A Novel Assay Reveals a Maturation Process during Ascospore Wall Formation



Abstract
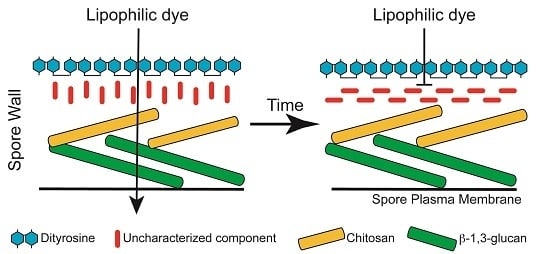
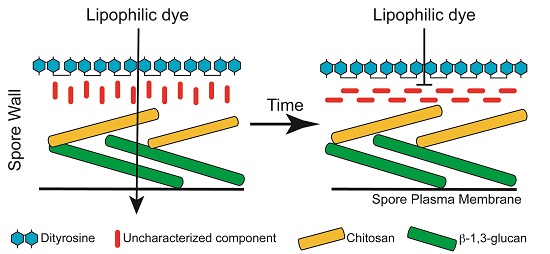
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Plasmids
2.2. MDH Staining Assay
2.3. Dityrosine Fluorescence Assay
2.4. Microscopy
2.5. Ether Tests
3. Results
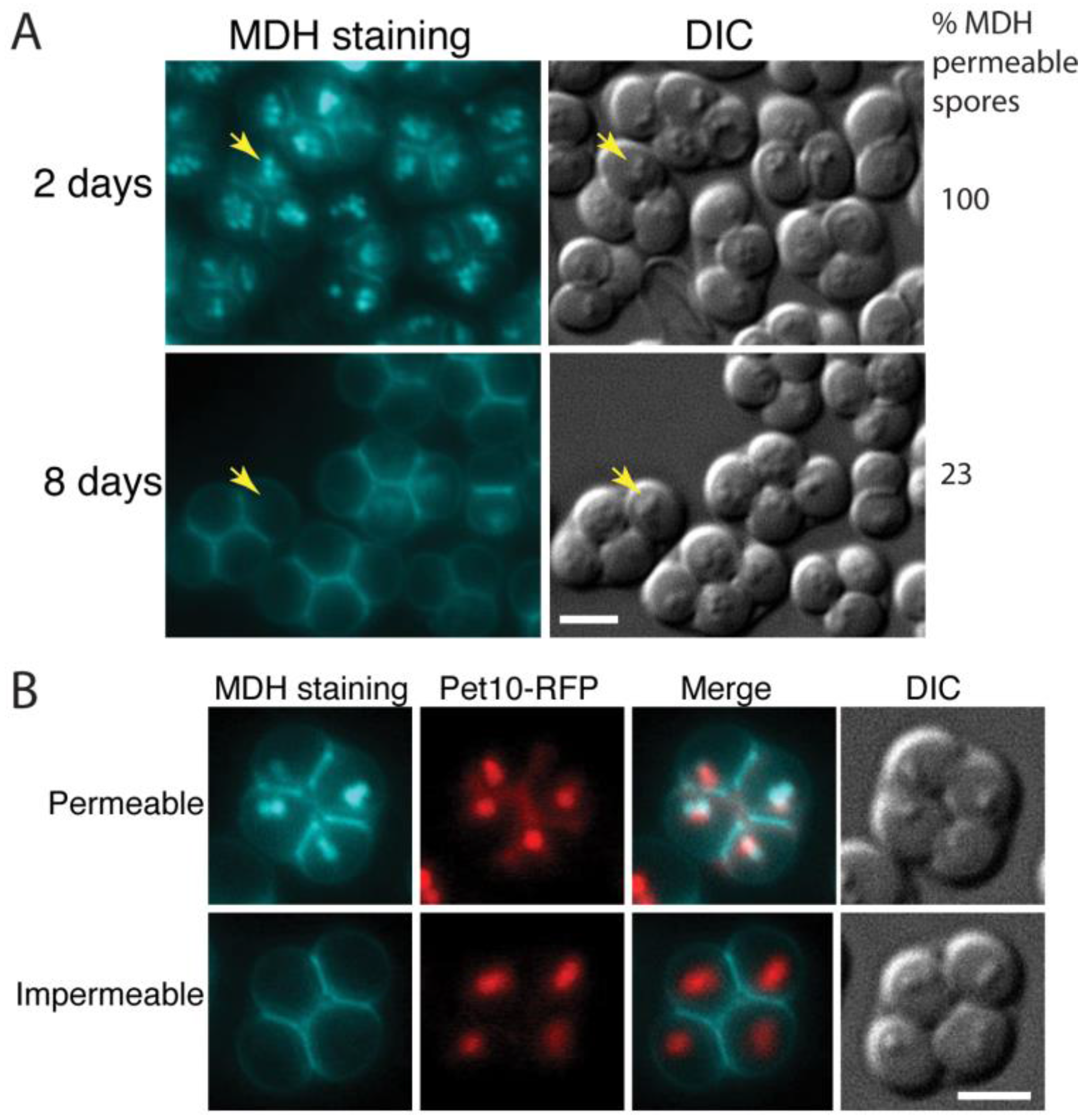
3.1. Staining with Monodansylpentane Reveals a Maturation Process during Spore Wall Development
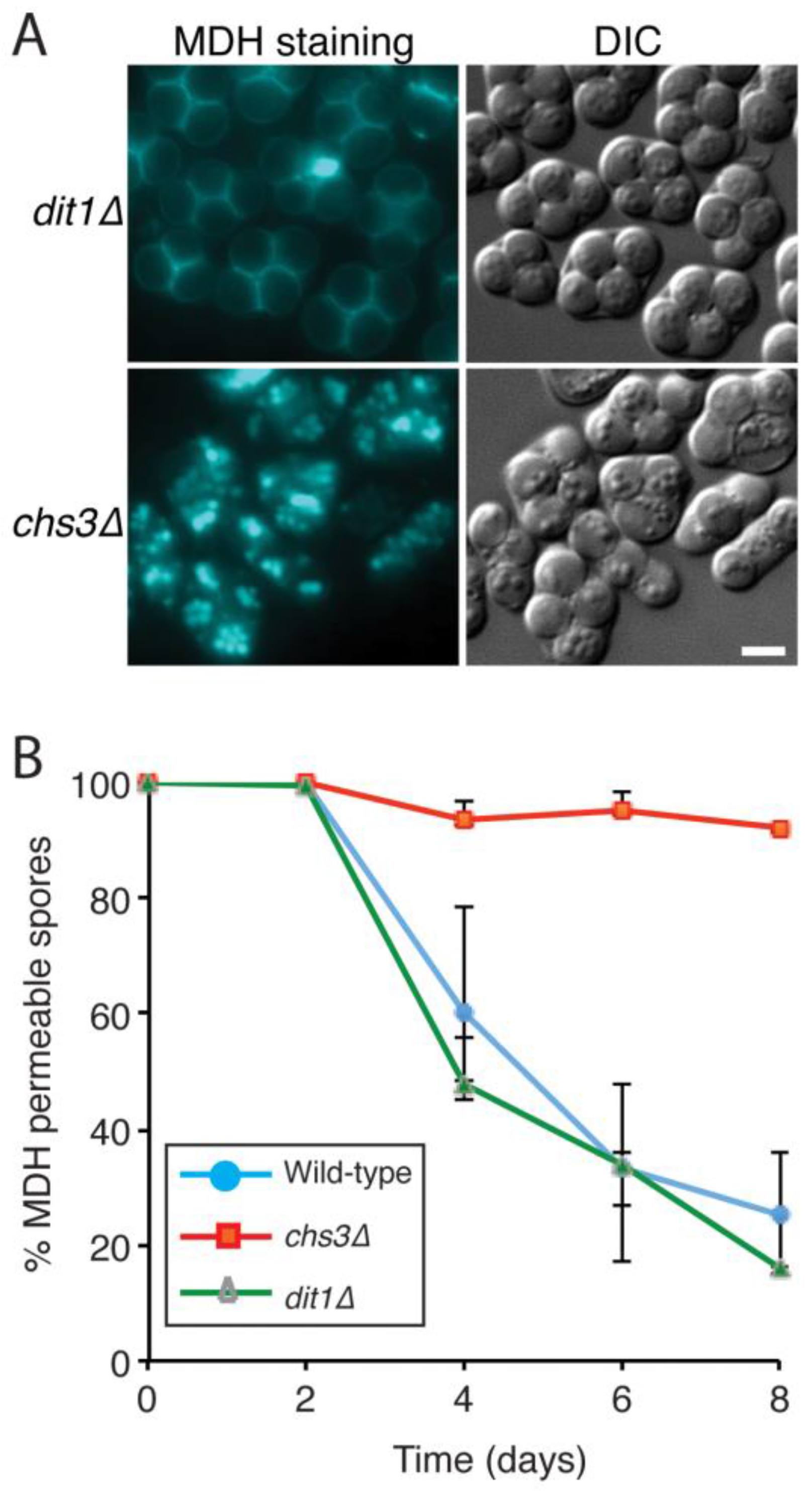
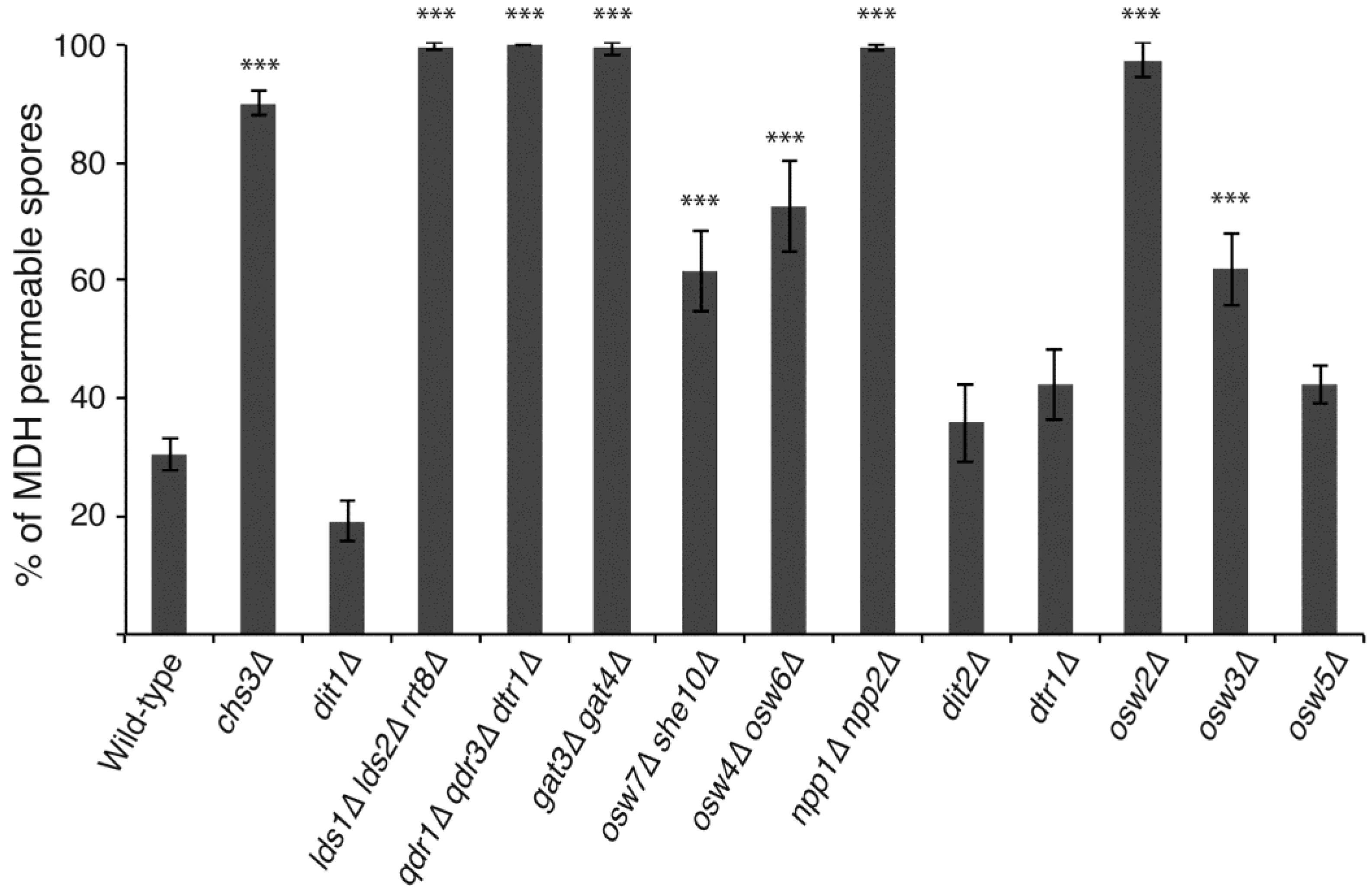
3.2. Acquisition of Impermeability Discriminates between Mutants with Outer Spore Wall Defects
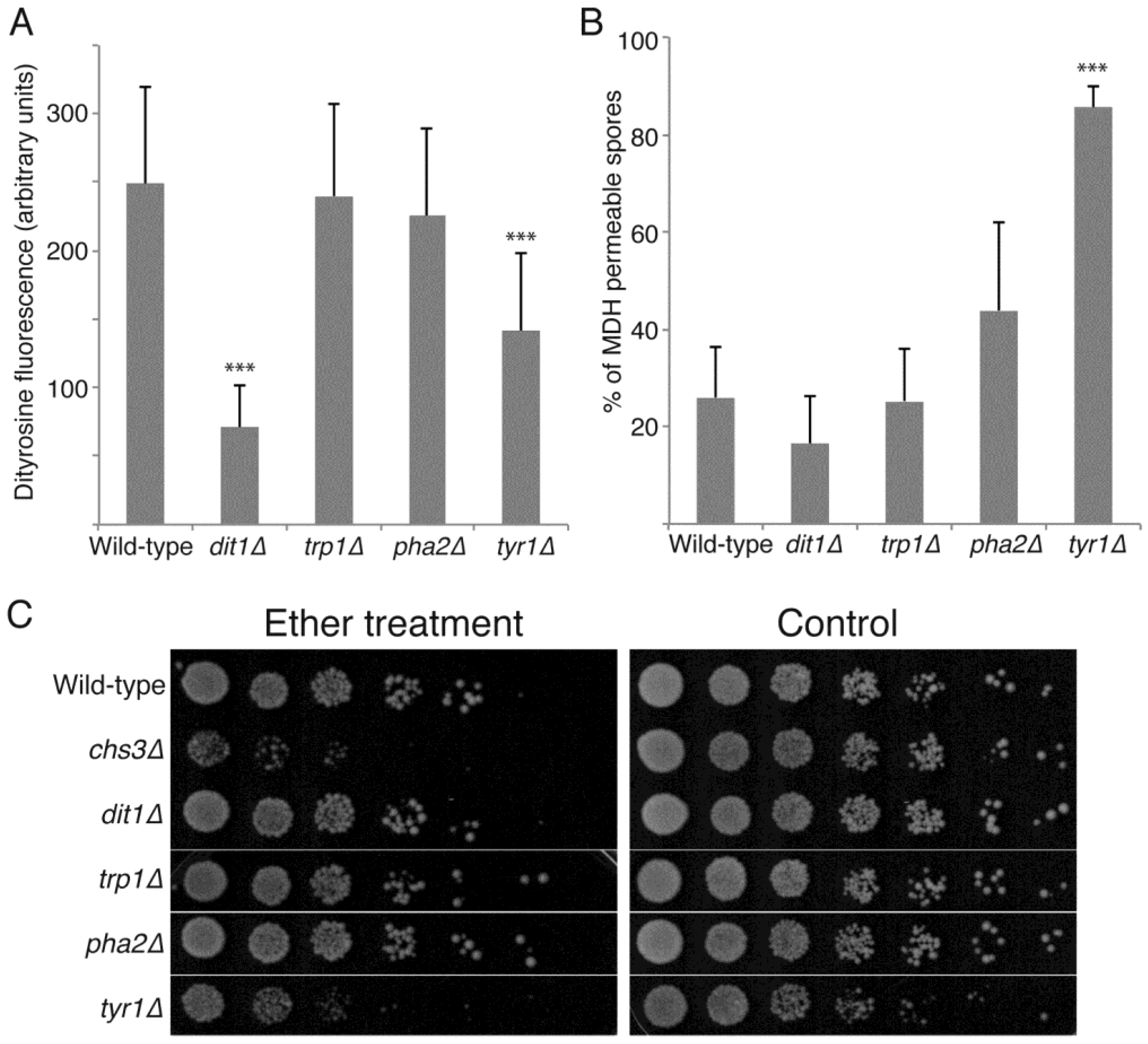
3.3. De Novo Synthesis of Tyrosine Contributes to Dityrosine Layer Assembly and to Additional Properties of the Spore Wall
4. Discussion
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Carlile, M.J.; Watkinson, S.C. The Fungi, 4th ed.; Academic Press: San Diego, CA, USA, 1994. [Google Scholar]
- Neiman, A.M. Sporulation in the budding yeast Saccharomyces cerevisiae. Genetics 2011, 189, 737–765. [Google Scholar] [CrossRef] [PubMed]
- Briza, P.; Ellinger, A.; Winkler, G.; Breitenbach, M. Chemical composition of the yeast ascospore wall—The second outer layer consists of chitosan. J. Biol. Chem. 1988, 263, 11569–11574. [Google Scholar] [PubMed]
- Briza, P.; Winkler, G.; Kalchhauser, H.; Breitenbach, M. Dityrosine is a prominent component of the yeast ascospore wall—A proof of its structure. J. Biol. Chem. 1986, 261, 4288–4294. [Google Scholar] [PubMed]
