

Sunken Riches: Ascomycete Diversity in the Western Mediterranean Coast through Direct Plating and Flocculation, and Description of Four New Taxa



Abstract

1. Introduction
2. Materials and Methods
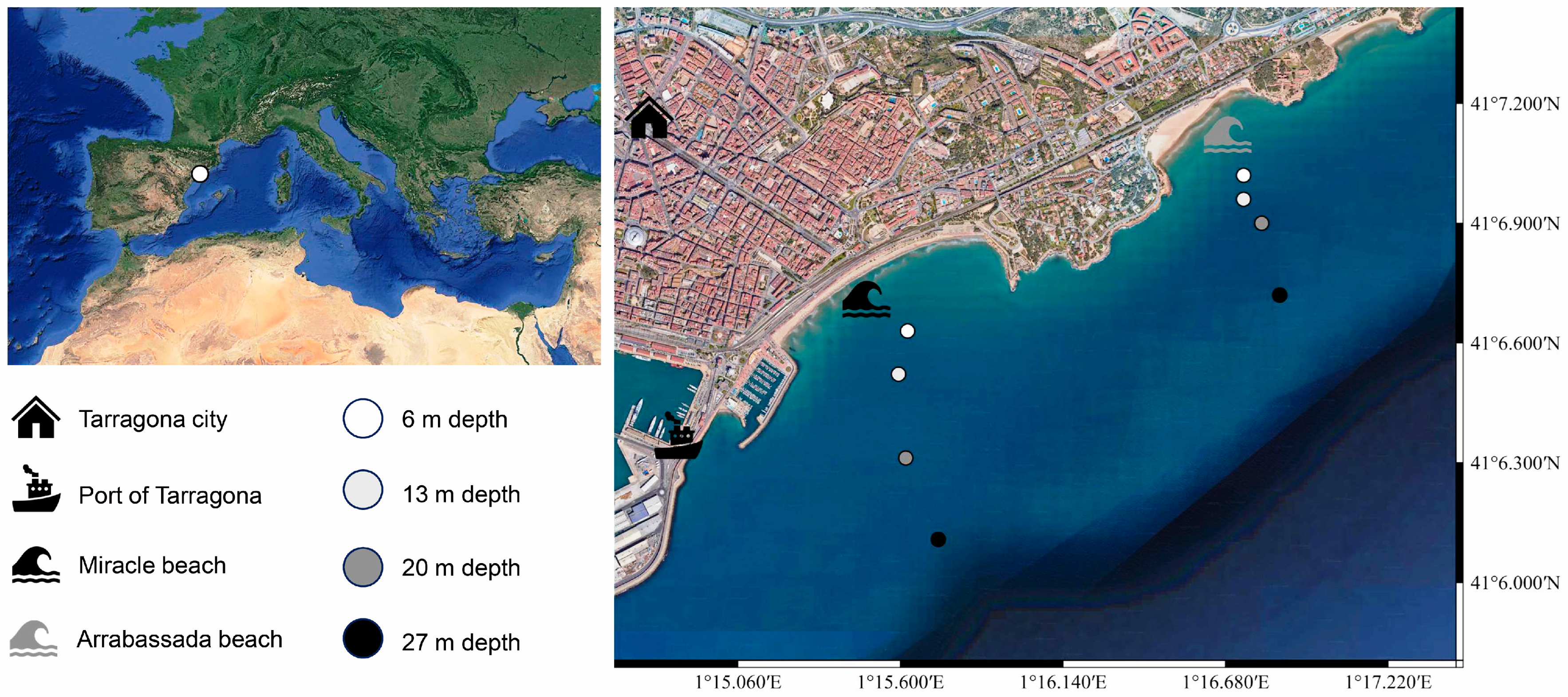
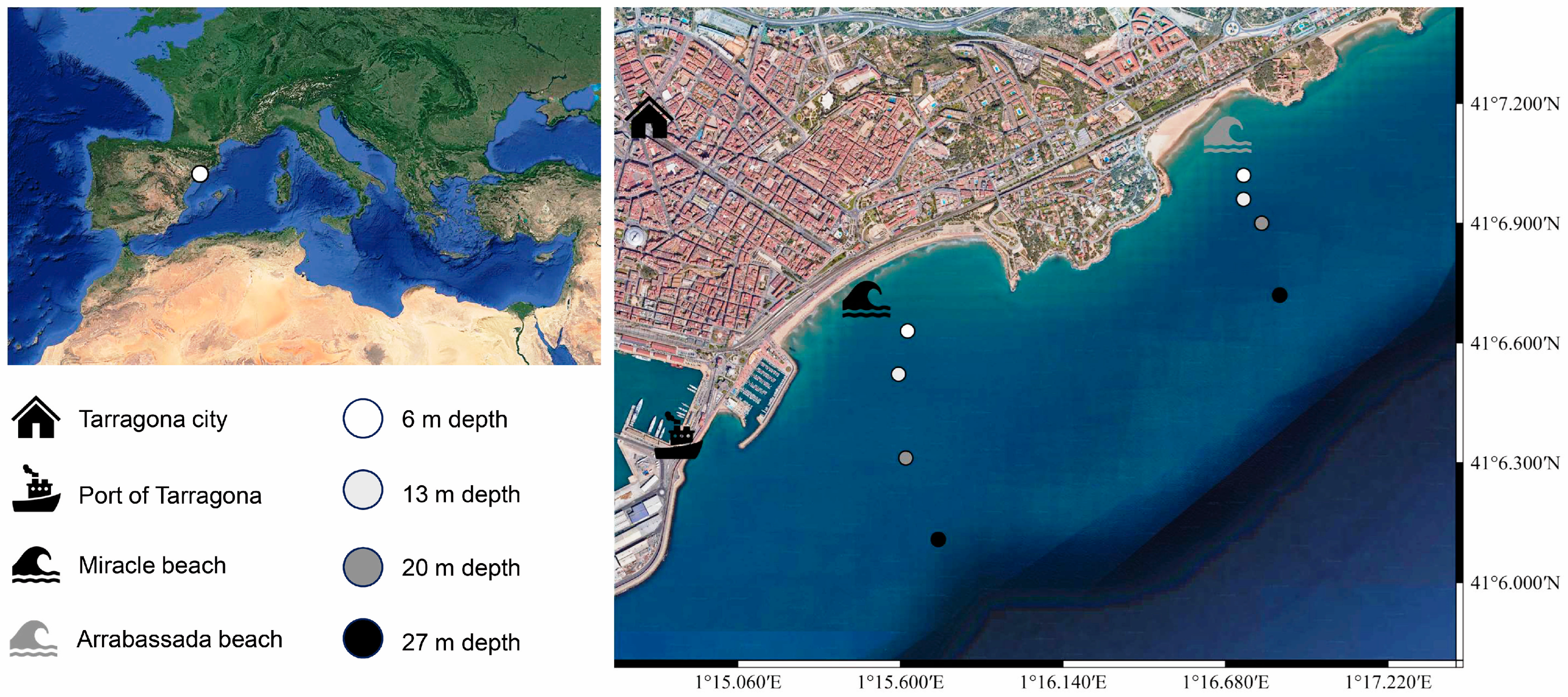
2.1. Study Area and Sampling
2.2. Direct Plating, Flocculation Pre-Treatment, Culturing, and Isolation of Fungi
2.3. Morphological Analyses
2.4. DNA Extraction, PCR Amplification, Sequencing, and Strain Identification
2.5. Phylogenetic Analyses
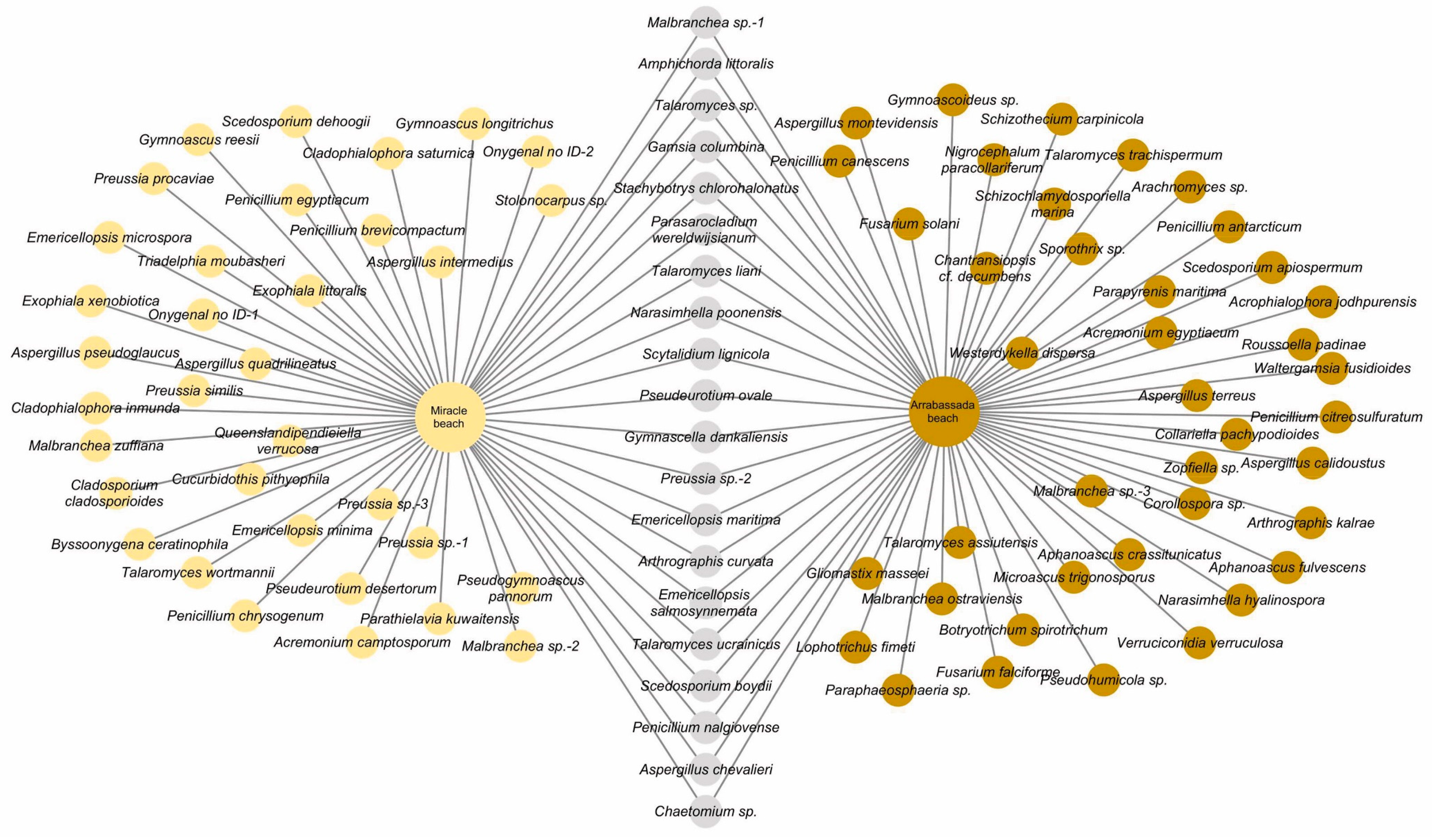
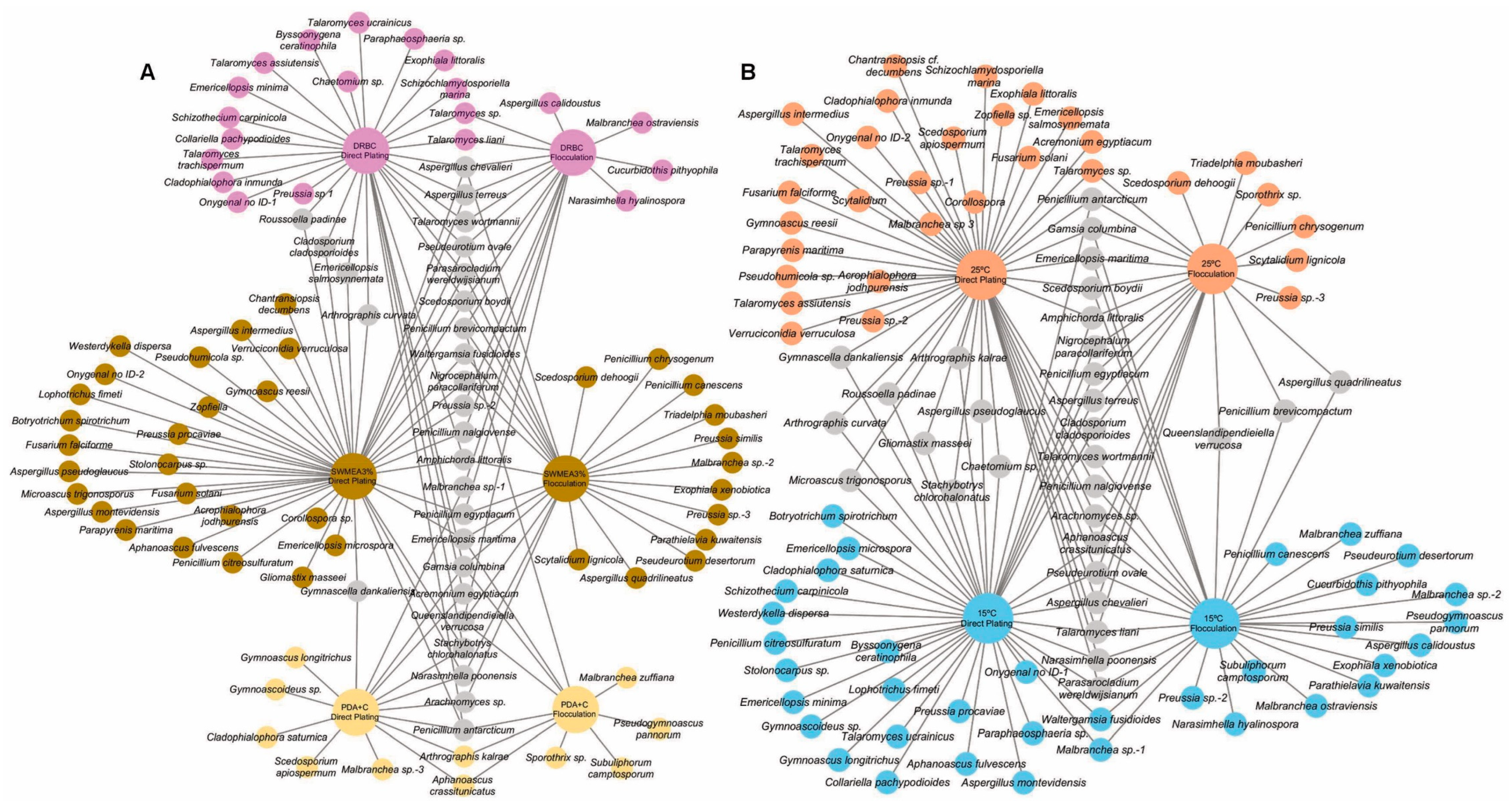
2.6. Representation of Diversity Data
3. Results
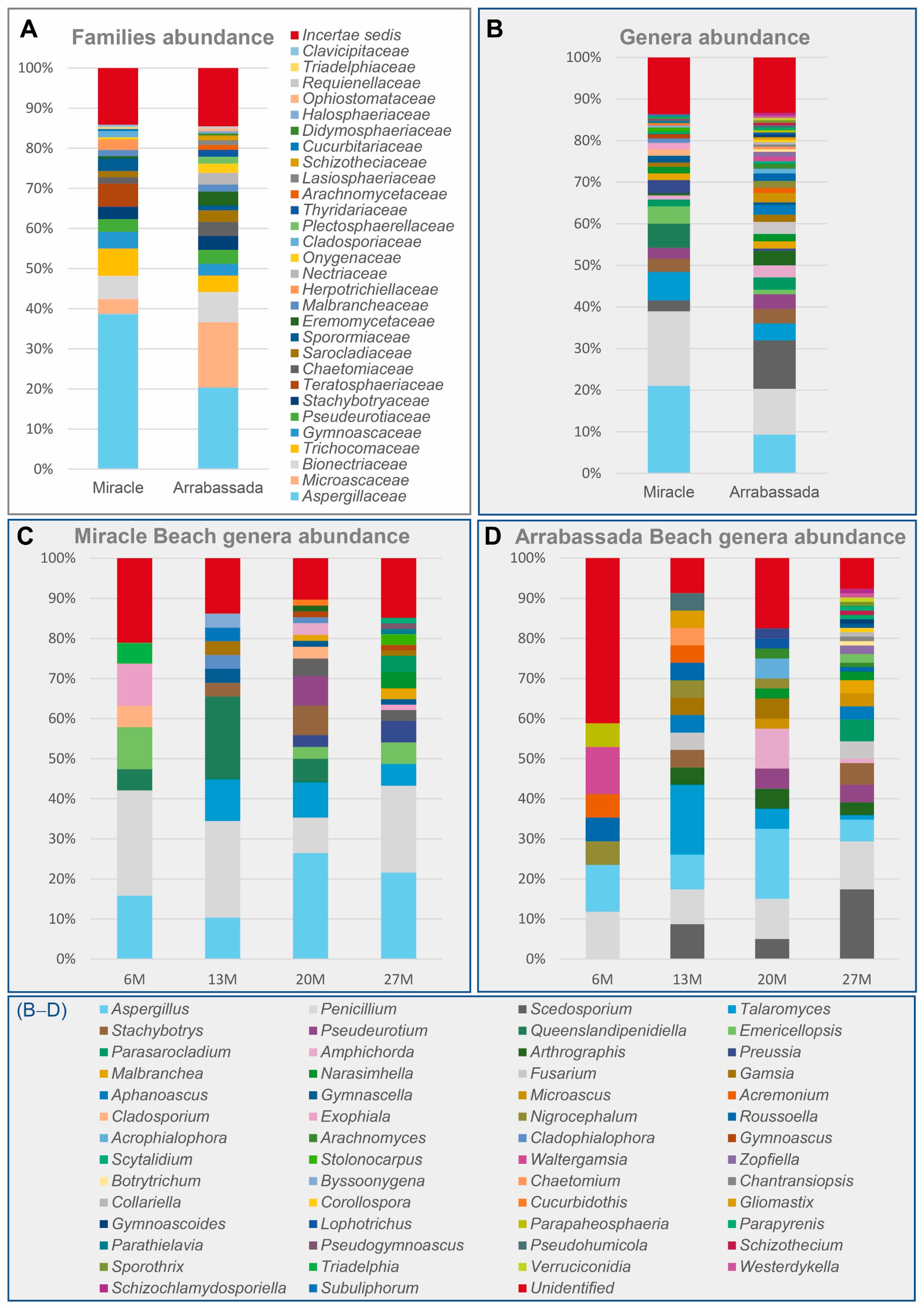
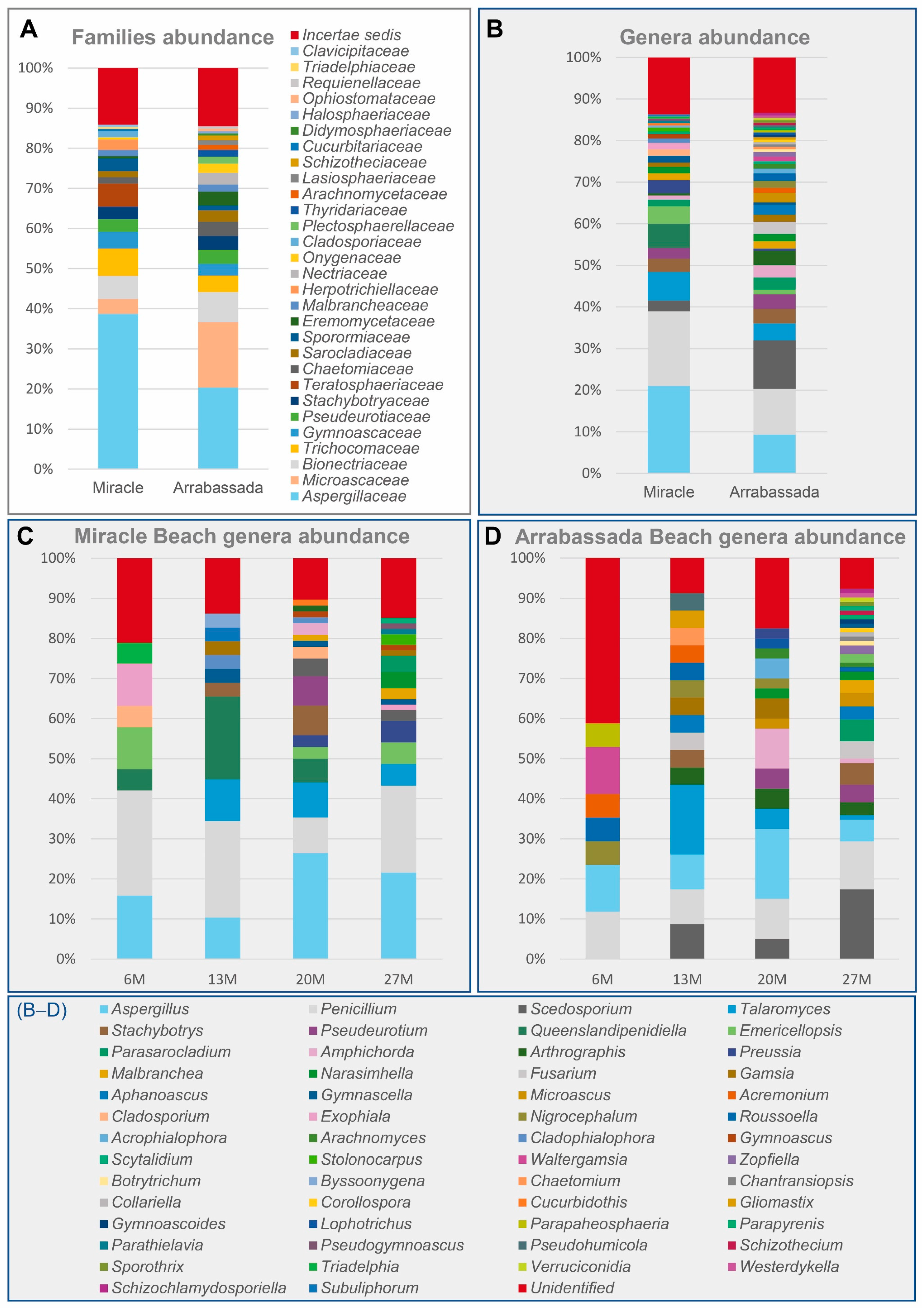
3.1. Diversity at the Family and Genus Level
3.2. Diversity at the Species Level
3.2.1. Species Detected in Miracle and Arrabassada Beaches
3.2.2. Direct Plating and SMF Pre-Treatment: Effect of Culture Media and Temperature
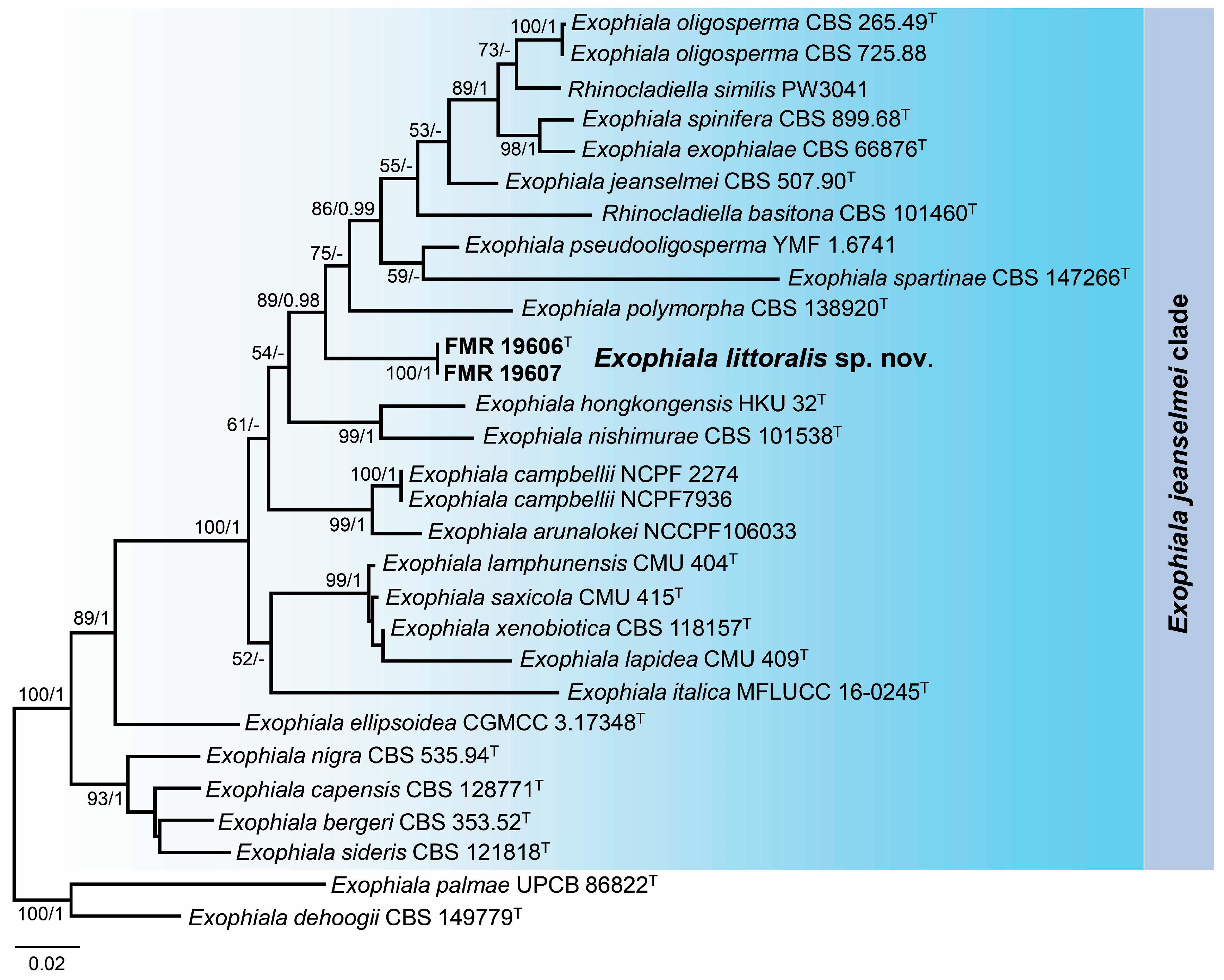
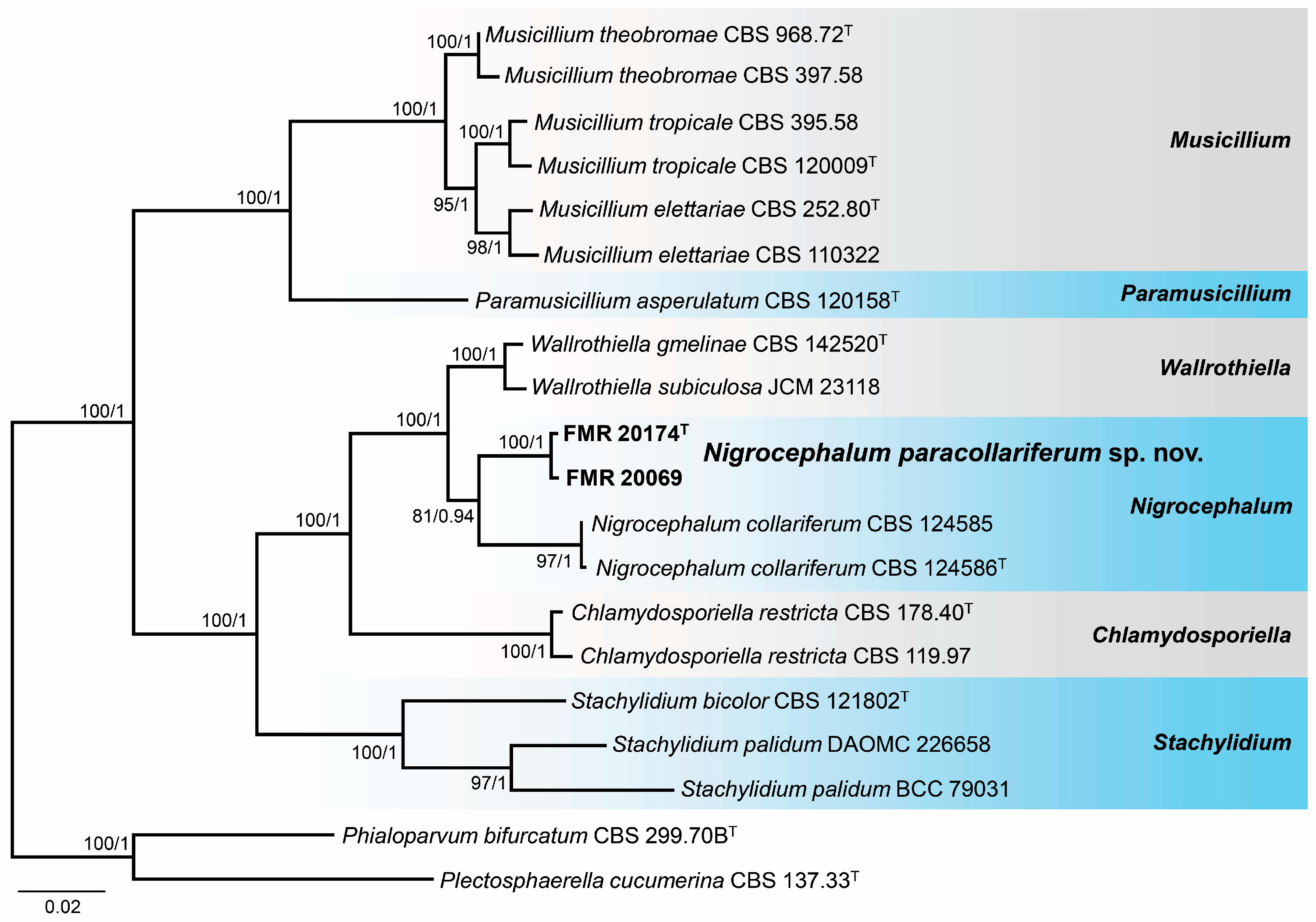
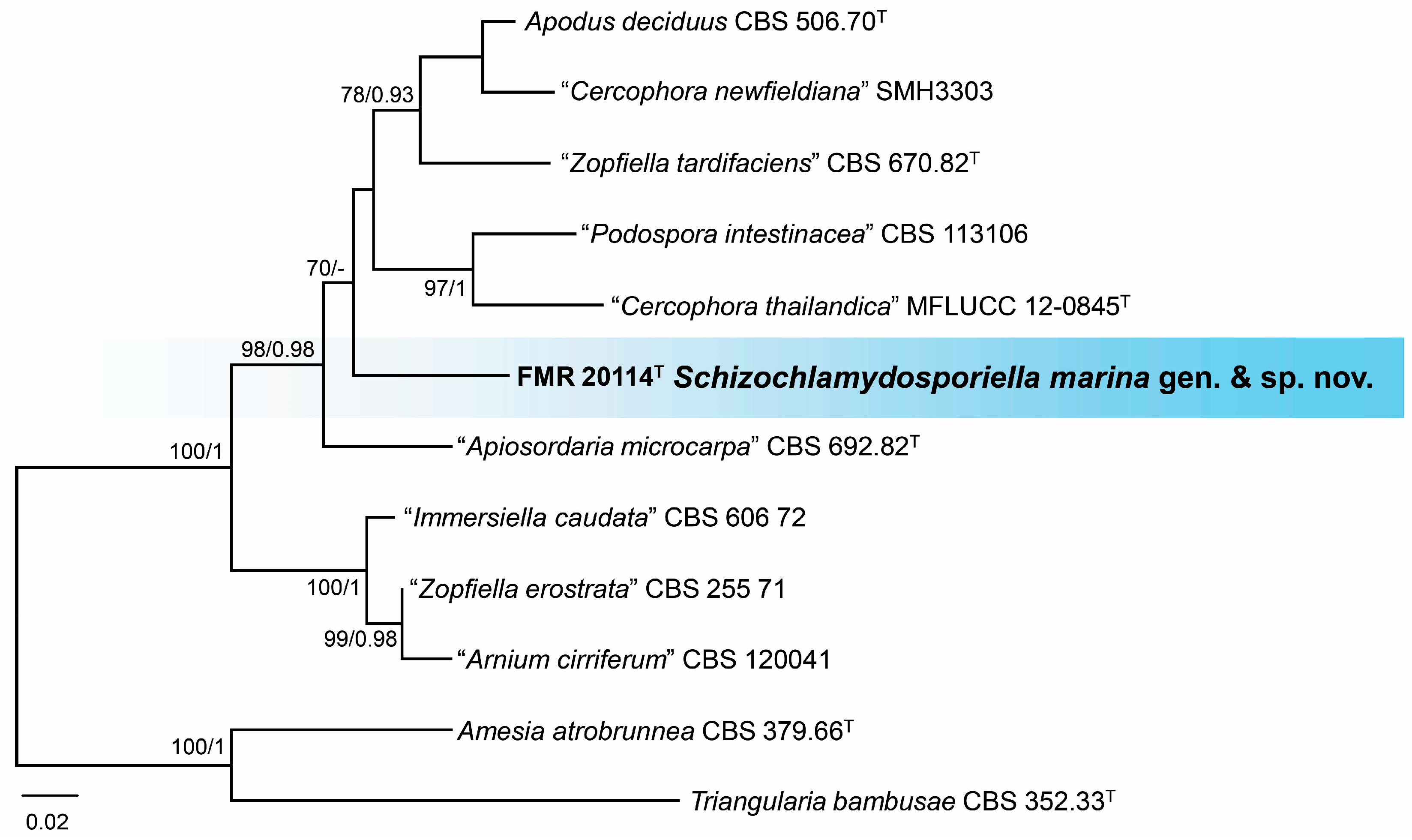
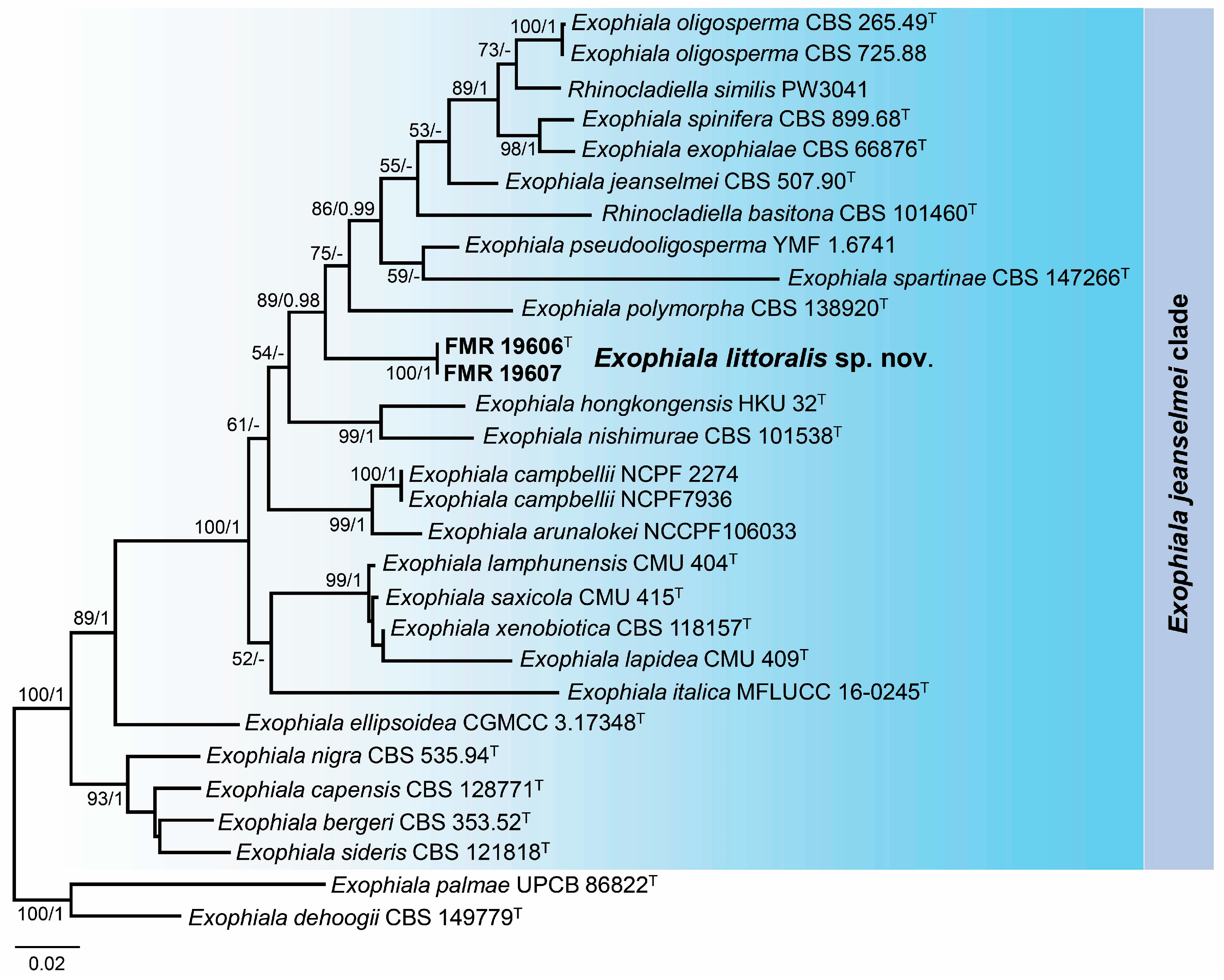
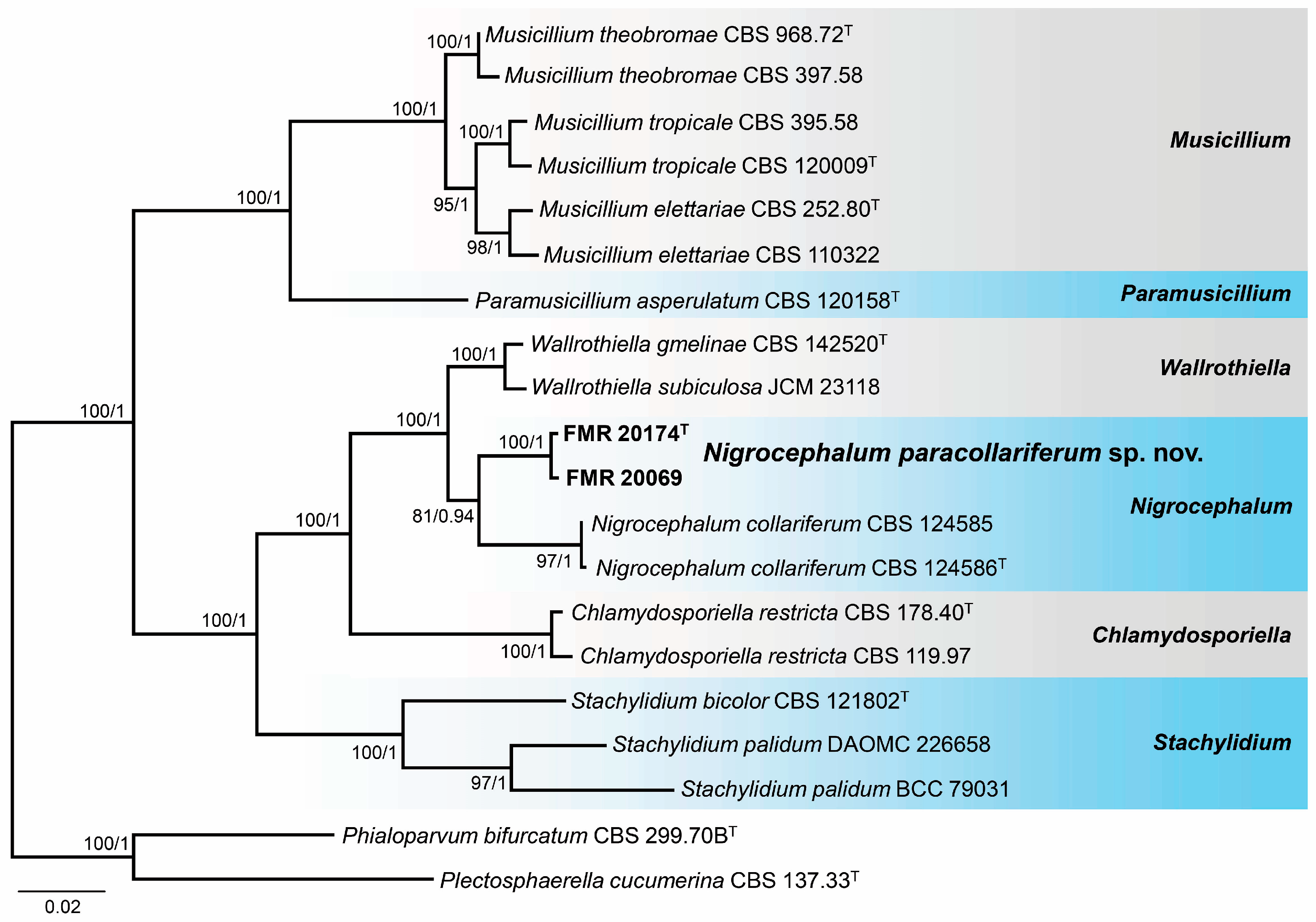
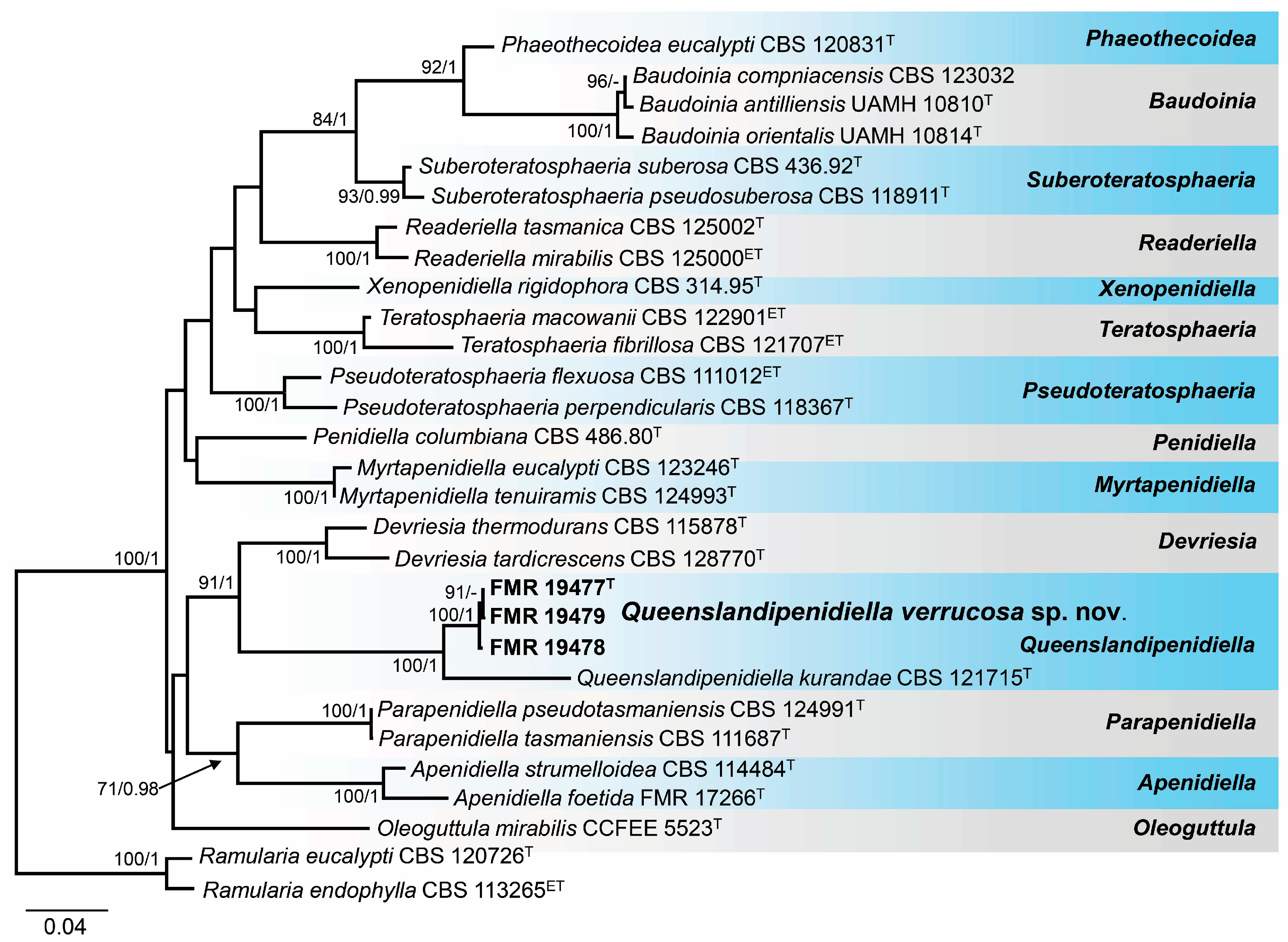
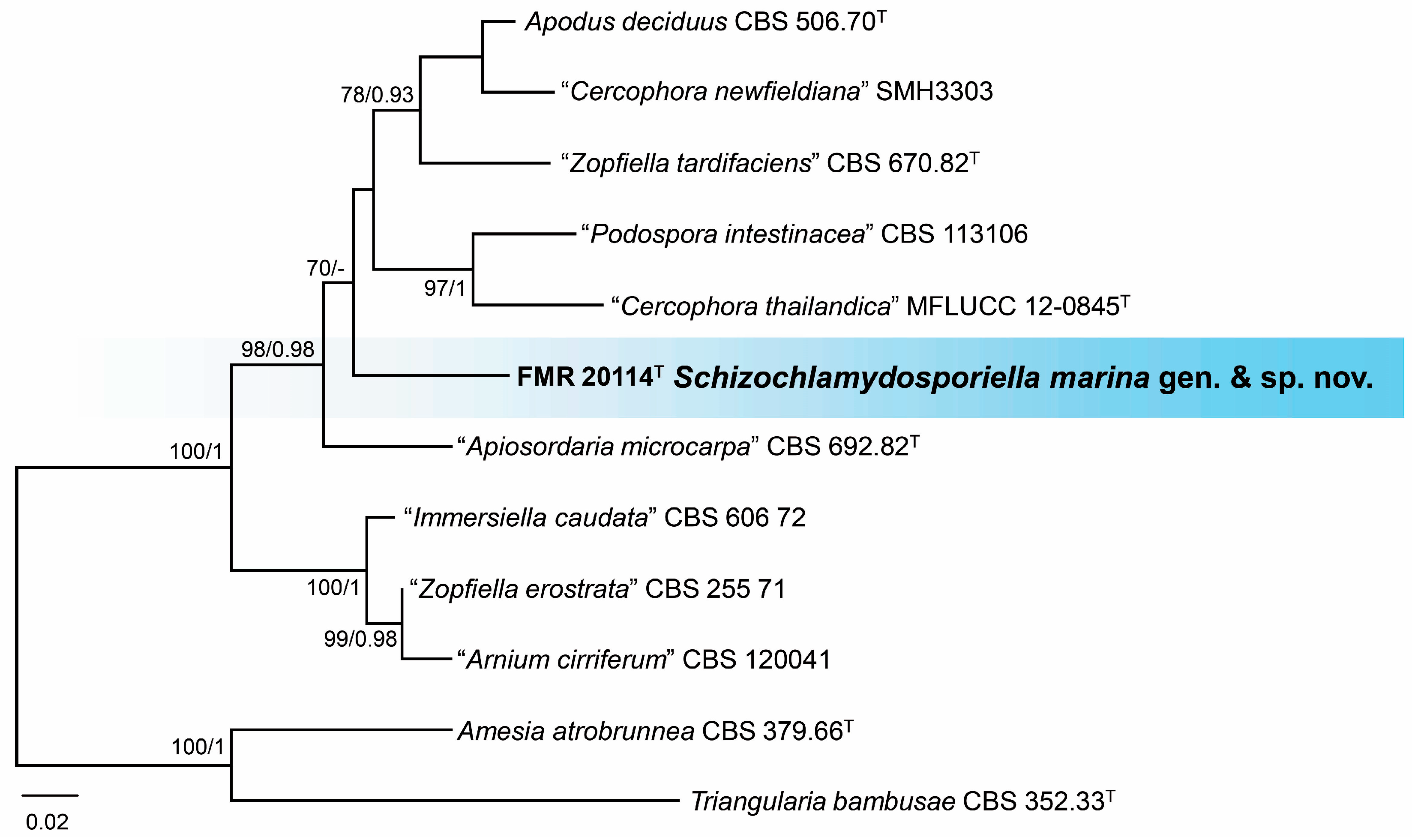
3.3. Phylogeny
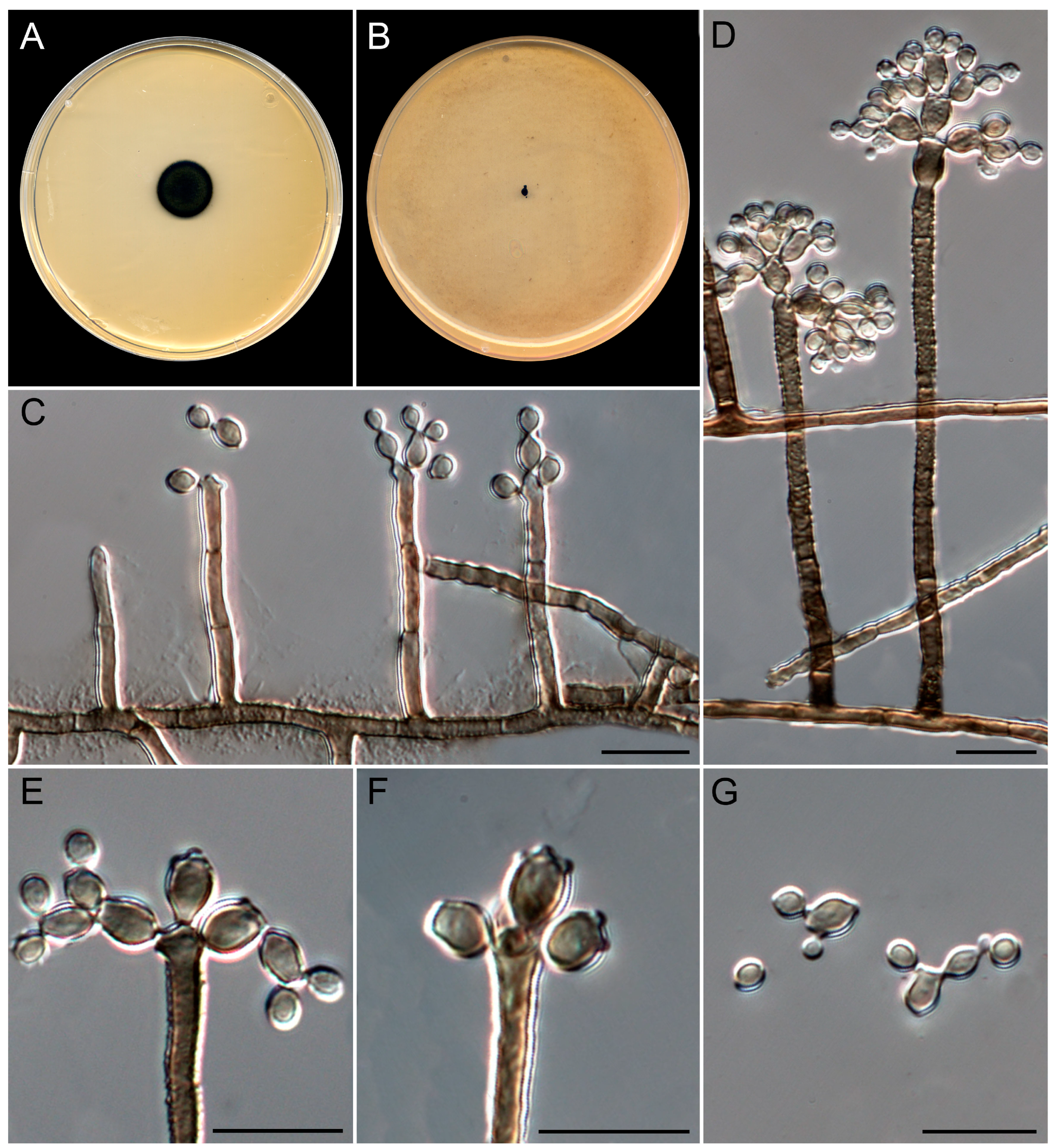
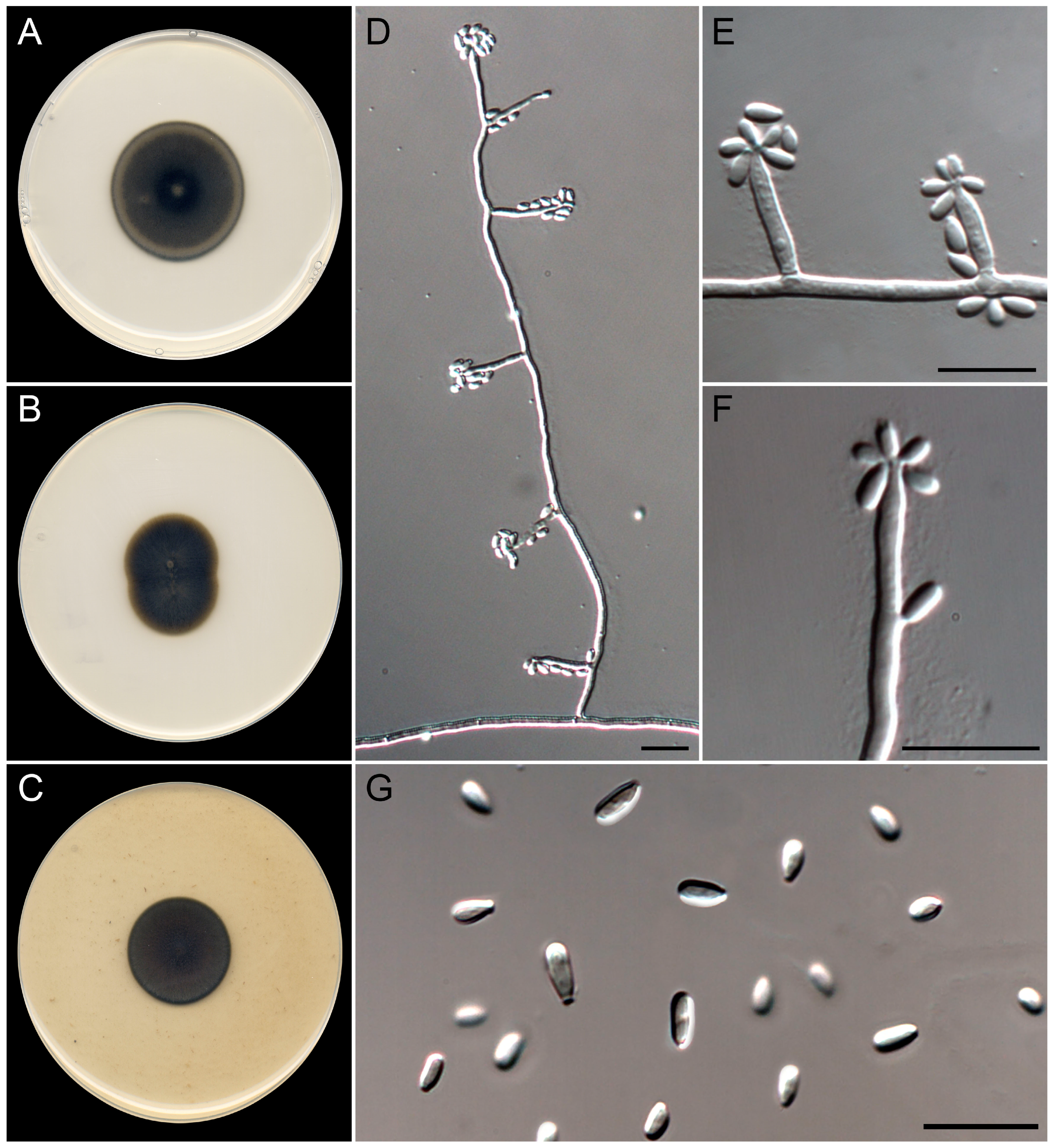
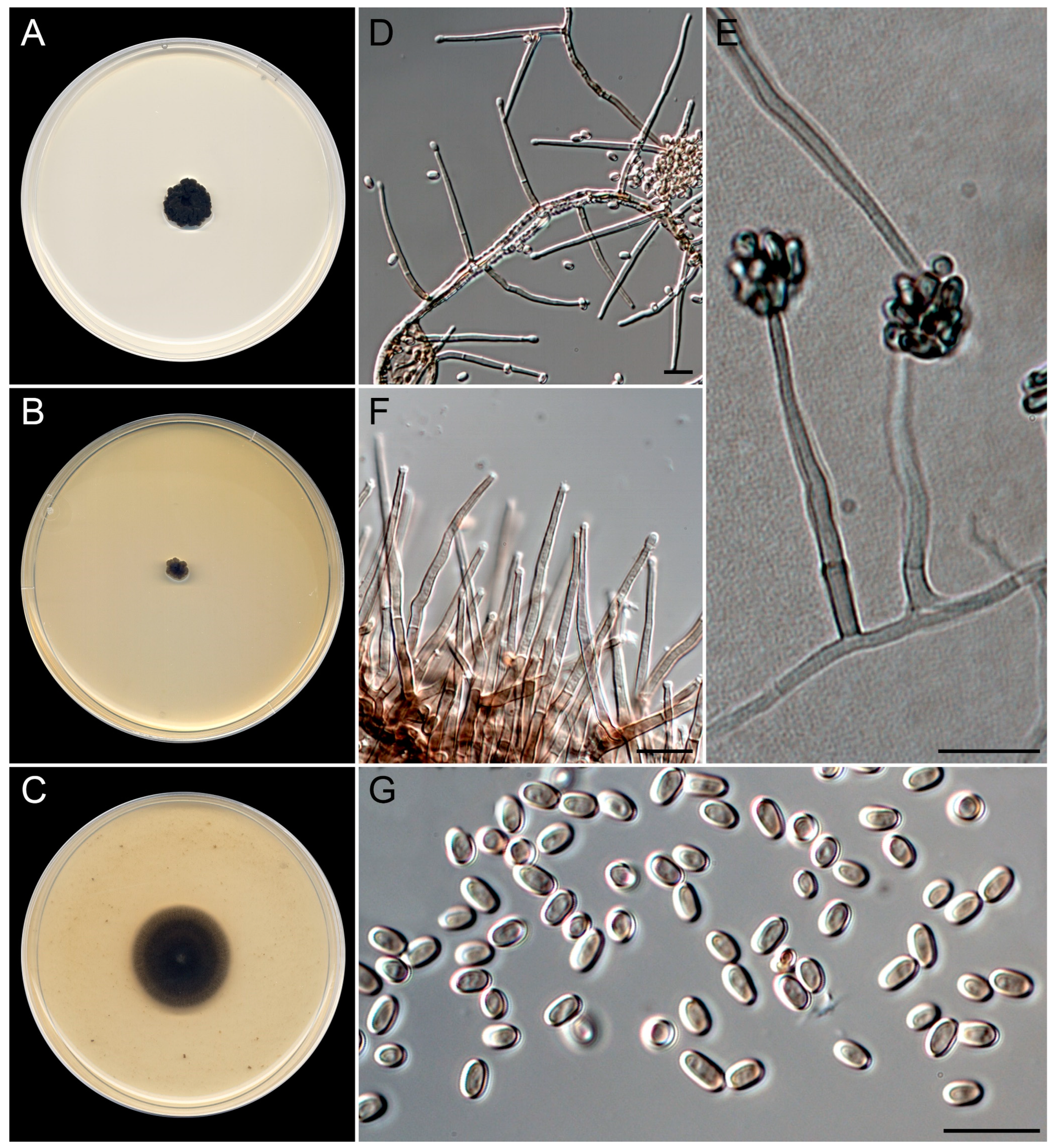
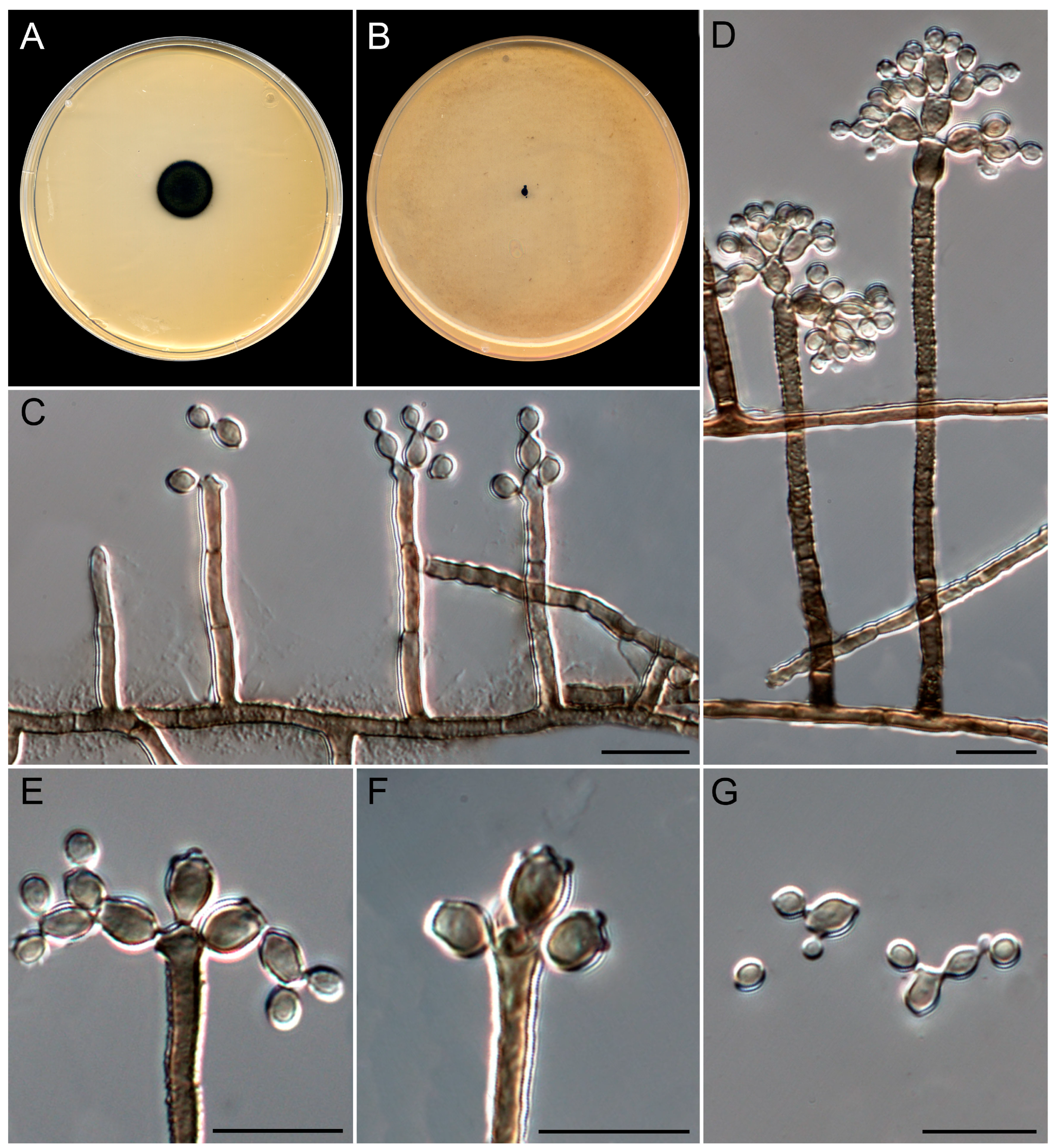
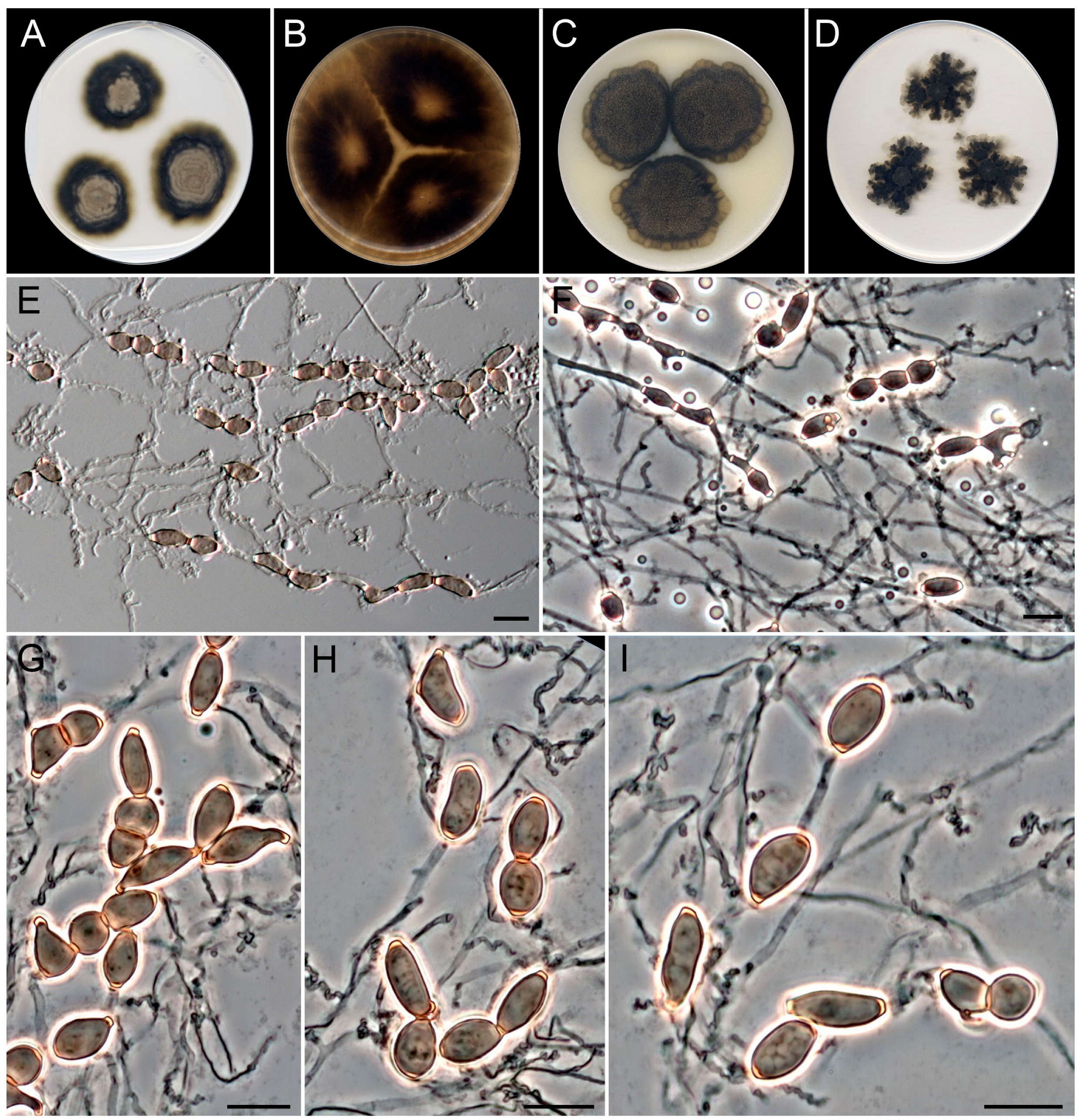
3.4. Taxonomy
4. Discussion
4.1. Effect of In Vitro Conditions for Isolation of Marine Fungi
4.2. Diversity and Insights of the Recovered Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niskanen, T.; Lücking, R.; Dahlberg, A.; Gaya, E.; Suz, L.M.; Mikryukov, V.; Liimatainen, K.; Druzhinina, I.; Westrip, J.R.S.; Mueller, G.M.; et al. Pushing the Frontiers of Biodiversity Research: Unveiling the Global Diversity, Distribution, and Conservation of Fungi. Annu. Rev. Environ. Resour. 2023, 48, 149–176. [Google Scholar] [CrossRef]
- Antonelli, A.; Smith, R.J.; Fry, C.; Simmonds, M.S.J.; Kersey, P.J.; Pritchard, H.W.; Abbo, M.S.; Acedo, C.; Adams, J.; Ainsworth, A.M.; et al. State of the World’s Plants and Fungi; Royal Botanic Gardens (Kew); Sfumato Foundation: London, UK, 2020. [Google Scholar]
- Jones, E.B.G. History of Marine Mycology—A Personal Perspective. Bot. Mar. 2023, 66, 453–470. [Google Scholar] [CrossRef]
