
Variation of Arbuscular Mycorrhizal Fungi Communities Between Root and Rhizosphere Soil of Endangered Plant Heptacodium miconioides Along Elevation Gradient

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
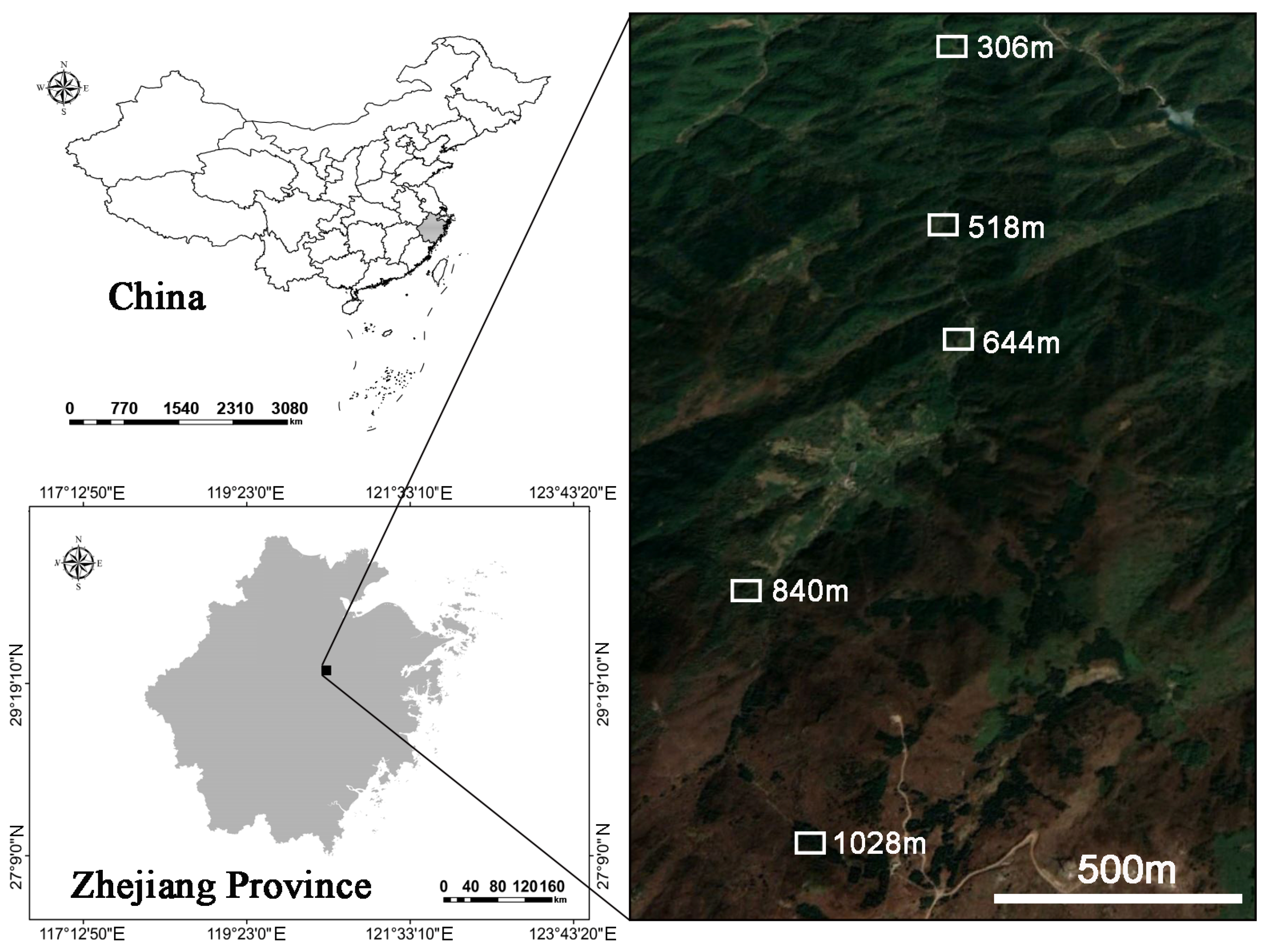
2.1. Study Area
2.2. Root and Rhizosphere Soil Sampling
2.3. Soil Properties and Analysis
2.4. AMF Colonization and Spore Density
2.5. DNA Extraction, MiSeq Sequencing, and Bioinformatic Analysis
2.6. Statistical Analysis
3. Results
3.1. Soil Characteristics Along the Elevation Gradient
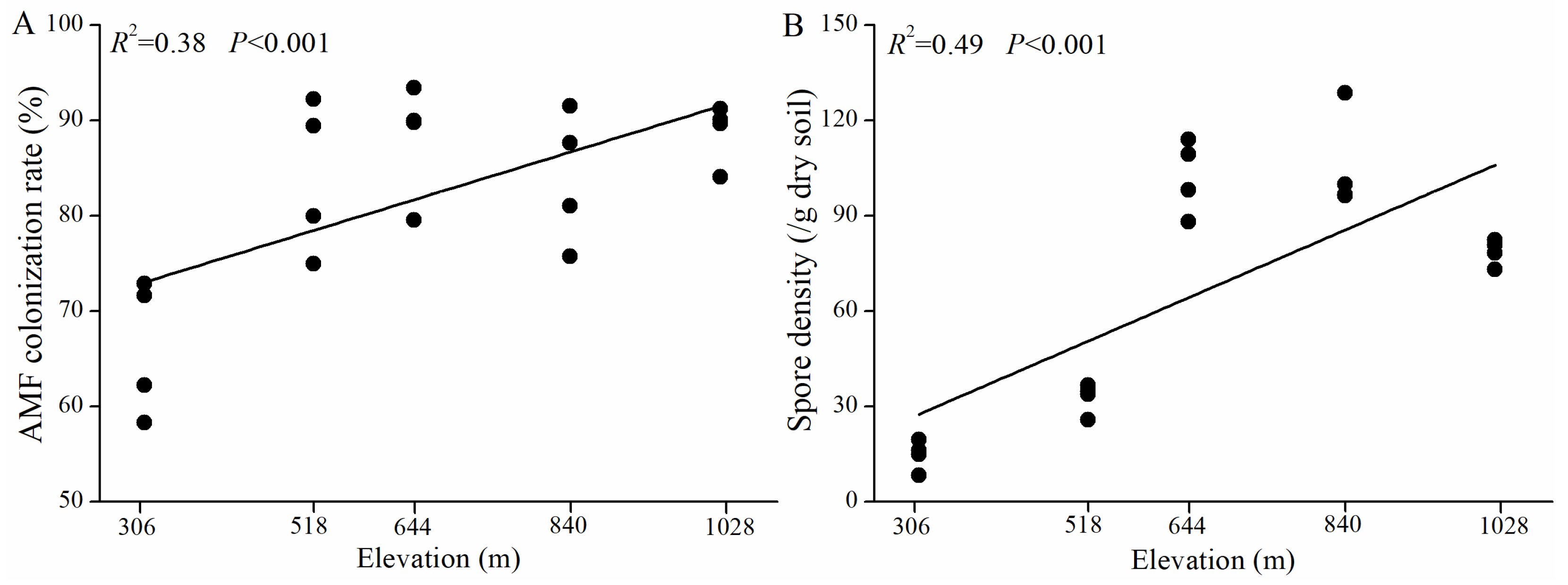
3.2. AMF Colonization and Spore Density Along the Elevation Gradient
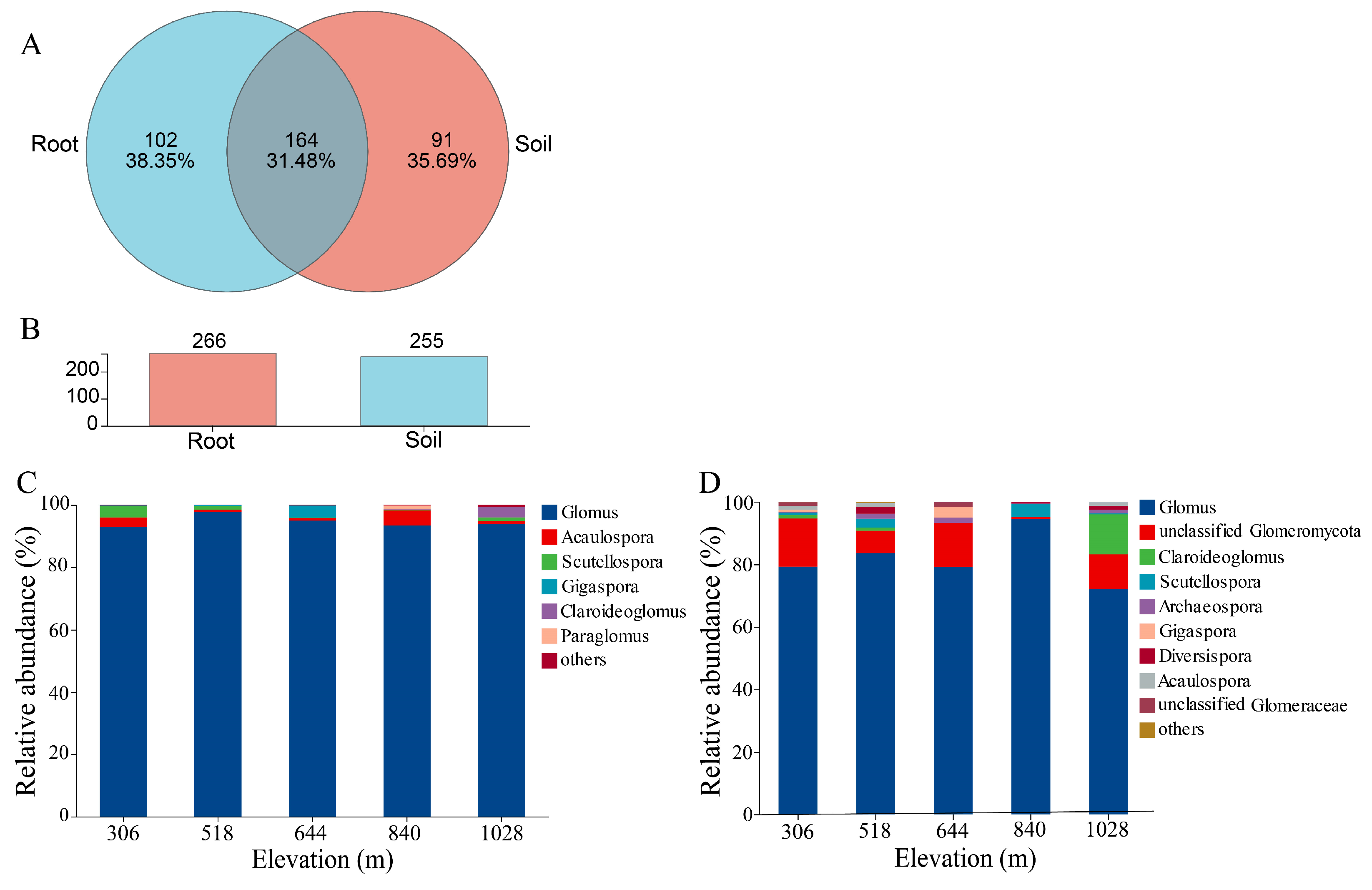
3.3. Overall Sequencing and Taxonomic Assignments in Root and Rhizosphere Soil
3.4. Alpha Diversity of AMF in Root and Rhizosphere Soil
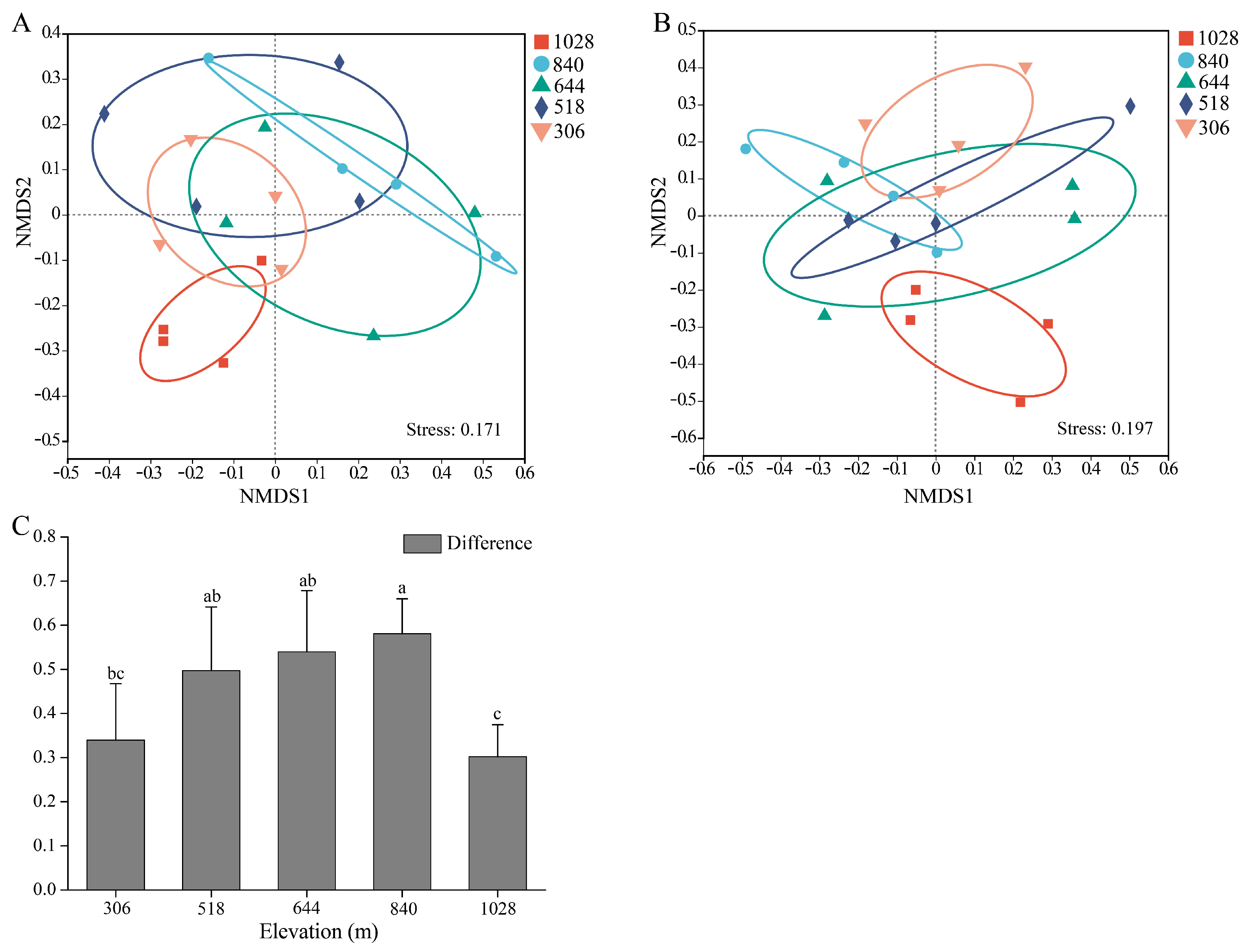
3.5. Beta Diversity of AMF in Root and Rhizosphere Soil
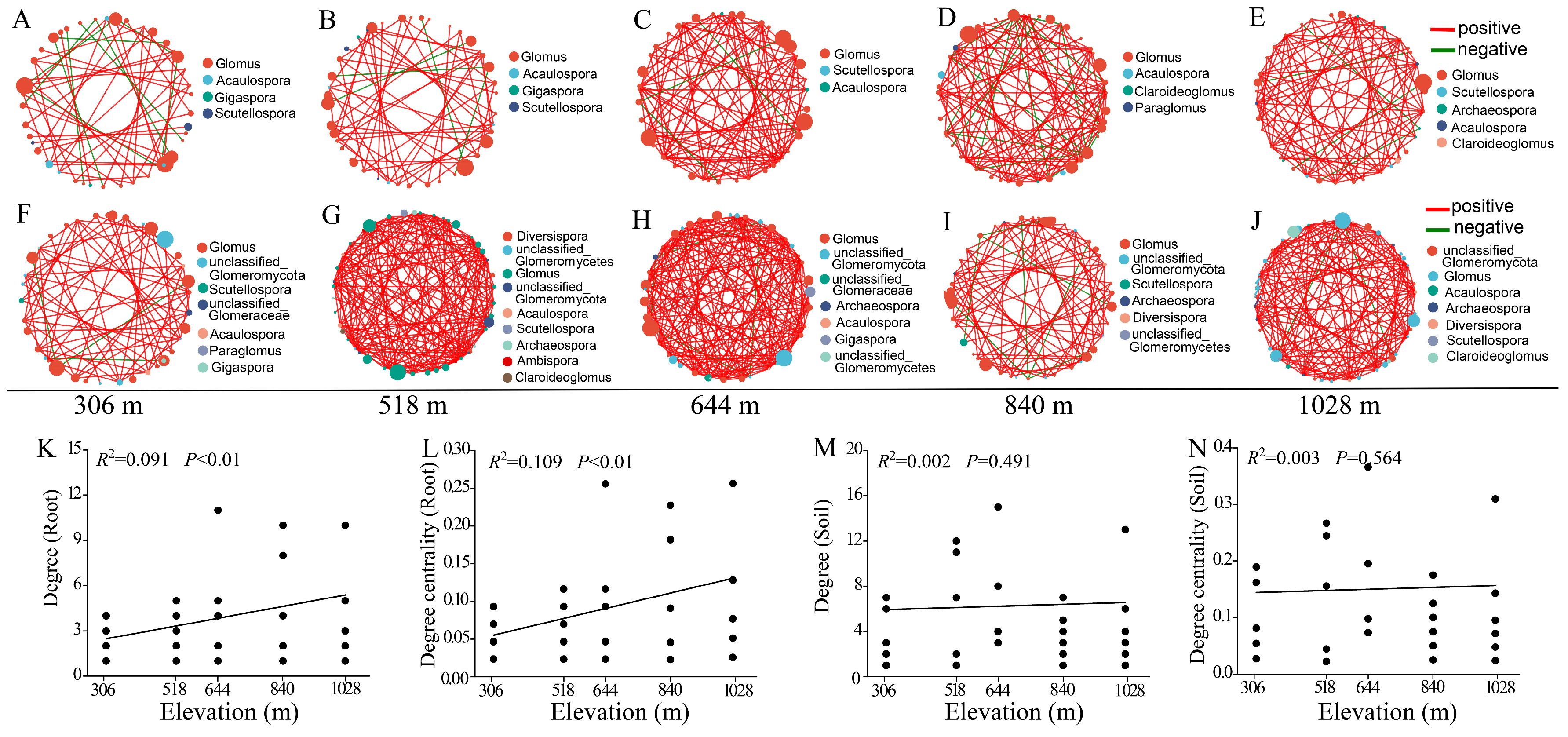
3.6. Co-Occurrence Network Analysis of AMF Community in Root and Rhizosphere Soil
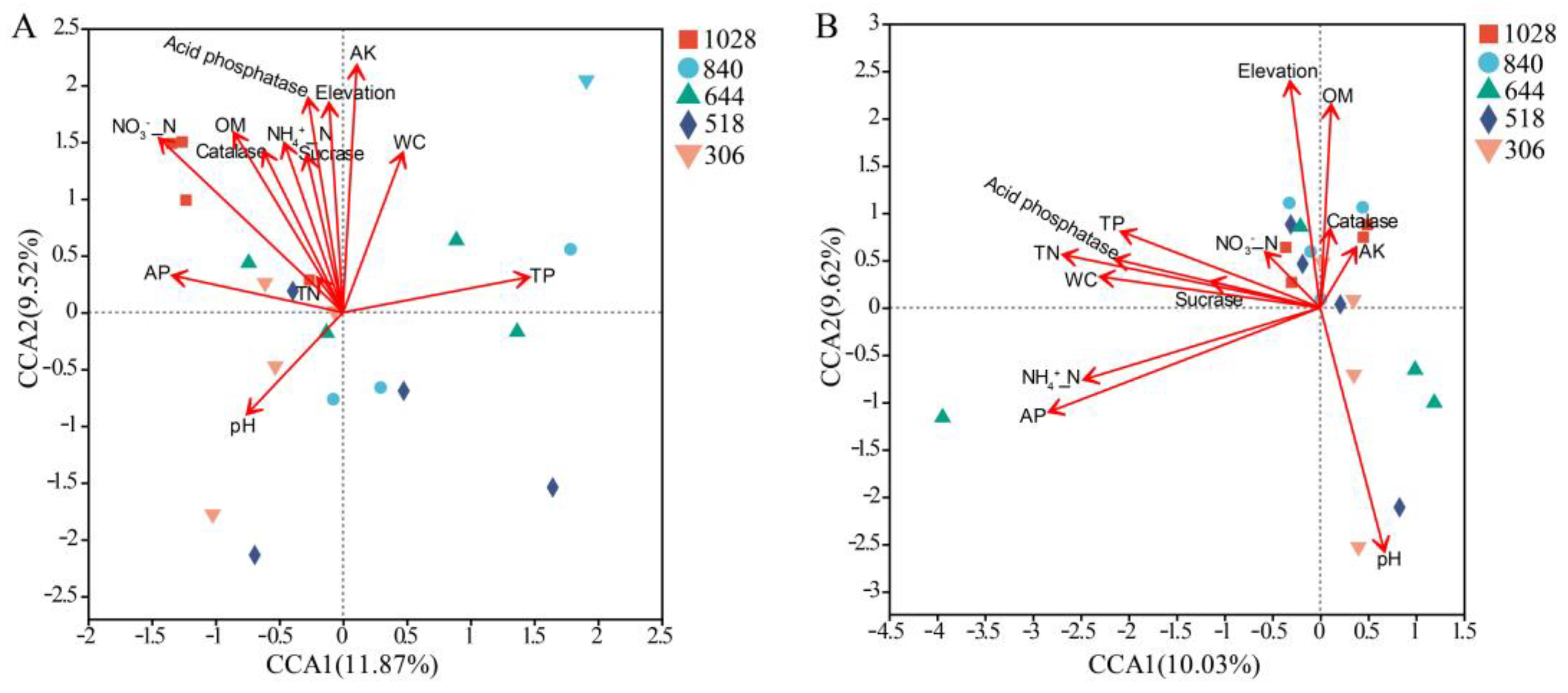
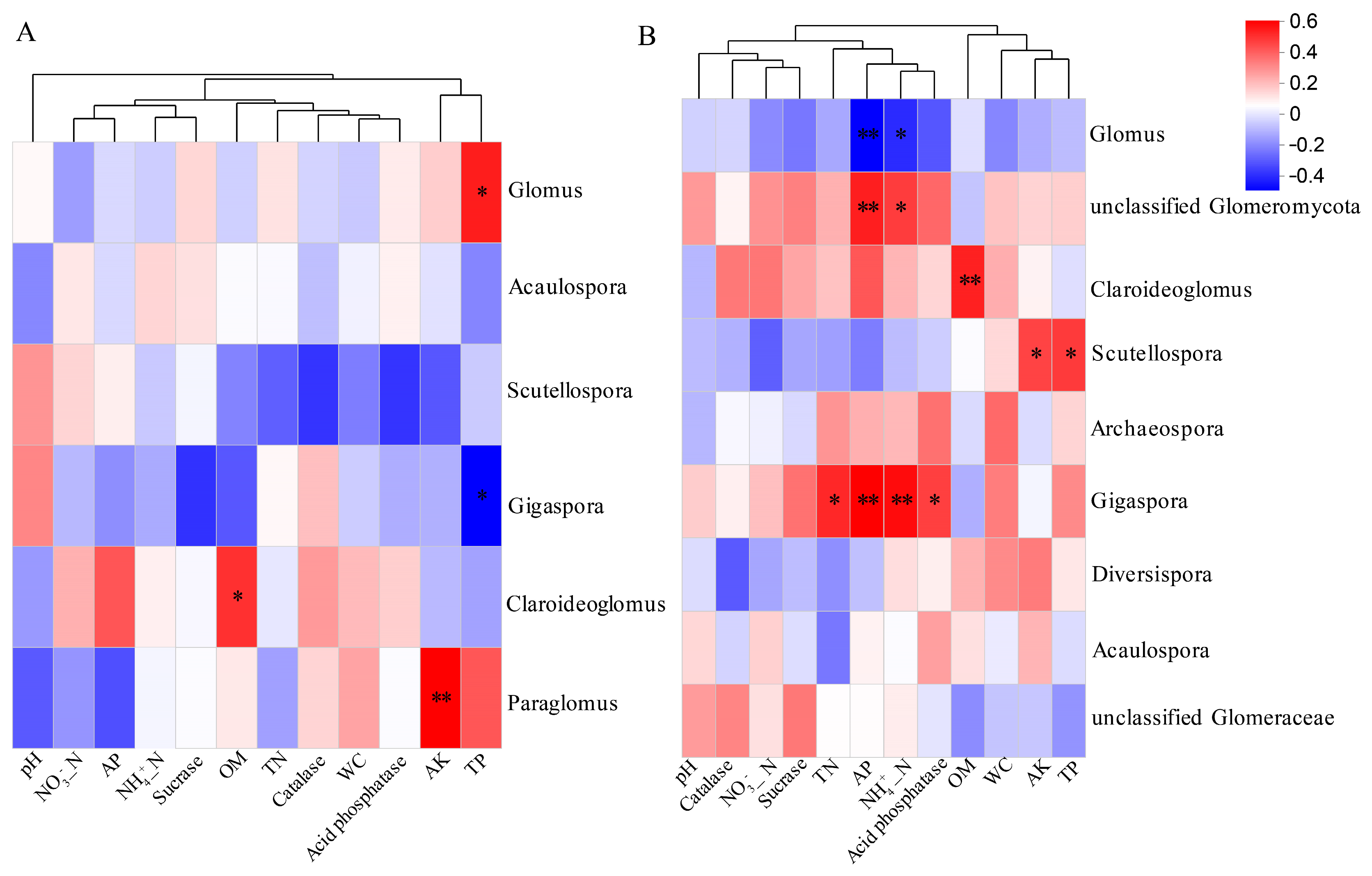
3.7. Relationships Between Soil Characteristics and AMF Community
4. Discussion
4.1. Changes in AMF Colonization and Spore Density of H. miconioides Along Elevation Gradients
4.2. Comparison of AMF Communities in Root and Rhizosphere Soil of H. miconioides Along Elevation Gradients
4.3. Relationship Between Environmental Factors and AMF Communities in Root and Rhizosphere Soil of H. miconioides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boonman, C.C.F.; Serra-Diaz, J.M.; Hoeks, S.; Guo, W.Y.; Enquist, B.J.; Maitner, B.; Malhi, Y.; Merow, C.; Buitenwerf, R.; Svenning, J. More than 17,000 tree species are at risk from rapid global change. Nat. Commun. 2024, 15, 166. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- de Vries, F.; Lau, J.; Hawkes, C.; Semchenko, M. Plant-soil feedback under drought: Does history shape the future? Trends Ecol. Evol. 2023, 8, 708–718. [Google Scholar] [CrossRef]
- Willing, C.E.; Wan, J.; Yeam, J.J.; Cessna, A.M.; Peay, K.G. Arbuscular mycorrhizal fungi equalize differences in plant fitness and facilitate plant species coexistence through niche differentiation. Nat. Ecol. Evol. 2024, 8, 2058–2071. [Google Scholar] [CrossRef]
- Smith, S.; Facelli, E.; Pope, S.; Andrew Smith, F. Plant performance in stressful environments: Interpreting new and established knowledge of the roles of arbuscular mycorrhizas. Plant Soil 2010, 326, 3–20. [Google Scholar] [CrossRef]
