Adaptive Immunity to Cryptococcus neoformans Infections

Abstract
1. Introduction
1.1. Dendritic Cells and Macrophages Connect the Innate and Adaptive Immune Systems during C. Neoformans Infection
1.2. Dendritic Cells Are the Primary Antigen Presenting Cells during Cryptococcal Infection
2. Cell-Mediated Immunity: T Cells
2.1. Importance of T Cells during Cryptococcal Infections
2.2. Cryptococcal Antigens Activate T Cell Maturation and Proliferation
2.3. Regulatory T Cells (Tregs)
2.4. Natural Killer T(NKT) Cells
2.5. Gamma Delta (γδ) T Cells
2.6. Memory T Cells
3. Antibody-Mediated Immunity against Cryptococcal Infections
3.1. B Cells and Antibody-Mediated Immune Responses in Human Cryptococcal Infections
3.2. B Cells and Antibody-Mediated Immune Responses in Non-Human Cryptococcal Infections
4. Cytokine Responses during C. neoformans Infections
4.1. Protective Cytokines
4.2. Non-Protective Cytokines
Cytokines Associated with IRIS
4.3. Cytokines/Chemokines with Varying/Conflicting Roles
4.3.1. Cytokines/Chemokines with Beneficial Role in Mice, but Detrimental in Humans
4.3.2. Cytokines with Contradictory Roles in Both Mouse and Human C. neoformans Infections
4.3.3. Cytokines with Contradictory Roles in Only One System
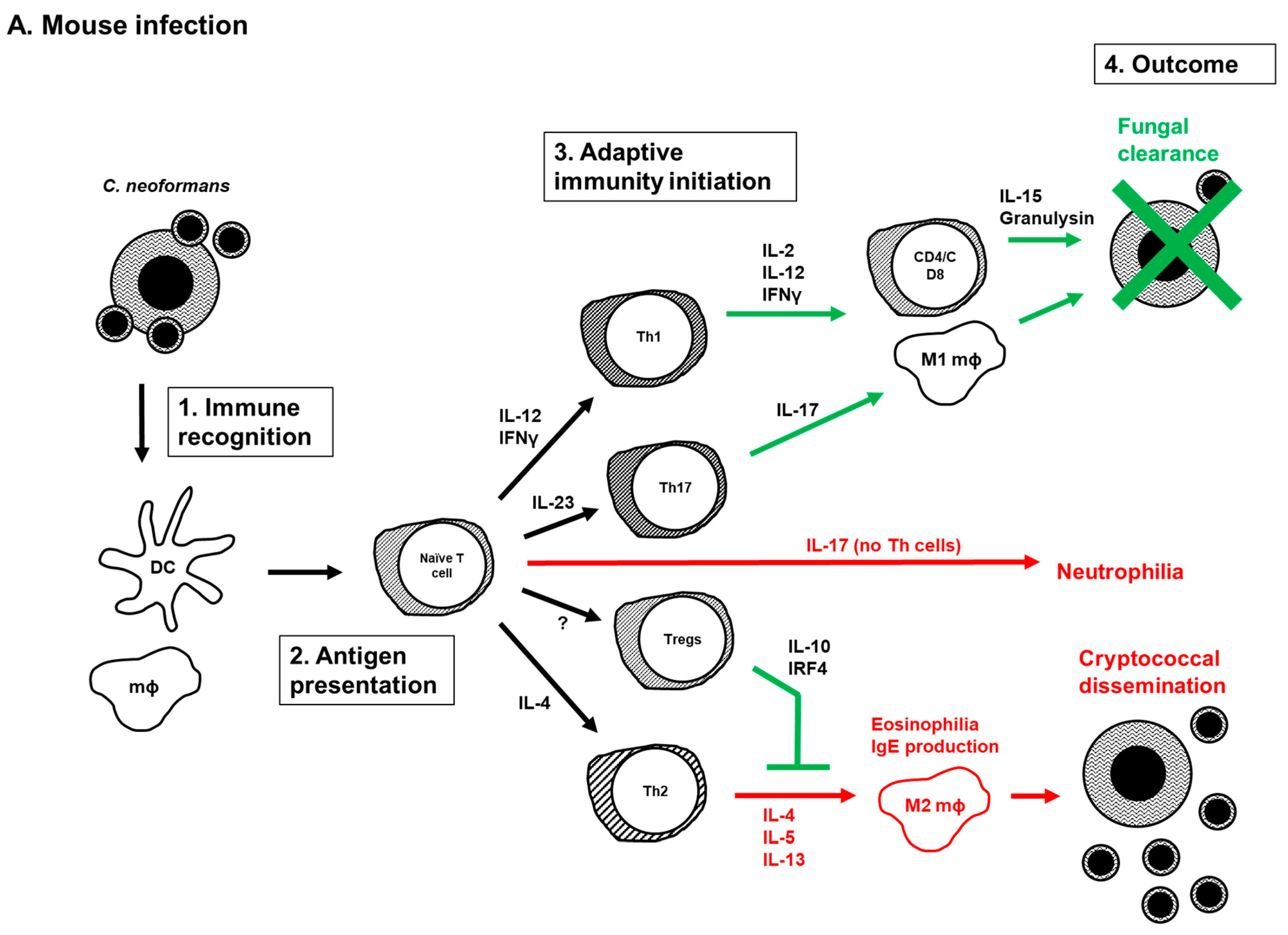
5. Current Model of the Adaptive Immune Response to Cryptococcus Infection
6. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hagen, F.; Khayhan, K.; Theelen, B.; Kolecka, A.; Polacheck, I.; Sionov, E.; Falk, R.; Parnmen, S.; Lumbsch, H.T.; Boekhout, T. Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species complex. Fungal Genet. Biol. 2015, 78, 16–48. [Google Scholar] [CrossRef] [PubMed]
- Park, B.J.; Wannemuehler, K.A.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 2009, 23, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Perfect, J.R.; Cloud, G.A.; Larsen, R.A.; Pankey, G.A.; Lancaster, D.J.; Henderson, H.; Kauffman, C.A.; Haas, D.W.; Saccente, M.; et al. Cryptococcosis in human immunodeficiency virus-negative patients in the era of effective azole therapy. Clin. Infect. Dis. 2001, 33, 690–699. [Google Scholar] [CrossRef] [PubMed]
- Pyrgos, V.; Seitz, A.E.; Steiner, C.A.; Prevots, D.R.; Williamson, P.R. Epidemiology of cryptococcal meningitis in the US: 1997–2009. PLoS ONE 2013, 8, e56269. [Google Scholar] [CrossRef] [PubMed]
- Marr, K.A.; Datta, K.; Pirofski, L.A.; Barnes, R. Cryptococcus gattii infection in healthy hosts: A sentinel for subclinical immunodeficiency? Clin. Infect. Dis. 2012, 54, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Hoang, L.; Kibsey, P.; Morshed, M.; Phillips, P. Clinical presentation, diagnosis and management of Cryptococcus gattii cases: Lessons learned from British Columbia. Can. J. Infect. Dis. Med. Microbiol. 2009, 20, 23–28. [Google Scholar] [PubMed]
- Hoang, L.M.; Maguire, J.A.; Doyle, P.; Fyfe, M.; Roscoe, D.L. Cryptococcus neoformans infections at Vancouver Hospital and Health Sciences Centre (1997–2002): Epidemiology, microbiology and histopathology. J. Med. Microbiol. 2004, 53, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Kidd, S.E.; Hagen, F.; Tscharke, R.L.; Huynh, M.; Bartlett, K.H.; Fyfe, M.; MacDougall, L.; Boekhout, T.; Kwon-Chung, K.J.; Meyer, W. A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proc. Natl. Acad. Sci. USA 2004, 101, 17258–17263. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.; Galanis, E.; MacDougall, L.; Chong, M.Y.; Balshaw, R.; Cook, V.J.; Bowie, W.; Steiner, T.; Hoang, L.; Morshed, M.; et al. Longitudinal clinical findings and outcome among patients with Cryptococcus gattii infection in British Columbia. Clin. Infect. Dis. 2015, 60, 1368–1376. [Google Scholar] [PubMed]
- Bennett, J.E.; Kwon-Chung, K.J.; Howard, D.H. Epidemiologic differences among serotypes of Cryptococcus neoformans. Am. J. Epidemiol. 1977, 105, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.M.; Lazera, M.S.; Barbosa, G.G.; Trilles, L.; Balassiano, B.R.; Macedo, R.C.; Bezerra, C.C.; Perez, M.A.; Cardarelli, P.; Wanke, B. Serotyping of 467 Cryptococcus neoformans isolates from clinical and environmental sources in Brazil: Analysis of host and regional patterns. J. Clin. Microbiol. 2003, 41, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D. The role of chemokines in linking innate and adaptive immunity. Curr. Opin. Immunol. 2002, 14, 129–135. [Google Scholar] [CrossRef]
- Svanborg, C.; Godaly, G.; Hedlund, M. Cytokine responses during mucosal infections: Role in disease pathogenesis and host defence. Curr. Opin. Microbiol. 1999, 2, 99–105. [Google Scholar] [CrossRef]
- Wozniak, K.L.; Vyas, J.M.; Levitz, S.M. In vivo role of dendritic cells in a murine model of pulmonary cryptococcosis. Infect. Immun. 2006, 74, 3817–3824. [Google Scholar] [CrossRef] [PubMed]
- Vecchiarelli, A.; Dottorini, M.; Pietrella, D.; Monari, C.; Retini, C.; Todisco, T.; Bistoni, F. Role of human alveolar macrophages as antigen-presenting cells in Cryptococcus neoformans infection. Am. J. Respir. Cell Mol. Biol. 1994, 11, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Osterholzer, J.J.; Milam, J.E.; Chen, G.H.; Toews, G.B.; Huffnagle, G.B.; Olszewski, M.A. Role of dendritic cells and alveolar macrophages in regulating early host defense against pulmonary infection with Cryptococcus Neoformans. Infect. Immun. 2009, 77, 3749–3758. [Google Scholar] [CrossRef] [PubMed]
- Guillot, L.; Carroll, S.F.; Badawy, M.; Qureshi, S.T. Cryptococcus neoformans induces IL-8 secretion and CXCL1 expression by human bronchial epithelial cells. Respir. Res. 2008, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Roseff, S.A.; Levitz, S.M. Effect of endothelial cells on phagocyte-mediated anticryptococcal activity. Infect. Immun. 1993, 61, 3818–3824. [Google Scholar] [PubMed]
