
Production of Xylanase by Trichoderma Species Growing on Olive Mill Pomace and Barley Bran in a Packed-Bed Bioreactor


Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Species Isolates and Inoculum Preparation
2.2. Preparation of OMP
2.3. Flask Solid State Fermentation for Xylanase Production
2.4. Xylanase Assay
2.5. Assessment of Reducing Sugars
2.6. Total Carbohydrates
2.7. Optimization of Xylanase Production by Trichoderma sp.
2.8. Packed-Bed Column Solid State Fermentation of OMP and BB for Xylanase Production
2.9. Percentage of Degradation
2.10. Times Profile of the Process
2.11. Partial Purification of Xylanase
2.11.1. Ammonium Sulfate Precipitation and Dialysis
2.11.2. Size-Exclusion Chromatography
2.12. Statistical Analysis
3. Results
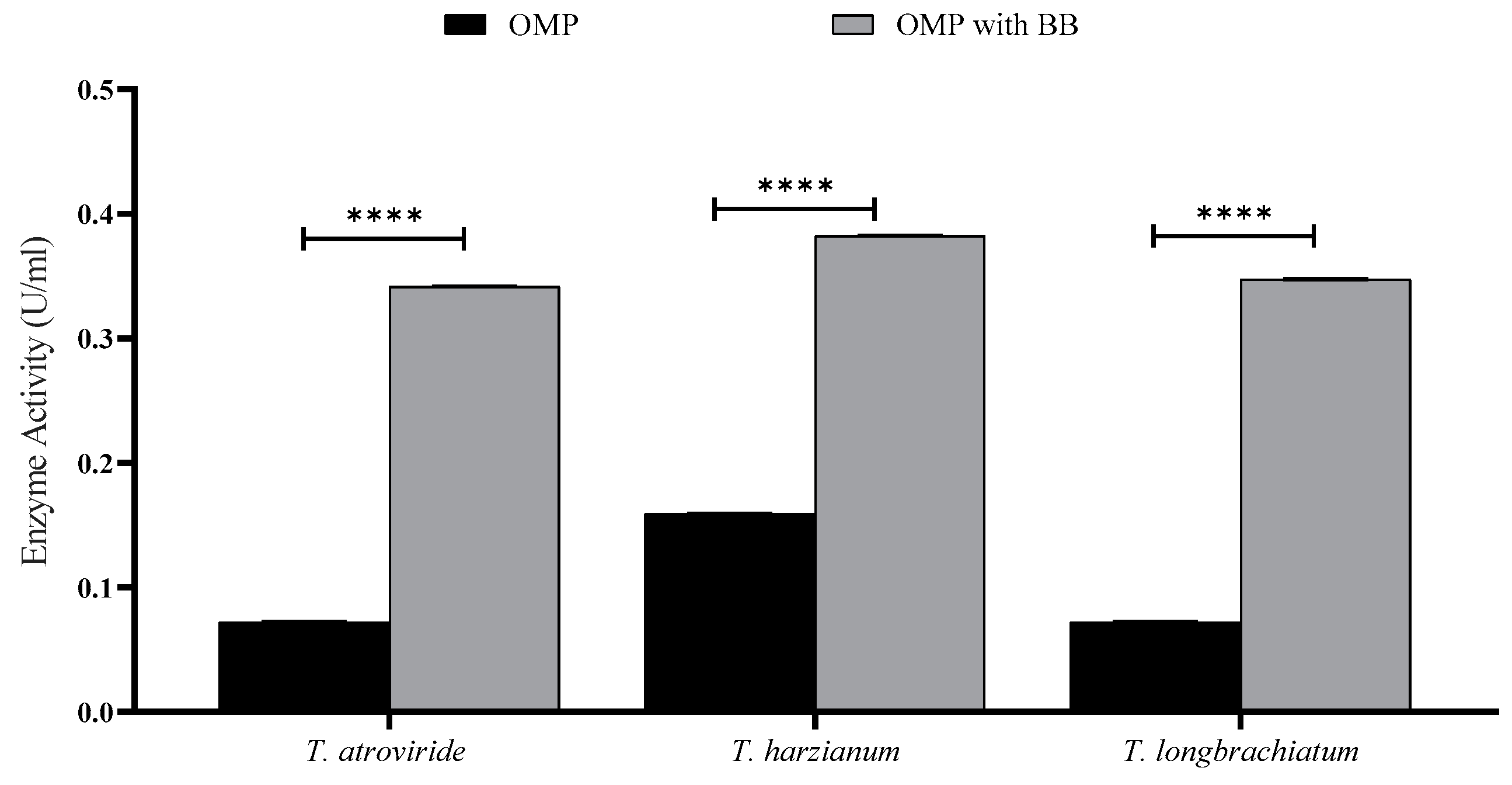
3.1. Flask Solid State Fermentation (SSF) for Xylanase Production by Trichoderma sp.
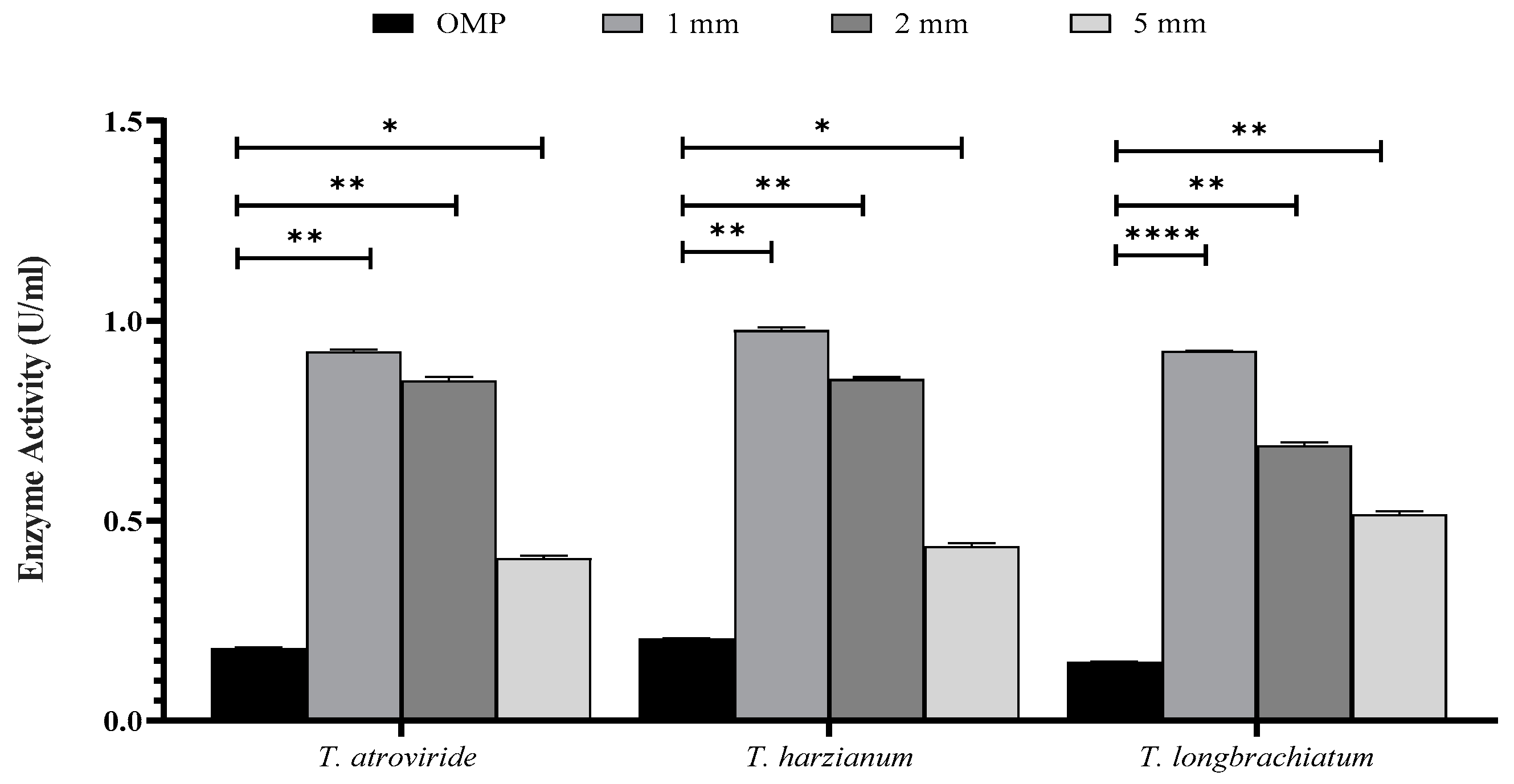
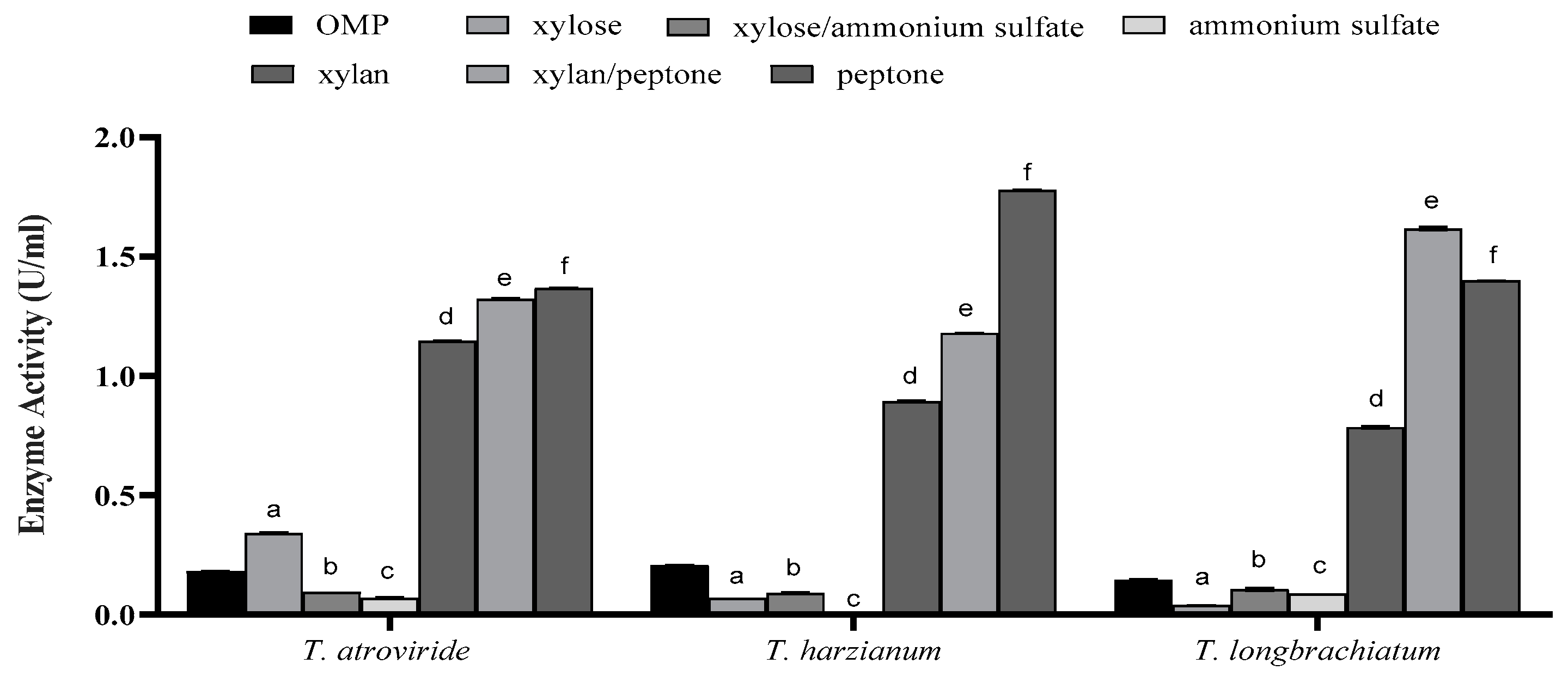
3.2. Optimization of SSF for Xylanase Production
3.3. Time Profile of the Process
3.4. Packed-Bed Column Solid State Fermentation
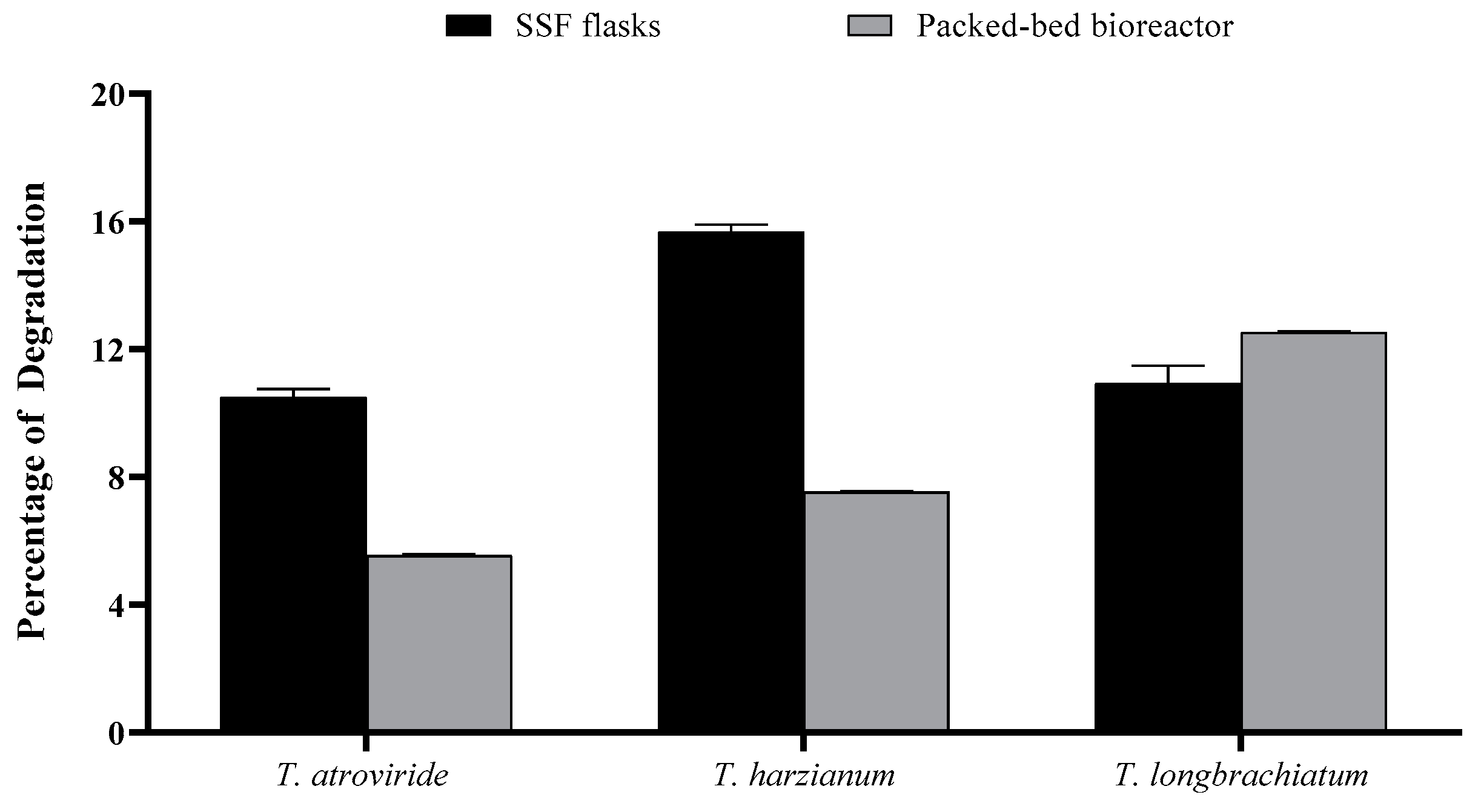
3.5. Percentage of OMP Degradation
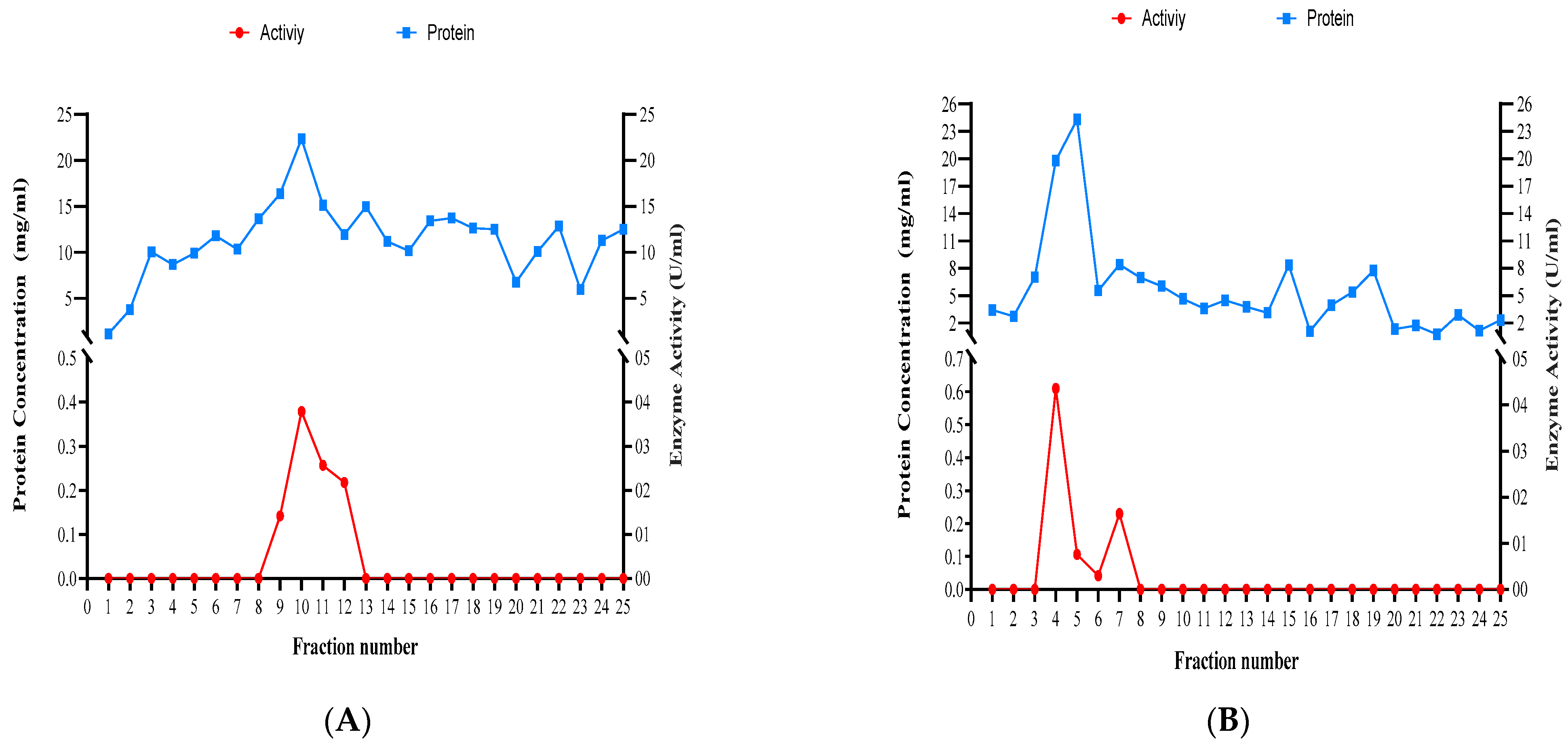
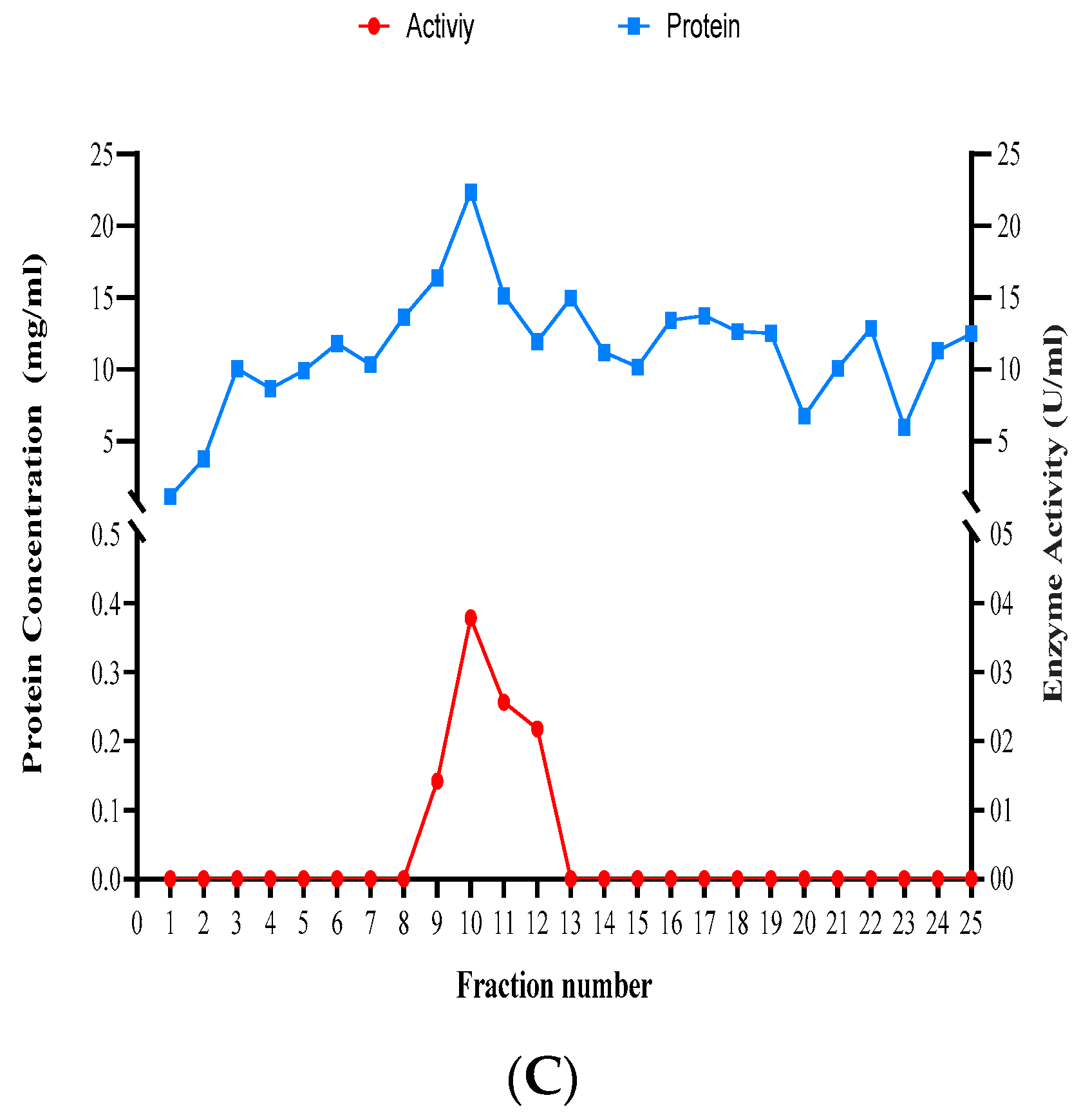
3.6. Partial Purification of Xylanase
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions | Trichoderma species | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T. atroviride | T. harzianum | T. longibrachiatum | ||||||||||
| OD280 | Protein mg/mL | Activity U/mL | Specific Enzyme Activity U/mg | OD280 | Protein mg/mL | Activity U/mL | Specific Enzyme Activity U/mg | OD280 | Protein mg/mL | Activity U/mL | Specific Enzyme Activity U/mg | |
| 1 | -0.035 | -1.750 | 0.000 | 0.000 | 0.068 | 3.410 | 0.000 | 0.000 | 0.006 | 0.300 | 0.000 | 0.000 |
| 2 | -0.030 | -1.510 | 0.000 | 0.000 | 0.055 | 2.735 | 0.000 | 0.000 | 0.009 | 0.450 | 0.000 | 0.000 |
| 3 | -0.001 | -0.050 | 0.000 | 0.000 | 0.140 | 7.015 | 0.000 | 0.000 | 0.065 | 3.250 | 0.000 | 0.000 |
| 4 | 0.005 | 0.260 | 0.000 | 0.000 | 0.396 | 19.820 | 0.610 | 0.031 | 0.149 | 7.440 | 0.000 | 0.000 |
