
UV-Induced Mutants of Metarhizium anisopliae: Improved Biological Parameters, Resistance to Stressful Factors, and Comparative Transcriptomic Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Entomopathogenic Fungi and Insects
2.2. Monospore Strain Isolation
2.3. Conidial Production and Conidial Suspension
2.4. UV-Induced Fungal Mutants
2.5. Fungal Growth and Conidial Productivity of UV-Induced Mutants
2.6. The Tolerance of UV Mutant to Environmental Stresses
2.7. Bioassay
2.8. RNA Isolation, Library Preparation, and RNA Sequencing
2.9. Quantitative PCR Verification
2.10. Statistical Analysis
3. Results
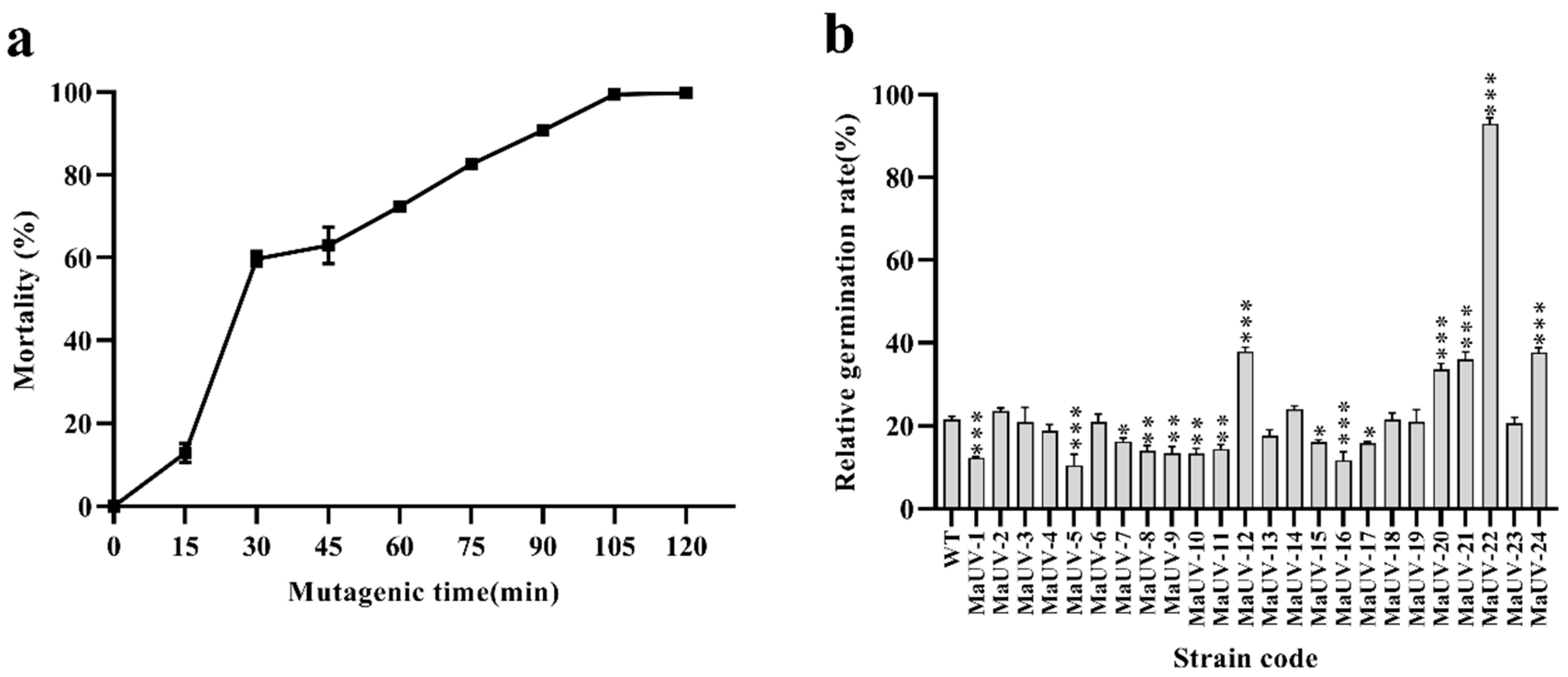
3.1. UV-Induced Mutants
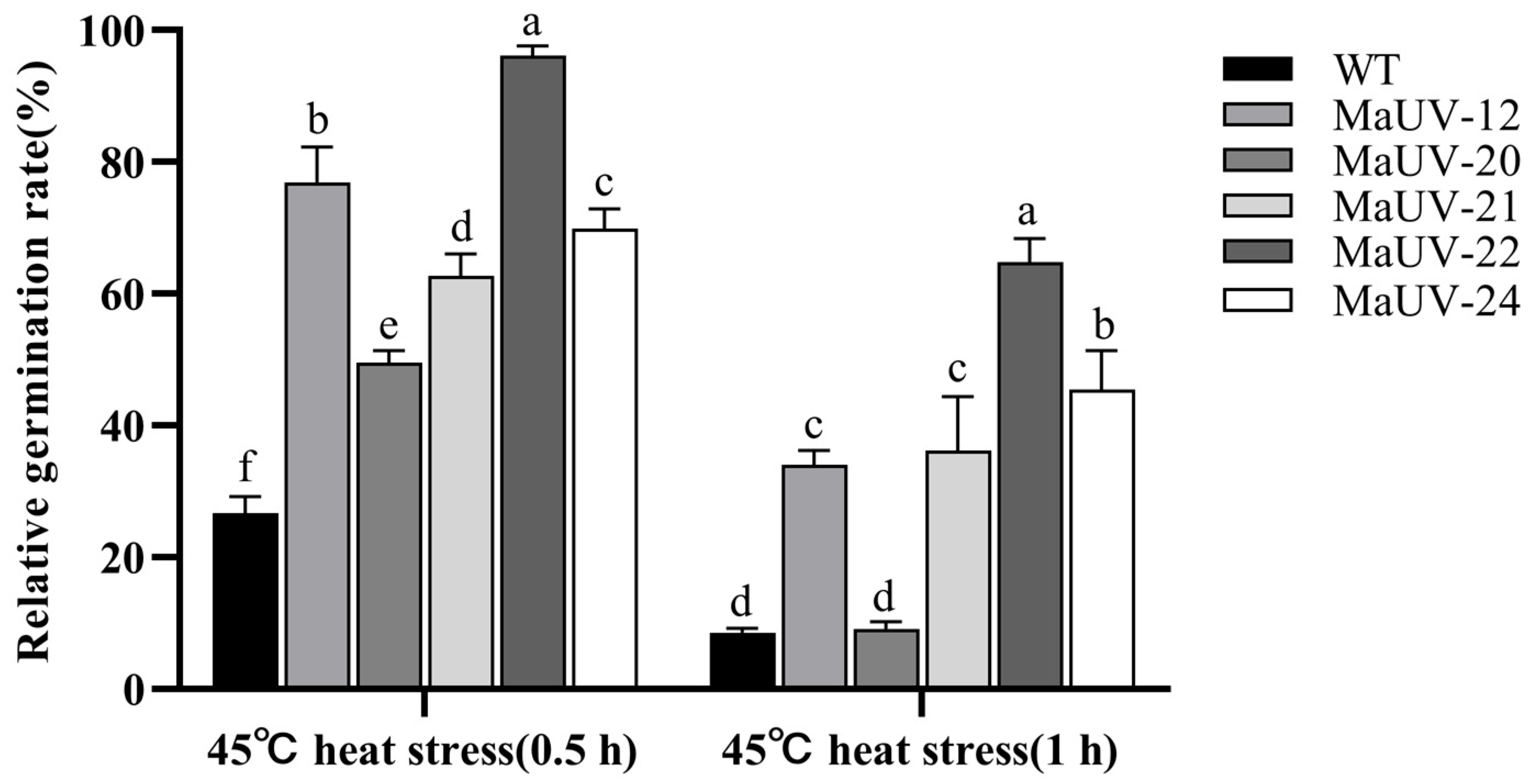
3.2. Fungal Growth, Conidial Productivity, and Resistance to Environmental Stresses of the UV-Induced Mutants
3.3. Fungal Bioassays
3.4. RNA-Sequencing and DEG Analyses
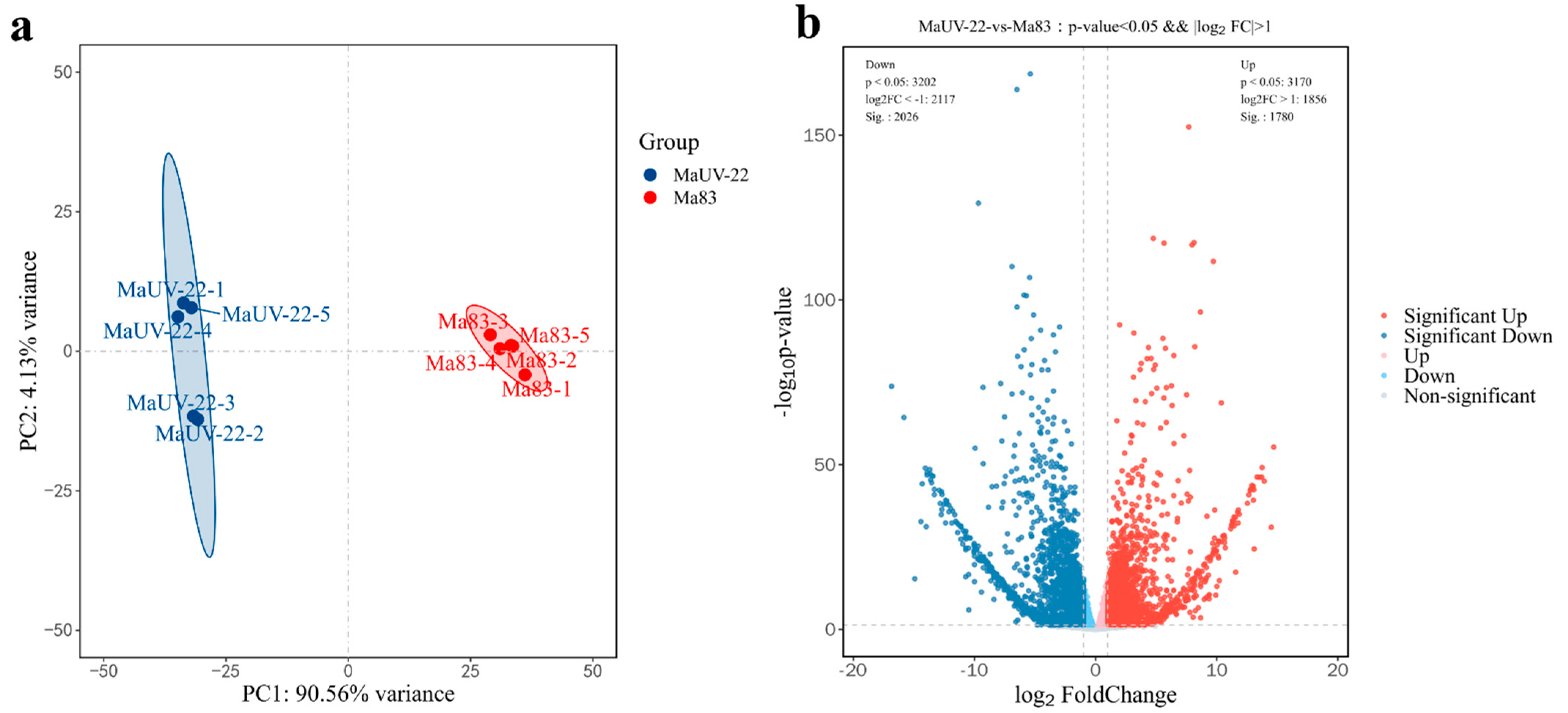
3.4.1. RNA-Sequencing
3.4.2. Enrichment Analysis of DEGs
3.4.3. Quantitative PCR Validation of DEGs
3.4.4. Expression Levels of Stress-Resistant and Virulence-Related Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oskoei, P.; Marçal, R.; Oliveira, H.; Guilherme, S. Hitting two targets with one shot on pesticide genotoxicity assessment–Identifying risk while unveiling ex vivo approach as a throughput tool in gill-breathing animals. J. Hazard. Mater. 2024, 476, 134948. [Google Scholar] [CrossRef] [PubMed]
- Humber, R.A. Evolution of entomopathogenicity in fungi. J. Invertebr. Pathol. 2008, 98, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Ortiz Urquiza, A.; Riveiro Miranda, L.; Santiago Álvarez, C.; Quesada Moraga, E.; Quesada Moraga, E. Insect-toxic secreted proteins and virulence of the entomopathogenic fungus Beauveria bassiana. J. Invertebr. Pathol. 2010, 105, 270–278. [Google Scholar] [CrossRef]
- Ortiz Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Pedrini, N.; Ortiz-Urquiza, A.; Huarte-Bonnet, C.; Fan, Y.; Juárez, M.P.; Keyhani, N.O. Tenebrionid secretions and a fungal benzoquinone oxidoreductase form competing components of an arms race between a host and pathogen. Proc. Natl. Acad. Sci. USA 2015, 112, E3651–E3660. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef] [PubMed]
