
Optimization of Protoplast Preparation and Establishment of PEG-Mediated Genetic Transformation Method in Cordyceps cicadae

Abstract
1. Introduction
2. Materials and Methods
2.1. C. cicadae Strain and Main Reagents
2.2. Preparation of Protoplasts from C. cicadae
2.3. Optimization of Protoplast Preparation from C. cicadae
2.3.1. Influence of Enzymatic Systems on Protoplast Preparation
2.3.2. Influence of Enzymatic Hydrolysis Temperature on Protoplast Preparation
2.3.3. Influence of Enzymatic Hydrolysis Time on Protoplast Preparation
2.3.4. Influence of Fungal Age on Protoplast Preparation
2.4. Orthogonal Tests for Protoplast Preparation from C. cicadae
2.5. Screening of Regeneration Media for C. cicadae Protoplasts
2.6. PEG-Mediated Protoplast Transformation of pCas9-EGFP
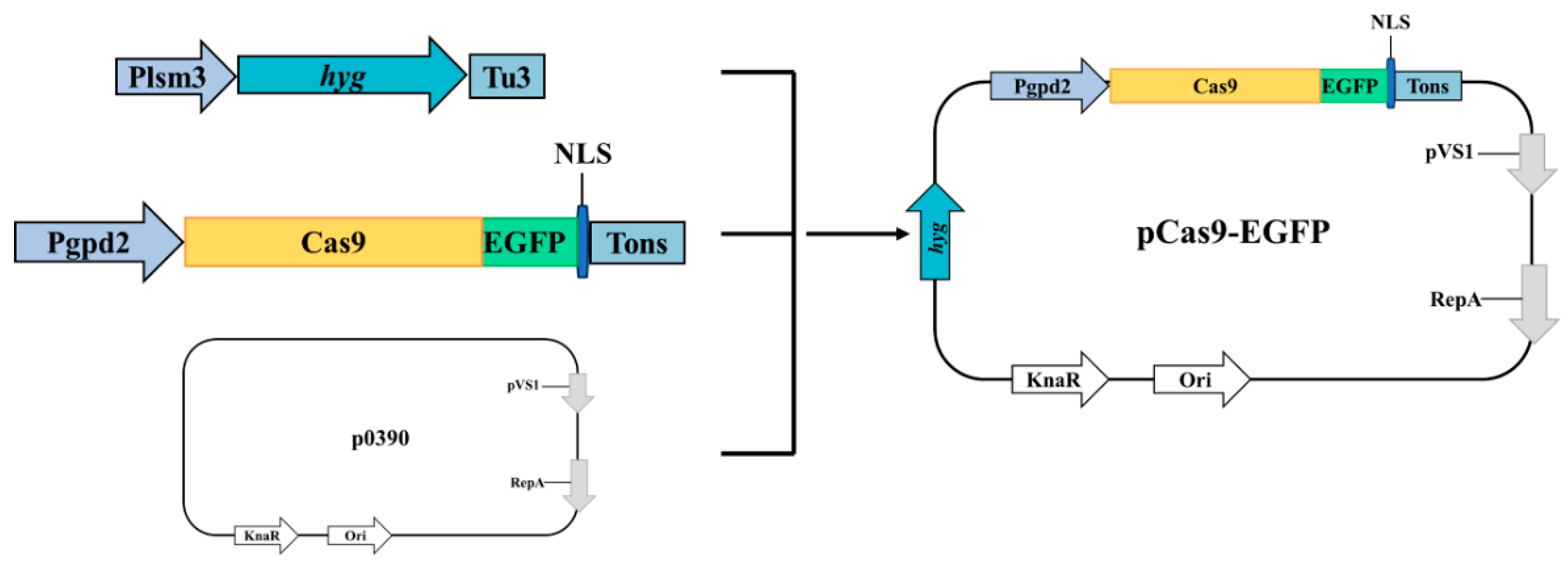
2.6.1. Construction of pCas9-EGFP Plasmid
2.6.2. Screening of Hygromycin B and Geneticin (G418) Concentrations
2.6.3. PEG-Mediated Protoplast Transformation of pCas9-EGFP Plasmid
2.6.4. Microscopic Observation of pCas9-EGFP Plasmid Expression
2.7. PEG-Mediated Protoplast Transformation of the G418 Gene Expression Cassette
2.7.1. Construction of the G418 Gene Expression Cassette
2.7.2. Validation of the G418 Gene Expression Cassette
2.8. Statistical Analysis
3. Results
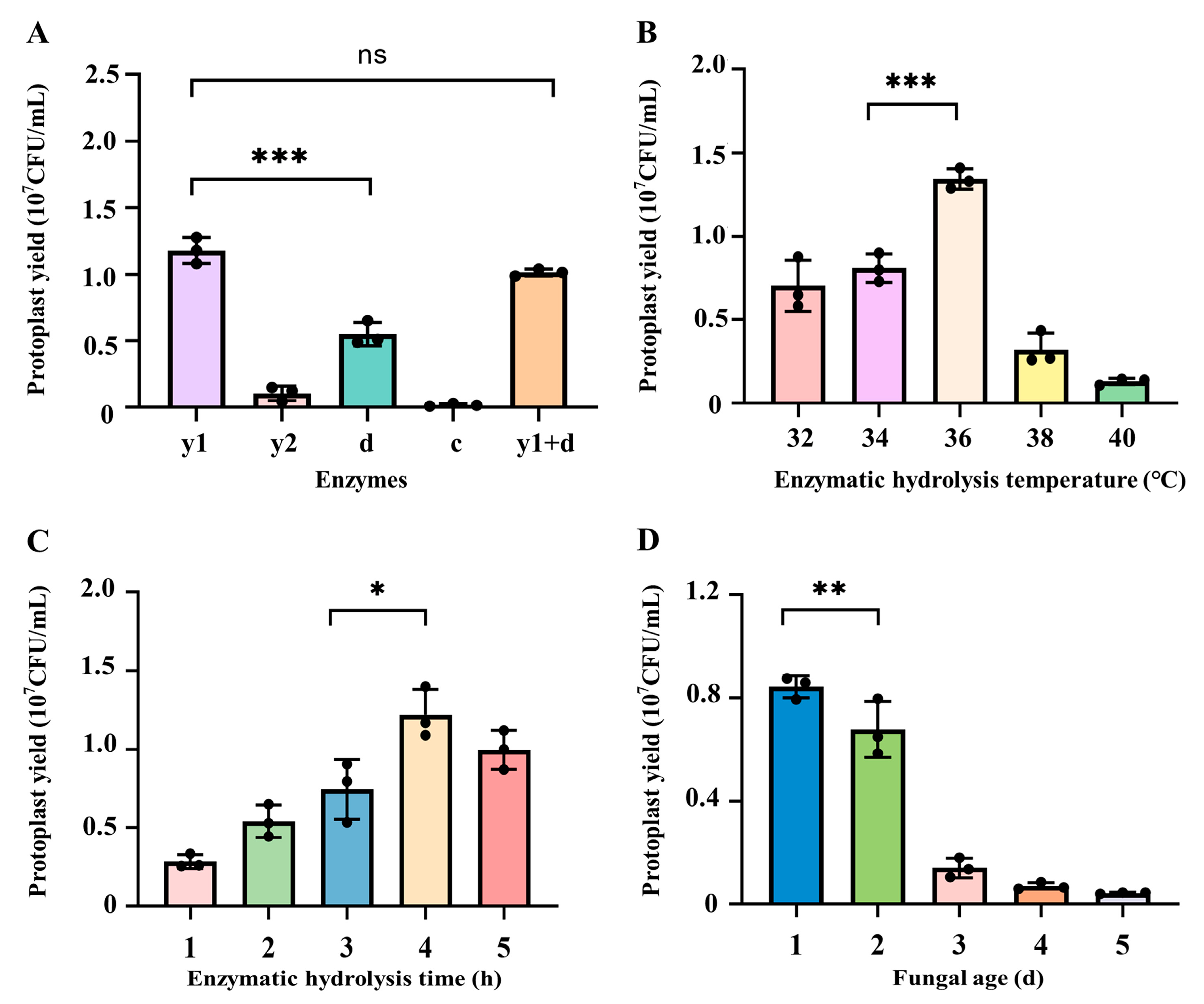
3.1. Influence of Different Factors on Protoplast Preparation from C. cicadae
3.1.1. Influence of Enzymes on the Preparation of Protoplasts from C. cicadae
3.1.2. Influence of Enzymatic Hydrolysis Temperature on the Preparation of Protoplasts from C. cicadae
3.1.3. Influence of Enzymatic Hydrolysis Time on the Preparation of Protoplasts from C. cicadae
3.1.4. Influence of Fungal Age on the Preparation of Protoplasts from C. cicadae
3.2. Optimization of Protoplast Preparation from C. cicadae by Orthogonal Tests
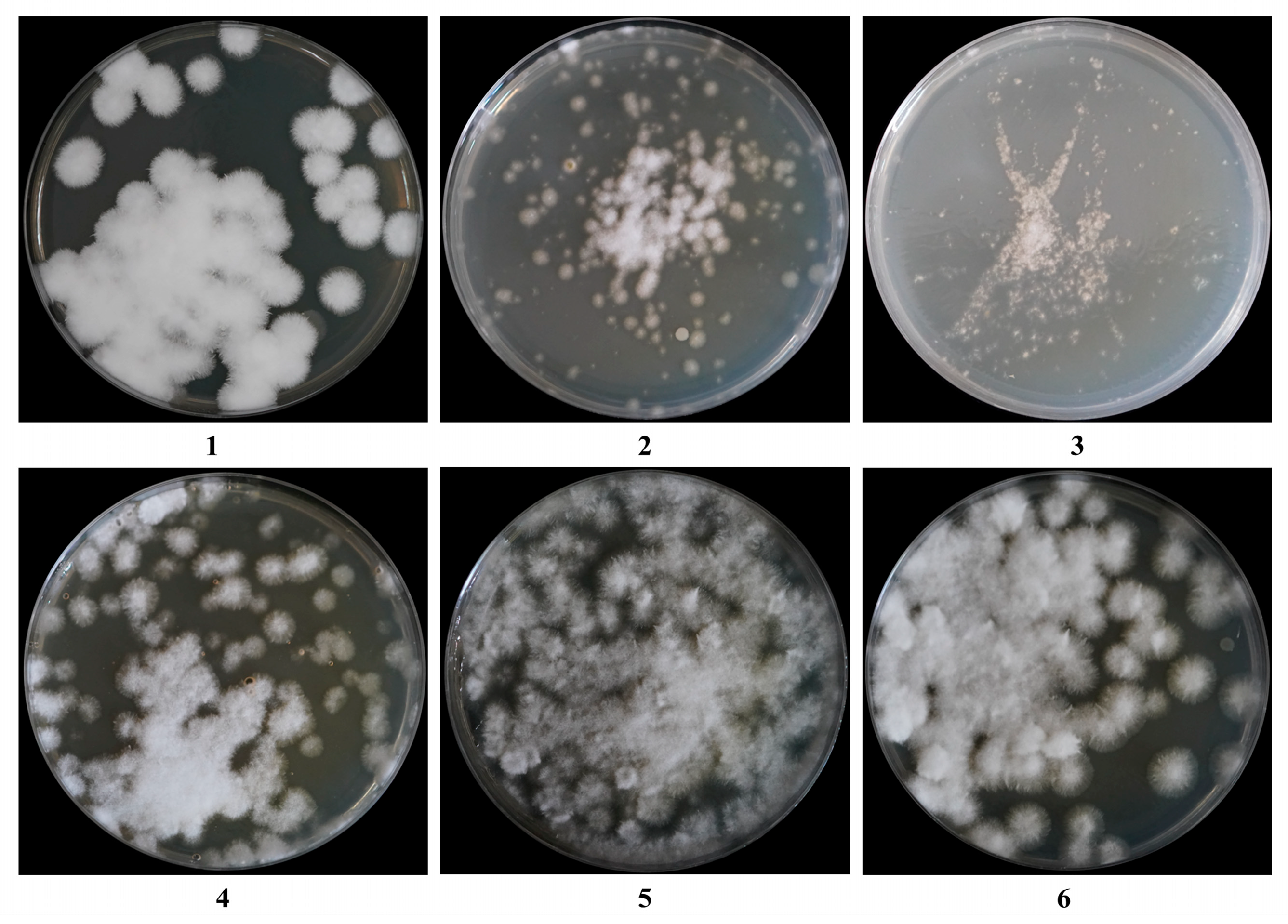
3.3. Influence of Regeneration Medium on the Regeneration of C. cicadae Protoplasts
3.4. Application of PEG-Mediated Protoplasts Transformation Method in the C. cicadae