| 5 | 0.013 | 0.640 | 0.000 | 0.000 | 0.487 | 24.325 | 0.106 | 0.004 | 0.212 | 10.595 | 0.000 | 0.000 |
| 6 | 0.070 | 3.486 | 0.000 | 0.000 | 0.111 | 5.550 | 0.042 | 0.007 | 0.213 | 10.670 | 0.000 | 0.000 |
| 7 | 0.016 | 0.796 | 0.000 | 0.000 | 0.168 | 8.415 | 0.230 | 0.027 | 0.300 | 15.005 | 0.000 | 0.000 |
| 8 | 0.035 | 1.750 | 0.000 | 0.000 | 0.140 | 6.980 | 0.000 | 0.000 | 0.309 | 15.440 | 0.000 | 0.000 |
| 9 | 0.159 | 7.950 | 0.511 | 0.064 | 0.121 | 6.040 | 0.000 | 0.000 | 0.397 | 19.840 | 0.059 | 0.003 |
| 10 | 0.200 | 10.005 | 0.093 | 0.009 | 0.093 | 4.635 | 0.000 | 0.000 | 0.485 | 24.235 | 0.469 | 0.019 |
| 11 | 0.350 | 17.500 | 0.106 | 0.006 | 0.072 | 3.600 | 0.000 | 0.000 | 0.391 | 19.540 | 0.140 | 0.007 |
| 12 | 0.456 | 22.780 | 0.041 | 0.002 | 0.090 | 4.475 | 0.000 | 0.000 | 0.286 | 14.315 | 0.250 | 0.017 |
| 13 | 0.388 | 19.400 | 0.000 | 0.000 | 0.076 | 3.775 | 0.000 | 0.000 | 0.209 | 10.425 | 0.597 | 0.057 |
| 14 | 0.300 | 15.000 | 0.000 | 0.000 | 0.062 | 3.120 | 0.000 | 0.000 | 0.244 | 12.220 | 0.050 | 0.004 |
| 15 | 0.298 | 14.900 | 0.000 | 0.000 | 0.167 | 8.345 | 0.000 | 0.000 | 0.304 | 15.205 | 0.000 | 0.000 |
| 16 | 0.266 | 13.300 | 0.000 | 0.000 | 0.022 | 1.120 | 0.000 | 0.000 | 0.300 | 15.005 | 0.000 | 0.000 |
| 17 | 0.255 | 12.750 | 0.000 | 0.000 | 0.079 | 3.935 | 0.000 | 0.000 | 0.223 | 11.130 | 0.000 | 0.000 |
| 18 | 0.250 | 12.500 | 0.000 | 0.000 | 0.108 | 5.390 | 0.000 | 0.000 | 0.270 | 13.480 | 0.000 | 0.000 |
| 19 | 0.203 | 10.165 | 0.000 | 0.000 | 0.155 | 7.745 | 0.000 | 0.000 | 0.257 | 12.830 | 0.000 | 0.000 |
| 20 | 0.100 | 5.005 | 0.000 | 0.000 | 0.007 | 0.345 | 0.000 | 0.000 | 0.270 | 13.500 | 0.000 | 0.000 |
| 21 | 0.125 | 6.250 | 0.000 | 0.000 | 0.034 | 1.715 | 0.000 | 0.000 | 0.200 | 10.005 | 0.000 | 0.000 |
| 22 | 0.200 | 10.000 | 0.000 | 0.000 | 0.015 | 0.765 | 0.000 | 0.000 | 0.190 | 9.480 | 0.000 | 0.000 |
| 23 | 0.100 | 5.010 | 0.000 | 0.000 | 0.058 | 2.885 | 0.000 | 0.000 | 0.156 | 7.775 | 0.000 | 0.000 |
| 24 | 0.109 | 5.450 | 0.000 | 0.000 | 0.023 | 1.170 | 0.000 | 0.000 | 0.098 | 4.910 | 0.000 | 0.000 |
| 25 | 0.100 | 5.000 | 0.000 | 0.000 | 0.047 | 2.335 | 0.000 | 0.000 | 0.085 | 4.260 | 0.000 | 0.000 |