- Schon-Hegrad, M.A.; Oliver, J.; McMenamin, P.G.; Holt, P.G. Studies on the density, distribution, and surface phenotype of intraepithelial class II major histocompatibility complex antigen (Ia)-bearing dendritic cells (DC) in the conducting airways. J. Exp. Med. 1991, 173, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, E.S.; Mellman, I. Cell biology of antigen processing in vitro and in vivo. Annu. Rev. Immunol. 2005, 23, 975–1028. [Google Scholar] [CrossRef] [PubMed]
- Upham, J.W. The role of dendritic cells in immune regulation and allergic airway inflammation. Respirology 2003, 8, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Unutmaz, D.; Wong, P.; Sano, G.; de los Santos, K.; Sparwasser, T.; Wu, S.; Vuthoori, S.; Ko, K.; Zavala, F.; et al. In vivo depletion of CD11c+ dendritic cells abrogates priming of CD8+ T cells by exogenous cell-associated antigens. Immunity 2002, 17, 211–220. [Google Scholar] [CrossRef]
- Berger, A. Th1 and Th2 responses: What are they? Br. Med. J. 2000, 321, 424. [Google Scholar] [CrossRef]
- Guglani, L.; Khader, S.A. Th17 cytokines in mucosal immunity and inflammation. Curr. Opin. HIV AIDS 2010, 5, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 2005, 201, 233–240. [Google Scholar] [CrossRef] [PubMed]
- De Becker, G.; Moulin, V.; Tielemans, F.; de Mattia, F.; Urbain, J.; Leo, O.; Moser, M. Regulation of T helper cell differentiation in vivo by soluble and membrane proteins provided by antigen-presenting cells. Eur. J. Immunol. 1998, 28, 3161–3171. [Google Scholar] [CrossRef]
- Vieira, P.L.; de Jong, E.C.; Wierenga, E.A.; Kapsenberg, M.L.; Kalinski, P. Development of Th1-inducing capacity in myeloid dendritic cells requires environmental instruction. J. Immunol. 2000, 164, 4507–4512. [Google Scholar] [CrossRef] [PubMed]
- Ranger, A.M.; Das, M.P.; Kuchroo, V.K.; Glimcher, L.H. B7-2 (CD86) is essential for the development of IL-4-producing T cells. Int. Immunol. 1996, 8, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, Y.; Yang, L.P.; Uchiyama, T.; Tanaka, Y.; Baum, P.; Sergerie, M.; Hermann, P.; Delespesse, G. OX40 costimulation enhances interleukin-4 (IL-4) expression at priming and promotes the differentiation of naive human CD4(+) T cells into high IL-4-producing effectors. Blood 1998, 92, 3338–3345. [Google Scholar] [PubMed]
- Lambrecht, B.N.; de Veerman, M.; Coyle, A.J.; Gutierrez-Ramos, J.-C.; Thielemans, K.; Pauwels, R.A. Myeloid dendritic cells induce Th2 responses to inhaled antigen, leading to eosinophilic airway inflammation. J. Clin. Investig. 2000, 106, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Haczku, A.; Takeda, K.; Redai, I.; Hamelmann, E.; Cieslewicz, G.; Joetham, A.; Loader, J.; Lee, J.J.; Irvin, C.; Gelfand, E.W. Anti-CD86 (B7.2) treatment abolishes allergic airway hyperresponsiveness in mice. Am. J. Respir. Crit. Care Med. 1999, 159, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 2005, 6, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; de Luca, A.; D’Angelo, C.; Moretti, S.; Romani, L. IL-17/Th17 in anti-fungal immunity: What’s new? Eur. J. Immunol. 2009, 39, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Na, L.; Fidel, P.L.; Schwarzenberger, P. Requirement of interleukin-17A for systemic anti-Candida albicans host defense in mice. J. Infect. Dis. 2004, 190, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Van de Veerdonk, F.L.; Kullberg, B.J.; Verschueren, I.C.; Hendriks, T.; van der Meer, J.W.; Joosten, L.A.; Netea, M.G. Differential effects of IL-17 pathway in disseminated candidiasis and zymosan-induced multiple organ failure. Shock 2010, 34, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Santos, N.; Gaffen, S.L. Th17 cells in immunity to Candida albicans. Cell Host Microbe 2012, 11, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Delsing, C.E.; Becker, K.L.; Simon, A.; Kullberg, B.J.; Bleeker-Rovers, C.P.; van de Veerdonk, F.L.; Netea, M.G. Th17 cytokine deficiency in patients with Aspergillus skull base osteomyelitis. BMC Infect. Dis. 2015, 15, 140. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, D.L.; Smith, K.D.; Kashem, S.W.; Bohjanen, P.R.; Nielsen, K. Different Lymphocyte Populations Direct Dichotomous Eosinophil or Neutrophil Responses to Pulmonary Cryptococcus Infection. J. Immunol. 2017, 198, 1627–1637. [Google Scholar] [CrossRef] [PubMed]
- Hoag, K.A.; Lipscomb, M.F.; Izzo, A.A.; Street, N.E. IL-12 and IFN-γ are required for initiating the protective Th1 response to pulmonary cryptococcosis in resistant C.B-17 mice. Am. J. Respir. Cell Mol. Biol. 1997, 17, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Herring, A.C.; Lee, J.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Induction of interleukin-12 and γ interferon requires tumor necrosis factor α for protective T1-cell-mediated immunity to pulmonary Cryptococcus neoformans infection. Infect. Immun. 2002, 70, 2959–2964. [Google Scholar] [CrossRef] [PubMed]
- Hoag, K.A.; Street, N.E.; Huffnagle, G.B.; Lipscomb, M.F. Early cytokine production in pulmonary Cryptococcus neoformans infections distinguishes susceptible and resistant mice. Am. J. Respir. Cell Mol. Biol. 1995, 13, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Meintjes, G.; Rebe, K.; Williams, G.N.; Bicanic, T.; Williams, A.; Schutz, C.; Bekker, L.G.; Wood, R.; Harrison, T.S. Adjunctive interferon-γ immunotherapy for the treatment of HIV-associated cryptococcal meningitis: A randomized controlled trial. Aids 2012, 26, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Decken, K.; Kohler, G.; Palmer-Lehmann, K.; Wunderlin, A.; Mattner, F.; Magram, J.; Gately, M.K.; Alber, G. Interleukin-12 is essential for a protective Th1 response in mice infected with Cryptococcus neoformans. Infect. Immun. 1998, 66, 4994–5000. [Google Scholar] [PubMed]
