
Novel Epidithiodiketopiperazine Derivatives in the Mutants of the Filamentous Fungus Trichoderma hypoxylon

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain, Media, and Culture Conditions
2.2. Large-Scale Fermentation, Extraction, and Isolation of Differential Compounds
2.3. HPLC and LC-MS Analysis
2.4. NMR Analysis
3. Results
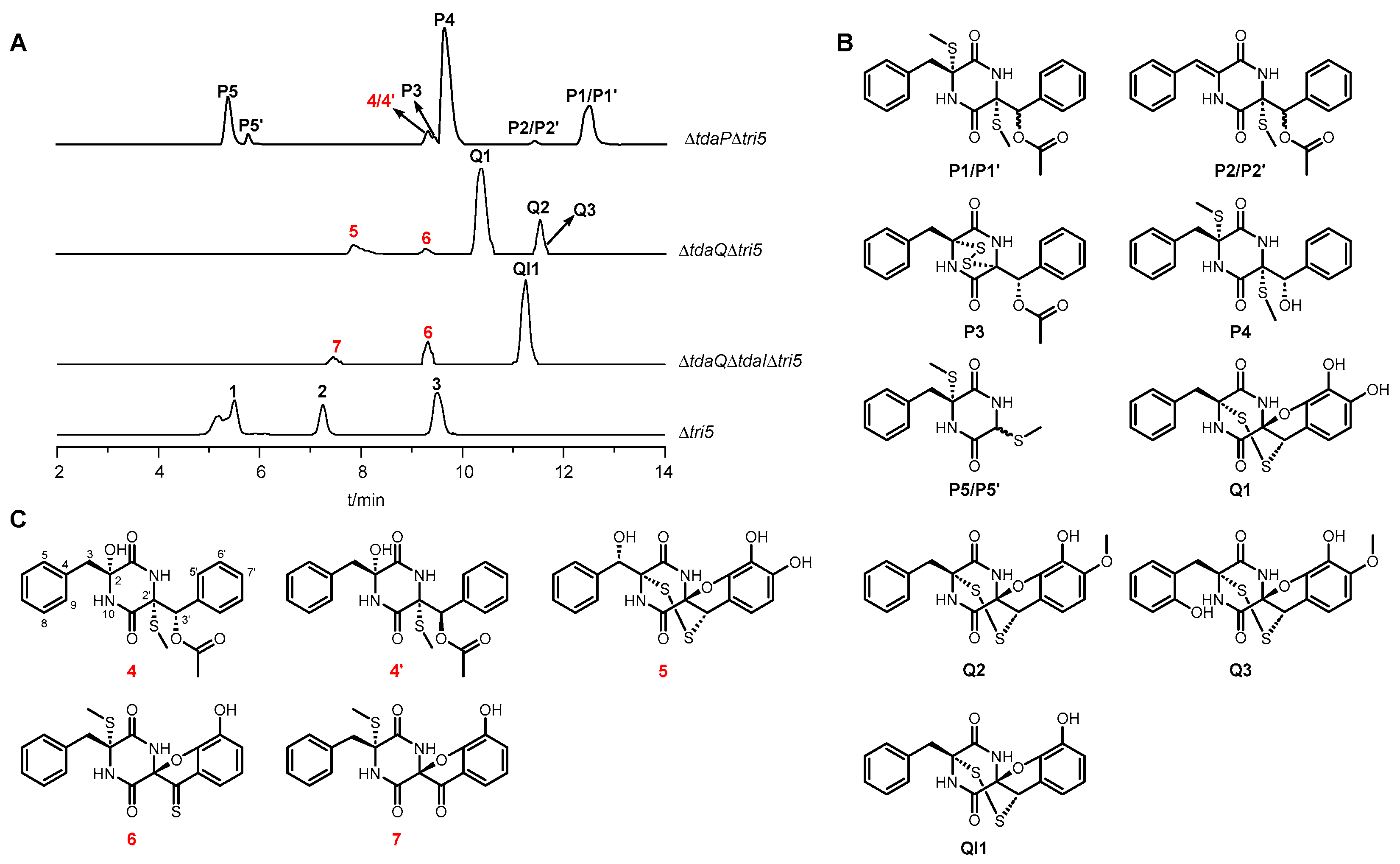
3.1. Analysis of Differential Compounds in the Gene Deletion Mutants of T. hypoxylon
3.2. Characterization of Five Novel ETP Derivatives from Gene Deletion Mutants of T. hypoxylon
3.3. Plausible Biosynthetic Pathway of Compounds 4–7 and 4′
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Orlova, T.I.; Bulgakova, V.G.; Polin, A.N. Secondary metabolites from marine microorganisms. II. Marine fungi and their habitats. Antibiot. Khimioter. 2016, 61, 52–63. [Google Scholar] [PubMed]
- Fan, J.; Wei, P.-L.; Yin, W.-B. Formation of bridged disulfide in epidithiodioxopiperazines. ChemBioChem 2024, 25, e202300770. [Google Scholar] [CrossRef]
- Huber, E.M. Epipolythiodioxopiperazine-based natural products: Building blocks, biosynthesis and biological activities. ChemBioChem 2022, 23, e202200341. [Google Scholar] [CrossRef]
- Dunbar, K.L.; Scharf, D.H.; Litomska, A.; Hertweck, C. Enzymatic carbon–sulfur bond formation in natural product biosynthesis. Chem. Rev. 2017, 117, 5521–5577. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhang, X.; Huang, X.; Wang, H.; Anjum, K.; Gu, Q.; Zhu, T.; Zhang, G.; Li, D. Irregularly bridged epipolythiodioxopiperazines and related analogues: Sources, structures, and biological activities. J. Nat. Prod. 2020, 83, 2045–2053. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, Q.; Chen, S.; Wang, S.; Lu, J.; Gao, X.; Zhang, D.-D.; Jin, X. Natural epidithiodiketopiperazine alkaloids as potential anticancer agents: Recent mechanisms of action, structural modification, and synthetic strategies. Bioorg. Chem. 2023, 137, 106642. [Google Scholar] [CrossRef]
- Esteban, P.; Redrado, S.; Comas, L.; Domingo, M.P.; Millán-Lou, M.I.; Seral, C.; Algarate, S.; Lopez, C.; Rezusta, A.; Pardo, J.; et al. In vitro and in vivo antibacterial activity of gliotoxin alone and in combination with antibiotics against Staphylococcus aureus. Toxins 2021, 13, 85. [Google Scholar] [CrossRef]
- Li, R.; Zhou, Y.; Zhang, X.; Yang, L.; Liu, J.; Wightman, S.M.; Lv, L.; Liu, Z.; Wang, C.Y.; Zhao, C. Identification of marine natural product Pretrichodermamide B as a STAT3 inhibitor for efficient anticancer therapy. Mar. Life Sci. Technol. 2023, 5, 94–101. [Google Scholar] [CrossRef]