- Nuortila, C.; Kytöviita, M.; Tuomi, J. Mycorrhizal symbiosis has contrasting effects on fitness components in Campanula rotundifolia. New Phytol. 2004, 164, 543–553. [Google Scholar] [CrossRef]
- Zubek, S.; Turnau, K.; Tsimilli-Michael, M.; Strasser, R.J. Response of endangered plant species to inoculation with arbuscular mycorrhizal fungi and soil bacteria. Mycorrhiza 2009, 19, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Rigg, J.L.; Offord, C.A.; Singh, B.K.; Anderso, I.; Clarke, S.; Powell, J.R. Soil microbial communities influence seedling growth of a rare conifer independent of plant–soil feedback. Ecology 2016, 97, 3346–3358. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Arunprasad, T. Growth response of Pterocarpus santalinus seedlings to native microbial symbionts (arbuscular mycorrhizal fungi and Rhizobium aegyptiacum) under nursery conditions. J. For. Res. 2021, 32, 225–231. [Google Scholar] [CrossRef]
- Wang, Y.H.; Shao, C.L.; Qiu, Y.J.; Yu, S.Q.; Xia, L.; He, X.B.; Wu, A.P.; Zhang, N.L. Arbuscular mycorrhizal fungi protect a subtropical tree species exposed to simulated acid rain by accelerating photosynthetic ability, antioxidant enzymes and osmolyte accumulation. J. Plant Ecol. 2022, 15, 1036–1048. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Bååth, E.; Reischke, S.; Salinas, N.; Meir, P. Adaptation of soil microbial growth to temperature: Using a tropical elevation gradient to predict future changes. Global Change Biol. 2019, 25, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Gai, J.P.; Tian, H.; Yang, F.Y.; Christie, P.; Li, X.L.; Klironomos, J.N. Arbuscular mycorrhizal fungal diversity along a Tibetan elevation gradient. Pedobiologia 2012, 55, 145–151. [Google Scholar] [CrossRef]
- Liu, L.; Hart, M.M.; Zhang, J.; Cai, X.; Gai, J.; Christie, P.; Li, X.; Klironomos, J.N. Altitudinal distribution patterns of Am fungal assemblages in a Tibetan alpine grassland. FEMS Microbiol. Ecol. 2015, 91, fiv078. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.C.; da Silva, D.K.A.; de Melo, M.A.C.; Escobar, I.E.C.; Oehl, F.; da Silva, G.A. Edaphic factors influence the distribution of arbuscular mycorrhizal fungi along an altitudinal gradient of a Tropical Mountain. Microb. Ecol. 2019, 78, 904–913. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, Z.; Zhou, L.; Huang, K. Effects of Altitude and Continuous Cropping on Arbuscular Mycorrhizal Fungi Community in Siraitia grosvenorii Rhizosphere. Agriculture 2023, 13, 1548. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, Z.; Yang, M.; Lu, S.; Cao, L.; Wang, X. Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi at Different Elevations in Mt. Taibai of Qinling Mountain. Front. Microbiol. 2021, 12, 609386. [Google Scholar] [CrossRef]
- Maherali, H.; Klironomos, J.N. Influence of Phylogeny on Fungal Community Assembly and Ecosystem Functioning. Science 2007, 316, 1746–1748. [Google Scholar] [CrossRef]
- Chagnon, P.L.; Bradley, R.L.; Maherali, H.; Klironomos, J.N. A trait-based framework to understand life history of mycorrhizal fungi. Trends Plant Sci. 2013, 18, 484–491. [Google Scholar] [CrossRef]
- Pascual, J.; Blanco, S.; Ramos, J.L.; van Dillewijn, P. Responses of bulk and rhizosphere soil microbial communities to thermoclimatic changes in a Mediterranean ecosystem. Soil Biol. Biochem. 2018, 118, 130–144. [Google Scholar] [CrossRef]
- Stevens, B.M.; Propster, J.R.; Öpik, M.; Wilson, G.W.; Alloway, S.L.; Mayemba, E.; Johnson, N.C. Arbuscular mycorrhizal fungi in roots and soil respond differently to biotic and abiotic factors in the Serengeti. Mycorrhiza 2020, 30, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Zhang, Y.; Yang, Y.C.; Yang, L.X.; Yang, N.; Zhang, D.P. Long-term effects of mixed planting on arbuscular mycorrhizal fungal communities in the roots and soils of Juglans mandshurica plantations. BMC Microbiol. 2020, 20, 304. [Google Scholar] [CrossRef]
- Mirás-Avalos, J.M.; Antunes, P.M.; Koch, A.; Khosla, K.; Klironomos, J.N.; Dunfield, K.E. The influence of tillage on the structure of rhizosphere and root-associated arbuscular mycorrhizal fungal communities. Pedobiologia 2011, 54, 235–241. [Google Scholar] [CrossRef]
- Hempel, S.; Renker, C.; Buscot, F. Differences in the species composition of arbuscular mycorrhizal fungi in spore, root and soil communities in a grassland ecosystem. Environ. Microbiol. 2007, 9, 1930–1938. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Sun, Z.S.; Jin, Z.X.; Li, J.M. Genome sequencing provides insights into Caprifoliaceae genome evolution and the mechanism underlying second blooming phenomenon in Heptacodium miconioides. Hortic. Plant J. 2024, 10, 897–910. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Chen, C.; Jin, Z.X.; Yang, Z.N.; Li, Y.L. Leaf anatomy, photosynthesis, and chloroplast ultrastructure of Heptacodium miconioides seedlings reveal adaptation to light environment. Environ. Exp. Bot. 2022, 195, 104780. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Mian, S.; Lambkin, D. 1057. Heptacodium miconioides Rehder. Curtis’s Bot. Mag. 2023, 40, 65–73. [Google Scholar] [CrossRef]