- Voelz, K.; Lammas, D.A.; May, R.C. Cytokine signaling regulates the outcome of intracellular macrophage parasitism by Cryptococcus neoformans. Infect. Immun. 2009, 77, 3450–3457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, F.; Tompkins, K.C.; McNamara, A.; Jain, A.V.; Moore, B.B.; Toews, G.B.; Huffnagle, G.B.; Olszewski, M.A. Robust Th1 and Th17 immunity supports pulmonary clearance but cannot prevent systemic dissemination of highly virulent Cryptococcus neoformans H99. Am. J. Pathol. 2009, 175, 2489–2500. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, D.L.; Specht, C.A.; Lee, C.K.; Smith, K.D.; Mukaremera, L.; Lee, S.T.; Lee, C.G.; Elias, J.A.; Nielsen, J.N.; Boulware, D.R.; et al. Chitin Recognition via Chitotriosidase Promotes Pathologic Type-2 Helper T Cell Responses to Cryptococcal Infection. PLoS Pathog. 2015, 11, e1004701. [Google Scholar] [CrossRef] [PubMed]
- Murdock, B.J.; Huffnagle, G.B.; Olszewski, M.A.; Osterholzer, J.J. Interleukin-17A Enhances Host Defense against Cryptococcal Lung Infection through Effects Mediated by Leukocyte Recruitment, Activation, and γ Interferon Production. Infect. Immun. 2014, 82, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Bauman, S.K.; Nichols, K.L.; Murphy, J.W. Dendritic cells in the induction of protective and nonprotective anticryptococcal cell-mediated immune responses. J. Immunol. 2000, 165, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Choi, H.K.; Son, J.; Kim, K.H.; Lee, S.H. Cryptococcal Meningitis in Patients with or without Human Immunodeficiency Virus: Experience in a Tertiary Hospital. Yonsei Med. J. 2011, 52, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Kofteridis, D.P.; Saridaki, Z.; Kazakou, I.; Lazaridou, S.; Alegakis, D.; Milaki, G.; Gikas, A. Idiopathic CD4+ T lymphocytopenia disclosed by recurrent cryptococcal meningitis. First case report from Greece. Int. J. Infect. Dis. 2005, 9, 347–348. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Lal, V.; Modi, M.; Khurana, D.; Bal, S.; Prabhakar, S. Idiopathic CD4 lymphocytopenia presenting as refractory cryptococcal meningitis. Ann. Indian Acad. Neurol. 2010, 13, 136–138. [Google Scholar] [PubMed]
- Jo, E.K.; Kim, H.S.; Lee, M.Y.; Iseki, M.; Lee, J.H.; Song, C.H.; Park, J.K.; Hwang, T.J.; Kook, H. X-linked hyper-IgM syndrome associated with Cryptosporidium parvum and Cryptococcus neoformans infections: The first case with molecular diagnosis in Korea. J. Korean Med. Sci. 2002, 17, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Winkelstein, J.A.; Marino, M.C.; Ochs, H.; Fuleihan, R.; Scholl, P.R.; Geha, R.; Stiehm, E.R.; Conley, M.E. The X-linked hyper-IgM syndrome: Clinical and immunologic features of 79 patients. Medicine 2003, 82, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Atkinson, T.P.; Lipsky, P.E.; Slater, J.E.; Nelson, D.L.; Strober, W. Defects of T-cell effector function and post-thymic maturation in X-linked hyper-IgM syndrome. J. Clin. Investig. 1999, 103, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.L.; Spurrell, J.C.; Wang, J.F.; Neely, G.G.; Epelman, S.; Krensky, A.M.; Mody, C.H. CD8 T cell-mediated killing of Cryptococcus neoformans requires granulysin and is dependent on CD4 T cells and IL-15. J. Immunol. 2002, 169, 5787–5795. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.F.; Ma, L.L.; Jones, G.J.; Gill, M.J.; Krensky, A.M.; Kubes, P.; Mody, C.H. Cytotoxic CD4+ T cells use granulysin to kill Cryptococcus neoformans, and activation of this pathway is defective in HIV patients. Blood 2007, 109, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.M.; Moore, T.A.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Generation of antifungal effector CD8+ T cells in the absence of CD4+ T cells during Cryptococcus neoformans infection. J. Immunol. 2005, 174, 7920–7928. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.M.; Ballinger, M.N.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Diversity of the T-Cell Response to Pulmonary Cryptococcus neoformans Infection. Infect. Immun. 2006, 74, 4538–4548. [Google Scholar] [CrossRef] [PubMed]
- Syme, R.M.; Wood, C.J.; Wong, H.; Mody, C.H. Both CD4+ and CD8+ human lymphocytes are activated and proliferate in response to Cryptococcus neoformans. Immunology 1997, 92, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Huffnagle, G.B.; Lipscomb, M.F.; Lovchik, J.A.; Hoag, K.A.; Street, N.E. The role of CD4+ and CD8+ T cells in the protective inflammatory response to a pulmonary cryptococcal infection. J. Leukoc. Biol. 1994, 55, 35–42. [Google Scholar] [PubMed]
- Lindell, D.M.; Moore, T.A.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Distinct Compartmentalization of CD4(+) T-Cell Effector Function Versus Proliferative Capacity during Pulmonary Cryptococcosis. Am. J. Pathol. 2006, 168, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Schulze, B.; Piehler, D.; Eschke, M.; von Buttlar, H.; Kohler, G.; Sparwasser, T.; Alber, G. CD4(+) FoxP3(+) regulatory T cells suppress fatal T helper 2 cell immunity during pulmonary fungal infection. Eur. J. Immunol. 2014, 44, 3596–3604. [Google Scholar] [CrossRef] [PubMed]
