
Sequential Induction of Drug Resistance and Characterization of an Initial Candida albicans Drug-Sensitive Isolate

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Candida albicans Isolates
2.2. DNA Extraction
2.3. DNA Sequencing and Sequence Analysis
2.3.1. Single-Nucleotide Polymorphism Detection
2.3.2. Annotation
2.4. Quantification of Ergosterol Content in the Plasma Membrane
2.5. Virulence Assay
2.6. Biofilm Formation Assay
2.7. Quantification of Cell Wall Chitin
2.8. Growth Kinetics
2.9. Statistical Analysis
3. Results
3.1. Ergosterol Quantification Assay
3.2. Chitin Quantification Assay
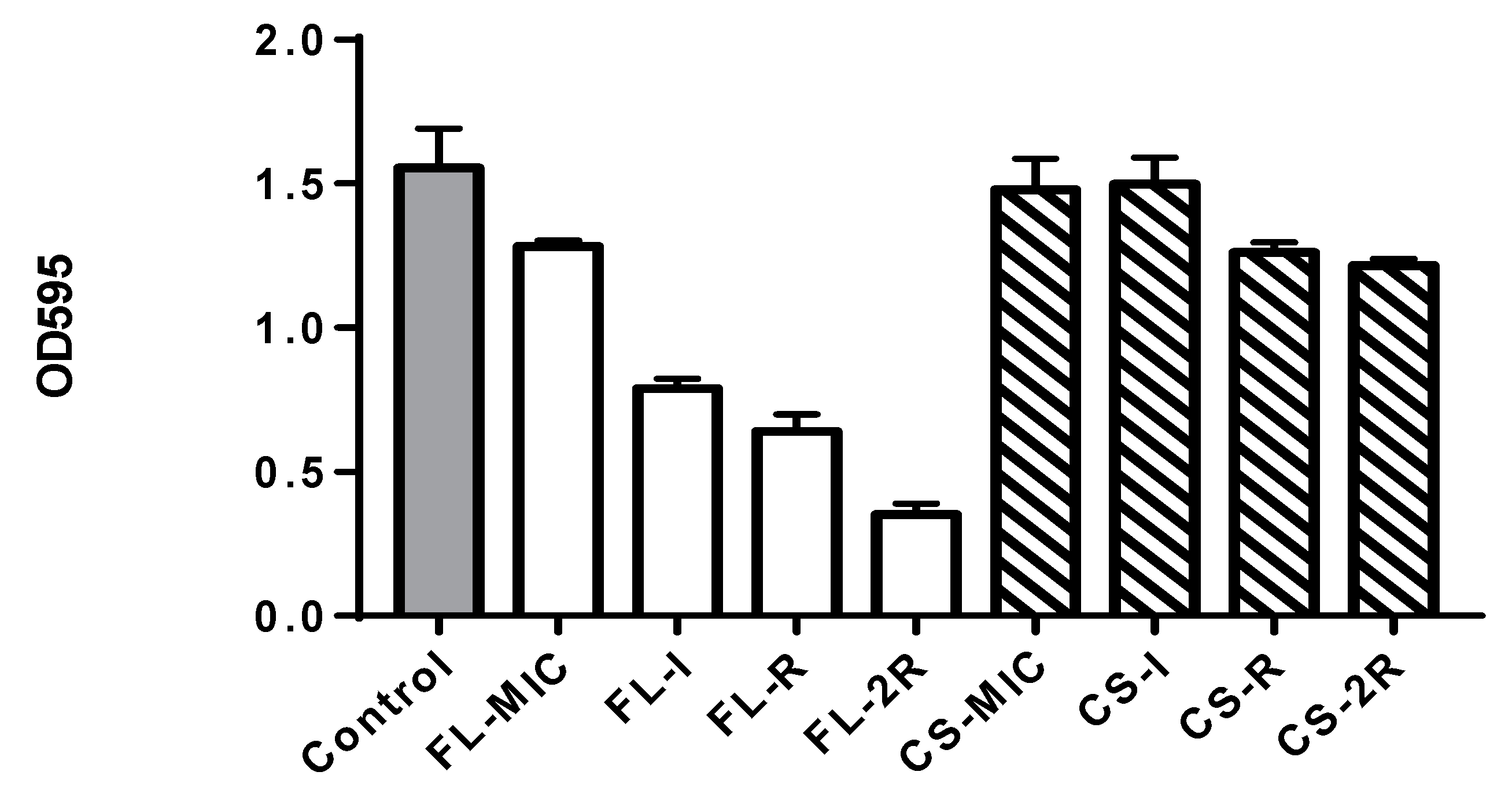
3.3. Biofilm Quantification Assay
3.4. Growth Rate Assay
3.5. Virulence Assay
3.6. WGS Analysis
3.6.1. Fluconazole Isolates
3.6.2. Caspofungin Isolates
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fattouh, N.; Hdayed, D.; Geukgeuzian, G.; Tokajian, S.; Khalaf, R.A. Molecular mechanism of fluconazole resistance and pathogenicity attributes of Lebanese Candida albicans hospital isolates. Fungal Genet. Biol. 2021, 153, 103575. [Google Scholar] [CrossRef] [PubMed]
- Mora Carpio, A.L.; Climaco, A. Fungemia Candidiasis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK436012/ (accessed on 1 February 2024).
