
Design of a Multi-Epitope Vaccine against Histoplasma capsulatum through Immunoinformatics Approaches


 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Retrieval of Antigenic Sequences
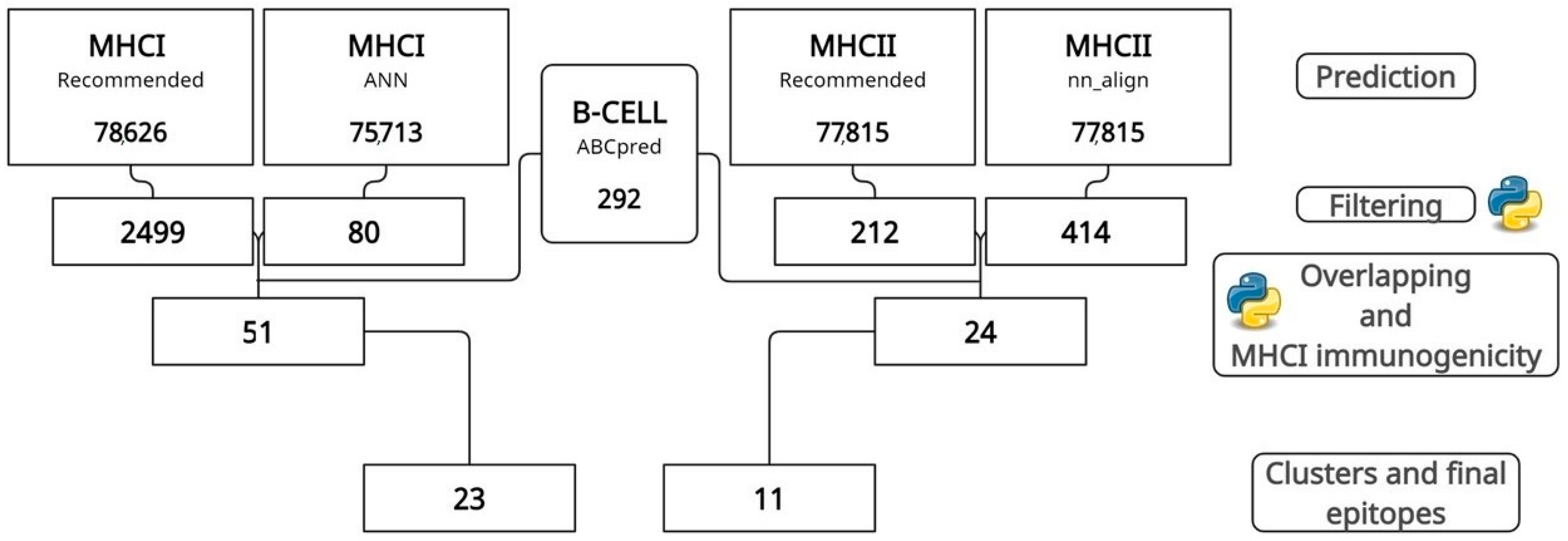
2.2. MHCI, MHCII, and B-Cell Epitope Prediction, Filtering, and Overlapping
2.3. Cluster Analysis, Epitope Conservation between Strains, and Population Coverage
2.4. Evaluation of the Immunogenicity of MHCI Epitopes for Dogs and Horses
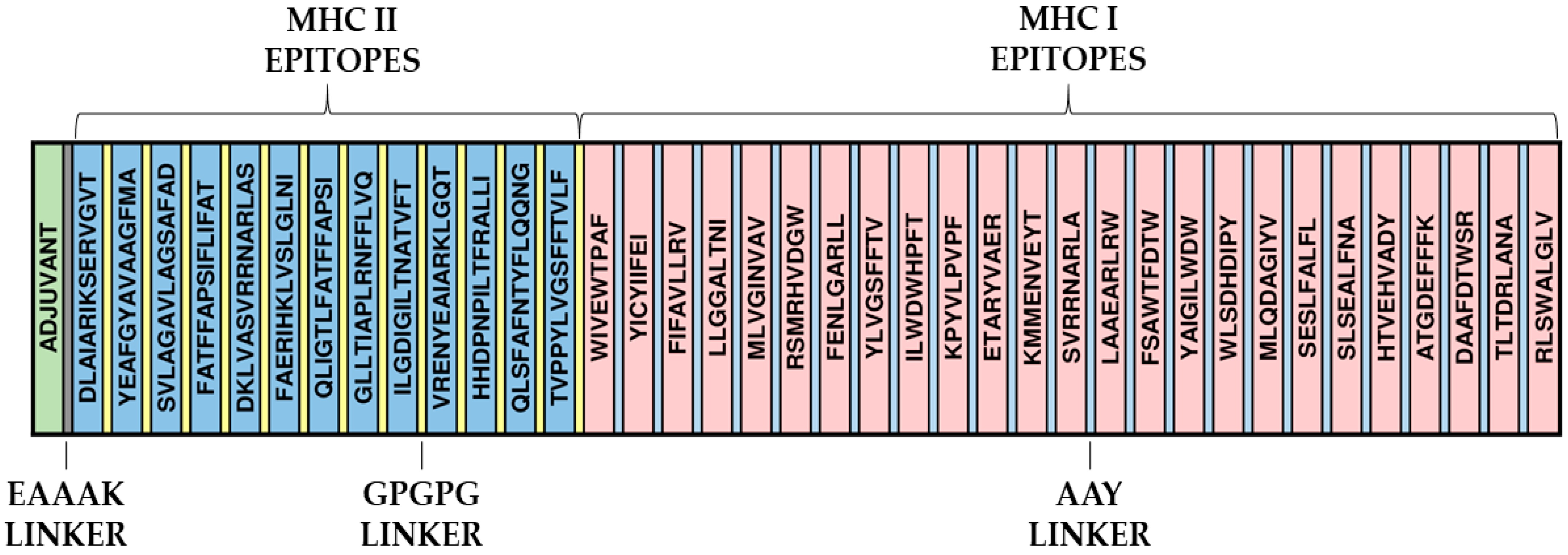
2.5. Construction of the Primary Structure of the Immunogen and Evaluation of Its Characteristics