- Lynn, R.R.; Magee, P.T. Development of the spore wall during ascospore formation in Saccharomyces cerevisiae. J. Cell Biol. 1970, 44, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Christodoulidou, A.; Bouriotis, V.; Thireos, G. Two sporulation-specific chitin deacetylase-encoding genes are required for the ascospore wall rigidity of Saccharomyces cerevisiae. J. Biol. Chem. 1996, 271, 31420–31425. [Google Scholar] [CrossRef] [PubMed]
- Christodoulidou, A.; Briza, P.; Ellinger, A.; Bouriotis, V. Yeast ascospore wall assembly requires two chitin deacetylase isozymes. FEBS Lett. 1999, 460, 275–279. [Google Scholar] [CrossRef]
- Pammer, M.; Briza, P.; Ellinger, A.; Schuster, T.; Stucka, R.; Feldmann, H.; Breitenbach, M. Dit101 (csd2, cal1), a cell cycle-regulated yeast gene required for synthesis of chitin in cell walls and chitosan in spore walls. Yeast 1992, 8, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Briza, P.; Ellinger, A.; Winkler, G.; Breitenbach, M. Characterization of a dl-dityrosine-containing macromolecule from yeast ascospore walls. J. Biol. Chem. 1990, 265, 15118–15123. [Google Scholar] [PubMed]
- Briza, P.; Kalchhauser, H.; Pittenauer, E.; Allmaier, G.; Breitenbach, M. N,N′-bisformyl dityrosine is an in vivo precursor of the yeast ascospore wall. Eur. J. Biochem. 1996, 239, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Briza, P.; Eckerstorfer, M.; Breitenbach, M. The sporulation-specific enzymes encoded by the DIT1 and DIT2 genes catalyze a two-step reaction leading to a soluble ll-dityrosine-containing precursor of the yeast spore wall. Proc. Natl. Acad. Sci. USA 1994, 91, 4524–4528. [Google Scholar] [CrossRef] [PubMed]