- Villa, S.; Hamideh, M.; Weinstock, A.; Qasim, M.N.; Hazbun, T.R.; Sellam, A.; Hernday, A.D.; Thangamani, S. Transcriptional control of hyphal morphogenesis in Candida albicans. FEMS Yeast Res. 2020, 20, foaa005. [Google Scholar] [CrossRef] [PubMed]
- Kadosh, D. Regulatory Mechanisms Controlling Morphology and Pathogenesis in Candida albicans. Curr. Opin. Microbiol. 2019, 52, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.A.; Gow, N.A.R. Mannosylation in Candida albicans: Role in cell wall function and immune recognition. Mol. Microbiol. 2013, 90, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Robbins, N.; Wright, G.D.; Cowen, L.E. Antifungal Drugs: The Current Armamentarium and Development of New Agents. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Lakhani, P.; Patil, A.; Majumdar, S. Challenges in the Polyene- and Azole-Based Pharmacotherapy of Ocular Fungal Infections. J. Ocul. Pharmacol. Ther. 2019, 35, 6–22. [Google Scholar] [CrossRef] [PubMed]
- Sucher, A.J.; Chahine, E.B.; Balcer, H.E. Echinocandins: The newest class of antifungals. Ann. Pharmacother. 2009, 43, 1647–1657. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of Antifungal Drug Resistance. Cold Spring Harb. Perspect. Med. 2015, 5, a019752. [Google Scholar] [CrossRef] [PubMed]
- Suchodolski, J.; Muraszko, J.; Korba, A.; Bernat, P.; Krasowska, A. Lipid composition and cell surface hydrophobicity of Candida albicans influence the efficacy of fluconazole–gentamicin treatment. Yeast 2020, 37, 117–129. [Google Scholar] [CrossRef]
- Sardari, A.; Zarrinfar, H.; Mohammadi, R. Detection of ERG11 point mutations in Iranian fluconazole-resistant Candida albicans isolates. Curr. Med. Mycol. 2019, 5, 7–14. [Google Scholar] [CrossRef]
- Pham, C.D.; Iqbal, N.; Bolden, C.B.; Kuykendall, R.J.; Harrison, L.H.; Farley, M.M.; Schaffner, W.; Beldavs, Z.G.; Chiller, T.M.; Park, B.J.; et al. Role of FKS Mutations in Candida glabrata: MIC Values, Echinocandin Resistance, and Multidrug Resistance. Antimicrob. Agents Chemother. 2014, 58, 4690–4696. [Google Scholar] [CrossRef] [PubMed]