References
- Sorrenti, V.; Burò, I.; Consoli, V.; Vanella, L. Recent advances in health benefits of bioactive compounds from food wastes and by-products: Biochemical aspects. Int. J. Mol. Sci. 2023, 24, 2019. [Google Scholar] [CrossRef] [PubMed]
- Knob, A.; Fortkamp, D.; Prolo, T.; Izidoro, S.C.; Almeida, J.M. Agro-residues as an alternative for xylanase production by filamentous fungi. BioResources 2014, 9, 5738–5773. [Google Scholar]
- Gontard, N.; Sonesson, U.; Birkved, M.; Majone, M.; Bolzonella, D.; Celli, A.; Angellier, H.; Jang, H.; Verniquet, A.; Brosze, J.; et al. A Research challenge vision regarding management of agricultural waste in circular bio-based economy. Crit. Rev. Environ. Sci. Technol. 2018, 48, 614–654. [Google Scholar] [CrossRef]
- Basit, A.; Jiang, W.; Rahim, K. Xylanase and its industrial applications. In Biotechnological Applications of Biomass; Basso, T.P., Basso, T.O., Basso, L.C., Eds.; Intechopen: London, UK, 2020; p. 638. [Google Scholar]
- Ocreto, J.B.; Chen, W.H.; Rollon, A.P.; Ong, H.C.; Pétrissans, A.; Pétrissans, M.; De Luna, M.D.G. Ionic liquid dissolution utilized for biomass conversion into biofuels, value-added chemicals and advanced materials: A comprehensive review. Chem. Eng. J. 2022, 445, 136733. [Google Scholar] [CrossRef]
- Rebouillat, S.; Pla, F. A Review: On smart materials based on some polysaccharides; within the contextual bigger data, insiders, “improvisation,” and said artificial intelligence trends. J. Biomater. Nanobiotech. 2019, 10, 41–77. [Google Scholar] [CrossRef]
- Kumar, A.; Naraian, R. Chapter 6—Differential expression of the microbial β-1,4-Xylanase and β-1,4-Endoglucanase genes. In New and Future Developments in Microbial Biotechnology and Bioengineering; Singh, H.B., Gupta, V.K., Jogaiah, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 95–111. [Google Scholar]