2.6. Modeling the Secondary and Tertiary Structure of the Multi-Epitope Vaccine
2.7. Molecular Docking of the Multi-Epitope Vaccine with Innate Immune System Receptor TLR4
2.8. Prediction of Discontinuous B-Cell Epitopes
2.9. Simulation of the Immune Response of the Multi-Epitope Vaccine
2.10. Codon Adaptation and In Silico Cloning
3. Results
3.1. Prediction, Filtering, and Analysis of CTL, HTL, and B-Cell Epitopes
3.2. The Final Epitopes Are Shown to Be Highly Conserved among Histoplasma capsulatum Genomes and Present a High Population Coverage
3.3. Prediction of Antigenicity, Allergenicity, Toxicity, and Physicochemical Properties
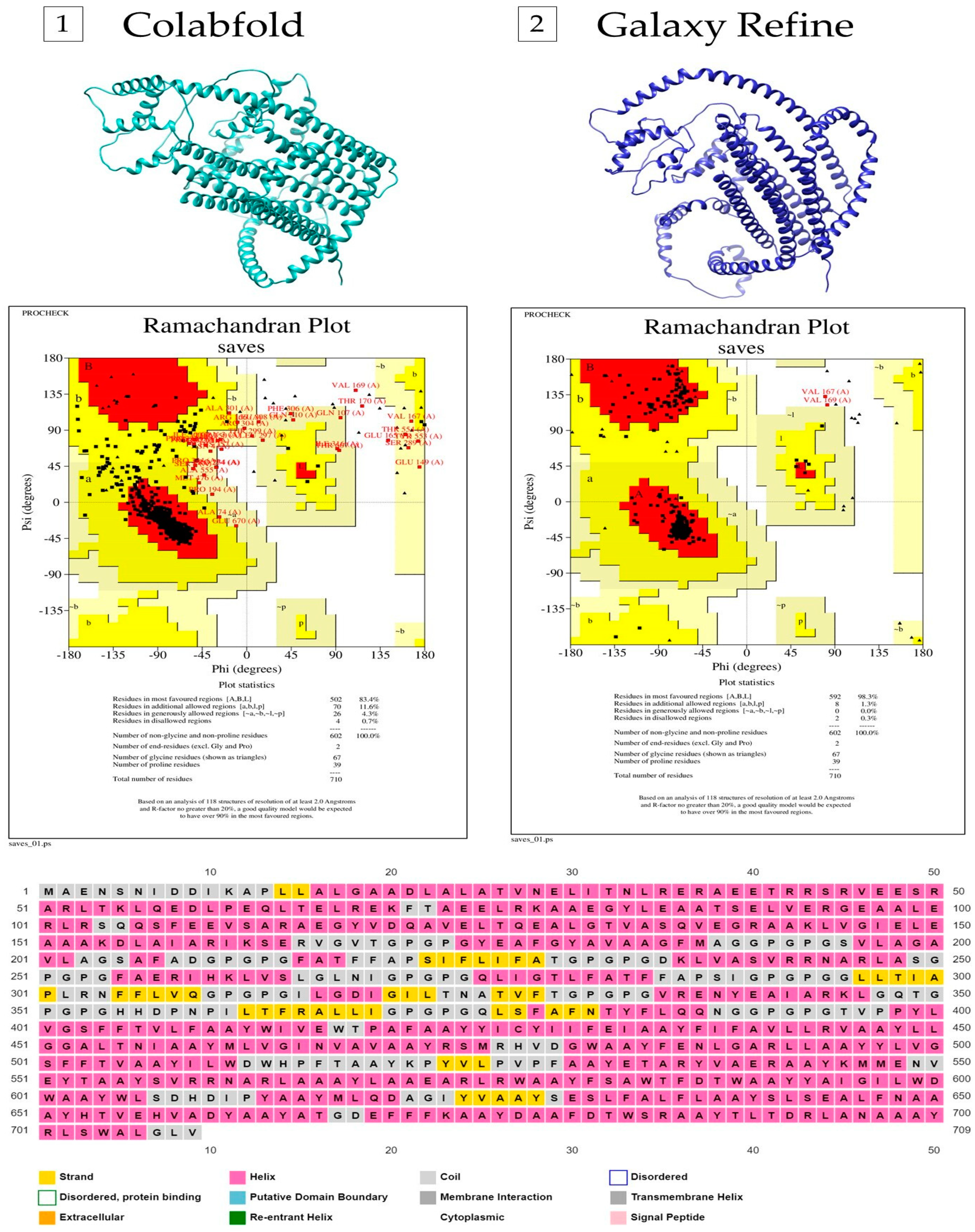
3.4. Secondary and Tertiary Structural Properties of the Multi-Epitope Vaccine
3.5. Docking between the Vaccine and the TLR4 Receptor Shows High Affinity
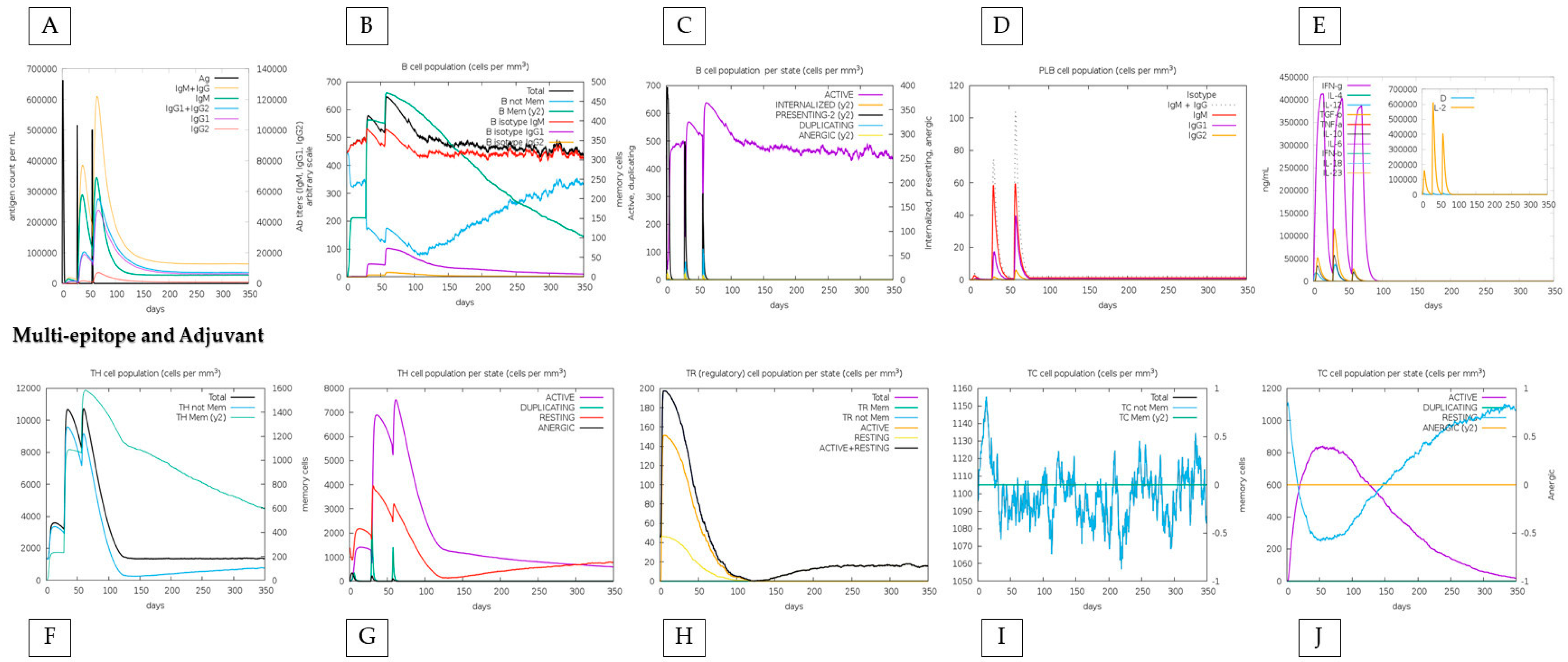
3.6. The “HistoVAC-TSFM” Vaccine Demonstrates High Immunogenicity, Possessing B-Cell Conformational Epitopes, and can Induce the Production of IFN-γ, TNF-α, and Essential Interleukins
3.7. HistoVAC-TSFM Has 10 and 25 MHCI-Inducing Epitopes for Canine and Equine Alleles
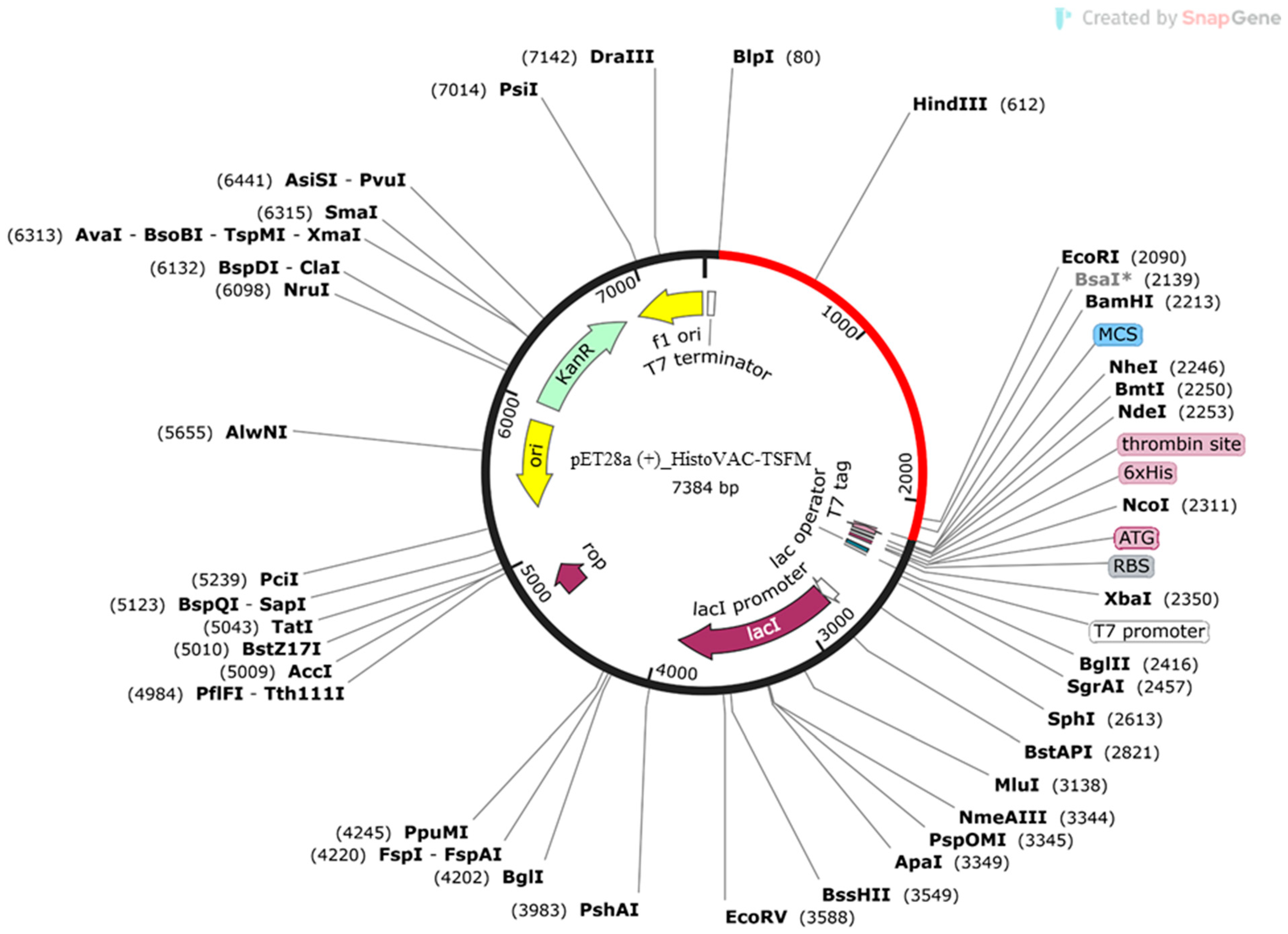