- Felder, T.; Bogengruber, E.; Tenreiro, S.; Ellinger, A.; Sa-Correia, I.; Briza, P. Dtrlp, a multidrug resistance transporter of the major facilitator superfamily, plays an essential role in spore wall maturation in Saccharomyces cerevisiae. Eukaryot. Cell 2002, 1, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.P.; Kim, C.; Smith, S.O.; Neiman, A.M. A highly redundant gene network controls assembly of the outer spore wall in S. cerevisiae. PLoS Genet. 2013, 9, e1003700. [Google Scholar] [CrossRef] [PubMed]
- Coluccio, A.E.; Rodriguez, R.K.; Kernan, M.J.; Neiman, A.M. The yeast spore wall enables spores to survive passage through the digestive tract of Drosophila. PLoS ONE 2008, 3, e2873. [Google Scholar] [CrossRef] [PubMed]
- Dawes, I.W.; Hardie, I.D. Selective killing of vegetative cells in sporulated yeast cultures by exposure to diethyl ether. Mol. Gen. Genet. 1974, 131, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Suda, Y.; Rodriguez, R.K.; Coluccio, A.E.; Neiman, A.M. A screen for spore wall permeability mutants identifies a secreted protease required for proper spore wall assembly. PLoS ONE 2009, 4, e7184. [Google Scholar] [CrossRef] [PubMed]
- Coluccio, A.; Bogengruber, E.; Conrad, M.N.; Dresser, M.E.; Briza, P.; Neiman, A.M. Morphogenetic pathway of spore wall assembly in Saccharomyces cerevisiae. Eukaryot. Cell 2004, 3, 1464–1475. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.D.; Fink, G.R. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1990. [Google Scholar]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [PubMed]