3.4.1. C. cicadae Sensitivity Tests
3.4.2. Evaluation of PEG-Mediated Transformation of pCas9-EGFP in Protoplasts from C. cicadae
3.4.3. Evaluation of PEG-Mediated Transformation of G418 Expression Cassette in Protoplasts from C. cicadae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, Y.C.; Yang, Z.L. A Revised Checklist of Medicinal Fungi in China. Mycosystema 2008, 27, 801–824. [Google Scholar]
- Qin, D.; Han, C. Medicinal and Edible Fungi as an Alternative Medicine for Treating Age-Related Disease. Evid.-Based Complement. Altern. Med. 2014, 2014, 638561. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Wu, T.; Wu, X.; Zhang, B.; Shen, Y.; Wang, S.; Song, W.; Ruan, H. Analysis of Internal and External Microorganism Community of Wild Cicada Flowers and Identification of the Predominant Cordyceps cicadae Fungus. Front. Microbiol. 2021, 12, 752791. [Google Scholar] [CrossRef]
- Nxumalo, W.; Elateeq, A.A.; Sun, Y. Can Cordyceps cicadae be Used as an Alternative to Cordyceps militaris and Cordyceps sinensis?—A Review. J. Ethnopharmacol. 2020, 257, 112879. [Google Scholar] [CrossRef]
- Wang, Y.; Ni, Z.; Li, J.; Shao, Y.; Yong, Y.; Lv, W.; Zhang, S.; Fu, T.; Chen, A. Cordyceps cicadae Polysaccharides Alleviate Hyperglycemia by Regulating Gut Microbiota and its Mmetabolites in High-Fat Diet/Streptozocin-Induced Diabetic Mice. Front. Nutr. 2023, 10, 1203430. [Google Scholar] [CrossRef]
- Xie, H.; Li, X.; Chen, Y.; Lang, M.; Shen, Z.; Shi, L. Ethanolic Extract of Cordyceps Cicadae Exerts Antitumor Effect On Human Gastric Cancer SGC-7901 Cells by Inducing Apoptosis, Cell Cycle Arrest and Endoplasmic Reticulum Stress. J. Ethnopharmacol. 2019, 231, 230–240. [Google Scholar] [CrossRef]
- Wang, X.; Qin, A.; Xiao, F.; Olatunji, O.J.; Zhang, S.; Pan, D.; Han, W.; Wang, D.; Ni, Y. N(6) -(2-Hydroxyethyl)-Adenosine From Cordyceps cicadae Protects Against Diabetic Kidney Disease Via Alleviation of Oxidative Stress and Inflammation. J. Food Biochem. 2019, 2, e12727. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, X.; Ge, Q.; Li, J.; Wang, D.; Wei, Y.; Ouyang, Z. Antioxidant and Anti-Aging Activities of Polysaccharides From Cordyceps cicadae. Int. J. Biol. Macromol. 2020, 157, 394–400. [Google Scholar] [CrossRef]