- Muñiz Paredes, F.; Miranda Hernández, F.; Loera, O. Production of conidia by entomopathogenic fungi: From inoculants to final quality tests. World J. Microbiol. Biotechnol. 2017, 33, 57. [Google Scholar] [CrossRef]
- Hunter, D.M.; Milner, R.J.; Spurgin, P.A. Aerial treatment of the Australian plague locust, Chortoicetes terminifera (Orthoptera: Acrididae) with Metarhizium anisopliae (Deuteromycotina: Hyphomycetes). Bull. Entomol. Res. 2001, 91, 93–99. [Google Scholar] [CrossRef]
- Peng, G.X.; Wang, Z.K.; Yin, Y.P.; Zeng, D.Y.; Xia, Y.X. Field trials of Metarhizium anisopliae var. acridum (Ascomycota: Hypocreales) against oriental migratory locusts, Locusta migratoria manilensis (Meyen) in Northern China. Crop Protect. 2008, 27, 1244–1250. [Google Scholar] [CrossRef]
- Price, R.E.; Bateman, R.P.; Brown, H.D.; Butler, E.T.; Müller, E.J. Aerial spray trials against brown locust (Locustana pardalina, Walker) nymphs in South Africa using oil-based formulations of Metarhizium flavoviride. Crop Protect. 1997, 16, 345–351. [Google Scholar] [CrossRef]
- Méndez González, F.; Castillo Minjarez, J.M.; Loera, O.; Favela Torres, E. Current developments in the resistance, quality, and production of entomopathogenic fungi. World J. Microbiol. Biotechnol. 2022, 38, 115. [Google Scholar] [CrossRef] [PubMed]
- Ekesi, S.; Maniania, N.K.; Ampong Nyarko, K. Effect of temperature on germination, radial growth and virulence of Metarhizium anisopliae and Beauveria bassiana on Megalurothrips sjostedti. Biocontrol Sci. Technol. 1999, 9, 177–185. [Google Scholar] [CrossRef]
- Santos, M.P.; Dias, L.P.; Ferreira, P.C.; Pasin, L.A.; Rangel, D.E. Cold activity and tolerance of the entomopathogenic fungus Tolypocladium spp. to UV-B irradiation and heat. J. Invertebr. Pathol. 2011, 108, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, É.K.K.; Rangel, D.E.N.; Moraes, Á.M.L.; Bittencourt, V.R.E.P.; Roberts, D.W. Variability in tolerance to UV-B radiation among Beauveria spp. isolates. J. Invertebr. Pathol. 2007, 96, 237–243. [Google Scholar] [CrossRef]
- Fernandes, E.K.; Moraes, A.M.; Pacheco, R.S.; Rangel, D.E.; Miller, M.P.; Bittencourt, V.R.; Roberts, D.W. Genetic diversity among Brazilian isolates of Beauveria bassiana: Comparisons with non-Brazilian isolates and other Beauveria species. J. Appl. Microbiol. 2009, 107, 760–774. [Google Scholar] [CrossRef]