- Harwoko, H.; Daletos, G.; Stuhldreier, F.; Lee, J.; Wesselborg, S.; Feldbrugge, M.; Muller, W.E.G.; Kalscheuer, R.; Ancheeva, E.; Proksch, P. Dithiodiketopiperazine derivatives from endophytic fungi Trichoderma harzianum and Epicoccum nigrum. Nat. Prod. Res. 2021, 35, 257–265. [Google Scholar] [CrossRef]
- Fox, E.M.; Howlett, B.J. Biosynthetic gene clusters for epipolythiodioxopiperazines in filamentous fungi. Mycol. Res. 2008, 112, 162–169. [Google Scholar] [CrossRef]
- Guo, C.-J.; Yeh, H.-H.; Chiang, Y.-M.; Sanchez, J.F.; Chang, S.-L.; Bruno, K.S.; Wang, C.C.C. Biosynthetic pathway for the epipolythiodioxopiperazine acetylaranotin in Aspergillus terreus revealed by genome-based deletion analysis. J. Am. Chem. Soc. 2013, 135, 7205–7213. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhao, R.; Ge, M.; Sun, Y.; Li, Y.; Shan, L. Gliotoxin, a natural product with ferroptosis inducing properties. Bioorg. Chem. 2023, 133, 106415. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Li, Y.; Xiang, X.; Tang, Z.; Liu, K.; Su, Q.; Zhang, X.; Li, L. Chaetocin: A review of its anticancer potentials and mechanisms. Eur. J. Pharmacol. 2021, 910, 174459. [Google Scholar] [CrossRef] [PubMed]
- Fortune, J.A.; Bingol, E.; Qi, A.; Baker, D.; Ritchie, F.; Karandeni Dewage, C.S.; Fitt, B.D.L.; Huang, Y.J. Leptosphaeria biglobosa inhibits the production of sirodesmin PL by L. maculans. Pest Manag. Sci. 2024, 80, 2416–2425. [Google Scholar] [CrossRef]
- Xu, G.B.; He, G.; Bai, H.H.; Yang, T.; Zhang, G.L.; Wu, L.W.; Li, G.Y. Indole akaloids from Chaetomium globosum. J. Nat. Prod. 2015, 78, 1479–1485. [Google Scholar] [CrossRef]
- Fan, J.; Ran, H.; Wei, P.-L.; Li, Y.; Liu, H.; Li, S.-M.; Hu, Y.; Yin, W.-B. Pretrichodermamide A biosynthesis reveals the hidden diversity of epidithiodiketopiperazines. Angew. Chem. Int. Ed. 2023, 62, e202217212. [Google Scholar] [CrossRef]
- Wei, W.; Jiang, N.; Mei, Y.N.; Chu, Y.L.; Ge, H.M.; Song, Y.C.; Ng, S.W.; Tan, R.X. An antibacterial metabolite from Lasiodiplodia pseudotheobromae F2. Phytochemistry 2014, 100, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mándi, A.; Li, X.-M.; Meng, L.-H.; Kurtán, T.; Wang, B.-G. Peniciadametizine A, a dithiodiketopiperazine with a unique spiro [furan-2, 7′-pyrazino [1, 2-b][1, 2] oxazine] skeleton, and a related analogue, peniciadametizine B, from the marine sponge-derived fungus Penicillium adametzioides. Mar. Drugs 2015, 13, 3640–3652. [Google Scholar] [CrossRef]