- Mardawati, E.; Sinurat, Y.; Yuliana, T. Production of crude xylanase from Trichoderma sp. using Reutealis trisperma exocarp substrate in solid state fermentation. In Proceedings of the International Conference of Sustainability Agriculture and Biosystem, Padang, Indonesia, 12–13 November 2019; IOP Publishing: Bristol, UK, 2020; p. 012024. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Kumar, B.; Verma, P. A Detailed overview of xylanases: An emerging biomolecule for current and future prospective. Bioresour. Bioprocess. 2019, 6, 40. [Google Scholar] [CrossRef]
- Dutta, P.D.; Neog, B.; Goswami, T. Xylanase enzyme production from Bacillus australimaris P5 for prebleaching of bamboo (Bambusa tulda) pulp. Mater. Chem. Phys. 2020, 243, 122227. [Google Scholar] [CrossRef]
- Fang, H.Y.; Chang, S.M.; Hsieh, M.C.; Fang, T.J. Production, Optimization growth conditions, and properties of the xylanase from Aspergillus carneus M34. J. Mol. Catal. B Enzym. 2007, 49, 36–42. [Google Scholar] [CrossRef]
- Rojas, L.F.; Zapata, P.; Ruiz-Tirado, L. Agro-industrial waste enzymes: Perspectives in circular economy. Curr. Opin. Green Sustain. Chem. 2022, 34, 100585. [Google Scholar] [CrossRef]
- Danso, B.; Ali, S.S.; Xie, R.; Sun, J. Valorization of wheat straw and bioethanol production by a novel xylanase- and cellulase-producing Streptomyces strain isolated from the wood-feeding termite, Microcerotermes Species. Fuel 2022, 310, 122333. [Google Scholar] [CrossRef]
- Bajpai, P. Xylanases. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Lederberg, J., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 600–612. [Google Scholar]
- Azzouz, Z.; Bettache, A.; Boucherba, N.; Amghar, Z.; Benallaoua, S. Optimization of xylanase production by newly isolated strain Trichoderma afroharzianum isolate AZ 12 in solid state fermentation using response surface methodology. Cellul. Chem. Technol. 2020, 54, 451–462. [Google Scholar] [CrossRef]
- Haddadin, M.S.; Haddadin, J.; Arabiyat, O.I.; Hattar, B. Biological conversion of olive pomace into compost by using Trichoderma harzianum and Phanerochaete chrysosporium. Bioresour. Technol. 2009, 100, 4773–4782. [Google Scholar] [CrossRef] [PubMed]
- Polizeli, M.; Rizzatti, A.; Monti, R.; Terenzi, H.; Jorge, J.A.; Amorim, D. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Abdelhadi, S.O.; Dosoretz, C.G.; Rytwo, G.; Gerchman, Y.; Azaizeh, H. Production of biochar from olive mill solid waste for heavy metal removal. Bioresour. Technol. 2017, 244, 759–767. [Google Scholar] [CrossRef]