- Calabon, M.S.; Jones, E.B.G.; Pang, K.-L.; Abdel-Wahab, M.A.; Jin, J.; Devadatha, B.; Sadaba, R.B.; Apurillo, C.C.; Hyde, K.D. Updates on the Classification and Numbers of Marine Fungi. Bot. Mar. 2023, 66, 213–238. [Google Scholar] [CrossRef]
- Debeljak, P.; Baltar, F. Fungal Diversity and Community Composition across Ecosystems. J. Fungi 2023, 9, 510. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.B.G.; Pang, K.-L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J.; et al. An Online Resource for Marine Fungi. Fungal Divers. 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, B.R.; Joppa, L.N.; Pimm, S.L.; Laurance, W.F. What We Know and Don’t Know about Earth’s Missing Biodiversity. Trends Ecol. Evol. 2012, 27, 501–510. [Google Scholar] [CrossRef]
- Bovio, E.; Gnavi, G.; Prigione, V.; Spina, F.; Denaro, R.; Yakimov, M.; Calogero, R.; Crisafi, F.; Varese, G.C. The Culturable Mycobiota of a Mediterranean Marine Site after an Oil Spill: Isolation, Identification and Potential Application in Bioremediation. Sci. Total Environ. 2017, 576, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Garzoli, L.; Gnavi, G.; Tamma, F.; Tosi, S.; Varese, G.C.; Picco, A.M. Sink or Swim: Updated Knowledge on Marine Fungi Associated with Wood Substrates in the Mediterranean Sea and Hints about Their Potential to Remediate Hydrocarbons. Prog. Oceanogr. 2015, 137, 140–148. [Google Scholar] [CrossRef]
- Cecchi, G.; Cutroneo, L.; Di Piazza, S.; Capello, M.; Zotti, M. Culturable Fungi from Dredged and Marine Sediments from Six Ports Studied in the Framework of the SEDITERRA Project. J. Soils Sediments 2021, 21, 1563–1573. [Google Scholar] [CrossRef]
- Marchese, P.; Garzoli, L.; Gnavi, G.; O’Connell, E.; Bouraoui, A.; Mehiri, M.; Murphy, J.M.; Varese, G.C. Diversity and Bioactivity of Fungi Associated with the Marine Sea Cucumber Holothuria Poli: Disclosing the Strains Potential for Biomedical Applications. J. Appl. Microbiol. 2020, 129, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Greco, G.; Cecchi, G.; Di Piazza, S.; Cutroneo, L.; Capello, M.; Zotti, M. Fungal Characterisation of a Contaminated Marine Environment: The Case of the Port of Genoa (North-Western Italy). Webbia 2018, 73, 97–106. [Google Scholar] [CrossRef]
- Hassett, B.T.; Vonnahme, T.R.; Peng, X.; Jones, E.B.G.; Heuzé, C. Global Diversity and Geography of Planktonic Marine Fungi. Bot. Mar. 2020, 63, 121–139. [Google Scholar] [CrossRef]