- Zhai, Y.-J.; Huo, G.-M.; Zhang, Q.; Li, D.; Wang, D.-C.; Qi, J.-Z.; Han, W.-B.; Gao, J.-M. Phaeosphaones: Tyrosinase inhibitory thiodiketopiperazines from an endophytic Phaeosphaeria fuckelii. J. Nat. Prod. 2020, 83, 1592–1597. [Google Scholar] [CrossRef]
- Yamazaki, H.; Takahashi, O.; Kirikoshi, R.; Yagi, A.; Ogasawara, T.; Bunya, Y.; Rotinsulu, H.; Uchida, R.; Namikoshi, M. Epipolythiodiketopiperazine and trichothecene derivatives from the NaI-containing fermentation of marine-derived Trichoderma cf. brevicompactum. J. Antibiot. 2020, 73, 559–567. [Google Scholar] [CrossRef]
- Howell, C.R.; Stipanovic, R.D. Gliovirin, a new antibiotic from Gliocladium virens, and its role in the biological control of Pythium ultimum. Can. J. Microbiol. 1983, 29, 321–324. [Google Scholar] [CrossRef]
- Khan, R.A.A.; Najeeb, S.; Hussain, S.; Xie, B.; Li, Y. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic fungi. Microorganisms 2020, 8, 817. [Google Scholar] [CrossRef]
- Khan, R.A.A.; Najeeb, S.; Mao, Z.; Ling, J.; Yang, Y.; Li, Y.; Xie, B. Bioactive secondary metabolites from Trichoderma spp. against phytopathogenic bacteria and root-knot nematode. Microorganisms 2020, 8, 401. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef]
- Sun, J.; Pei, Y.; Li, E.; Li, W.; Hyde, K.D.; Yin, W.-B.; Liu, X. A new species of Trichoderma hypoxylon harbours abundant secondary metabolites. Sci. Rep. 2016, 6, 37369. [Google Scholar] [CrossRef]
- Liu, H.; Fan, J.; Zhang, P.; Hu, Y.; Liu, X.; Li, S.-M.; Yin, W.-B. New insights into the disulfide bond formation enzymes in epidithiodiketopiperazine alkaloids. Chem. Sci. 2021, 12, 4132–4138. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Ran, H.; Wei, P.-L.; Li, Y.; Liu, H.; Li, S.-M.; Yin, W.-B. An ortho-quinone methide mediates disulfide migration in the biosynthesis of epidithiodiketopiperazines. Angew. Chem. Int. Ed. 2023, 62, e202304252. [Google Scholar] [CrossRef]
- Liu, H.; Pu, Y.-H.; Ren, J.-W.; Li, E.-W.; Guo, L.-X.; Yin, W.-B. Genetic dereplication driven discovery of a tricinoloniol acid biosynthetic pathway in Trichoderma hypoxylon. Org. Biomol. Chem. 2020, 18, 5344–5348. [Google Scholar] [CrossRef]
- Scharf, D.H.; Chankhamjon, P.; Scherlach, K.; Heinekamp, T.; Roth, M.; Brakhage, A.A.; Hertweck, C. Epidithiol formation by an unprecedented twin carbon-sulfur lyase in the gliotoxin pathway. Angew. Chem. Int. Ed. 2012, 51, 10064–10068. [Google Scholar] [CrossRef]