- Krenek, P.; Samajova, O.; Luptovciak, I.; Doskocilova, A.; Komis, G.; Samaj, J. Transient Plant Transformation Mediated by Agrobacterium Tumefaciens: Principles, Methods and Applications. Biotechnol. Adv. 2015, 6 Pt 2, 1024–1042. [Google Scholar] [CrossRef]
- Sayari, M.; van der Nest, M.A.; Steenkamp, E.T.; Adegeye, O.O.; Marincowitz, S.; Wingfield, B.D. Agrobacterium-Mediated Transformation of Ceratocystis albifundus. Microbiol. Res. 2019, 226, 55–64. [Google Scholar] [CrossRef]
- Jiang, D.; Zhu, W.; Wang, Y.; Sun, C.; Zhang, K.; Yang, J. Molecular Tools for Functional Genomics in Filamentous Fungi: Recent Advances and New Strategies. Biotechnol. Adv. 2013, 8, 1562–1574. [Google Scholar] [CrossRef]
- Bai, S.; Luo, H.; Tong, H.; Wu, Y.; Yuan, Y. Advances On Transfer and Maintenance of Large DNA in Bacteria, Fungi, and Mammalian Cells. Biotechnol. Adv. 2024, 76, 108421. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.L.; Magana-Ortiz, D.; Gomez-Lim, M.; Fernandez, F.; Loske, A.M. Physical Methods for Genetic Transformation of Fungi and Yeast. Phys. Life Rev. 2014, 2, 184–203. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, R.; Ye, T.; Guan, R.; Xu, L.; Ma, X.; Zhang, J.; Xiao, S.; Yuan, D. Isolation, Purification and PEG-Mediated Transient Expression of Mesophyll Protoplasts in Camellia oleifera. Plant Methods 2022, 1, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, J.; Du, G.; Chen, J.; Takahashi, S.; Liu, S. Developing Aspergillus niger as a Cell Factory for Food Enzyme Production. Biotechnol. Adv. 2020, 44, 107630. [Google Scholar] [CrossRef]
- Rachmawati, R.; Kinoshita, H.; Nihira, T. Establishment of Transformation System in Cordyceps Militaris by Using Integration Vector with Benomyl Resistance Gene. Procedia Environ. Sci. 2013, 17, 142–149. [Google Scholar] [CrossRef]