- Barone, G.; Rastelli, E.; Corinaldesi, C.; Tangherlini, M.; Danovaro, R.; Dell’Anno, A. Benthic Deep-Sea Fungi in Submarine Canyons of the Mediterranean Sea. Prog. Oceanogr. 2018, 168, 57–64. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.-R.; Wijesinghe, S.N.; Raza, M.; Bao, D.-F.; Lu, L.; Tibpromma, S.; et al. The Numbers of Fungi: Contributions from Traditional Taxonomic Studies and Challenges of Metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Zhang, Z.-F.; Liu, F.; Liu, L.-R.; Li, M.; Cai, L.; Liu, S.; Mao, J. Culturing the Uncultured Marine Fungi in the Omics Age: Opportunities and Challenges. Fungal Biol. Rev. 2024, 48, 100353. [Google Scholar] [CrossRef]
- Calgua, B.; Mengewein, A.; Grunert, A.; Bofill-Mas, S.; Clemente-Casares, P.; Hundesa, A.; Wyn-Jones, A.P.; López-Pila, J.M.; Girones, R. Development and Application of a One-Step Low Cost Procedure to Concentrate Viruses from Seawater Samples. J. Virol. Methods 2008, 153, 79–83. [Google Scholar] [CrossRef]
- Calgua, B.; Rodriguez-Manzano, J.; Hundesa, A.; Suñen, E.; Calvo, M.; Bofill-Mas, S.; Girones, R. New Methods for the Concentration of Viruses from Urban Sewage Using Quantitative PCR. J. Virol. Methods 2013, 187, 215–221. [Google Scholar] [CrossRef]
- Calgua, B.; Fumian, T.; Rusiñol, M.; Rodriguez-Manzano, J.; Mbayed, V.A.; Bofill-Mas, S.; Miagostovich, M.; Girones, R. Detection and Quantification of Classic and Emerging Viruses by Skimmed-Milk Flocculation and PCR in River Water from Two Geographical Areas. Water Res. 2013, 47, 2797–2810. [Google Scholar] [CrossRef]
- Rusiñol, M.; Hundesa, A.; Cárdenas-Youngs, Y.; Fernández-Bravo, A.; Pérez-Cataluña, A.; Moreno-Mesonero, L.; Moreno, Y.; Calvo, M.; Alonso, J.L.; Figueras, M.J.; et al. Microbiological Contamination of Conventional and Reclaimed Irrigation Water: Evaluation and Management Measures. Sci. Total Environ. 2020, 710, 136298. [Google Scholar] [CrossRef]
- Bofill-Mas, S.; Hundesa, A.; Calgua, B.; Rusiñol, M.; de Motes, C.M.; Girones, R. Cost-Effective Method for Microbial Source Tracking Using Specific Human and Animal Viruses. JoVE 2011, 58, e2820. [Google Scholar] [CrossRef]
- Gonzales-Gustavson, E.; Cárdenas-Youngs, Y.; Calvo, M.; da Silva, M.F.M.; Hundesa, A.; Amorós, I.; Moreno, Y.; Moreno-Mesonero, L.; Rosell, R.; Ganges, L.; et al. Characterization of the Efficiency and Uncertainty of Skimmed Milk Flocculation for the Simultaneous Concentration and Quantification of Water-Borne Viruses, Bacteria and Protozoa. J. Microbiol. Methods 2017, 134, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lozano, C.; Pintó, J. Current Status and Future Restoration of Coastal Dune Systems on the Catalan Shoreline (Spain, NW Mediterranean Sea). J. Coast Conserv. 2018, 22, 519–532. [Google Scholar] [CrossRef]