- Toutounji, M.; Tokajian, S.; Khalaf, R.A. Genotypic and phenotypic characterization of Candida albicans Lebanese hospital isolates resistant and sensitive to caspofungin. Fungal Genet. Biol. 2019, 127, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Hickman, R.A.; Leangapichart, T.; Lunha, K.; Jiwakanon, J.; Angkititrakul, S.; Magnusson, U.; Sunde, M.; Järhult, J.D. Exploring the Antibiotic Resistance Burden in Livestock, Livestock Handlers and Their Non-Livestock Handling Contacts: A One Health Perspective. Front. Microbiol. 2021, 12, 651461. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Ter-Hovhannisyan, V.; Lomsadze, A.; Chernoff, Y.O.; Borodovsky, M. Gene prediction in novel fungal genomes using an ab initio algorithm with unsupervised training. Genome Res. 2008, 18, 1979–1990. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Blachowicz, A.; Chiang, A.J.; Elsaesser, A.; Kalkum, M.; Ehrenfreund, P.; Stajich, J.E.; Torok, T.; Wang, C.C.C.; Venkateswaran, K. Proteomic and Metabolomic Characteristics of Extremophilic Fungi under Simulated Mars Conditions. Front. Microbiol. 2019, 10, 1013. [Google Scholar] [CrossRef] [PubMed]
- Vasquez-Gross, H.; Kaur, S.; Epstein, L.; Dubcovsky, J. A haplotype-phased genome of wheat stripe rust pathogen Puccinia striiformis f. sp. tritici, race PST-130 from the Western USA. PLoS ONE 2020, 15, e0238611. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Zdobnov, E.M. BUSCO: Assessing Genomic Data Quality and Beyond. Curr. Protoc. 2021, 1, e323. [Google Scholar] [CrossRef] [PubMed]
- Arthington-Skaggs, B.A.; Jradi, H.; Desai, T.; Morrison, C.J. Quantitation of Ergosterol Content: Novel Method for Determination of Fluconazole Susceptibility of Candida albicans. J. Clin. Microbiol. 1999, 37, 3332–3337. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef]