- Fernandes, É.K.K.; Rangel, D.E.N.; Braga, G.U.L.; Roberts, D.W. Tolerance of entomopathogenic fungi to ultraviolet radiation: A review on screening of strains and their formulation. Curr. Genet. 2015, 61, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Rangel, D.E.; Braga, G.U.; Fernandes, É.K.; Keyser, C.A.; Hallsworth, J.E.; Roberts, D.W. Stress tolerance and virulence of insect-pathogenic fungi are determined by environmental conditions during conidial formation. Curr. Genet. 2015, 61, 383–404. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Anderson, A.J.; Roberts, D.W. Growth of Metarhizium anisopliae on non-preferred carbon sources yields conidia with increased UV-B tolerance. J. Invertebr. Pathol. 2006, 93, 127–134. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Fernandes, É.K.K.; Anderson, A.J.; Roberts, D.W. Culture of Metarhizium robertsii on salicylic-acid supplemented medium induces increased conidial thermotolerance. Fungal Biol. 2012, 116, 438–442. [Google Scholar] [CrossRef]
- Huarte-Bonnet, C.; Pereira-Junior, R.A.; Paixão, F.R.A.; Braga, G.U.L.; Roberts, D.W.; Luz, C.; Pedrini, N.; Fernandes, É.K.K. Metarhizium robertsii and M. acridum conidia produced on riboflavin-supplemented medium have increased UV-A tolerance and upregulated photoprotection and photoreactivation genes. BioControl 2020, 65, 211–222. [Google Scholar] [CrossRef]
- Hallsworth, J.E.; Magan, N. Culture age, temperature, and pH affect the polyol and trehalose contents of fungal propagules. Appl. Environ. Microbiol. 1996, 62, 2435–2442. [Google Scholar] [CrossRef]
- Dias, L.P.; Souza, R.K.F.; Pupin, B.; Rangel, D.E.N. Conidiation under illumination enhances conidial tolerance of insect-pathogenic fungi to environmental stresses. Fungal Biol. 2021, 125, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Lu, H.L.; Fang, W.; St Leger, R.J. Overexpression of a Metarhizium robertsii HSP25 gene increases thermotolerance and survival in soil. Appl. Microbiol. Biotechnol. 2014, 98, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Wei, X.Y.; Wang, S.J.; Wang, J.J. Characterization of trehalose-6-phosphate phosphatase in trehalose biosynthesis, asexual development, stress resistance and virulence of an insect mycopathogen. Pestic. Biochem. Physiol. 