- Zheng, Z.; Huang, C.; Cao, L.; Xie, C.; Han, R. Agrobacterium Tumefaciens-Mediated Transformation as a Tool for Insertional Mutagenesis in Medicinal Fungus Cordyceps militaris. Fungal Biol. 2011, 3, 265–274. [Google Scholar] [CrossRef]
- Nai, Y.S.; Lee, M.R.; Kim, S.; Lee, S.J.; Kim, J.C.; Yang, Y.T.; Kim, J.S. Relationship Between Expression Level of Hygromycin B-Resistant Gene and Agrobacterium Tumefaciens-Mediated Transformation Efficiency in Beauveria bassiana JEF-007. J. Appl. Microbiol. 2017, 3, 724–731. [Google Scholar] [CrossRef]
- Shin, T.Y.; Lee, M.R.; Lee, S.J.; Kim, J.C.; Kim, S.; Park, S.E.; Kim, W.J.; Kim, J.S. Gene-Disruption of the Entomopathogenic Fungus Beauveria bassiana Incubated with DsRNA. J. Basic Microbiol. 2021, 7, 642–651. [Google Scholar] [CrossRef]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The Role of the Fungal Cell Wall in the Infection of Plants. Trends Microbiol. 2017, 12, 957–967. [Google Scholar] [CrossRef]
- Kemppainen, M.; Chowdhury, J.; Lundberg-Felten, J.; Pardo, A. Fluorescent Protein Expression in the Ectomycorrhizal Fungus Laccaria Bicolor: A Plasmid Toolkit for Easy Use of Fluorescent Markers in Basidiomycetes. Curr. Genet. 2020, 4, 791–811. [Google Scholar] [CrossRef] [PubMed]
- Aloulou-Abdelkefi, M.; Trigui-Lahiani, H.; Gargouri, A. Autoclaved Mycelium Induces Efficiently the Production of Hydrolytic Enzymes for Protoplast Preparation of Autologous Fungus. Appl. Biochem. Microbiol. 2017, 53, 230–236. [Google Scholar] [CrossRef]
- Lou, H.-W.; Ye, Z.-W.; Yu, Y.-H.; Lin, J.-F.; Guo, L.-Q.; Chen, B.-X.; Tang, H.-B.; Wei, T.; Chen, L.-T.; Yun, F. The Efficient Genetic Transformation of Cordyceps militaris by Using Mononuclear Protoplasts. Sci. Hortic. 2019, 243, 307–313. [Google Scholar] [CrossRef]
- Wu, J.; Chou, J. Optimization of Protoplast Preparation and Regeneration of a Medicinal Fungus Antrodia cinnamomea. Mycobiology 2019, 4, 483–493. [Google Scholar] [CrossRef]
- Li, D.; Tang, Y.; Lin, J.; Cai, W. Methods for Genetic Transformation of Filamentous Fungi. Microb. Cell Factories 2017, 16, 168. [Google Scholar] [CrossRef]