- Khdair, A.I.; Abu-Rumman, G.; Khdair, S.I. Pollution estimation from olive mills wastewater in Jordan. Heliyon 2019, 5, e02386. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, K.; Nayak, R.; Shukla, A.; Parmar, P.; Goswami, D.; Saraf, M. Polyhydroxyalkanoates: An exotic gleam in the gloomy tale of plastics. J. Polym. Environ. 2021, 29, 2013–2032. [Google Scholar] [CrossRef]
- Abdollahi, F.; Jahadi, M.; Ghavami, M. Thermal stability of natural pigments produced by Monascus purpureus in submerged fermentation. Food Sci. Nutr. 2021, 9, 4855–4862. [Google Scholar] [CrossRef]
- Singh, A.; Shahid, M.; Srivastava, M.; Pandey, S.; Sharma, A.; Kumar, V. Optimal physical parameters for growth of Trichoderma species at varying pH, temperature, and agitation. Virol Mycol. 2014, 3, 1–7. [Google Scholar]
- Khanahmadi, M.; Arezi, I.; Amiri, M.-S.; Miranzadeh, M. Bioprocessing of agro-industrial residues for optimization of xylanase production by solid-state fermentation in flask and tray bioreactor. Biocatal. Agric. Biotechnol. 2018, 13, 272–282. [Google Scholar] [CrossRef]
- Al Sheikh, A.H.M. Utilization of Dry Mill Residues (DOR) for the Production of Xylanase Enzyme by Aspergillus terreus in Solid State Fermentation. Master’s Thesis, Hashemite University, Zarqa, Jordan, 2011. [Google Scholar]
- Assamoi, A.A.; Destain, J.; Delvigne, F.; Lognay, G.; Thonart, P. Solid-state fermentation of xylanase from Penicillium canescens 10-10c in a multi-layer-packed bed reactor. Appl. Biochem. Biotechnol. 2007, 145, 87–98. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Teixeira, R.S.; da Silva, A.S.; Ferreira-Leitao, V.S.; da Silva Bon, E.P. Amino acids interference on the quantification of reducing sugars by the 3,5-dinitrosalicylic acid assay mislead carbohydrase activity measurements. Carbohydr. Res. 2012, 363, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Ranjitha, P.; Karthy, E.; Mohankumar, A. Purification and characterization of the lipase from marine Vibrio fischeri. Int. J. Biol. 2009, 1, 48. [Google Scholar] [CrossRef]