2020, 163, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Colvin, M.E.; Hatch, F.T.; Felton, J.S. Chemical and biological factors affecting mutagen potency. Mutat. Res. 1998, 400, 479–492. [Google Scholar] [CrossRef]
- Fitriana, Y.; Satoh, K.; Narumi, I.; Saito, T. Ion-beam and gamma-ray irradiations induce thermotolerant mutants in the entomopathogenic fungus Metarhizium anisopliae s.l. Biocontrol Sci. Technol. 2014, 24, 1052–1061. [Google Scholar] [CrossRef]
- Sun, Y.X.; Zhang, B.X.; Zhang, W.T.; Wang, Q.; Toufeeq, S.; Rao, X.J. UV-induced mutagenesis of Beauveria bassiana (Hypocreales: Clavicipitaceae) yields two hypervirulent isolates with different transcriptomic profiles. Pest Manag. Sci. 2023, 79, 2762–2779. [Google Scholar] [CrossRef]
- Qiu, L.; Nie, S.X.; Hu, S.J.; Wang, S.J.; Wang, J.J.; Guo, K. Screening of Beauveria bassiana with high biocontrol potential based on ARTP mutagenesis and high-throughput FACS. Pestic. Biochem. Physiol. 2021, 171, 104732. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Araújo, C.A.S.; Dias, L.P.; Ferreira, P.C.; Mittmann, J.; Pupin, B.; Brancini, G.T.P.; Braga, G.Ú.L.; Rangel, D.E.N. Responses of entomopathogenic fungi to the mutagen 4-nitroquinoline 1-oxide. Fungal Biol. 2018, 122, 621–628. [Google Scholar] [CrossRef]
- St. Leger, R.J.; Wang, C. Genetic engineering of fungal biocontrol agents to achieve greater efficacy against insect pests. Appl. Microbiol. Biotechnol. 2010, 85, 901–907. [Google Scholar] [CrossRef]
- Wang, L.; Li, C.; Xiong, P.; Gershenwald, J.E.; Prieto, V.G.; Duvic, M.; Lee, J.E.; Grimm, E.A.; Hsu, T.C.; Wei, Q. 4-Nitroquinoline-1-oxide-induced mutagen sensitivity and risk of cutaneous melanoma: A case–control analysis. Melanoma Res. 2016, 26, 181–187. [Google Scholar] [CrossRef]
- Bai, P.; Hegedus, C.; Erdélyi, K.; Szabó, E.; Bakondi, E.; Gergely, S.; Szabó, C.; Virág, L. Protein tyrosine nitration and poly(ADP-ribose) polymerase activation in N-methyl-N-nitro-N-nitrosoguanidine-treated thymocytes: Implication for cytotoxicity. Toxicol. Lett. 2007, 170, 203–213. [Google Scholar] [CrossRef]