- Huh, W.K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.L.; McCusker, J.H. Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 1999, 15, 1541–1553. [Google Scholar] [CrossRef]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Neiman, A.M.; Katz, L.; Brennwald, P.J. Identification of domains required for developmentally regulated SNARE function in Saccharomyces cerevisiae. Genetics 2000, 155, 1643–1655. [Google Scholar] [PubMed]
- Coluccio, A.; Neiman, A.M. Interspore bridges: A new feature of the Saccharomyces cerevisiae spore wall. Microbiology 2004, 150, 3189–3196. [Google Scholar] [CrossRef] [PubMed]
- Rabitsch, K.P.; Toth, A.; Galova, M.; Schleiffer, A.; Schaffner, G.; Aigner, E.; Rupp, C.; Penkner, A.M.; Moreno-Borchart, A.C.; Primig, M.; et al. A screen for genes required for meiosis and spore formation based on whole-genome expression. Curr. Biol. 2001, 11, 1001–1009. [Google Scholar] [CrossRef]
- Currie, E.; Guo, X.; Christiano, R.; Chitraju, C.; Kory, N.; Harrison, K.; Haas, J.; Walther, T.C.; Farese, R.V., Jr. High confidence proteomic analysis of yeast LDs identifies additional droplet proteins and reveals connections to dolichol synthesis and sterol acetylation. J. Lipid Res. 2014, 55, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Pei-Chen Lin, C.; Pathak, M.C.; Temple, B.R.; Nile, A.H.; Mousley, C.J.; Duncan, M.C.; Eckert, D.M.; Leiker, T.J.; Ivanova, P.T.; et al. A phosphatidylinositol transfer protein integrates phosphoinositide signaling with lipid droplet metabolism to regulate a developmental program of nutrient stress-induced membrane biogenesis. Mol. Biol. Cell. 2014, 25, 712–727. [Google Scholar] [CrossRef] [PubMed]