- Lou, H.; Ye, Z.; Yun, F.; Lin, J.; Guo, L.; Chen, B.; Mu, Z. Targeted Gene Deletion in Cordyceps militaris Using the Split-Marker Approach. Mol. Biotechnol. 2018, 5, 380–385. [Google Scholar] [CrossRef]
- Meng, G.; Wang, X.; Liu, M.; Wang, F.; Liu, Q.; Dong, C. Efficient CRISPR/Cas9 System Based On Autonomously Replicating Plasmid with an AMA1 Sequence and Precisely Targeted Gene Deletion in the Edible Fungus, Cordyceps militaris. Microb. Biotechnol. 2022, 10, 2594–2606. [Google Scholar] [CrossRef]
- Ning, Y.; Hu, B.; Yu, H.; Liu, X.; Jiao, B.; Lu, X. Optimization of Protoplast Preparation and Establishment of Genetic Transformation System of an Arctic-Derived Fungus Eutypella sp. Front. Microbiol. 2022, 13, 769008. [Google Scholar] [CrossRef]
- Xu, X.; Zhu, H.; Ren, Y.; Feng, C.; Ye, Z.; Cai, H.; Wan, X.; Peng, C. Efficient Isolation and Purification of Tissue-Specific Protoplasts From Tea Plants (Camellia sinensis (L.) O. Kuntze). Plant Methods 2021, 1, 84. [Google Scholar] [CrossRef]
- Chen, B.; Wei, T.; Ye, Z.; Yun, F.; Kang, L.; Tang, H.; Guo, L.; Lin, J. Efficient CRISPR-Cas9 Gene Disruption System in Edible-Medicinal Mushroom Cordyceps militaris. Front. Microbiol. 2018, 9, 1157. [Google Scholar] [CrossRef]
- Lim, F.-H.; Rasid, O.A.; Idris, A.S.; As’wad, A.W.M.; Vadamalai, G.; Parveez, G.K.A.; Wong, M. Enhanced Polyethylene Glycol (PEG)-Mediated Protoplast Transformation System for the Phytopathogenic Fungus, Ganoderma Boninense. Folia Microbiol. 2021, 4, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Flor-Parra, I.; Zhurinsky, J.; Bernal, M.; Gallardo, P.; Daga, R.R. A Lallzyme MMX-Based Rapid Method for Fission Yeast Protoplast Preparation. Yeast 2014, 2, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.H.; Naing, A.H.; Kim, C.K. Protoplast Isolation and Shoot Regeneration From Protoplast-Derived Callus of Petunia hybrida Cv. Mirage Rose. Biology 2020, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Xiao, Y.; Xing, J.; Yu, K.; Li, X.; Chai, Y. Establishment of a Genetic Transformation System for Cordycipitoid Fungus Cordyceps chanhua. Front. Microbiol. 2024, 15, 1333793. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, M.; Xie, C.; Guo, X. Establishment of Genetic Transformation System of Cordyceps Sinensis by PEG Method. China Biotechnol. 2017, 4, 76–82. [Google Scholar]