- Philips, J.G.; Martin Avila, E.; Robold, A.V. Horizontal gene transfer from genetically modified plants-Regulatory considerations. Front. Bioeng. Biotechnol. 2022, 10, 971402. [Google Scholar] [CrossRef]
- Eckerstorfer, M.F.; Dolezel, M.; Heissenberger, A.; Miklau, M.; Reichenbecher, W.; Steinbrecher, R.A.; Waßmann, F. An EU perspective on biosafety considerations for plants developed by genome editing and other new genetic modification techniques (nGMs). Front. Bioeng. Biotechnol. 2019, 7, 31. [Google Scholar]
- Eckerstorfer, M.F.; Engelhard, M.; Heissenberger, A.; Simon, S.; Teichmann, H. Plants developed by new genetic modification techniques—Comparison of existing regulatory frameworks in the EU and Non-EU countries. Front. Bioeng. Biotechnol. 2019, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yao, R.; Wei, Y.; Huang, S.; Keyhani, N.O.; Huang, Z. Screening of Metarhizium anisopliae UV-induced mutants for faster growth yields a hyper-virulent isolate with greater UV and thermal tolerances. Appl. Microbiol. Biotechnol. 2016, 100, 9217–9228. [Google Scholar] [CrossRef]
- Erkorkmaz, B.A.; Kırtel, O.; Abaramak, G.; Nikerel, E.; Toksoy Öner, E. UV and chemically induced Halomonas smyrnensis mutants for enhanced levan productivity. J. Biotechnol. 2022, 356, 19–29. [Google Scholar] [CrossRef]
- Rangel, D.E.N.; Finlay, R.D.; Hallsworth, J.E.; Dadachova, E.; Gadd, G.M. Fungal strategies for dealing with environment- and agriculture-induced stresses. Fungal Biol. 2018, 122, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A.; Keyhani, N.O. Stress response signaling and virulence: Insights from entomopathogenic fungi. Curr. Genet. 2015, 61, 239–249. [Google Scholar] [CrossRef]
- Agustinho, D.P.; Miller, L.C.; Li, L.X.; Doering, T.L. Peeling the onion: The outer layers of Cryptococcus neoformans. Memórias Inst. Oswaldo Cruz 2018, 113, e180040. [Google Scholar] [CrossRef]
- Douglas, L.M.; Konopka, J.B. Plasma membrane organization promotes virulence of the human fungal pathogen Candida albicans. J. Microbiol. 2016, 54, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Li, S.Y.; Wang, J.; Liu, Y.L.; Deng, Y. Roles of high osmolarity glycerol and cell wall Integrity pathways in cadmium toxicity in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 6169. [Google Scholar] [CrossRef]