- Schulze, B.; Piehler, D.; Eschke, M.; Heyen, L.; Protschka, M.; Kohler, G.; Alber, G. Therapeutic expansion of CD4+FoxP3+ regulatory T cells limits allergic airway inflammation during pulmonary fungal infection. Pathog. Dis. 2016, 74, ftw020. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, D.L.; Smith, K.D.; Kotov, D.I.; Nielsen, J.N.; Bohjanen, P.R.; Nielsen, K. Regulatory T Cell Induction and Retention in the Lungs Drives Suppression of Detrimental Type 2 Th Cells During Pulmonary Cryptococcal Infection. J. Immunol. 2016, 196, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Kinjo, Y.; Uezu, K.; Yara, S.; Miyagi, K.; Koguchi, Y.; Nakayama, T.; Taniguchi, M.; Saito, A. Monocyte Chemoattractant Protein-1-Dependent Increase of Vα14 NKT Cells in Lungs and Their Roles in Th1 Response and Host Defense in Cryptococcal Infection. J. Immunol. 2001, 167, 6525–6532. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Kinjo, Y.; Yara, S.; Koguchi, Y.; Uezu, K.; Nakayama, T.; Taniguchi, M.; Saito, A. Activation of Vα14(+) Natural Killer T Cells by α-Galactosylceramide Results in Development of Th1 Response and Local Host Resistance in Mice Infected with Cryptococcus neoformans. Infect. Immun. 2001, 69, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, T.; Nakamura, K.; Miyazato, A.; Nakagawa, K.; Miyazawa, T.; Kinjyo, Y.; Kronenberg, M.; Kaku, M.; Kawakami, K. CD1d-dependent activation of NKT cells by Cryptococcus neoformans-pulsed dendritic cells (B172). J. Immunol. 2007, 178 (Suppl. 1), LB36. [Google Scholar]
- Blackstock, R.; Murphy, J.W. Age-related resistance of C57BL/6 mice to Cryptococcus neoformans is dependent on maturation of NKT cells. Infect. Immun. 2004, 72, 5175–5180. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Nakamura, Y.; Sato, K.; Takahashi, Y.; Nomura, T.; Miyasaka, T.; Ishii, K.; Hara, H.; Yamamoto, N.; Kanno, E.; et al. Defect of CARD9 leads to impaired accumulation of γ interferon-producing memory phenotype T cells in lungs and increased susceptibility to pulmonary infection with Cryptococcus neoformans. Infect. Immun. 2014, 82, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, K.L.; Kolls, J.K.; Wormley, F.L. Depletion of neutrophils in a protective model of pulmonary cryptococcosis results in increased IL-17A production by γ/δ T cells. BMC Immunol. 2012, 13, 65. [Google Scholar] [CrossRef] [PubMed]
- Uezu, K.; Kawakami, K.; Miyagi, K.; Kinjo, Y.; Kinjo, T.; Ishikawa, H.; Saito, A. Accumulation of γδ T Cells in the Lungs and Their Regulatory Roles in Th1 Response and Host Defense against Pulmonary Infection with Cryptococcus neoformans. J. Immunol. 2004, 172, 7629–7634. [Google Scholar] [CrossRef] [PubMed]
- Mody, C.H.; Sims, K.L.; Wood, C.J.; Syme, R.M.; Spurrell, J.C.; Sexton, M.M. Proteins in the cell wall and membrane of Cryptococcus neoformans stimulate lymphocytes from both adults and fetal cord blood to proliferate. Infect. Immun. 1996, 64, 4811–4819. [Google Scholar] [PubMed]
- Mody, C.H.; Wood, C.J.; Syme, R.M.; Spurrell, J.C. The cell wall and membrane of Cryptococcus neoformans possess a mitogen for human T lymphocytes. Infect. Immun. 1999, 67, 936–941. [Google Scholar] [PubMed]
- Syme, R.M.; Spurrell, J.C.; Ma, L.L.; Green, F.H.; Mody, C.H. Phagocytosis and protein processing are required for presentation of Cryptococcus neoformans mitogen to T lymphocytes. Infect. Immun. 2000, 68, 6147–6153. [Google Scholar] [CrossRef] [PubMed]
- Huffnagle, G.B.; Yates, J.L.; Lipscomb, M.F. Immunity to a pulmonary Cryptococcus neoformans infection requires both CD4+ and CD8+ T cells. J. Exp. Med. 1991, 173, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Levitz, S.M.; Dupont, M.P. Phenotypic and functional characterization of human lymphocytes activated by interleukin-2 to directly inhibit growth of Cryptococcus neoformans in vitro. J. Clin. Investig. 1993, 91, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.W.; Hidore, M.R.; Wong, S.C. Direct interactions of human lymphocytes with the yeast-like organism, Cryptococcus neoformans. J. Clin. Investig. 1993, 91, 1553–1566. [Google Scholar] [CrossRef] [PubMed]
- Workman, C.J.; Szymczak-Workman, A.L.; Collison, L.W.; Pillai, M.R.; Vignali, D.A.A. The Development and Function of Regulatory T Cells. Cell. Mol. Life Sci. CMLS 2009, 66, 2603–2622. [Google Scholar] [CrossRef] [PubMed]
- Corthay, A. How do Regulatory T Cells Work? Scand. J. Immunol. 2009, 70, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Kroetz, D.N.; Deepe, G.S., Jr. CCR5 dictates the equilibrium of proinflammatory IL-17+ and regulatory Foxp3+ T cells in fungal infection. J. Immunol. 2010, 184, 5224–5231. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Sutmuller, R.; Hermann, C.; van der Graaf, C.A.; van der Meer, J.W.; van Krieken, J.H.; Hartung, T.; Adema, G.; Kullberg, B.J. Toll-like receptor 2 suppresses immunity against Candida albicans through induction of IL-10 and regulatory T cells. J. Immunol. 2004, 172, 3712–3718. [Google Scholar] [CrossRef] [PubMed]
- Pandiyan, P.; Conti, H.R.; Zheng, L.; Peterson, A.C.; Mathern, D.R.; Hernández-Santos, N.; Edgerton, M.; Gaffen, S.L.; Lenardo, M.J. CD4(+) CD25(+) Foxp3(+) regulatory T cells promote Th17 cells in vitro and enhance host resistance in mouse Candida albicans Th17 cell infection model. Immunity 2011, 34, 422–434. [Google Scholar] [CrossRef] [PubMed]
- McKinley, L.; Logar, A.J.; McAllister, F.; Zheng, M.; Steele, C.; Kolls, J.K. Regulatory T Cells Dampen Pulmonary Inflammation and Lung Injury in an Animal Model of Pneumocystis Pneumonia. J. Immunol. 2006, 177, 6215–6226. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.C.; Goldman, D.L.; Doering, T.L.; Pirofski, L.; Casadevall, A. Antibody response to Cryptococcus neoformans proteins in rodents and humans. Infect. Immun. 1999, 67, 2218–2224. [Google Scholar] [PubMed]
- Deshaw, M.; Pirofski, L.A. Antibodies to the Cryptococcus neoformans capsular glucuronoxylomannan are ubiquitous in serum from HIV+ and HIV− individuals. Clin. Exp. Immunol. 1995, 99, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Goldman, D.L.; Khine, H.; Abadi, J.; Lindenberg, D.J.; Pirofski, L.; Niang, R.; Casadevall, A. Serologic evidence for Cryptococcus neoformans infection in early childhood. Pediatrics 2001, 107, E66. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, K.; Metzger, B.; Hanau, L.H.; Guh, A.; Rucker, L.; Badri, S.; Pirofski, L.-A. IgM(+) Memory B Cell Expression Predicts HIV-Associated Cryptococcosis Status. J. Infect. Dis. 2009, 200, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Ellis, M.; Cesario, T.; Ruhling, M.; Vayuvegula, B. Disseminated cryptococcal infection in a patient with hypogammaglobulinemia and normal T cell functions. Am. J. Med. 1987, 82, 129–131. [Google Scholar] [CrossRef]