- Expósito, N.; Rovira, J.; Sierra, J.; Folch, J.; Schuhmacher, M. Microplastics Levels, Size, Morphology and Composition in Marine Water, Sediments and Sand Beaches. Case Study of Tarragona Coast (Western Mediterranean). Sci. Total Environ. 2021, 786, 147453. [Google Scholar] [CrossRef]
- Folch, J. La Mar de Tarragona: Una Aproximació Naturalista al Litoral Més Desconegut; Publicacions Universitat Rovira i Virgili: Tarragona, Spain, 2021; pp. 1–200. [Google Scholar]
- Ruiz-Fernández, J.M.; Guillén, J.E.; Ramos-Segura, A.; Otero, M.; Tello-Antón, M.O.; Mateo, M.A.; Bernardeau-Esteller, J.; Rueda, J.L.; Urra, J.; Mateo, A.; et al. Atlas de las Praderas Marinas de España; IEO: Murcia, Spain; IEL: Alicante, Spain; UICN: Málaga, Spain, 2016; pp. 1–681. [Google Scholar]
- Folch, J.; Pascual, X. Inventario de Especies. Parc Subaquàtic. Tarragona. Societat d’Exploracions Submarines—Autoridad Portuaria de Tarragona. Available online: https://www.porttarragona.cat/es/catalogo-de-publicaciones/item/inventario-de-especies (accessed on 14 February 2024).
- Capdevila, M.; Folch, J. Fauna Malacològica Del Parc Subaquàtic de Tarragona (El Tarragonès, Catalunya, Espanya). Spira 2009, 3, 33–51. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. 2024. Available online: https://www.qgis.org (accessed on 20 January 2024).
- Kossuga, M.H.; Romminger, S.; Xavier, C.; Milanetto, M.C.; do Valle, M.Z.; Pimenta, E.F.; Morais, R.P.; de Carvalho, E.; Mizuno, C.M.; Coradello, L.F.C.; et al. Evaluating Methods for the Isolation of Marine-Derived Fungal Strains and Production of Bioactive Secondary Metabolites. Rev. Bras. Farm. 2012, 22, 257–267. [Google Scholar] [CrossRef]
- Wingfield, B.D.; Wingfield, M.J.; Duong, T.A. Molecular Basis of Cycloheximide Resistance in the Ophiostomatales Revealed. Curr. Genet. 2022, 68, 505–514. [Google Scholar] [CrossRef]
- Gannibal, P.B. Polyphasic Approach to Fungal Taxonomy. Biol. Bull. Rev. 2022, 12, 18–28. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen: London, UK, 1978; p. 256. [Google Scholar]