- Li, Y.L.; Wang, X.Y.; Chen, X.Y.; Lu, J.Y.; Jin, Z.X.; Li, J.M. Functions of arbuscular mycorrhizal fungi in regulating endangered species Heptacodium miconioides growth and drought stress tolerance. Plant Cell Rep. 2023, 42, 1967–1986. [Google Scholar] [CrossRef]
- Li, Y.L.; Jin, Z.X.; Luo, G.Y.; Chen, C.; Sun, Z.S.; Wang, X.Y. Effects of arbuscular mycorrhizal fungi inoculation on non-structural carbohydrate contents and C:N:P stoichiometry of Heptacodium miconioides under drought stress. Chin. J. Appl. Ecol. 2022, 33, 963–971. [Google Scholar]
- Mebius, L. A rapid method for the determination of organic carbon in soil. Anal. Chim. Acta 1960, 22, 120–124. [Google Scholar] [CrossRef]
- Liu, G.; Liu, R.L.; Zhang, W.G.; Yang, B.; Bi, X.Q.; Li, M.Z.; Chen, X.Y.; Nie, H.; Zhu, Z.H. Arbuscular mycorrhizal colonization rate of an exotic plant, Galinsoga quadriradiata, in mountain ranges changes with altitude. Mycorrhiza 2021, 31, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.; Wang, S.B.; Luo, S.W.; Ren, L.; Liang, Y.Q.; Yang, R.C.; Li, Y.T.; Zhang, Y.Q.; Deng, S.Q.; Zou, L.N. Significant impacts of both total amount and availability of heavy metals on the functions and assembly of soil microbial communities in different land use patterns. Front. Microbiol. 2019, 10, 2293. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Brit. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Brundrett, M.; Melville, L.; Peterson, L. Practical Methods in Mycorrhiza Research: Based on a Workshop Organized in Conjunction with the Ninth North American Conference on Mycorrhizae; University of Guelph, Canada, Mycologue Publication: Guelph, ON, Canada, 1994. [Google Scholar]
- Lumini, E.; Orgiazzi, A.; Borriello, R.; Bonfante, P.; Bianciotto, V. Disclosing arbuscular mycorrhizal fungal biodiversity in soil through a land-use gradient using a Pyrosequencing approach. Environ. Microbiol. 2010, 12, 2165–2179. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microb. 2009, 75, 7537. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Feng, X.X.; Guo, Y.X.; Ren, C.J.; Wang, J.; Doughty, R. Elevation gradients affect the differences of arbuscular mycorrhizal fungi diversity between root and rhizosphere soil. Agric. For. Meteorol. 2020, 284, 107894. [Google Scholar] [CrossRef]