- Kaida, R.; Satoh, Y.; Bulone, V.; Yamada, Y.; Kaku, T.; Hayashi, T.; Kaneko, T.S. Activation of Beta-Glucan Synthases by Wall-Bound Purple Acid Phosphatase in Tobacco Cells. Plant Physiol. 2009, 4, 1822–1830. [Google Scholar] [CrossRef]
- Jin, L.; Xu, Z.; Men, X.; Zhang, B.; Liu, Z.Q.; Zheng, Y. Enhancement of Protoplast Preparation and Regeneration of Hirsutella sinensis Based on Process Optimization. Biotechnol. Lett. 2020, 11, 2357–2366. [Google Scholar] [CrossRef]
- Rehman, L.; Su, X.; Guo, H.; Qi, X.; Cheng, H. Protoplast Transformation as a Potential Platform for Exploring Gene Function in Verticillium dahliae. BMC Biotechnol. 2016, 1, 57. [Google Scholar] [CrossRef]
- Choi, G.C.; Avila-Peltroche, J.; Won, B.Y.; Cho, T.O. Optimization of Protoplast Isolation and Subsequent Regeneration From the Economically Important Brown Alga Ecklonia Cava Using Response Surface Methodology. Algal Res. 2024, 80, 103525. [Google Scholar] [CrossRef]
- Agrawal, R.; Satlewal, A.; Gaur, R.; Mathur, A.; Kumar, R.; Gupta, R.P.; Tuli, D.K. Pilot Scale Pretreatment of Wheat Straw and Comparative Evaluation of Commercial Enzyme Preparations for Biomass Saccharification and Fermentation. Biochem. Eng. J. 2015, 102, 54–61. [Google Scholar] [CrossRef]
- Hu, Z.; Liu, Q.; Ouyang, B.; Wang, G.; Wei, C.; Zhao, X. Recent Advances in Genetic Engineering to Enhance Plant-Polysaccharide-Degrading Enzyme Expression in Penicillium Oxalicum: A Brief Review. Int. J. Biol. Macromol. 2024, 278 Pt 2, 134775. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qin, Y.; Kong, Y.; Karunarathna, S.C.; Liang, Y.; Xu, J. Optimization of Protoplast Preparation Conditions in Lyophyllum decastes and Transcriptomic Analysis Throughout the Process. J. Fungi 2024, 12, 886. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Govindaraj, L.; Dhanasekaran, M.; Vetrivel, S.; Kumar, K.K.; Ebenezar, E. Combination of Driselase and Lysing Enzyme in One Molar Potassium Chloride is Effective for the Production of Protoplasts from Germinated Conidia of Fusarium Verticillioides. J. Microbiol. Methods 2015, 111, 127–134. [Google Scholar] [CrossRef]
- Zhang, C.; Bozileva, E.; Frits, V.D.K.; Dong, Y.; Sanders, J.P.M.; Bruins, M.E. Integration of Galacturonic Acid Extraction with Alkaline Protein Extraction From Green Tea Leaf Residue. Ind. Crops Prod. 2016, 89, 95–102. [Google Scholar] [CrossRef]
- Cheng-Jin, G.; Run, Z. Study on Protoplast Preparation and Regeneration of Cordyceps sinensis. Food Sci. 2009, 30, 166–170. [Google Scholar]
- Liu, X.; Xia, Y.; Zhang, Y.; Liang, L.; Xiong, Z.; Wang, G.; Song, X.; Ai, L. Enhancement of Antroquinonol Production Via the Overexpression of 4-Hydroxybenzoate Polyprenyltransferase Biosynthesis-Related Genes in Antrodia Cinnamomea. Phytochemistry 2021, 184, 112677. [Google Scholar] [CrossRef]
- Huang, J.; Wang, A.; Huang, C.; Sun, Y.; Song, B.; Zhou, R.; Li, L. Generation of Marker-Free Pbd-2 Knock-in Pigs Using the CRISPR/Cas9 and Cre/Loxp Systems. Genes 2020, 8, 951. [Google Scholar] [CrossRef]
- Li, S.; Liu, Q.; Zhong, Z.; Deng, Z.; Sun, Y. Exploration of Hygromycin B Biosynthesis Utilizing CRISPR-Cas9-Associated Base Editing. Acs. Chem. Biol. 2020, 6, 1417–1423. [Google Scholar] [CrossRef]
- Liu, Z.; Friesen, T.L. Polyethylene Glycol (PEG)-Mediated Transformation in Filamentous Fungal Pathogens. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 365–375. [Google Scholar]
- Wu, F.; Hanzawa, Y. A Simple Method for Isolation of Soybean Protoplasts and Application to Transient Gene Expression Analyses. JOVE-J. Vis. Exp. 2018, 131, 57258. [Google Scholar]
- Yoo, S.; Cho, Y.; Sheen, J. Arabidopsis Mesophyll Protoplasts: A Versatile Cell System for Transient Gene Expression Analysis. Nat. Protoc. 2007, 7, 1565–1572. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Symbol | Levels | ||