- Saryono, S.; Novianty, R.; Suraya, N.; Piska, F.; Devi, S.; Pratiwi, N.W.; Ardhi, A. Molecular identification of cellulase-producing thermophilic fungi isolated from Sungai Pinang hot spring, Riau Province, Indonesia. Biodivers. J. 2022, 23. [Google Scholar] [CrossRef]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Pathania, S.; Sharma, N.; Verma, S.K. Optimization of cellulase-free xylanase produced by a potential thermoalkalophilic Paenibacillus sp. N1 isolated from hot springs of Northern Himalayas in India. J. Microbiol. Biotechnol. Food Sci. 2020, 9, 1–24. [Google Scholar]
- Oliveira, F.; Salgado, J.M.; Abrunhosa, L.; Pérez-Rodríguez, N.; Domínguez, J.M.; Venâncio, A.; Belo, I. Optimization of lipase production by solid-state fermentation of olive pomace: From flask to laboratory-scale packed-bed bioreactor. Bioprocess Biosyst. Eng. 2017, 40, 1123–1132. [Google Scholar] [CrossRef]
- Dhivahar, J.; Khusro, A.; Ahamad Paray, B.; Rehman, M.U.; Agastian, P. Production and partial purification of extracellular Xylanase from Pseudomonas nitroreducens using frugivorous bat (Pteropus giganteus) faeces as an ideal substrate and its role in poultry feed digestion. J. King Saud Univ. Sci. 2020, 32, 2474–2479. [Google Scholar] [CrossRef]
- Siddiqui, M.A.H.; Biswas, M.; Faruk, M.O.; Roy, M.; Asaduzzaman, A.K.M.; Sharma, S.C.D.; Biswas, T.; Roy, N. Optimization, Isolation and characterization of cellulase–free thermostable xylanase from Paenibacillus sp. Am. J. Life Sci. 2016, 4, 93–98. [Google Scholar] [CrossRef]
- Abusara, W.A. Beta-Glucosidase Enzyme Production in Solid-State Fermentation Using Olive Mill Pomace (OMP) and Yeast Industry Wastewater (YIW); MSc. Hashemite University: Zarqa, Jordan, 2019. [Google Scholar]
- Le Strat, Y.; Mandin, M.; Ruiz, N.; Robiou du Pont, T.; Ragueneau, E.; Barnett, A.; Déléris, P.; Dumay, J. Quantification of xylanolytic and cellulolytic activities of fungal strains isolated from Palmaria palmata to enhance R-phycoerythrin extraction of Palmaria palmata: From seaweed to seaweed. Mar. Drugs 2023, 21, 393. [Google Scholar] [CrossRef]