- Bothe, H.; Turnau, K.; Regvar, M. The potential role of arbuscular mycorrhizal fungi in protecting endangered plants and habitats. Mycorrhiza 2010, 20, 445–457. [Google Scholar] [CrossRef]
- Kotilinek, M.; Hiiesalu, I.; Košnar, J.; Šmilauerová, M.; Šmilauer, P.; Altman, J.; Dvorský, M.; Kopecký, M.; Doležal, J. Fungal root symbionts of high-altitude vascular plants in the Himalayas. Sci. Rep. 2017, 7, 6562. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, M.; Shi, Z.; Gao, J.; Wang, X. Biodiversity and Variations of Arbuscular Mycorrhizal Fungi Associated with Roots along Elevations in Mt. Taibai of China. Diversity 2022, 14, 626. [Google Scholar] [CrossRef]
- Wahl, A.L.; Spiegelberger, T. Arbuscular mycorrhizal fungi in changing mountain grassland ecosystems: A challenge for research. Botany 2016, 94, 435–458. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, H.B.; Chai, Y.B.; Xie, H.J.; Mi, N.N.; Li, X.L.; Jin, Z.B.; Gai, J.P. Elevational distribution and occurrence of arbuscular mycorrhizal fungi in non-host Carex capillacea. Mycorrhiza 2021, 31, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, E.S.; Fernandes, G.W.; Berbara, R.L.L.; Valério, H.M.; Goto, B.T. Variation of arbuscular mycorrhizal fungal communities along an altitudinal gradient in rupestrian grasslands in Brazil. Mycorrhiza 2015, 25, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.C.; Lin, W.R.; Lin, L.C.; Wang, P.H. Arbuscular mycorrhizal fungal diversity associated with an endangered species, Chamaecyparis formosensis, in the nature habitat. Trop. Ecol. 2024, 65, 271–277. [Google Scholar] [CrossRef]
- Lisboa, F.J.G.; Chaer, G.; Fernandes, M.F.; Berbara, R.L.L.; Madari, B. The match between microbial community structure and soil properties is modulated by land use types and sample origin within an integrated agroecosystem. Soil Biol. Biochem. 2014, 78, 97–118. [Google Scholar] [CrossRef]
- Coughlan, A.P.; Dalpé, Y.; Lapointe, L.; Piché, Y. Soil pH-induced changes in root colonization, diversity, and reproduction of symbiotic arbuscular mycorrhizal fungi from healthy and declining maple forests. Can. J. Forest Res. 2000, 30, 1543–1554. [Google Scholar] [CrossRef]