- De Gorgolas, M.; Erice, A.; Gil, A.; Gutierrez, J.; Rivas, P.; Hernando, C.; Rodriguez, M.C. Cryptococcal meningitis in a patient with X-linked hyper-IgM1 syndrome. Scand. J. Infect. Dis. 2005, 37, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Iseki, M.; Anzo, M.; Yamashita, N.; Matsuo, N. Hyper-IgM immunodeficiency with disseminated cryptococcosis. Acta Paediatr. 1994, 83, 780–782. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Wuthrich, M.; Deepe, G.; Klein, B. Adaptive immunity to fungi. Cold Spring Harb. Perspect. Med. 2014, 5, a019612. [Google Scholar] [CrossRef] [PubMed]
- Notarangelo, L.D.; Hayward, A.R. X-linked immunodeficiency with hyper-IgM (XHIM). Clin. Exp. Immunol. 2000, 120, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Wahab, J.A.; Hanifah, M.J.; Choo, K.E. Bruton’s agammaglobulinaemia in a child presenting with cryptococcal empyema thoracis and periauricular pyogenic abscess. Singapore Med. J. 1995, 36, 686–689. [Google Scholar] [PubMed]
- Guo, L.-Y.; Liu, L.-L.; Liu, Y.; Chen, T.-M.; Li, S.-Y.; Yang, Y.-H.; Liu, G. Characteristics and outcomes of cryptococcal meningitis in HIV seronegative children in Beijing, China, 2002–2013. BMC Infect. Dis. 2016, 16, 635. [Google Scholar] [CrossRef] [PubMed]
- Browne, S.K.; Burbelo, P.D.; Chetchotisakd, P.; Suputtamongkol, Y.; Kiertiburanakul, S.; Shaw, P.A.; Kirk, J.L.; Jutivorakool, K.; Zaman, R.; Ding, L.; et al. Adult-onset immunodeficiency in Thailand and Taiwan. N. Engl. J. Med. 2012, 367, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Monga, D.P.; Kumar, R.; Mohapatra, L.N.; Malaviya, A.N. Experimental cryptococcosis in normal and B-cell-deficient mice. Infect. Immun. 1979, 26, 1–3. [Google Scholar] [PubMed]
- Rivera, J.; Zaragoza, O.; Casadevall, A. Antibody-mediated protection against Cryptococcus neoformans pulmonary infection is dependent on B cells. Infect. Immun. 2005, 73, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Szymczak, W.A.; Davis, M.J.; Lundy, S.K.; Dufaud, C.; Olszewski, M.; Pirofski, L.A. X-linked immunodeficient mice exhibit enhanced susceptibility to Cryptococcus neoformans Infection. MBio 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, O.; Alvarez, M.; Telzak, A.; Rivera, J.; Casadevall, A. The relative susceptibility of mouse strains to pulmonary Cryptococcus neoformans infection is associated with pleiotropic differences in the immune response. Infect. Immun. 2007, 75, 2729–2739. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, N.; Murphy, J.W. Antibody-dependent natural killer cell-mediated growth inhibition of Cryptococcus neoformans. Infect. Immun. 1986, 51, 556–562. [Google Scholar] [PubMed]
- Aguirre, K.M.; Johnson, L.L. A role for B cells in resistance to Cryptococcus neoformans in mice. Infect. Immun. 1997, 65, 525–530. [Google Scholar] [PubMed]
- Kelly, R.M.; Chen, J.; Yauch, L.E.; Levitz, S.M. Opsonic requirements for dendritic cell-mediated responses to Cryptococcus neoformans. Infect. Immun. 2005, 73, 592–598. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kozel, T.R.; Follette, J.L. Opsonization of encapsulated Cryptococcus neoformans by specific anticapsular antibody. Infect. Immun. 1981, 31, 978–984. [Google Scholar] [PubMed]
- Netski, D.; Kozel, T.R. Fc-dependent and Fc-independent opsonization of Cryptococcus neoformans by anticapsular monoclonal antibodies: Importance of epitope specificity. Infect. Immun. 2002, 70, 2812–2819. [Google Scholar] [CrossRef] [PubMed]
- Taborda, C.P.; Casadevall, A. CR3 (CD11b/CD18) and CR4 (CD11c/CD18) are involved in complement-independent antibody-mediated phagocytosis of Cryptococcus neoformans. Immunity 2002, 16, 791–802. [Google Scholar] [CrossRef]
- Vecchiarelli, A.; Pietrella, D.; Lupo, P.; Bistoni, F.; McFadden, D.C.; Casadevall, A. The polysaccharide capsule of Cryptococcus neoformans interferes with human dendritic cell maturation and activation. J. Leukoc. Biol. 2003, 74, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Feldmesser, M.; Casadevall, A. J774 murine macrophage-like cell interactions with Cryptococcus neoformans in the presence and absence of opsonins. J. Infect. Dis. 1996, 173, 1222–1231. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, J.; Scharff, M.D.; Casadevall, A. Protective murine monoclonal antibodies to Cryptococcus neoformans. Infect. Immun. 1992, 60, 4534–4541. [Google Scholar] [PubMed]
- Mukherjee, S.; Lee, S.C.; Casadevall, A. Antibodies to Cryptococcus neoformans glucuronoxylomannan enhance antifungal activity of murine macrophages. Infect. Immun. 1995, 63, 573–579. [Google Scholar] [PubMed]
- Rachini, A.; Pietrella, D.; Lupo, P.; Torosantucci, A.; Chiani, P.; Bromuro, C.; Proietti, C.; Bistoni, F.; Cassone, A.; Vecchiarelli, A. An anti-beta-glucan monoclonal antibody inhibits growth and capsule formation of Cryptococcus neoformans in vitro and exerts therapeutic, anticryptococcal activity in vivo. Infect. Immun. 2007, 75, 5085–5094. [Google Scholar] [CrossRef] [PubMed]
- Muller, U.; Stenzel, W.; Kohler, G.; Werner, C.; Polte, T.; Hansen, G.; Schutze, N.; Straubinger, R.K.; Blessing, M.; McKenzie, A.N.; et al. IL-13 induces disease-promoting type 2 cytokines, alternatively activated macrophages and allergic inflammation during pulmonary infection of mice with Cryptococcus neoformans. J. Immunol. 2007, 179, 5367–5377. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Dayrit, J.K.; Davis, M.J.; Carolan, J.F.; Osterholzer, J.J.; Curtis, J.L.; Olszewski, M.A. Scavenger Receptor A Modulates the Immune Response to Pulmonary Cryptococcus neoformans Infection. J. Immunol. 2013, 191, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Hernandez, Y.; Erb-Downward, J.R.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Role of IFN- γ in regulating T2 immunity and the development of alternatively activated macrophages during allergic bronchopulmonary mycosis. J. Immunol. 2005, 174, 6346–6356. [Google Scholar] [CrossRef] [PubMed]