- Naeimi, S.; Khosravi, V.; Varga, A.; Vágvölgyi, C.; Kredics, L. Screening of organic substrates for solid-state fermentation, viability and bioefficacy of Trichoderma harzianum AS12-2, a biocontrol strain against rice sheath blight disease. J. Agron. 2020, 10, 1258. [Google Scholar] [CrossRef]
- Medouni-Haroune, L.; Zaidi, F.; Medouni-Adrar, S.; Kecha, M. Olive pomace: From an olive mill waste to a resource, an overview of the new treatments. J. Crit. Rev. 2018, 5, 1–6. [Google Scholar] [CrossRef]
- Boutiche, M.; Sahir-Halouane, F.; Meziant, L.; Boulaouad, I.; Saci, F.; Fiala, S.; Bekrar, A.; Mesbah, R.; Abdessemed, A. Characterization and valorization of olive pomace for production of cellulase from Trichoderma reesei RUT C30 in solid-state fermentation. Alger. J. Environ. Sci. Technol. 2020, 6, 1381–1387. [Google Scholar]
- Soliman, H.M.; Sherief, A.; EL-Tanash, A.B. Production of xylanase by Aspergillus niger and Trichoderma viride using some agricultural residues. Int. J. Agric. Res. 2012, 7, 46–57. [Google Scholar] [CrossRef]
- Lakshmi, G.S.; Rao, C.S.; Rao, R.S.; Hobbs, P.J.; Prakasham, R.S. Enhanced production of xylanase by a newly isolated Aspergillus terreus under solid state fermentation using palm industrial waste: A statistical optimization. Biochem. Eng. J. 2009, 48, 51–57. [Google Scholar] [CrossRef]
- Gautério, G.V.; da Silva, L.G.G.; Hübner, T.; da Rosa Ribeiro, T.; Kalil, S.J. Xylooligosaccharides production by crude and partially purified xylanase from Aureobasidium pullulans: Biochemical and thermodynamic properties of the enzymes and their application in xylan hydrolysis. Process Biochem. 2021, 104, 161–170. [Google Scholar] [CrossRef]
- Seyis, I.; Aksoz, N. Xylanase production from Trichoderma harzianum 1073 D3 with alternative carbon and nitrogen sources. Food Technol. Biotechnol. 2005, 43, 37–40. [Google Scholar]
- El-Gendi, H.; Badawy, A.S.; Bakhiet, E.K.; Rawway, M.; Ali, S.G. Valorization of lignocellulosic wastes for sustainable xylanase production from locally isolated Bacillus subtilis exploited for xylooligosaccharides’ production with potential antimicrobial activity. Arch. Microbiol. 2023, 205, 315. [Google Scholar] [CrossRef]