- Song, J.; Chen, L.; Chen, F.M.; Ye, J.R. Edaphic and host plant factors are linked to the composition of arbuscular mycorrhizal fungal communities in the root zone of endangered Ulmus chenmoui Cheng in China. Ecol. Evol. 2019, 9, 8900–8910. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Vyas, D.; Jha, A. Soil depth: An overriding factor for distribution of arbuscular mycorrhizal fungi. J. Soil Sci. Plant Nutr. 2013, 13, 23–33. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Vályi, K.; Mardhiah, U.; Rillig, M.C.; Hempel, S. Community assembly and coexistence in communities of arbuscular mycorrhizal fungi. ISME J. 2016, 10, 2341–2351. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.F.; Wang, Q.; Guo, X.Y.; Jiang, X.R.; Cheng, Q.Q.; Fu, L.; Liu, W.; Zhang, L. Local patterns of arbuscular mycorrhizal fungal diversity and community structure in a natural Toona ciliata var. pubescens forest in South Central China. PeerJ 2021, 9, e11331. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yu, X.; Chen, J.; Liu, H.; Zheng, Y.; Qu, H.; Bao, Y. Arbuscular Mycorrhizae Fungi Diversity in the Root–Rhizosphere–Soil of Tetraena mongolica, Sarcozygium xanthoxylon, and Nitraria tangutorum Bobr in Western Ordos, China. Agronomy 2023, 13, 1485. [Google Scholar] [CrossRef]
- Weber, S.E.; Diez, J.M.; Andrews, L.V.; Goulden, M.L.; Aronson, E.L.; Allen, M.F. Responses of arbuscular mycorrhizal fungi to multiple coinciding global change drivers. Fungal Ecol. 2019, 40, 62–71. [Google Scholar] [CrossRef]
- Soteras, F.; Moreira, B.C.; Grilli, G.; Pastor, N.; Mendes, F.C.; Mendes, D.R.; Renison, D.; Kasuya, M.C.M.; de Souza, F.A.; Becerra, A. Arbuscular mycorrhizal fungal diversity in rhizosphere spores versus roots of an endangered endemic tree from Argentina: Is fungal diversity similar among forest disturbance types? Appl. Soil Ecol. 2016, 98, 272–277. [Google Scholar] [CrossRef]
- Bonfim, J.A.; Vasconcellos, R.L.F.; Gumiere, T.; Mescolotti, D.D.L.C.; Oehl, F.; Cardoso, E.J.B.N. Diversity of arbuscular mycorrhizal fungi in a Brazilian atlantic forest toposequence. Microb. Ecol. 2016, 71, 164–177. [Google Scholar] [CrossRef]
- Ma, B.; Wang, H.Z.; Dsouza, M.; Lou, J.; He, Y.; Dai, Z.M.; Brookes, P.C.; Xu, J.M.; Gilbert, J.A. Geographic patterns of co-occurrence network topological features for soil microbiota at continental scale in eastern China. ISME J. 2016, 10, 1891–1901. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef]
