Pro-Leu-Aib-Aib-Gln-Leuol |
| D | Ac-Aib-Gly-Ala-Leu-Iva-Gln-Iva-Leu-Aib-Gly-Iva-Aib-Pro-Leu-Aib-Aib-Gln-Leuol |
| Peptaivirin | Composition |
|---|---|
| A | Ac-Phe-Aib-Ala-Aib-Iva-Leu-Gln-Gly-Aib-Aib-Ala-Ala-Aib-Pro-Iva-Aib-Aib-Gln-Trpol |
| B | Ac-Phe-Aib-Ser-Aib-Iva-Leu-Gln-Gly-Aib-Aib-Ala-Ala-Aib-Pro-Iva-Aib-Aib-Gln-Pheol |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, X.; Thaochan, N.; Hu, Q. Diversity of Linear Non-Ribosomal Peptide in Biocontrol Fungi. J. Fungi 2020, 6, 61. https://doi.org/10.3390/jof6020061
Niu X, Thaochan N, Hu Q. Diversity of Linear Non-Ribosomal Peptide in Biocontrol Fungi. Journal of Fungi. 2020; 6(2):61. https://doi.org/10.3390/jof6020061
Chicago/Turabian StyleNiu, Xiaoyan, Narit Thaochan, and Qiongbo Hu. 2020. "Diversity of Linear Non-Ribosomal Peptide in Biocontrol Fungi" Journal of Fungi 6, no. 2: 61. https://doi.org/10.3390/jof6020061
APA StyleNiu, X., Thaochan, N., & Hu, Q. (2020). Diversity of Linear Non-Ribosomal Peptide in Biocontrol Fungi. Journal of Fungi, 6(2), 61. https://doi.org/10.3390/jof6020061