- Pandey, S.; Shahid, M.; Srivastava, M.; Sharma, A.; Singh, A.; Kumar, V.; Srivastava, Y. Isolation and optimized production of xylanase under solid state fermentation condition from Trichoderma sp. Int. J. Adv. Res. 2014, 2, 263–273. [Google Scholar]
- De los Santos-Villalobos, S.; Hernández-Rodríguez, L.E.; Villaseñor-Ortega, F.; Peña-Cabriales, J.J. Production of Trichoderma asperellum T8a spores by a” home-made” solid-state fermentation of mango industrial wastes. BioResources 2012, 7, 4938–4951. [Google Scholar] [CrossRef]
- Massadeh, M.I.; Fandi, K.; Al-Abeid, H.; Alsharafat, O.; Abu-Elteen, K. Production of citric acid by Aspergillus niger cultivated in olive mill wastewater using a two-stage packed column bioreactor. Fermentation 2022, 8, 153. [Google Scholar] [CrossRef]
- Silva, L.d.O.; Terrasan, C.R.F.; Carmona, E.C. Purification and characterization of xylanases from Trichoderma inhamatum. Electron. J. Biotechnol. 2015, 18, 307–313. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alananbeh, K.M.; Alkfoof, R.; Muhaidat, R.; Massadeh, M. Production of Xylanase by Trichoderma Species Growing on Olive Mill Pomace and Barley Bran in a Packed-Bed Bioreactor. J. Fungi 2024, 10, 49. https://doi.org/10.3390/jof10010049
Alananbeh KM, Alkfoof R, Muhaidat R, Massadeh M. Production of Xylanase by Trichoderma Species Growing on Olive Mill Pomace and Barley Bran in a Packed-Bed Bioreactor. Journal of Fungi. 2024; 10(1):49. https://doi.org/10.3390/jof10010049
Chicago/Turabian StyleAlananbeh, Kholoud M., Rana Alkfoof, Riyadh Muhaidat, and Muhannad Massadeh. 2024. "Production of Xylanase by Trichoderma Species Growing on Olive Mill Pomace and Barley Bran in a Packed-Bed Bioreactor" Journal of Fungi 10, no. 1: 49. https://doi.org/10.3390/jof10010049
APA StyleAlananbeh, K. M., Alkfoof, R., Muhaidat, R., & Massadeh, M. (2024). Production of Xylanase by Trichoderma Species Growing on Olive Mill Pomace and Barley Bran in a Packed-Bed Bioreactor. Journal of Fungi, 10(1), 49. https://doi.org/10.3390/jof10010049