- Kapteyn, J.C.; Hoyer, L.L.; Hecht, J.E.; Müller, W.H.; Andel, A.; Verkleij, A.J.; Makarow, M.; Van Den Ende, H.; Klis, F.M. The cell wall architecture of Candida albicans wild-type cells and cell wall-defective mutants. Mol. Microbiol. 2000, 35, 601–611. [Google Scholar] [CrossRef]
- Chaffin, W.L. Candida albicans Cell Wall Proteins. Microbiol. Mol. Biol. Rev. MMBR 2008, 72, 495–544. [Google Scholar] [CrossRef] [PubMed]
- Akins, R.A. An update on antifungal targets and mechanisms of resistance in Candida albicans. Med. Mycol. 2005, 43, 285–318. [Google Scholar] [CrossRef]
- Yu, P.K.; Moron-Espiritu, L.S.; Lao, A.R. Mathematical Modeling of Fluconazole Resistance in the Ergosterol Pathway of Candida albicans. mSystems 2022, 7, e0069122. [Google Scholar] [CrossRef]
- Peron, I.H.; Reichert-Lima, F.; Busso-Lopes, A.F.; Nagasako, C.K.; Lyra, L.; Moretti, M.L.; Schreiber, A.Z. Resistance Surveillance in Candida albicans: A Five-Year Antifungal Susceptibility Evaluation in a Brazilian University Hospital. PLoS ONE 2016, 11, e0158126. [Google Scholar] [CrossRef]
- Knafler, H.C.; Smaczynska-de Rooij, I.I.; Walker, L.A.; Lee, K.K.; Gow, N.A.; Ayscough, K.R. AP-2-dependent endocytic recycling of the chitin synthase Chs3 regulates polarized growth in Candida albicans. MBio 2019, 10, e02421-18. [Google Scholar] [CrossRef] [PubMed]
- Bruzual, I.; Riggle, P.; Hadley, S.; Kumamoto, C.A. Biofilm formation by fluconazole-resistant Candida albicans strains is inhibited by fluconazole. J. Antimicrob. Chemother. 2007, 59, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Iyer, K.R.; Robbins, N.; Cowen, L.E. The role of Candida albicans stress response pathways in antifungal tolerance and resistance. iScience 2022, 25, 103953. [Google Scholar] [CrossRef] [PubMed]
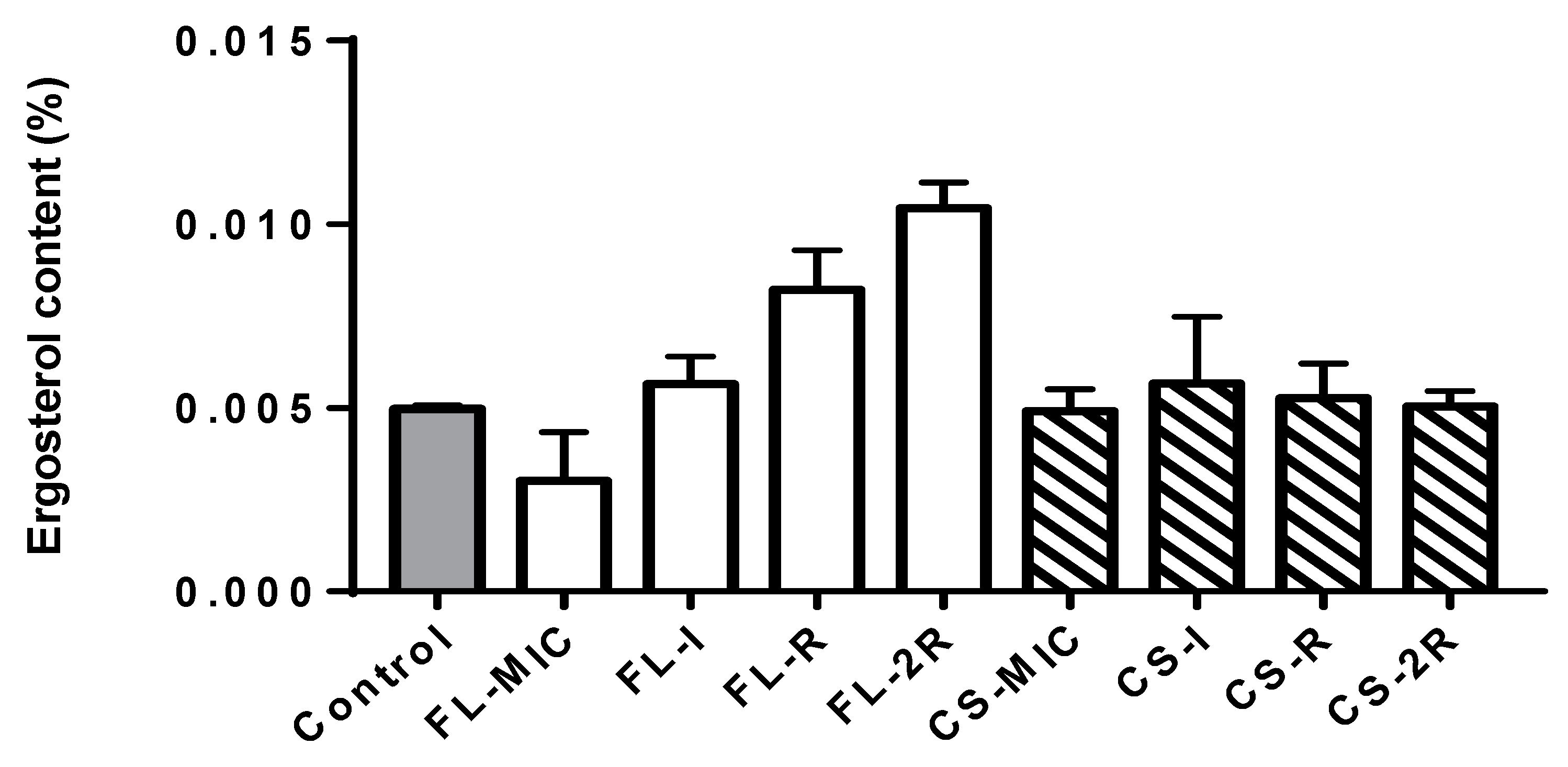
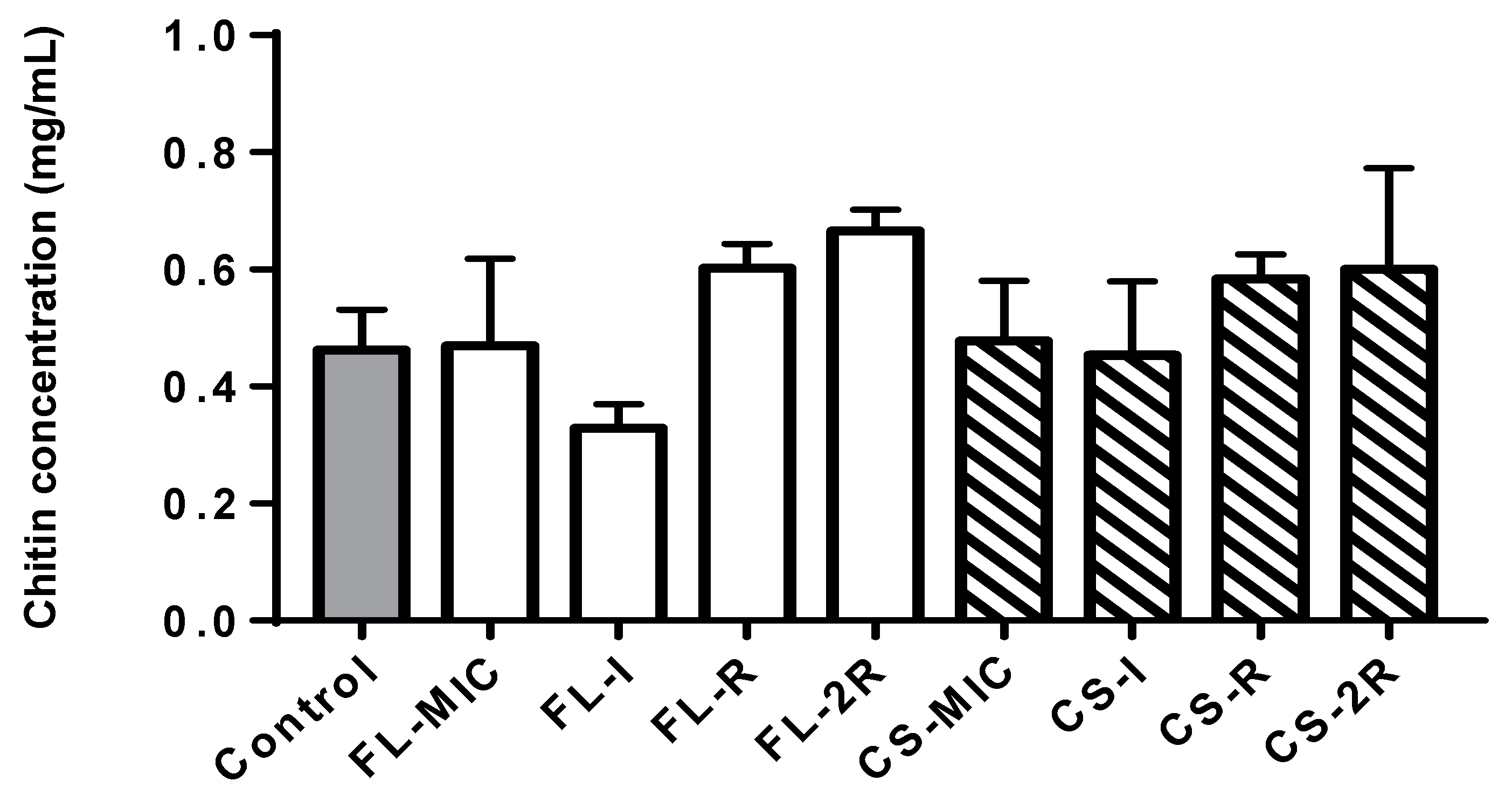



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Fluconazole Concentration |