- Scharf, D.H.; Chankhamjon, P.; Scherlach, K.; Heinekamp, T.; Willing, K.; Brakhage, A.A.; Hertweck, C. Epidithiodiketopiperazine biosynthesis: A four-enzyme cascade converts glutathione conjugates into transannular disulfide bridges. Angew. Chem. Int. Ed. 2013, 52, 11092–11095. [Google Scholar] [CrossRef]
- Scharf, D.H.; Remme, N.; Habel, A.; Chankhamjon, P.; Scherlach, K.; Heinekamp, T.; Hortschansky, P.; Brakhage, A.A.; Hertweck, C. A dedicated glutathione S-transferase mediates carbon-sulfur bond formation in gliotoxin biosynthesis. J. Am. Chem. Soc. 2011, 133, 12322–12325. [Google Scholar] [CrossRef] [PubMed]
- Scherlach, K.; Kuttenlochner, W.; Scharf, D.H.; Brakhage, A.A.; Hertweck, C.; Groll, M.; Huber, E.M. Structural and mechanistic insights into C-S bond formation in gliotoxin. Angew. Chem. Int. Ed. 2021, 60, 14188–14194. [Google Scholar] [CrossRef]
- Marion, A.; Groll, M.; Scharf, D.H.; Scherlach, K.; Glaser, M.; Sievers, H.; Schuster, M.; Hertweck, C.; Brakhage, A.A.; Antes, I.; et al. Gliotoxin biosynthesis: Structure, mechanism, and metal promiscuity of carboxypeptidase GliJ. ACS Chem. Biol. 2017, 12, 1874–1882. [Google Scholar] [CrossRef]
- Tsunematsu, Y.; Maeda, N.; Sato, M.; Hara, K.; Hashimoto, H.; Watanabe, K.; Hertweck, C. Specialized flavoprotein promotes sulfur migration and spiroaminal formation in aspirochlorine biosynthesis. J. Am. Chem. Soc. 2021, 143, 206–213. [Google Scholar] [CrossRef]
- Tsunematsu, Y.; Maeda, N.; Yokoyama, M.; Chankhamjon, P.; Watanabe, K.; Scherlach, K.; Hertweck, C. Enzymatic amide tailoring expedites unusual Retro-Aldol-type amino acid conversion to form antifungal cyclopeptide. Angew. Chem. Int. Ed. 2018, 57, 14051–14054. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.-J.; Wang, K.-T.; Xu, M.-Y.; Zhang, J.-J.; Lam, C.-K.; Zhong, G.-H.; Xu, J.; Yang, D.-P.; Li, H.-J.; Wang, L.-Y. Secondary metabolites with chemical diversity from the marine-derived fungus Pseudallescheria boydii F19-1 and their cytotoxic activity. RSC Adv. 2016, 6, 76206–76213. [Google Scholar] [CrossRef]
- Song, Z.; Hou, Y.; Yang, Q.; Li, X.; Wu, S. Structures and biological activities of diketopiperazines from marine organisms: A review. Mar. Drugs 2021, 19, 403. [Google Scholar] [CrossRef]
- Sherkhane, P.D.; Bansal, R.; Banerjee, K.; Chatterjee, S.; Oulkar, D.; Jain, P.; Rosenfelder, L.; Elgavish, S.; Horwitz, B.A.; Mukherjee, P.K. Genomics-driven discovery of the gliovirin biosynthesis gene cluster in the plant beneficial fungus Trichoderma virens. ChemistrySelect 2017, 2, 3347–3352. [Google Scholar] [CrossRef]