- Carvalho, F.; Souza, F.A.; Carrenho, R.; Moreira, F.M.S.; Jesus, E.C.; Fernandes, G.W. The mosaic of habitats in the high-altitude Brazilian rupestrian fields is a hotspot for arbuscular mycorrhizal fungi. Appl. Soil Ecol. 2012, 52, 9–19. [Google Scholar] [CrossRef]
- Camenzind, T.; Hempel, S.; Homeier, J.; Horn, S.; Velescu, A.; Wilcke, W.; Rillig, M.C. Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob. Change Biol. 2014, 20, 3646–3659. [Google Scholar] [CrossRef] [PubMed]
- Van Geel, M.; Jacquemyn, H.; Plue, J.; Saar, L.; Kasari, L.; Peeters, G.; van Acker, K.; Honnay, O.; Ceulemans, T. Abiotic rather than biotic filtering shapes the arbuscular mycorrhizal fungal communities of European seminatural grasslands. New Phytol. 2018, 220, 1262–1272. [Google Scholar] [CrossRef] [PubMed]
- Helgason, T.; Fitter, A.H. Natural selection and the evolutionary ecology of the arbuscular mycorrhizal fungi (phylum Glomeromycota). J. Exp. Bot. 2009, 60, 2465–2480. [Google Scholar] [CrossRef]
- Gai, J.; Liu, R. Effects of soil factors on arbuscular mycorrhizae (AM) fungi around roots of wild plants. Chin. J. Appl. Ecol. 2003, 14, 470. [Google Scholar]
- Johnson, N.C.; Wilson, G.W.T.; Wilson, J.A.; Miller, R.M.; Bowker, M.A. Mycorrhizal phenotypes and the law of the minimum. New Phytol. 2015, 205, 1473–1484. [Google Scholar] [CrossRef]
- Zhao, L.L.; Zhang, K.X.; Sun, X.; He, X.L. Dynamics of arbuscular mycorrhizal fungi and glomalin in the rhizosphere of Gymnocarpos przewalskii in Northwest Desert China. Appl. Soil Ecol. 2022, 170, 104251. [Google Scholar] [CrossRef]
- Ouimet, R.; Camiré, C.; Furlan, V. Effect of soil K, Ca and Mg saturation andendo mycorrhization on growth and nutrient uptake of sugar maple seedlings. Plant Soil 1996, 179, 207–216. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Elevation | Regression | ||||
|---|---|---|---|---|---|---|
| 306 m | 518 m | 644 m | 840 m | 1028 m | R2 | |
| NH4+-N | 49.14 ± 7.80 ab | 31.37 ± 3.19 c | 57.36 ± 15.76 a | 36.36 ± 9.31 bc | 54.92 ± 5.61 a | 0.09 |
| NO3−-N | 38.77 ± 3.42 b | 5.70 ± 2.73 d | 31.34 ± 5.61 bc | 24.37 ± 3.94 c | 51.29 ± 5.41 a | 0.57 ** |
| AK | 0.76 ± 0.19 c | 0.86 ± 0.14 bc | 0.88 ± 0.13 bc | 1.31 ± 0.33 a | 1.24 ± 0.29 ab | 0.31 ** |
| pH | 5.52 ± 0.05 a | 5.19 ± 0.13 b | 5.37 ± 0.10 ab | 4.86 ± 0.10 c | 4.91 ± 0.13 c | 0.63 ** |
| OM | 82.21 ± 12.1 c | 100.22 ± 25.92 c | 89.34 ± 16.24 c | 137.65 ± 7.83 b | 206.55 ± 13.31 a | 0.85 ** |
| WC | 8.96 ± 3.47 c | 17.93 ± 1.82 bc | 28.58 ± 9.07 ab | 24.17 ± 7.88 ab | 33.85 ± 10.34 a | 0.44 ** |