- Briza, P.; Breitenbach, M.; Ellinger, A.; Segall, J. Isolation of two developmentally regulated genes involved in spore wall maturation in Saccharomyces cerevisiae. Genes Dev. 1990, 4, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.; Letisse, F.; Peyriga, L.; Alkim, C.; Liu, Y.; Lardenois, A.; Martin-Yken, H.; Portais, J.C.; Primig, M.; Francois, J. Developmental stage dependent metabolic regulation during meiotic differentiation in budding yeast. BMC Biol. 2014, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.; Santore, E.; Lavoie, E.; Needleman, L.; Fiacco, N.; Kim, C.; Neiman, A.M. A visual screen of protein localization during sporulation identifies new components of prospore membrane-associated complexes in budding yeast. Eukaryot. Cell 2014, 13, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Arellano, M.; Cartagena-Lirola, H.; Nasser Hajibagheri, M.A.; Duran, A.; Henar Valdivieso, M. Proper ascospore maturation requires the chs1+ chitin synthase gene in Schizosaccharomyces pombe. Mol. Microbiol. 2000, 35, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Tanaka, K.; Matsuda, H.; Kawamukai, M. cda1+, encoding chitin deacetylase is required for proper spore formation in Schizosaccharomyces pombe. FEBS Lett. 2005, 579, 2737–2743. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Tanaka, K.; Nakagawa, T.; Matsuda, H.; Kawamukai, M. Genetic analysis of chs1+ and chs2+ encoding chitin synthases from Schizosaccharomyces pombe. Biosci. Biotechnol. Biochem. 