- Lovchik, J.A.; Wilder, J.A.; Huffnagle, G.B.; Riblet, R.; Lyons, C.R.; Lipscomb, M.F. Ig heavy chain complex-linked genes influence the immune response in a murine cryptococcal infection. J. Immunol. 1999, 163, 3907–3913. [Google Scholar] [PubMed]
- Kawakami, K.; Tohyama, M.; Xie, Q.; Saito, A. IL-12 protects mice against pulmonary and disseminated infection caused by Cryptococcus neoformans. Clin. Exp. Immunol. 1996, 104, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Beenhouwer, D.O.; Shapiro, S.; Feldmesser, M.; Casadevall, A.; Scharff, M.D. Both Th1 and Th2 cytokines affect the ability of monoclonal antibodies to protect mice against Cryptococcus neoformans. Infect. Immun. 2001, 69, 6445–6455. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, K.L.; Doyle, H.A. Requirement for CD4(+) T lymphocytes in host resistance against Cryptococcus neoformans in the central nervous system of immunized mice. Infect. Immun. 2000, 68, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Gault, R.A.; Kozel, T.R.; Murphy, W.J. Immunomodulation with CD40 stimulation and interleukin-2 protects mice from disseminated cryptococcosis. Infect. Immun. 2006, 74, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Gault, R.A.; Kozel, T.R.; Murphy, W.J. Protection from Direct Cerebral Cryptococcus Infection by Interferon-γ-Dependent Activation of Microglial Cells. J. Immunol. 2007, 178, 5753–5761. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Brouwer, A.E.; Hoogendoorn, E.H.; van der Meer, J.W.; Koolen, M.; Verweij, P.E.; Kullberg, B.J. Two patients with cryptococcal meningitis and idiopathic CD4 lymphopenia: Defective cytokine production and reversal by recombinant interferon-γ therapy. Clin. Infect. Dis. 2004, 39, e83–e87. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Bustamante, B.; Ticona, E.; Hamill, R.J.; Johnson, P.C.; Reboli, A.; Aberg, J.; Hasbun, R.; Hsu, H.H. Recombinant interferon-γ 1b as adjunctive therapy for AIDS-related acute cryptococcal meningitis. J. Infect. Dis. 2004, 189, 2185–2191. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz-Demirdag, Y.; Wilson, B.; Lowery-Nordberg, M.; Bocchini, J.A., Jr.; Bahna, S.L. Interleukin-2 treatment for persistent cryptococcal meningitis in a child with idiopathic CD4(+) T lymphocytopenia. Allergy Asthma Proc. 2008, 29, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Mora, D.J.; Fortunato, L.R.; Andrade-Silva, L.E.; Ferreira-Paim, K.; Rocha, I.H.; Vasconcelos, R.R.; Silva-Teixeira, D.N.; Nascentes, G.A.; Silva-Vergara, M.L. Cytokine profiles at admission can be related to outcome in AIDS patients with cryptococcal meningitis. PLoS ONE 2015, 10, e0120297. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.M.; Reiner, S.L. The lineage decisions of helper T cells. Nat. Rev. Immunol. 2002, 2, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.J.; Sullivan, B.M.; Peng, S.L.; Glimcher, L.H. Molecular mechanisms regulating Th1 immune responses. Annu. Rev. Immunol. 2003, 21, 713–758. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Lim, A.; Omarjee, S.; Levitz, S.M.; Gosnell, B.I.; Spelman, T.; Elliott, J.H.; Carr, W.H.; Moosa, M.Y.; Ndung’u, T.; et al. Cryptococcosis-IRIS is associated with lower Cryptococcus-specific IFN-γ responses before antiretroviral therapy but not higher T-cell responses during therapy. J. Infect. Dis. 2013, 208, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.A.; Brouwer, A.E.; Wuthiekanun, V.; Jaffar, S.; Shattock, R.; Irving, D.; Sheldon, J.; Chierakul, W.; Peacock, S.; Day, N.; et al. IFN-γ at the site of infection determines rate of clearance of infection in cryptococcal meningitis. J. Immunol. 2005, 174, 1746–1750. [Google Scholar] [CrossRef] [PubMed]
- Lortholary, O.; Dromer, F.; Mathoulin-Pelissier, S.; Fitting, C.; Improvisi, L.; Cavaillon, J.M.; Dupont, B. Immune mediators in cerebrospinal fluid during cryptococcosis are influenced by meningeal involvement and human immunodeficiency virus serostatus. J. Infect. Dis. 2001, 183, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Boulware, D.R.; Bonham, S.C.; Meya, D.B.; Wiesner, D.L.; Park, G.S.; Kambugu, A.; Janoff, E.N.; Bohjanen, P.R. Paucity of initial cerebrospinal fluid inflammation in cryptococcal meningitis is associated with subsequent immune reconstitution inflammatory syndrome. J. Infect. Dis. 2010, 202, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Meintjes, G.; Bicanic, T.; Buffa, V.; Hogan, L.; Mo, S.; Tomlinson, G.; Kropf, P.; Noursadeghi, M.; Harrison, T.S. Cerebrospinal fluid cytokine profiles predict risk of early mortality and immune reconstitution inflammatory syndrome in HIV-associated cryptococcal meningitis. PLoS Pathog. 2015, 11, e1004754. [Google Scholar] [CrossRef] [PubMed]
- Blasi, E.; Barluzzi, R.; Mazzolla, R.; Pitzurra, L.; Puliti, M.; Saleppico, S.; Bistoni, F. Biomolecular events involved in anticryptococcal resistance in the brain. Infect. Immun. 1995, 63, 1218–1222. [Google Scholar] [PubMed]
- Kleinschek, M.A.; Muller, U.; Brodie, S.J.; Stenzel, W.; Kohler, G.; Blumenschein, W.M.; Straubinger, R.K.; McClanahan, T.; Kastelein, R.A.; Alber, G. IL-23 enhances the inflammatory cell response in Cryptococcus neoformans infection and induces a cytokine pattern distinct from IL-12. J. Immunol. 2006, 176, 1098–1106. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Qureshi, M.H.; Zhang, T.; Okamura, H.; Kurimoto, M.; Saito, A. IL-18 protects mice against pulmonary and disseminated infection with Cryptococcus neoformans by inducing IFN-γ production. J. Immunol. 1997, 159, 5528–5534. [Google Scholar] [PubMed]