|---|---|
| FL–MIC | 0.190 µg/mL |
| FL–Int | 4.095 µg/mL |
| FL–R | 8.000 µg/mL |
| FL–2R | 16.000 µg/mL |
| Isolate | Caspofungin Concentration |
|---|---|
| CS–MIC | 0.190 µg/mL |
| CS–Int | 0.595 µg/mL |
| CS–R | 1.000 µg/mL |
| CS–2R | 2.000 µg/mL |
| FL-MIC | FL-I | FL-R | FL-2R | ||
|---|---|---|---|---|---|
| Function | Genes | AA Change | AA Change | AA Change | AA Change |
| Drug Resistance | ERG11 | L220L | L220L–L480L | L220L–L480L | L220L–T128K–L264S |
| CDR2 | I704M–R365Q | I704M–M617L | I704M–M617L | R365Q–I704M–E374K | |
| MSH2 | D73D | T141T | V127 | V127A | |
| CHS3 | - | R762P | L321P–D388G–R762P | L321P–D388G–R762P | |
| ERG4 | - | - | - | M157V | |
| Stress response | SOD5 | - | A210P | A210P | A210P–G214D |
| SOD6 | - | S225P | S225P | S225P | |
| HSP60 | K67K | N281N | S267A | S267A | |
| MNL1 | H349R | H349R–I175T | H349R–I175T | H349R–I175T | |
| Phenotypic Switching/dimorphism | SAP1 | L53 | L53I | L53I | L53I |
| CPH1 | T510A | T510A | T510A | T510A | |
| EFG1 | K251K | K251K | I384T | I384T | |
| MFG1 | - | A68P | A68P–Q76_Q80del | A68P–Q76_Q80del | |
| SIR2 | L213 | L213 | L213 | L213S | |
| RFG1 | - | - | Q118del | Q118del | |
| FGR15 | V131A | V131A | V131A–S361L | V131A–S361L | |
| Biofilm Formation and Immune Evasion | HWP1 | - | - | T296A–S441P | T296A–S441P |
| ALS4 | I79V–V83I | I79V–V83I–V111A | I79V–V83I–V111A– S186P–D198N | I79V–V83I–V111A– S186P–D198N | |
| NAG3 | - | R158* | Y278H | Y278H | |
| UME6 | A146A | N257S–M267I | N257S–M267I | N257S–M267I –N396S | |
| SFL1 | V691I | V691I | V691I–S373L | V691I–S373L | |
| CS-MIC | CS-I | CS-R | CS-2R | ||
|---|---|---|---|---|---|
| Function | Genes | AA Change | AA Change | AA Change | AA Change |
| Drug Resistance | CDR2 | I704M | I704M–M617L | I704M–M617L–E374K | I704M–M617L–E374K–R365Q |
| CRZ2 | - | - | S330A | S330A | |
| GSC2 | A455A | A462P | A462P | A462P | |
| Stress response | SOD5 | - | A210P | A210P–G214D | A210P–G214D |