2004, 68, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Martin-Castellanos, C.; Blanco, M.; Rozalen, A.E.; Perez-Hidalgo, L.; Garcia, A.I.; Conde, F.; Mata, J.; Ellermeier, C.; Davis, L.; San-Segundo, P.; et al. A large-scale screen in S. pombe identifies seven novel genes required for critical meiotic events. Curr. Biol. 2005, 15, 2056–2062. [Google Scholar] [CrossRef] [PubMed]
- Fukunishi, K.; Miyakubi, K.; Hatanaka, M.; Otsuru, N.; Hirata, A.; Shimoda, C.; Nakamura, T. The fission yeast spore is coated by a proteinaceous surface layer comprising mainly Isp3. Mol. Biol. Cell 2014, 25, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Smail, E.H.; Briza, P.; Panagos, A.; Berenfeld, L. Candida albicans cell walls contain the fluorescent cross-linking amino acid dityrosine. Infect. Immun. 1995, 63, 4078–4083. [Google Scholar] [PubMed]
- Prillinger, H.; Schweigkofler, W.; Breitenbach, M.; Briza, P.; Staudacher, E.; Lopandic, K.; Molnar, O.; Weigang, F.; Ibl, M.; Ellinger, A. Phytopathogenic filamentous (Ashbya, Eremothecium) and dimorphic fungi (Holleya, Nematospora) with needle-shaped ascospores as new members within the Saccharomycetaceae. Yeast 1997, 13, 945–960. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| AN120 | MATa/MATα ura3/ura3 leu2/leu2 his3∆SK/his3∆SK trp1::hisG/trp1::hisG ARG4/arg4-NspI lys2/lys2 hoΔ::LYS2/hoΔ::LYS2 RME1/rme1Δ::LEU2 | [24] |
| AN117-4B | MATα ura3 leu2 his3∆SK trp1::hisG arg4-NspI lys2 ho∆::LYS2 rme1Δ::LEU2 | [24] |
| AN117-16D | MATa ura3 leu2 his3∆SK trp1::hisG lys2 hoΔ::LYS2 | [24] |