- Tang, R.; Kimishima, A.; Ishida, R.; Setiawan, A.; Arai, M. Selective cytotoxicity of epidithiodiketopiperazine DC1149B, produced by marine-derived Trichoderma lixii on the cancer cells adapted to glucose starvation. J. Nat. Med. 2020, 74, 153–158. [Google Scholar] [CrossRef]
- Liu, H.; Wang, G.; Li, W.; Liu, X.; Li, E.; Yin, W.-B. A highly efficient genetic system for the identification of a harzianum B biosynthetic gene cluster in Trichoderma hypoxylon. Microbiology 2018, 164, 769–778. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 4 | 4′ | No. | 5 | |||
|---|---|---|---|---|---|---|---|
| δC, Type | δH (J, Hz) | δC, Type | δH (J, Hz) | δC, Type | δH (J, Hz) | ||
| 1 | 167.1, C | - | 167.0, C | - | 1 | 167.3, C | - |
| 2 | 82.6, C | - | 81.5, C | - | 2 | 74.6, C | - |
| 3 | 44.6, CH2 | 3.45 (d, J = 12.6) 2.82 (d, J = 12.6) | 44.1, CH2 | 3.45 (d, J = 12.6) 2.83 (d, J = 12.6) | 3 | 71.6, CH | 5.11 (s) |
| 4 | 135.4, C | - | 135.1, C | - | 4 | 139.6, C | - |
| 5 | 130.8, CH | 7.13–7.35 (m a) | 130.4, CH | 7.13–7.35 (m a) | 5 | 128.9, CH | 7.58–7.62 (m) |
| 6 | 128.3, CH | 7.13–7.35 (m a) | 128.3, CH | 7.13–7.35 (m a) | 6 | 127.3, CH | 7.24–7.34 (m b) |
| 7 | 128.7, CH | 7.13–7.35 (m a) | 128.7, CH | 7.13–7.35 (m a) | 7 | 127.6, CH | 7.24–7.34 (m b) |
| 8 | 128.3, CH | 7.13–7.35 (m a) | 128.3, CH | 7.13–7.35 (m a) | 8 | 127.3, CH | 7.24–7.34 (m b) |
| 9 | 130.8, CH | 7.13–7.35 (m a) | 130.4, CH | 7.13–7.35 (m a) | 9 | 128.9, CH | 7.58–7.62 (m) |
| 1′ | 165.3, C | - | 164.6, C | - | 1′ | 165.4, C | - |
| 2′ | 71.2, C | - | 58.9, C | - | 2′ | 96.3, C | - |
| 3′ | 77.2, CH | 5.82 (s) | 76.6, CH | 5.81 (s) | 3′ | 54.4, CH | 4.60 (d, J = 1.2) |
| 4′ | 134.1, C | - | 134.0, C | - | 4′ | 113.7, C | - |
| 5′ | 128.0, CH | 7.14–7.36 (m a) | 128.3, CH | 7.13–7.35 (m a) | 5′ | 146.3, C | - |
| 6′ | 127.9, CH | 7.14–7.36 (m a) | 127.9, CH | 7.13–7.35 (m a) | 6′ | 129.7, C | - |
| 7′ | 126.7, CH | 7.13–7.35 (m a) | 126.7, CH | 7.13–7.35 (m a) | 7′ | 148.3, C | - |
| 8′ | 127.9, CH | 7.13–7.35 (m a) | 127.9, CH | 7.13–7.35 (m a) | 8′ | 109.0, CH | 6.38 (d, J = 8.0) |
| 9′ | 128.0, CH | 7.13–7.35 (m a) | 128.3, CH | 7.13–7.35 (m a) | 9′ | 113.0, CH | 6.43 (dd, J = 8.0, 1.2) |
| 11′ | 168.7, C | - | 168.7, C | - | 10-NH | - | 9.48 (s) |
| 12′ | 20.5, CH3 | 2.03 (s) | 20.3, CH3 | 1.56 (s) | 10′-NH | - | 9.34 (s) |
| 10-NH | - | 9.14 (s) | - | 8.95 (s) | 3-OH | - | 6.19 (s) |
| 10′-NH | - | 7.81 (s) | - | 7.56 (s) | 6′-OH | - | n.d. |