| SOD6 | - | S225P | S225P | S225P | |
| HSP60 | S267A | S267A | S267A | S267A | |
| MNL1 | H349R | H349R–I175T | H349R–I175T | R269 * | |
| ATG26 | - | Y401C | E269K–Y401C | E269K–Y401C–T414I | |
| Phenotypic Switching/dimorphism | CPH1 | P451P | I79I | T510A | T510A |
| MFG1 | - | - | Q80del | Q80del | |
| RFG1 | - | - | T39S | T39S | |
| SIR2 | - | - | L213S | L213S | |
| FGR15 | - | S361L | S361L | S361L | |
| Biofilm Formation and Immune Evasion | HWP1 | - | S441P | S441P–T296A | S441P–T296A |
| ALS4 | - | I79V–V83I–V111A | I79V–V83I–V111A–S186P | I79V–V83I–V111A–S186P–D198N | |
| NAG3 | - | - | Y278H | R158 * | |
| UME6 | N257S–M267I | N257S–M267I | N257S–M267I–N396S | N257S–M267I–N396S–N484del | |
| SFL1 | V691I–S373L | V691I–S373L | V691I–S373L–T75A | T75A–Q301fs | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Hachem, S.; Fattouh, N.; Chedraoui, C.; Finianos, M.; Bitar, I.; Khalaf, R.A. Sequential Induction of Drug Resistance and Characterization of an Initial Candida albicans Drug-Sensitive Isolate. J. Fungi 2024, 10, 347. https://doi.org/10.3390/jof10050347
El Hachem S, Fattouh N, Chedraoui C, Finianos M, Bitar I, Khalaf RA. Sequential Induction of Drug Resistance and Characterization of an Initial Candida albicans Drug-Sensitive Isolate. Journal of Fungi. 2024; 10(5):347. https://doi.org/10.3390/jof10050347
Chicago/Turabian StyleEl Hachem, Setrida, Nour Fattouh, Christy Chedraoui, Marc Finianos, Ibrahim Bitar, and Roy A. Khalaf. 2024. "Sequential Induction of Drug Resistance and Characterization of an Initial Candida albicans Drug-Sensitive Isolate" Journal of Fungi 10, no. 5: 347. https://doi.org/10.3390/jof10050347
APA StyleEl Hachem, S., Fattouh, N., Chedraoui, C., Finianos, M., Bitar, I., & Khalaf, R. A. (2024). Sequential Induction of Drug Resistance and Characterization of an Initial Candida albicans Drug-Sensitive Isolate. Journal of Fungi, 10(5), 347. https://doi.org/10.3390/jof10050347