|---|---|---|---|---|
| 1 | 2 | 3 | ||
| Fungal Age (d) | A | 1 | 2 | 3 |
| Enzymatic Hydrolysis Time (h) | B | 3 | 4 | 5 |
| Enzymatic Hydrolysis Temperature (°C) | C | 34 | 36 | 38 |
| Experimental Number | A Fungal Age/d | B Enzymatic Hydrolysis Time/h | C Enzymatic Hydrolysis Temperature/°C | Protoplast Yield (CFU/mL × 106) |
|---|---|---|---|---|
| 1 | 1 | 3 | 34 | 15.30 |
| 2 | 1 | 4 | 38 | 3.55 |
| 3 | 1 | 5 | 36 | 8.00 |
| 4 | 2 | 3 | 38 | 5.50 |
| 5 | 2 | 4 | 36 | 23.50 |
| 6 | 2 | 5 | 34 | 51.00 |
| 7 | 3 | 3 | 36 | 1.00 |
| 8 | 3 | 4 | 34 | 1.40 |
| 9 | 3 | 5 | 38 | 0.25 |
| k1 | 8.95 | 7.26 | 22.56 | |
| k2 | 26.67 | 9.48 | 3.10 | |
| k3 | 0.88 | 19.75 | 10.80 | |
| R | 25.78 | 12.49 | 19.46 |
| Regeneration Media | Protoplast Regeneration Rate (%) |
|---|---|
| PDA | control |
| PDA + KCl | 2.87 ± 0.51 d |
| PDA + NaCl | 1.39 ± 0.96 d |
| PDA + glucose | 4.78 ± 0.85 c |
| PDA + sucrose | 10.43 ± 1.18 a |
| PDA + mannitol | 7.11 ± 1.03 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, H.; Ruan, H.; Wu, T.; Zhang, H.; Dong, R.; Jiang, Y. Optimization of Protoplast Preparation and Establishment of PEG-Mediated Genetic Transformation Method in Cordyceps cicadae. J. Fungi 2025, 11, 219. https://doi.org/10.3390/jof11030219
Qi H, Ruan H, Wu T, Zhang H, Dong R, Jiang Y. Optimization of Protoplast Preparation and Establishment of PEG-Mediated Genetic Transformation Method in Cordyceps cicadae. Journal of Fungi. 2025; 11(3):219. https://doi.org/10.3390/jof11030219
Chicago/Turabian StyleQi, Haikun, Haihua Ruan, Tao Wu, Hongyang Zhang, Rui Dong, and Yanjun Jiang. 2025. "Optimization of Protoplast Preparation and Establishment of PEG-Mediated Genetic Transformation Method in Cordyceps cicadae" Journal of Fungi 11, no. 3: 219. https://doi.org/10.3390/jof11030219
APA StyleQi, H., Ruan, H., Wu, T., Zhang, H., Dong, R., & Jiang, Y. (2025). Optimization of Protoplast Preparation and Establishment of PEG-Mediated Genetic Transformation Method in Cordyceps cicadae. Journal of Fungi, 11(3), 219. https://doi.org/10.3390/jof11030219