- Micalizzi, E.W.; Golshani, A.; Smith, M.L. Propionic acid disrupts endocytosis, cell cycle, and cellular respiration in yeast. BMC Res. Notes 2021, 14, 335. [Google Scholar] [CrossRef]
- McMahon, H.T.; Boucrot, E. Molecular mechanism and physiological functions of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2011, 12, 517–533. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.C.; Wang, Q.; Khan, I.A.; Li, Y.; Wang, J.; Wang, J.Y.; Liu, X.H.; Lin, F.C.; Lu, J.P. The methylcitrate cycle and its crosstalk with the glyoxylate cycle and tricarboxylic acid cycle in pathogenic fungi. Molecules 2023, 28, 6667. [Google Scholar] [CrossRef]
- Lorenz, M.C.; Fink, G.R. The glyoxylate cycle is required for fungal virulence. Nature 2001, 412, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Brock, M.; Fischer, R.; Linder, D.; Buckel, W.; Buckel, W. Methylcitrate synthase from Aspergillus nidulans: Implications for propionate as an antifungal agent. Mol. Microbiol. 2000, 35, 961–973. [Google Scholar] [CrossRef]
- Wang, J.J.; Cai, Q.; Qiu, L.; Ying, S.H.; Feng, M.G. Additive roles of two TPS genes in trehalose synthesis, conidiation, multiple stress responses and host infection of a fungal insect pathogen. Appl. Microbiol. Biotechnol. 2017, 101, 3637–3651. [Google Scholar] [CrossRef]
- Sagini, J.P.N.; Ligabue Braun, R. Fungal heat shock proteins: Molecular phylogenetic insights into the host takeover. Sci. Nat. 2024, 111, 16. [Google Scholar] [CrossRef] [PubMed]
- Bui, D.C.; Lee, Y.; Lim, J.Y.; Fu, M.; Kim, J.C.; Choi, G.J.; Son, H.; Lee, Y.W. Heat shock protein 90 is required for sexual and asexual development, virulence, and heat shock response in Fusarium graminearum. Sci. Rep. 2016, 6, 28154. [Google Scholar] [CrossRef]
- Seymour, I.J.; Piper, P.W. Stress induction of HSP30, the plasma membrane heat shock protein gene of Saccharomyces cerevisiae, appears not to use known stress-regulated transcription factors. Microbiology 1999, 145 Pt 1, 231–239. [Google Scholar] [CrossRef]
- Harada, Y.; Garenáux, E.; Nagatsuka, T.; Uzawa, H.; Nishida, Y.; Sato, C.; Kitajima, K. Interaction of 70-kDa heat shock protein with glycosaminoglycans and acidic glycopolymers. Biochem. Biophys. Res. Commun. 2014, 453, 229–234. [Google Scholar] [CrossRef]