- Kawakami, K.; Koguchi, Y.; Qureshi, M.H.; Kinjo, Y.; Yara, S.; Miyazato, A.; Kurimoto, M.; Takeda, K.; Akira, S.; Saito, A. Reduced host resistance and Th1 response to Cryptococcus neoformans in interleukin-18 deficient mice. FEMS Microbiol. Lett. 2000, 186, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Uicker, W.C.; Doyle, H.A.; McCracken, J.P.; Langlois, M.; Buchanan, K.L. Cytokine and chemokine expression in the central nervous system associated with protective cell-mediated immunity against Cryptococcus neoformans. Med. Mycol. 2005, 43, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Olszewski, M.A.; Tsang, T.M.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Effect of cytokine interplay on macrophage polarization during chronic pulmonary infection with Cryptococcus neoformans. Infect. Immun. 2011, 79, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Boulware, D.R.; Meya, D.B.; Bergemann, T.L.; Wiesner, D.L.; Rhein, J.; Musubire, A.; Lee, S.J.; Kambugu, A.; Janoff, E.N.; Bohjanen, P.R. Clinical Features and Serum Biomarkers in HIV Immune Reconstitution Inflammatory Syndrome after Cryptococcal Meningitis: A Prospective Cohort Study. PLoS Med. 2010, 7, e1000384. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, K.L.; Hardison, S.E.; Kolls, J.K.; Wormley, F.L. Role of IL-17A on resolution of pulmonary C. neoformans infection. PLoS ONE 2011, 6, e17204. [Google Scholar]
- Huffnagle, G.B.; Boyd, M.B.; Street, N.E.; Lipscomb, M.F. IL-5 is required for eosinophil recruitment, crystal deposition, and mononuclear cell recruitment during a pulmonary Cryptococcus neoformans infection in genetically susceptible mice (C57BL/6). J. Immunol. 1998, 160, 2393–2400. [Google Scholar] [PubMed]
- Scriven, J.E.; Rhein, J.; Hullsiek, K.H.; von Hohenberg, M.; Linder, G.; Rolfes, M.A.; Williams, D.A.; Taseera, K.; Meya, D.B.; Meintjes, G.; et al. Early ART After Cryptococcal Meningitis Is Associated With Cerebrospinal Fluid Pleocytosis and Macrophage Activation in a Multisite Randomized Trial. J. Infect. Dis. 2015, 212, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Chaka, W.; Heyderman, R.; Gangaidzo, I.; Robertson, V.; Mason, P.; Verhoef, J.; Verheul, A.; Hoepelman, A.I. Cytokine profiles in cerebrospinal fluid of human immunodeficiency virus-infected patients with cryptococcal meningitis: No leukocytosis despite high interleukin-8 levels. University of Zimbabwe Meningitis Group. J. Infect. Dis. 1997, 176, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Schoffelen, T.; Illnait-Zaragozi, M.-T.; Joosten, L.A.B.; Netea, M.G.; Boekhout, T.; Meis, J.F.; Sprong, T. Cryptococcus gattii Induces a Cytokine Pattern That Is Distinct from Other Cryptococcal Species. PLoS ONE 2013, 8, e55579. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Kadota, J.; Ishimatsu, Y.; Iwashita, T.; Tomono, K.; Kawakami, K.; Kohno, S. Th1–Th2 cytokine kinetics in the bronchoalveolar lavage fluid of mice infected with Cryptococcus neoformans of different virulences. Microbiol. Immunol. 2000, 44, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Li, S.Y.; Luo, Y.F.; Zhai, Y.P.; Wei, X.Q. Changes of Th1/Th2 cytokines in immunocompetent patients with pulmonary cryptococcosis. Genet. Mol. Res. 2013, 12, 5733–5742. [Google Scholar] [CrossRef] [PubMed]
- Lortholary, O.; Sitbon, K.; Dromer, F. Evidence for human immunodeficiency virus and Cryptococcus neoformans interactions in the pro-inflammatory and anti-inflammatory responses in blood during AIDS-associated cryptococcosis. Clin. Microbiol. Infect. 2005, 11, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Huffnagle, G.B.; Strieter, R.M.; Standiford, T.J.; McDonald, R.A.; Burdick, M.D.; Kunkel, S.L.; Toews, G.B. The role of monocyte chemotactic protein-1 (MCP-1) in the recruitment of monocytes and CD4+ T cells during a pulmonary Cryptococcus neoformans infection. J. Immunol. 1995, 155, 4790–4797. [Google Scholar] [PubMed]
- Olszewski, M.A.; Huffnagle, G.B.; Traynor, T.R.; McDonald, R.A.; Cook, D.N.; Toews, G.B. Regulatory effects of macrophage inflammatory protein 1α/CCL3 on the development of immunity to Cryptococcus neoformans depend on expression of early inflammatory cytokines. Infect. Immun. 2001, 69, 6256–6263. [Google Scholar] [CrossRef] [PubMed]
- Gyetko, M.R.; Sud, S.; Chen, G.H.; Fuller, J.A.; Chensue, S.W.; Toews, G.B. Urokinase-type plasminogen activator is required for the generation of a type 1 immune response to pulmonary Cryptococcus neoformans infection. J. Immunol. 2002, 168, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Haddow, L.J.; Colebunders, R.; Meintjes, G.; Lawn, S.D.; Elliott, J.H.; Manabe, Y.C.; Bohjanen, P.R.; Sungkanuparph, S.; Easterbrook, P.J.; French, M.A.; et al. Cryptococcal immune reconstitution inflammatory syndrome in HIV-1-infected individuals: Proposed clinical case definitions. Lancet Infect. Dis. 2010, 10, 791–802. [Google Scholar] [CrossRef]
- Lortholary, O.; Fontanet, A.; Memain, N.; Martin, A.; Sitbon, K.; Dromer, F. Incidence and risk factors of immune reconstitution inflammatory syndrome complicating HIV-associated cryptococcosis in France. AIDS 2005, 19, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Shelburne, S.A.; Darcourt, J., 3rd; White, A.C., Jr.; Greenberg, S.B.; Hamill, R.J.; Atmar, R.L.; Visnegarwala, F. The role of immune reconstitution inflammatory syndrome in AIDS-related Cryptococcus neoformans disease in the era of highly active antiretroviral therapy. Clin. Infect. Dis. 2005, 40, 1049–1052. [Google Scholar] [CrossRef] [PubMed]