- Müller, F.M.C.; Werner, K.E.; Kasai, M.; Francesconi, A.; Chanock, S.J.; Walsh, T.J. Rapid Extraction of Genomic DNA from Medically Important Yeasts and Filamentous Fungi by High-Speed Cell Disruption. J. Clin. Microbiol. 1998, 36, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Mycological Society of America: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus Species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Aveskamp, M.M.; de Gruyter, J.; Spiers, A.G.; Crous, P.W. Multiple Didymella Teleomorphs Are Linked to the Phoma Clematidina Morphotype. Persoonia Mol. Phylogeny Evol. Fungi 2009, 22, 56–62. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E. Two Divergent Intragenomic RDNA ITS2 Types within a Monophyletic Lineage of the Fungusfusariumare Nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Stielow, J.B.; Lévesque, C.A.; Seifert, K.A.; Meyer, W.; Irinyi, L.; Smits, D.; Renfurm, R.; Verkley, G.J.M.; Groenewald, M.; Chaduli, D.; et al. One Fungus, Which Genes? Development and Assessment of Universal Primers for Potential Secondary Fungal DNA Barcodes. Persoonia Mol. Phylogeny Evol. Fungi 2015, 35, 242–263. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the Consolidated Species Concept to Resolve Species in the Teratosphaeriaceae. Persoonia Mol. Phylogeny Evol. Fungi 2014, 33, 1–40. [Google Scholar] [CrossRef]
- Torres-Garcia, D.; García, D.; Réblová, M.; Jurjević, Ž.; Hubka, V.; Gené, J. Diversity and Novel Lineages of Black Yeasts in Chaetothyriales from Freshwater Sediments in Spain. Persoonia Mol. Phylogeny Evol. Fungi 2023, 51, 194–228. [Google Scholar] [CrossRef]
- Giraldo, A.; Crous, P.W. Inside Plectosphaerellaceae. Stud. Mycol. 2019, 92, 227–286. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Miller, A.N.; Cano-Lira, J.F.; Guarro, J.; García, D.; Stadler, M.; Huhndorf, S.M.; Stchigel, A.M. Re-Evaluation of the Order Sordariales: Delimitation of Lasiosphaeriaceae s. str., and Introduction of the New Families Diplogelasinosporaceae, Naviculisporaceae, and Schizotheciaceae. Microorganisms 2020, 8, 1430. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Miller, A.N. Corrections to Recent Changes in the Taxonomy of the Sordariales. Mycol. Progr. 2022, 21, 69. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway: Enabling High-Impact Science for Phylogenetics Researchers with Limited Resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the eXtreme to the Campus and Beyond, New York, NY, USA, 16 July 2012; Association for Computing Machinery: New York, NY, USA, 2012; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Hespanhol, L.; Vallio, C.S.; Costa, L.M.; Saragiotto, B.T. Understanding and Interpreting Confidence and Credible Intervals around Effect Estimates. Braz. J. Phys. Ther. 2019, 23, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- de Hoog, G.S.; Vicente, V.A.; Najafzadeh, M.J.; Harrak, M.J.; Badali, H.; Seyedmousavi, S. Waterborne Exophiala Species Causing Disease in Cold-Blooded Animals. Persoonia Mol. Phylogeny Evol. Fungi 2011, 27, 46–72. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, M.R.; Padhye, A. A Exophiala jeanselmei, a New Combination for Phialophora jeanselmei. Mycotaxon 1977, 5, 341–352. [Google Scholar]
- Vitale, R.G.; Hoog, G.S. de Molecular Diversity, New Species and Antifungal Susceptibilities in the Exophiala spinifera Clade. Med. Mycol. 2002, 40, 545–556. [Google Scholar] [CrossRef] [PubMed]
- de Hoog, G.S.; Vicente, V.; Caligiorne, R.B.; Kantarcioglu, S.; Tintelnot, K.; Gerrits van den Ende, A.H.G.; Haase, G. Species Diversity and Polymorphism in the Exophiala spinifera Clade Containing Opportunistic Black Yeast-Like Fungi. J. Clin. Microbiol. 2003, 41, 4767–4778. [Google Scholar] [CrossRef]
- Yang, X.; Feng, M.; Yu, Z. Exophiala pseudooligosperma sp. nov., a Novel Black Yeast from Soil in Southern China. Int. J. Syst. Evol. Microbiol. 2021, 71, 5116. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.Y.; Ngan, A.H.Y.; Tsang, C.C.C.; Ling, I.W.H.; Chan, J.F.W.; Leung, S.-Y.; Yuen, K.-Y.; Lau, S.K.P. Clinical Spectrum of Exophiala Infections and a Novel Exophiala Species, Exophiala hongkongensis. J. Clin. Microbiol. 2020, 51, 260–267. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi, 4th ed.; Foundation Atlas of Clinical Fungi: Hilversum, The Netherlands, 2020. [Google Scholar]
- Weisenborn, J.L.; Kirschner, R.; Piepenbring, M.A. New Darkly Pigmented and Keratinolytic Species of Acremonium (Hyphomycetes) with Relationship to the Plectosphaerellaceae from Human Skin and Nail Lesions in Panama. Nova Hedwig. 2010, 90, 457–468. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z.; Stone, J.K. Penidiella kurandae. Fungal Planet 2007, 16. [Google Scholar]
- Taxonomy of Fungi Imperfecti: Proceedings of the First International Specialists’ Workshop Conference on Criteria and Terminology in the Classification of Fungi Imperfecti, Kananaskis, Alberta, Canada. In Taxonomy of Fungi Imperfecti; University of Toronto Press: Toronto, ON, Canada, 1971; ISBN 978-1-4875-8916-5.