- Verghese, J.; Abrams, J.; Wang, Y.; Morano, K.A. Biology of the heat shock response and protein chaperones: Budding yeast (Saccharomyces cerevisiae) as a model system. Microbiol. Mol. Biol. Rev. 2012, 76, 115–158. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Hu, Y.; Ying, S.-H.; Feng, M.-G. Roles of six Hsp70 genes in virulence, cell wall integrity, antioxidant activity and multiple stress tolerance of Beauveria bassiana. Fungal Genet. Biol. 2020, 144, 103437. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Masison, D.C. Hsp70-nucleotide exchange factor (NEF) Fes1 has non-NEF roles in degradation of gluconeogenic enzymes and cell wall integrity. PLoS Genet. 2019, 15, e1008219. [Google Scholar] [CrossRef]
- Leach, M.D.; Budge, S.; Walker, L.; Munro, C.; Cowen, L.E.; Brown, A.J.P. Hsp90 orchestrates transcriptional regulation by Hsf1 and cell wall remodelling by MAPK signalling during thermal adaptation in a pathogenic yeast. PLoS Path. 2012, 8, e1003069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.B.; Feng, M.G. Antioxidant enzymes and their contributions to biological control potential of fungal insect pathogens. Appl. Microbiol. Biotechnol. 2018, 102, 4995–5004. [Google Scholar] [CrossRef]
- Zhang, L.B.; Tang, L.; Ying, S.H.; Feng, M.G. Subcellular localization of six thioredoxins and their antioxidant activity and contributions to biological control potential in Beauveria bassiana. Fungal Genet. Biol. 2015, 76, 1–9. [Google Scholar] [CrossRef]
- Zhang, L.B.; Tang, L.; Ying, S.H.; Feng, M.G. Regulative roles of glutathione reductase and four glutaredoxins in glutathione redox, antioxidant activity, and iron homeostasis of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2016, 100, 5907–5917. [Google Scholar] [CrossRef] [PubMed]
- Tudzynski, P.; Heller, J.; Siegmund, U. Reactive oxygen species generation in fungal development and pathogenesis. Curr. Opin. Microbiol. 2012, 15, 653–659. [Google Scholar] [CrossRef]
- Slade, D.; Radman, M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef]
- Wang, J.; Ma, Y.; Liu, Y.; Tong, S.; Zhu, S.; Jin, D.; Pei, Y.; Fan, Y. A polyketide synthase, BbpksP, contributes to conidial cell wall structure and UV tolerance in Beauveria bassiana. J. Invertebr. Pathol. 2020, 169, 107280. [Google Scholar] [CrossRef]