| 2-OH | - | 6.74 (s) | - | 6.86 (s) | 7′-OH | - | n.d. |
| 2′-SCH3 | 9.6, CH3 | 0.86 (s) | 12.3, CH3 | 2.08 (s) | |||
| No. | 6 | 7 | ||
|---|---|---|---|---|
| δC, Type | δH (J, Hz) | δC, Type | δH (J, Hz) | |
| 1 | 165.8, C | - | 165.5, C | - |
| 2 | 67.6, C | - | 67.7, C | - |
| 3 | 43.9, CH2 | 3.04 (d, J = 13.0) 3.50 (d, J = 13.0) | 43.4, CH2 | 3.03 (d, J = 13.3) 3.50 (d, J = 13.3) |
| 4 | 134.3, C | - | 134.5, C | - |
| 5 | 130.4, CH | 7.23–7.25 (m a) | 130.4, CH | 7.23–7.25 (m a) |
| 6 | 128.3, CH | 7.26–7.34 (m b) | 128.3, CH | 7.26–7.35 (m b) |
| 7 | 127.2, CH | 7.26–7.34 (m b) | 127.3, CH | 7.26–7.35 (m b) |
| 8 | 128.3, CH | 7.26–7.34 (m b) | 128.3, CH | 7.26–7.35 (m b) |
| 9 | 130.4, CH | 7.23–7.25 (m a) | 130.4, CH | 7.23–7.25 (m a) |
| 1′ | 160.2, C | - | 160.4, C | - |
| 2′ | 99.0, C | - | 88.3, C | - |
| 3′ | 227.5, C | - | 194.9, C | - |
| 4′ | 134.0, C | - | 119.9, C | - |
| 5′ | 160.1, C | - | 160.5, C | - |
| 6′ | 143.6, C | - | 143.5, C | - |
| 7′ | 124.7, CH | 7.18 (dd, J = 7.8, 1.2) | 124.4, CH | 7.15 (dd, J = 7.7, 1.2) |
| 8′ | 123.7, CH | 6.92 (t, J = 7.8) | 123.3, CH | 6.96 (t, J = 7.7) |
| 9′ | 114.0, CH | 7.08 (dd, J = 7.8, 1.2) | 114.1, CH | 7.03 (dd, J = 7.7, 1.2) |
| 10-NH | - | 9.54 (s) | - | 9.67 (s) |
| 10′-NH | - | 9.49 (s) | - | 9.41 (s) |
| 2-SCH3 | 13.5, CH3 | 2.30 (s) | 13.0, CH3 | 2.31 (s) |
| 6′-OH | - | 10.12 (s) | - | 10.15 (s) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Z.; Li, Y.; Wei, P.-L.; Zhang, S.; Wang, D.; Fan, J.; Yin, W.-B. Novel Epidithiodiketopiperazine Derivatives in the Mutants of the Filamentous Fungus Trichoderma hypoxylon. J. Fungi 2025, 11, 241. https://doi.org/10.3390/jof11040241
Ren Z, Li Y, Wei P-L, Zhang S, Wang D, Fan J, Yin W-B. Novel Epidithiodiketopiperazine Derivatives in the Mutants of the Filamentous Fungus Trichoderma hypoxylon. Journal of Fungi. 2025; 11(4):241. https://doi.org/10.3390/jof11040241
Chicago/Turabian StyleRen, Zedong, Yuanyuan Li, Peng-Lin Wei, Shengquan Zhang, Dong Wang, Jie Fan, and Wen-Bing Yin. 2025. "Novel Epidithiodiketopiperazine Derivatives in the Mutants of the Filamentous Fungus Trichoderma hypoxylon" Journal of Fungi 11, no. 4: 241. https://doi.org/10.3390/jof11040241
APA StyleRen, Z., Li, Y., Wei, P.-L., Zhang, S., Wang, D., Fan, J., & Yin, W.-B. (2025). Novel Epidithiodiketopiperazine Derivatives in the Mutants of the Filamentous Fungus Trichoderma hypoxylon. Journal of Fungi, 11(4), 241. https://doi.org/10.3390/jof11040241