| yKZ70 | as AN117-16D plus PET10::PET10-RFP::TRP1 | this study |
| yKZ72 | as AN117-4B plus PET10::PET10-RFP::TRP1 | this study |
| yKZ104 | as AN120, plus PET10::PET10-RFP::TRP1/PET10::PET10-RFP::TRP1 | this study |
| AN262 | as AN120, plus chs3∆::HIS3MX6/chs3∆::HIS3MX6 | [25] |
| AN264 | as AN120, plus dit1∆::HIS3MX6/dit1∆::HIS3MX6 | [25] |
| CL6 | MATa/MATα HO/HO leu2/leu2 lys2/lys2 URA3::tet0224/URA3::tet0224 lds1∆::HIS3MX6/lds1∆::HIS3MX6 rrt8∆::kanMX6/rrt8∆::kanMX6 lds2∆::hphMX6/lds2∆::hphMX6 | [14] |
| CL7 | MATa/MATα HO/HO leu2/leu2 lys2/lys2 URA3::tet0224/URA3::tet0224 dtr1∆::HIS3MX6/dtr1∆::HIS3MX6 qdr3∆::kanMX6/qdr3∆::kanMX6 qdr1∆::hphMX6/qdr1∆::hphMX6 | [14] |
| CL15 | MATa/MATα HO/HO leu2/leu2 trp1/trp1 lys2/lys2 URA3::tet0224/URA3::tet0224 gat4∆::HIS3MX6/gat4∆::HIS3MX6 gat3∆::kanMX6/gat3∆::kanMX6 | [14] |
| CL26 | MATa/MATα HO/HO leu2/leu2 URA3::tet0224/URA3::tet0224 osw7∆::HISMX63/osw7∆::HIS3MX6 she10∆::kanMX6/she10∆::kanMX6 | [14] |
| CL35 | as AN120, plus osw4,6Δ::HIS3MX6/osw4,6Δ::HIS3MX6 | [14] |
| CL57 | MATa/MATα HO/HO leu2/leu2 lys2/lys2 URA3:tet0224/URA3:tet0224 npp2∆::HIS3MX6/npp2∆::HIS3MX6 npp1∆::kanMX6/npp1∆::kanMX6 | [14] |
| ySZ233 | as AN120, plus dit2∆::kanMX6/dit2∆::kanMX6 | this study |
| ySZ265 | as AN120, plus osw2∆::hphMX4/osw2∆hphMX4 | this study |
| MYA-1810 | MATa/MATa HO/HO his3/his3 trp1/trp1 lys2/lys2 LEU2::PURA3-tetR-GFP/LEU2::PURA3-tetR-GFP REC8::HA3-URA3/REC8::HA3-URA3 URA3::tetO224/URA3::tetO224 dtr1∆::HIS3MX6/dtr1∆::HIS3MX6 | [26] |