- Chen, J.F.; Tan, J.J.; Wang, J.Y.; Mao, A.J.; Xu, X.P.; Zhang, Y.; Zheng, X.L.; Liu, Y.; Jin, D.; Li, X.B.; et al. The zinc finger transcription factor BbCmr1 Regulates conidium maturation in Beauveria bassiana. Microbiol. Spectr. 2022, 10, e0206621. [Google Scholar] [CrossRef]
- Wang, J.J.; Qiu, L.; Cai, Q.; Ying, S.H.; Feng, M.G. Three α-1,2-mannosyltransferases contribute differentially to conidiation, cell wall integrity, multistress tolerance and virulence of Beauveria bassiana. Fungal Genet. Biol. 2014, 70, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Y.; Tong, S.M.; Guan, Y.; Ying, S.H.; Feng, M.G. The velvet protein VeA functions in asexual cycle, stress tolerance and transcriptional regulation of Beauveria bassiana. Fungal Genet. Biol. 2019, 127, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.J.; Mou, Y.N.; Tong, S.M.; Ying, S.H.; Feng, M.G. Subtilisin-like Pr1 proteases marking the evolution of pathogenicity in a wide-spectrum insect-pathogenic fungus. Virulence 2020, 11, 365–380. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, H.; Luo, Y.; Li, Q.; Guo, J.; Wang, B. UV-Induced Mutants of Metarhizium anisopliae: Improved Biological Parameters, Resistance to Stressful Factors, and Comparative Transcriptomic Analysis. J. Fungi 2025, 11, 412. https://doi.org/10.3390/jof11060412
Gao H, Luo Y, Li Q, Guo J, Wang B. UV-Induced Mutants of Metarhizium anisopliae: Improved Biological Parameters, Resistance to Stressful Factors, and Comparative Transcriptomic Analysis. Journal of Fungi. 2025; 11(6):412. https://doi.org/10.3390/jof11060412
Chicago/Turabian StyleGao, Hao, Yingjie Luo, Qiming Li, Jiaxuan Guo, and Bin Wang. 2025. "UV-Induced Mutants of Metarhizium anisopliae: Improved Biological Parameters, Resistance to Stressful Factors, and Comparative Transcriptomic Analysis" Journal of Fungi 11, no. 6: 412. https://doi.org/10.3390/jof11060412
APA StyleGao, H., Luo, Y., Li, Q., Guo, J., & Wang, B. (2025). UV-Induced Mutants of Metarhizium anisopliae: Improved Biological Parameters, Resistance to Stressful Factors, and Comparative Transcriptomic Analysis. Journal of Fungi, 11(6), 412. https://doi.org/10.3390/jof11060412