- Udagawa, S.I.; Muroi, T. Notes on some Japanese ascomycetes XVI. Nippon Kingakukai Kaiho 1981, 22, 11–26. [Google Scholar]
- Rédou, V.; Navarri, M.; Meslet-Cladière, L.; Barbier, G.; Burgaud, G. Species Richness and Adaptation of Marine Fungi from Deep-Subseafloor Sediments. Appl. Environ. Microbiol. 2015, 81, 3571–3583. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Revealing the Hidden Diversity of Marine Fungi in Portugal with the Description of Two Novel Species, Neoascochyta Fuci Sp. Nov. and Paraconiothyrium Salinum Sp. Nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 5337–5354. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Xu, W.; Luo, Z.-H.; Pang, K.-L. Diversity and Temperature Adaptability of Cultivable Fungi in Marine Sediments from the Chukchi Sea. Bot. Mar. 2020, 63, 197–207. [Google Scholar] [CrossRef]
- Garmendia, G.; Alvarez, A.; Villarreal, R.; Martinez Silveira, A.; Wisniewski, M.; Vero, S. Fungal Diversity in the Coastal Waters of King George Island (Maritime Antarctica). World J. Microbiol. Biotechnol. 2021, 37, 142. [Google Scholar] [CrossRef]
- Morales, S.E.; Biswas, A.; Herndl, G.J.; Baltar, F. Global Structuring of Phylogenetic and Functional Diversity of Pelagic Fungi by Depth and Temperature. Front. Mar. Sci. 2019, 6, 131. [Google Scholar] [CrossRef]
- Tisthammer, K.H.; Cobian, G.M.; Amend, A.S. Global Biogeography of Marine Fungi Is Shaped by the Environment. Fungal Ecol. 2016, 19, 39–46. [Google Scholar] [CrossRef]
- Li, W.; Wang, M.; Burgaud, G.; Yu, H.; Cai, L. Fungal Community Composition and Potential Depth-Related Driving Factors Impacting Distribution Pattern and Trophic Modes from Epi- to Abyssopelagic Zones of the Western Pacific Ocean. Microb. Ecol. 2019, 78, 820–831. [Google Scholar] [CrossRef] [PubMed]
- Baltar, F.; Zhao, Z.; Herndl, G.J. Potential and Expression of Carbohydrate Utilization by Marine Fungi in the Global Ocean. Microbiome 2021, 9, 106. [Google Scholar] [CrossRef]
- Nicoletti, R.; Trincone, A. Bioactive Compounds Produced by Strains of Penicillium and Talaromyces of Marine Origin. Mar. Drugs 2016, 14, 37. [Google Scholar] [CrossRef]
- Wang, M.-M.; Yang, S.-Y.; Li, Q.; Zheng, Y.; Ma, H.-H.; Tu, Y.-H.; Li, W.; Cai, L. Microascaceae from the Marine Environment, with Descriptions of Six New Species. J. Fungi 2024, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Guarro, J.; Punsola, L.; Cano, J. Byssoonygena ceratinophila, gen. et sp. nov. a New Keratinophilic Fungus from Spain. Mycopathologia 1987, 100, 159–161. [Google Scholar] [CrossRef]
- Thaxter, R. On Certain Peculiar Fungus-Parasites of Living Insects. Bot. Gaz. 1914, 58, 235–253. [Google Scholar] [CrossRef]
- Blackwell, M.; Haelewaters, D.; Pfister, D.H. Laboulbeniomycetes: Evolution, Natural History, and Thaxter’s Final Word. Mycologia 2020, 112, 1048–1059. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Guo, S.; Gong, L.-F.; He, G.; Pang, K.-L.; Luo, Z.-H. Cultivable Fungal Diversity in Deep-Sea Sediment of the East Pacific Ocean. Geomicrobiol. J. 2018, 35, 790–797. [Google Scholar] [CrossRef]
- Wang, X.W.; Han, P.J.; Bai, F.Y.; Luo, A.; Bensch, K.; Meijer, M.; Kraak, B.; Han, D.Y.; Sun, B.D.; Crous, P.W.; et al. Taxonomy, Phylogeny and Identification of Chaetomiaceae with Emphasis on Thermophilic Species. Stud. Mycol. 2022, 101, 121–243. [Google Scholar] [CrossRef] [PubMed]
- Gladfelter, A.S.; James, T.Y.; Amend, A.S. Marine Fungi. Curr. Biol. 2019, 29, R191–R195. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Marine Fungi: Opportunities and Challenges. Encyclopedia 2022, 2, 559–577. [Google Scholar] [CrossRef]
- Ekanayaka, A.H.; Tibpromma, S.; Dai, D.; Xu, R.; Suwannarach, N.; Stephenson, S.L.; Dao, C.; Karunarathna, S.C. A Review of the Fungi That Degrade Plastic. J. Fungi 2022, 8, 772. [Google Scholar] [CrossRef]
- Guerra-Mateo, D.; Gené, J.; Baulin, V.; Cano-Lira, J.F. Phylogeny and Taxonomy of the Genus Amphichorda (Bionectriaceae): An Update on Beauveria-like Strains and Description of a Novel Species from Marine Sediments. Diversity 2023, 15, 795. [Google Scholar] [CrossRef]
- Bonugli-Santos, R.C.; dos Santos Vasconcelos, M.R.; Passarini, M.R.Z.; Vieira, G.A.L.; Lopes, V.C.P.; Mainardi, P.H.; dos Santos, J.A.; de Azevedo Duarte, L.; Otero, I.V.R.; da Silva Yoshida, A.M.; et al. Marine-Derived Fungi: Diversity of Enzymes and Biotechnological Applications. Front. Microbiol. 2015, 6, 269. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sarma, V.V.; Thambugala, K.M.; Huang, J.-J.; Li, X.-Y.; Hao, G.-F. Ecology and Evolution of Marine Fungi with Their Adaptation to Climate Change. Front. Microbiol. 2021, 12, 719000. [Google Scholar] [CrossRef] [PubMed]
- Zeghal, E.; Vaksmaa, A.; Vielfaure, H.; Boekhout, T.; Niemann, H. The Potential Role of Marine Fungi in Plastic Degradation—A Review. Front. Mar. Sci. 2021, 8, 738877. [Google Scholar] [CrossRef]
- Barnes, N.M.; Damare, S.R.; Bhatawadekar, V.C.; Garg, A.; Lotlikar, N.P. Degradation of Crude Oil-Associated Polycyclic Aromatic Hydrocarbons by Marine-Derived Fungi. 3 Biotech. 2023, 13, 335. [Google Scholar] [CrossRef] [PubMed]
- Vaksmaa, A.; Polerecky, L.; Dombrowski, N.; Kienhuis, M.V.M.; Posthuma, I.; Gerritse, J.; Boekhout, T.; Niemann, H. Polyethylene Degradation and Assimilation by the Marine Yeast Rhodotorula mucilaginosa. ISME Commun. 2023, 3, 68. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Zeng, J.S.; Harrak, M.J.; Sutton, D.A. Exophiala Xenobiotica Sp. Nov., an Opportunistic Black Yeast Inhabiting Environments Rich in Hydrocarbons. Antonie Van Leeuwenhoek 2006, 90, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Isola, D.; Zucconi, L.; Onofri, S.; Caneva, G.; de Hoog, G.S.; Selbmann, L. Extremotolerant Rock Inhabiting Black Fungi from Italian Monumental Sites. Fungal Divers. 2016, 76, 75–96. [Google Scholar] [CrossRef]
- Madrid, H.; Hernández-Restrepo, M.; Gené, J.; Cano, J.; Guarro, J.; Silva, V. New and Interesting Chaetothyrialean Fungi from Spain. Mycol. Progr. 2016, 15, 1179–1201. [Google Scholar] [CrossRef]
- Zeng, J.S.; Sutton, D.A.; Fothergill, A.W.; Rinaldi, M.G.; Harrak, M.J.; de Hoog, G.S. Spectrum of Clinically Relevant Exophiala Species in the United States. J. Clini. Microbiol. 2007, 45, 3713–3720. [Google Scholar] [CrossRef]
- Zeng, J.; Feng, P.; van den Ende, A.H.G.G.; Xi, L.; Harrak, M.J.; de Hoog, G.S. Multilocus Analysis of the Exophiala jeanselmei Clade Containing Black Yeasts Involved in Opportunistic Disease in Humans. Fungal Divers. 2014, 65, 3–16. [Google Scholar] [CrossRef]
- Seyedmousavi, S.; Guillot, J.; de Hoog, G.S. Phaeohyphomycoses, Emerging Opportunistic Diseases in Animals. Clin. Microbiol. Rev. 2013, 26, 19–35. [Google Scholar] [CrossRef]
- Thitla, T.; Kumla, J.; Khuna, S.; Lumyong, S.; Suwannarach, N. Species Diversity, Distribution, and Phylogeny of Exophiala with the Addition of Four New Species from Thailand. J. Fungi 2022, 8, 766. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding Needles in Haystacks: Linking Scientific Names, Reference Specimens and Molecular Data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.M.; Fraser, M.; Szekely, A.; Larcombe, D.E.; Johnson, E.M. Rapid Identification of Clinically Relevant Members of the Genus Exophiala by Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry and Description of Two Novel Species, Exophiala campbellii and Exophiala lavatrina. J. Clin. Microbiol. 2017, 55, 1162–1176. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.M.; Fraser, M.; Schilling, W.; Jones, G.; Pearl, R.; Linton, C.J.; Johnson, E.M. Exophiala campbellii Causing a Subcutaneous Palmar Cyst in an Otherwise Healthy UK Resident. Med. Mycol. Case Rep. 2020, 29, 43–45. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Rudramurthy, S.M.; Padhye, A.A.; Hemashetter, B.M.; Iyer, R.; Hallur, V.; Sharma, A.; Agnihotri, S.; Gupta, S.; Ghosh, A.; et al. Clinical Spectrum, Molecular Characterization, Antifungal Susceptibility Testing of Exophiala spp. From India and Description of a Novel Exophiala Species, E. arunalokei sp. Nov. Front. Cell. Infect. Microbiol. 2021, 11, 686120. [Google Scholar] [CrossRef] [PubMed]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal Diversity Notes 840–928: Micro-Fungi Associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Yong, L.K.; Wiederhold, N.P.; Sutton, D.A.; Sandoval-Denis, M.; Lindner, J.R.; Fan, H.; Sanders, C.; Guarro, J. Morphological and Molecular Characterization of Exophiala polymorpha Sp. Nov. Isolated from Sporotrichoid Lymphocutaneous Lesions in a Patient with Myasthenia Gravis. J. Clin. Microbiol. 2015, 53, 2816–2822. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Moreno, L.F.; Stielow, B.J.; Muszewska, A.; Hainaut, M.; Gonzaga, L.; Abouelleil, A.; Patané, J.S.L.; Priest, M.; Souza, R.; et al. Exploring the Genomic Diversity of Black Yeasts and Relatives (Chaetothyriales, Ascomycota). Stud. Mycol. 2017, 86, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Thangavel, R.; Wingfield, M.J.; Noordeloos, M.E.; Dima, B.; Brandrud, T.E.; Jansen, G.M.; et al. Fungal Planet Description Sheets: 1182–1283. Persoonia Mol. Phylogeny Evol. Fungi 2021, 46, 313–528. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-Scale Generation and Analysis of Filamentous Fungal DNA Barcodes Boosts Coverage for Kingdom Fungi and Reveals Thresholds for Fungal Species and Higher Taxon Delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Gueidan, C.; Villaseñor, C.R.; de Hoog, G.S.; Gorbushina, A.A.; Untereiner, W.A.; Lutzoni, F. A Rock-Inhabiting Ancestor for Mutualistic and Pathogen-Rich Fungal Lineages. Stud. Mycol. 2008, 61, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Irinyi, L.; Serena, C.; Garcia-Hermoso, D.; Arabatzis, M.; Desnos-Ollivier, M.; Vu, D.; Cardinali, G.; Arthur, I.; Normand, A.-C.; Giraldo, A.; et al. International Society of Human and Animal Mycology (ISHAM)-ITS Reference DNA Barcoding Database—The Quality Controlled Standard Tool for Routine Identification of Human and Animal Pathogenic Fungi. Med. Mycol. 2015, 53, 313–337. [Google Scholar] [CrossRef] [PubMed]