| MYA-1824 | MATa/MATa HO/HO his3/his3 trp1/trp1 lys2/lys2 LEU2::PURA3-tetR-GFP/LEU2::PURA3-tetR-GFP REC8::HA3-URA3/REC8::HA3-URA3 URA3::tetO224/URA3::tetO224 osw3∆::HIS3MX6/osw3∆::HIS3MX6 | [26] |
| MYA-2022 | MATa/MATa HO/HO his3/his3 trp1/trp1 lys2/lys2 LEU2::PURA3-tetR-GFP/LEU2::PURA3-tetR-GFP REC8::HA3-URA3/REC8::HA3-URA3 URA3::tetO224/URA3::tetO224 osw5∆::HIS3MX6/osw5∆::HIS3MX6 | [26] |
| yKZ107 | as AN120, plus trp1::hisG::TRP1/trp1::hisG | this study |
| yKZ108 | as AN120, plus pha2∆::kanMX6/pha2∆::kanMX6 trp1::hisG::TRP1/trp1::hisG | this study |
| yKZ110 | as AN120, plus tyr1∆::kanMX6/tyr1∆::kanMX6 trp1::hisG::TRP1/trp1::hisG | this study |
| yZK111 | as AN262, plus trp1::hisG::TRP1/trp1::hisG | this study |
| yKZ112 | as AN264, plus trp1::hisG::TRP1/trp1::hisG | this study |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Needleman, L.; Zhou, S.; Neiman, A.M. A Novel Assay Reveals a Maturation Process during Ascospore Wall Formation. J. Fungi 2017, 3, 54. https://doi.org/10.3390/jof3040054
Zhang K, Needleman L, Zhou S, Neiman AM. A Novel Assay Reveals a Maturation Process during Ascospore Wall Formation. Journal of Fungi. 2017; 3(4):54. https://doi.org/10.3390/jof3040054
Chicago/Turabian StyleZhang, Kai, Leor Needleman, Sai Zhou, and Aaron M. Neiman. 2017. "A Novel Assay Reveals a Maturation Process during Ascospore Wall Formation" Journal of Fungi 3, no. 4: 54. https://doi.org/10.3390/jof3040054
APA StyleZhang, K., Needleman, L., Zhou, S., & Neiman, A. M. (2017). A Novel Assay Reveals a Maturation Process during Ascospore Wall Formation. Journal of Fungi, 3(4), 54. https://doi.org/10.3390/jof3040054