| AP | 11.83 ± 0.37 a | 9.29 ± 0.83 a | 10.85 ± 4.60 a | 9.52 ± 1.80 a | 12.25 ± 2.01 a | 0.02 |
| TN | 1.22 ± 0.07 b | 1.61 ± 0.49 b | 2.52 ± 0.56 a | 1.61 ± 0.57 b | 2.02 ± 0.45 ab | 0.12 |
| TP | 0.25 ± 0.03 b | 0.32 ± 0.02 ab | 0.27 ± 0.09 ab | 0.36 ± 0.07 a | 0.28 ± 0.04 ab | 0.01 |
| Catalase | 14.68 ± 1.96 bc | 14.57 ± 2.77 c | 17.94 ± 1.80 ab | 16.88 ± 0.70 ac | 18.57 ± 1.5 a | 0.27 * |
| Acid phosphatase | 0.03 ± 0.02 b | 0.03 ± 0.02 b | 0.11 ± 0.06 a | 0.08 ± 0.02 ab | 0.13 ± 0.06 a | 0.32 ** |
| Sucrase | 2.64 ± 0.62 a | 1.32 ± 0.31 b | 3.48 ± 0.87 a | 2.41 ± 0.47 ab | 3.15 ± 0.89 a | 0.04 |
| Soil Properties | Rhizosphere Soil | Root | ||
|---|---|---|---|---|
| R2 | p | R2 | p | |
| NH4+-N | 0.020 | 0.424 | 0.123 | 0.138 |
| NO3−-N | 0.056 | 0.306 | 0.147 | 0.058 |
| AK | −0.015 | 0.513 | 0.041 | 0.342 |
| pH | −0.003 | 0.497 | −0.081 | 0.861 |
| OM | 0.114 | 0.164 | 0.138 | 0.092 |
| WC | −0.066 | 0.689 | −0.084 | 0.785 |
| AP | 0.038 | 0.369 | 0.238 | 0.021 |
| TN | 0.233 | 0.029 | 0.177 | 0.036 |
| TP | −0.090 | 0.775 | 0.001 | 0.497 |
| Catalase | 0.164 | 0.072 | 0.050 | 0.296 |
| Acid phosphatase | 0.071 | 0.271 | 0.035 | 0.664 |
| Sucrase | 0.132 | 0.150 | −0.051 | −0.27 |
| Elevation | 0.104 | 0.122 | −0.009 | 0.534 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Luo, G.; Wu, S.; Yang, D.; Lv, P.; Jin, Z.; Li, J. Variation of Arbuscular Mycorrhizal Fungi Communities Between Root and Rhizosphere Soil of Endangered Plant Heptacodium miconioides Along Elevation Gradient. J. Fungi 2025, 11, 222. https://doi.org/10.3390/jof11030222
Li Y, Luo G, Wu S, Yang D, Lv P, Jin Z, Li J. Variation of Arbuscular Mycorrhizal Fungi Communities Between Root and Rhizosphere Soil of Endangered Plant Heptacodium miconioides Along Elevation Gradient. Journal of Fungi. 2025; 11(3):222. https://doi.org/10.3390/jof11030222
Chicago/Turabian StyleLi, Yueling, Guangyu Luo, Shijie Wu, Dang Yang, Pengpeng Lv, Zexin Jin, and Junmin Li. 2025. "Variation of Arbuscular Mycorrhizal Fungi Communities Between Root and Rhizosphere Soil of Endangered Plant Heptacodium miconioides Along Elevation Gradient" Journal of Fungi 11, no. 3: 222. https://doi.org/10.3390/jof11030222
APA StyleLi, Y., Luo, G., Wu, S., Yang, D., Lv, P., Jin, Z., & Li, J. (2025). Variation of Arbuscular Mycorrhizal Fungi Communities Between Root and Rhizosphere Soil of Endangered Plant Heptacodium miconioides Along Elevation Gradient. Journal of Fungi, 11(3), 222. https://doi.org/10.3390/jof11030222