- Arzanlou, M.; Groenewald, J.Z.; Gams, W.; Braun, U.; Shin, H.-D.; Crous, P.W. Phylogenetic and Morphotaxonomic Revision of Ramichloridium and Allied Genera. Stud. Mycol. 2007, 58, 57–93. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.-C.; Tang, J.Y.M.; Chan, K.-F.; Lee, C.-Y.; Chan, J.F.W.; Ngan, A.H.Y.; Cheung, M.; Lau, E.C.L.; Li, X.; Ng, R.H.Y.; et al. Diversity of Phenotypically Non-Dermatophyte, Non-Aspergillus Filamentous Fungi Causing Nail Infections: Importance of Accurate Identification and Antifungal Susceptibility Testing. Emerg. Microbes Infect. 2019, 8, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Su, L.; Yang, S.; Sun, J.; Liu, B.; Fu, R.; Wu, B.; Liu, X.; Cai, L.; Guo, L.; et al. Unveiling the Hidden Diversity of Rock-Inhabiting Fungi: Chaetothyriales from China. J. Fungi 2020, 6, 187. [Google Scholar] [CrossRef]
- Moussa, T.A.A.; Al-Zahrani, H.S.; Kadasa, N.M.S.; Moreno, L.F.; Gerrits van den Ende, A.H.G.; de Hoog, G.S.; Al-Hatmi, A.M.S. Nomenclatural Notes on Nadsoniella and the Human Opportunist Black Yeast Genus Exophiala. Mycoses 2017, 60, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, M.M.F.; Vicente, V.A.; Bittencourt, J.V.M.; Gelinski, J.M.L.; Prenafeta-Boldú, F.X.; Romero-Güiza, M.; Fornari, G.; Gomes, R.R.; Santos, G.D.; Gerrits Van Den Ende, A.H.G.; et al. Diversity of Opportunistic Black Fungi on Babassu Coconut Shells, a Rich Source of Esters and Hydrocarbons. Fungal Biol. 2017, 121, 488–500. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Cheewangkoon, R.; van der Bank, M.; Swart, W.J.; Stchigel, A.M.; Cano-Lira, J.F.; Roux, J.; Madrid, H.; et al. Fungal Planet Description Sheets: 154–213. Persoonia Mol. Phylogeny Evol. Fungi 2013, 31, 188–296. [Google Scholar] [CrossRef] [PubMed]
- Kiyuna, T.; An, K.-D.; Kigawa, R.; Sano, C.; Miura, S.; Sugiyama, J. Molecular Assessment of Fungi in “Black Spots” That Deface Murals in the Takamatsuzuka and Kitora Tumuli in Japan: Acremonium Sect. Gliomastix Including Acremonium tumulicola Sp. Nov. and Acremonium felinum Comb. Nov. Mycoscience 2011, 52, 1–17. [Google Scholar] [CrossRef]
- Réblová, M.; Hernández-Restrepo, M.; Fournier, J.; Nekvindová, J. New Insights into the Systematics of Bactrodesmium and Its Allies and Introducing New Genera, Species and Morphological Patterns in the Pleurotheciales and Savoryellales (Sordariomycetes). Stud. Mycol. 2020, 95, 415–466. [Google Scholar] [CrossRef]
- Crous, P.W.; Braun, U.; Groenewald, J.Z. Mycosphaerella Is Polyphyletic. Stud. Mycol. 2007, 58, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; de Hoog, G.S.; Isola, D.; Onofri, S.; Quaedvlieg, W.; de Vries, M.; Verkley, G.J.M.; Stielow, J.B.; Zucconi, L.; Selbmann, L. Phylogeny and Taxonomy of Meristematic Rock-Inhabiting Black Fungi in the Dothideomycetes Based on Multi-Locus Phylogenies. Fungal Divers. 2014, 65, 127–165. [Google Scholar] [CrossRef]
- Crous, P.W.; Groenewald, J.Z. Why Everlastings Don’t Last. Persoonia Mol. Phylogeny Evol. Fungi 2011, 26, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.A.; Ewaze, J.O.; Summerbell, R.C.; Arocha-Rosete, Y.; Maharaj, A.; Guardiola, Y.; Saleh, M.; Wong, B.; Bogale, M.; O’Hara, M.J.; et al. Multilocus DnA Sequencing of the Whiskey Fungus Reveals a Continental-Scale Speciation Pattern. Persoonia Mol. Phylogeny Evol. Fungi 2016, 37, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet Description Sheets: 868–950. Persoonia Mol. Phylogeny Evol. Fungi 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Hunter, G.C.; Wingfield, B.D.; Crous, P.W.; Wingfield, M.J. A Multi-Gene Phylogeny for Species of Mycosphaerella Occurring on Eucalyptus Leaves. Stud. Mycol. 2006, 55, 147–161. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.A.; Carnegie, A.J.; Wingfield, M.J.; Hunter, G.C.; Burgess, T.I.; Andjic, V.; Barber, P.A.; Groenewald, J.Z. Unravelling Mycosphaerella: Do You Believe in Genera? Persoonia Mol. Phylogeny Evol. Fungi 2009, 23, 99–118. [Google Scholar] [CrossRef]
- Wang, X.W.; Yang, F.Y.; Meijer, M.; Kraak, B.; Sun, B.D.; Jiang, Y.L.; Wu, Y.M.; Bai, F.Y.; Seifert, K.A.; Crous, P.W.; et al. Redefining Humicola Sensu Stricto and Related Genera in the Chaetomiaceae. Stud. Mycol. 2019, 93, 65–153. [Google Scholar] [CrossRef]
- Miller, A.N.; Huhndorf, S.M. Multi-Gene Phylogenies Indicate Ascomal Wall Morphology Is a Better Predictor of Phylogenetic Relationships than Ascospore Morphology in the Sordariales (Ascomycota, Fungi). Mol. Phylogen. Evol. 2005, 35, 60–75. [Google Scholar] [CrossRef]
- Dai, D.-Q.; Phookamsak, R.; Wijayawardene, N.; Li, W.; Bhat, D.J.; Xu, J.; Taylor, J.; Hyde, K.; Chukeatirote, E. Bambusicolous Fungi. Fungal Divers. 2016, 82, 1–105. [Google Scholar] [CrossRef]
- Cai, L.; Jeewon, R.; Hyde, K.D. Phylogenetic Investigations of Sordariaceae Based on Multiple Gene Sequences and Morphology. Mycol. Res. 2006, 110, 137–150. [Google Scholar] [CrossRef]
- Cai, L.; Jeewon, R.; Hyde, K.D. Phylogenetic Evaluation and Taxonomic Revision of Schizothecium Based on Ribosomal DNA and Protein Coding Genes. Fungal Divers. 2005, 19, 1–21. [Google Scholar]
- Kruys, Å.; Huhndorf, S.M.; Miller, A.N. Coprophilous Contributions to the Phylogeny of Lasiosphaeriaceae and Allied Taxa within Sordariales (Ascomycota, Fungi). Fungal Divers. 2015, 70, 101–113. [Google Scholar] [CrossRef]
- Miller, A.N.; Huhndorf, S.M. Using Phylogenetic Species Recognition to Delimit Species Boundaries within Lasiosphaeria. Mycologia 2004, 96, 1106–1127. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Houbraken, J.; Groenewald, J.Z.; Meijer, M.; Andersen, B.; Nielsen, K.F.; Crous, P.W.; Samson, R.A. Diversity and Taxonomy of Chaetomium and Chaetomium-like Fungi from Indoor Environments. Stud. Mycol. 2016, 84, 145–224. [Google Scholar] [CrossRef]
- Huhndorf, S.M.; Miller, A.N.; Fernández, F.A. Molecular Systematics of the Sordariales: The Order and the Family Lasiosphaeriaceae Redefined. Mycologia 2004, 96, 368–387. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Mahoney, D.; Debuchy, R. Podospora Bullata, a New Homothallic Ascomycete from Kangaroo Dung in Australia. Available online: https://ascomycete.org/Journal/Article/art-0180 (accessed on 14 February 2024).
- Harms, K.; Milic, A.; Stchigel, A.M.; Stadler, M.; Surup, F.; Marin-Felix, Y. Three New Derivatives of Zopfinol from Pseudorhypophila mangenotii Gen. et Comb. Nov. J. Fungi 2021, 7, 181. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primer Pairs | Taxa | Annealing Temp. (°C) | Orientation | Primer Reference |
|---|---|---|---|---|---|
| ITS–LSU (LSU D1-D3 region) | ITS5 | Every strain | 53 | Forward | [36] |
| LR5 | Reverse | [37] | |||
| β-tubulin (tub2) | Bt2a | Aspergillaceae, Bionectriaceae, Microascaceae, Trichocomaceae | 56 | Forward | [38] |
| Bt2b | Reverse | ||||
| Btub2f | Trichocomaceae | 56 | Forward | [39] | |
| Btub4R | Reverse | ||||
| T1 | Chaetomiaceae, Schizotheciaceae | 56 | Forward | [40] | |
| Btub4R | Reverse | [39] | |||
| Traslation elongation factor-1α (tef1-α) | EF-983F | Bionectriaceae, Plectosphaerellaceae | 57 | Forward | [41] |
| EF-2218R | Reverse | ||||
| EF-728F | Cladosporiaceae, Nectriaceae | 57 | Forward | [42] | |
| EF-986R | Reverse | ||||
| RNA polymerase II second largest subunit (rpb2) | RPB2-5F | Bionectriaceae, Chaetomiaceae, Cucurbitariaceae, Plectosphaerellaceae, Schizotheciaceae, Thyridariaceae | 60 | Forward | [43] |
| RPB2-7R | Reverse | ||||
| SSU | NS1 | Incertae sedis | 53 | Forward | [36] |
| NS4 | Reverse |
| Dataset | Parameters | Exophiala | Nigrocephalum | Queenslandipenidiella | Schizochlamydosporiella |
|---|---|---|---|---|---|
| ITS | Lenth (bp) | 625 | 511 | 529 | 545 |
| Conserved sites | 382 | 374 | 284 | 376 | |
| Variable sites | 226 | 130 | 240 | 159 | |
| Parsimony informative | 147 | 93 | 204 | 75 | |
| BI model | SYM+I+G | GTR+G | GTR+I+G | SYM+G | |
| LSU | Lenth (bp) | 873 | 791 | 751 | 827 |
| Conserved sites | 801 | 714 | 568 | 730 | |
| Variable sites | 60 | 77 | 180 | 94 | |
| Parsimony informative | 38 | 42 | 139 | 45 | |
| BI model | K80+I+G | K80+I | SYM+I+G | K80+I | |
| rpb2 | Lenth (bp) | - | 743 | - | 840 |
| Conserved sites | - | 484 | - | 460 | |
| Variable sites | - | 259 | - | 380 | |
| Parsimony informative | - | 219 | - | 223 | |
| BI model | - | GTR+I+G | - | GTR+I+G | |
| tef1-a | Lenth (bp) | - | 787 | - | - |
| Conserved sites | - | 624 | - | - | |
| Variable sites | - | 163 | - | - | |
| Parsimony informative | - | 132 | - | - | |
| BI model | - | GTR+G | - | - | |
| Concatenated | Lenth (bp) | 1498 | 2832 | 1280 | 2212 |
| Conserved sites | 1183 | 2196 | 852 | 1566 | |
| Variable sites | 286 | 629 | 420 | 633 | |
| Parsimony informative | 185 | 486 | 343 | 343 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra-Mateo, D.; Cano-Lira, J.F.; Fernández-Bravo, A.; Gené, J. Sunken Riches: Ascomycete Diversity in the Western Mediterranean Coast through Direct Plating and Flocculation, and Description of Four New Taxa. J. Fungi 2024, 10, 281. https://doi.org/10.3390/jof10040281
Guerra-Mateo D, Cano-Lira JF, Fernández-Bravo A, Gené J. Sunken Riches: Ascomycete Diversity in the Western Mediterranean Coast through Direct Plating and Flocculation, and Description of Four New Taxa. Journal of Fungi. 2024; 10(4):281. https://doi.org/10.3390/jof10040281
Chicago/Turabian StyleGuerra-Mateo, Daniel, José F. Cano-Lira, Ana Fernández-Bravo, and Josepa Gené. 2024. "Sunken Riches: Ascomycete Diversity in the Western Mediterranean Coast through Direct Plating and Flocculation, and Description of Four New Taxa" Journal of Fungi 10, no. 4: 281. https://doi.org/10.3390/jof10040281
APA StyleGuerra-Mateo, D., Cano-Lira, J. F., Fernández-Bravo, A., & Gené, J. (2024). Sunken Riches: Ascomycete Diversity in the Western Mediterranean Coast through Direct Plating and Flocculation, and Description of Four New Taxa. Journal of Fungi, 10(4), 281. https://doi.org/10.3390/jof10040281