- Kambugu, A.; Meya, D.B.; Rhein, J.; O’Brien, M.; Janoff, E.N.; Ronald, A.R.; Kamya, M.R.; Mayanja-Kizza, H.; Sande, M.A.; Bohjanen, P.R.; et al. Outcome of cryptococcal meningitis in Uganda before and after availability of HAART. Clin. Infect. Dis. 2008, 46, 1694–1701. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, D.M.; Venter, W.D.; Feldman, C.; van Rie, A. Incidence and risk factors for the immune reconstitution inflammatory syndrome in HIV patients in South Africa: A prospective study. AIDS 2008, 22, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Lawn, S.D.; Bekker, L.G.; Myer, L.; Orrell, C.; Wood, R. Cryptococcocal immune reconstitution disease: A major cause of early mortality in a South African antiretroviral programme. AIDS 2005, 19, 2050–2052. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.J.; Tsang, T.M.; Qiu, Y.; Dayrit, J.K.; Freij, J.B.; Huffnagle, G.B.; Olszewski, M.A. Macrophage M1/M2 polarization dynamically adapts to changes in cytokine microenvironments in Cryptococcus neoformans infection. MBio 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Koguchi, Y.; Kawakami, K. Cryptococcal infection and Th1–Th2 cytokine balance. Int. Rev. Immunol. 2002, 21, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Casazza, J.; Stone, H.H.; Meintjes, G.; Lawn, S.D.; Levitz, S.M.; Harrison, T.S.; Koup, R.A. The phenotype of the Cryptococcus-specific CD4+ memory T-cell response is associated with disease severity and outcome in HIV-associated cryptococcal meningitis. J. Infect. Dis. 2013, 207, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Okagaki, L.H.; Strain, A.K.; Nielsen, J.N.; Charlier, C.; Baltes, N.J.; Chretien, F.; Heitman, J.; Dromer, F.; Nielsen, K. Cryptococcal cell morphology affects host cell interactions and pathogenicity. PLoS Pathog. 2010, 6, e1000953. [Google Scholar] [CrossRef]
- Zaragoza, O.; Garcia-Rodas, R.; Nosanchuk, J.D.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L.; Casadevall, A. Fungal cell gigantism during mammalian infection. PLoS Pathog. 2010, 6, e1000945. [Google Scholar] [CrossRef]
- Okagaki, L.H.; Nielsen, K. Titan cells confer protection from phagocytosis in Cryptococcus neoformans infections. Eukaryot. Cell 2012, 11, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, J.N.; Okagaki, L.H.; Wiesner, D.L.; Strain, A.K.; Nielsen, J.N.; Nielsen, K. Titan cell production enhances the virulence of Cryptococcus neoformans. Infect. Immun. 2012, 80, 3776–3785. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, O.; Nielsen, K. Titan cells in Cryptococcus neoformans: Cells with a giant impact. Curr. Opin. Microbiol. 2013, 16, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Voelz, K.; Johnston, S.A.; Smith, L.M.; Hall, R.A.; Idnurm, A.; May, R.C. ‘Division of labour’ in response to host oxidative burst drives a fatal Cryptococcus gattii outbreak. Nat. Commun. 2014, 5, 5194. [Google Scholar] [CrossRef] [PubMed]
- Huston, S.M.; Li, S.S.; Stack, D.; Timm-McCann, M.; Jones, G.J.; Islam, A.; Berenger, B.M.; Xiang, R.F.; Colarusso, P.; Mody, C.H. Cryptococcus gattii is killed by dendritic cells, but evades adaptive immunity by failing to induce dendritic cell maturation. J. Immunol. 2013, 191, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Saijo, T.; Chen, J.; Chen, S.C.A.; Rosen, L.B.; Yi, J.; Sorrell, T.C.; Bennett, J.E.; Holland, S.M.; Browne, S.K.; Kwon-Chung, K.J. Anti-Granulocyte-Macrophage Colony-Stimulating Factor Autoantibodies Are a Risk Factor for Central Nervous System Infection by Cryptococcus gattii in Otherwise Immunocompetent Patients. MBio 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Ngamskulrungroj, P.; Chang, Y.; Sionov, E.; Kwon-Chung, K.J. The primary target organ of Cryptococcus gattii is different from that of Cryptococcus neoformans in a murine model. MBio 2012, 3. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Human | Mice | References | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Classification | Cytokines/Chemokines | Blood/Plasma/Serum | CSF | Lungs | Spleen | Brain | |||||
| Protective cytokines | IFNγ | [40,118,121,122,127,128,130,136] | |||||||||
| IL-12 | [40,100,116,124] | ||||||||||
| IL-2 | NA | NA | [119,120,123] | ||||||||
| Protection support cytokines | IL-6 | NA | NA | [100,117,128,129,130] | |||||||
| IL-18 | NA | NA | NA | [134,135] | |||||||
| IL-23 | NA | NA | NA | [133] | |||||||
| IP10 | NA | [127,130,136,137] | |||||||||
| G-CSF | NA | NA | [71,128,130,138,139] | ||||||||
| GM-CSF | NA | NA | NA | [128,131,138] | |||||||
| TNFα | NA | [41,121,128,130,136,138] | |||||||||
| Non-protective cytokines | IL-5 | NA | NA | [47,112,136,140] | |||||||
| IL-13 | NA | NA | [42,47,112,115,141] | ||||||||
| Cytokines/ chemokines with varying roles | IL-1β | NA | NA | [136,142,143] | |||||||
| IL-4 | [42,112,115,117,124,131,136,138,144,145] | ||||||||||
| IL-8 | NA | NA | NA | [124,128,129,130,146] | |||||||
| IL-10 | NA | [47,100,117,124,127,129,131,142,146] | |||||||||
| IL-17 | NA | [39,45,112,124,131,138] | |||||||||
| MCP-1 | NA | NA | [131,136,147,148] | ||||||||
| MIP-1α | NA | NA | NA | [40,131,136,148] | |||||||
| RANTES | NA | NA | NA | [47,136] | |||||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mukaremera, L.; Nielsen, K. Adaptive Immunity to Cryptococcus neoformans Infections. J. Fungi 2017, 3, 64. https://doi.org/10.3390/jof3040064
Mukaremera L, Nielsen K. Adaptive Immunity to Cryptococcus neoformans Infections. Journal of Fungi. 2017; 3(4):64. https://doi.org/10.3390/jof3040064
Chicago/Turabian StyleMukaremera, Liliane, and Kirsten Nielsen. 2017. "Adaptive Immunity to Cryptococcus neoformans Infections" Journal of Fungi 3, no. 4: 64. https://doi.org/10.3390/jof3040064
APA StyleMukaremera, L., & Nielsen, K. (2017). Adaptive Immunity to Cryptococcus neoformans Infections. Journal of Fungi, 3(4), 64. https://doi.org/10.3390/jof3040064