3.8. Codon Adaptation and In Silico Cloning of HistoVAC-TSFM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maiga, A.W.; Deppen, S.; Scaffidi, B.K.; Baddley, J.; Aldrich, M.C.; Dittus, R.S.; Grogan, E.L. Mapping Histoplasma capsulatum Exposure, United States. Emerg. Infect. Dis. 2018, 24, 1835–1839. [Google Scholar] [CrossRef]
- Quinones, F.; Koplan, J.P.; Pike, L.; Staine, F.; Ajello, L. Histoplasmosis in Belize, Central America. Am. J. Trop. Med. Hyg. 1978, 27, 558–561. [Google Scholar] [CrossRef]
- Oladele, R.O.; Ayanlowo, O.O.; Richardson, M.D.; Denning, D.W. Histoplasmosis in Africa: An Emerging or a Neglected Disease? PLoS Negl. Trop. Dis. 2018, 12, e0006046. [Google Scholar] [CrossRef]
- McLeod, D.S.A.; Mortimer, R.H.; Perry-Keene, D.A.; Allworth, A.; Woods, M.L.; Perry-Keene, J.; McBride, W.J.H.; Coulter, C.; Robson, J.M.B. Histoplasmosis in Australia: Report of 16 Cases and Literature Review. Medicine 2011, 90, 61–68. [Google Scholar] [CrossRef]
- Valdez, A.F.; Miranda, D.Z.; Guimarães, A.J.; Nimrichter, L.; Nosanchuk, J.D. Pathogenicity & Virulence of Histoplasma capsulatum—A Multifaceted Organism Adapted to Intracellular Environments. Virulence 2022, 13, 1900–1919. [Google Scholar] [CrossRef]
- Mittal, J.; Ponce, M.G.; Gendlina, I.; Nosanchuk, J.D. Histoplasma capsulatum: Mechanisms for Pathogenesis. In Fungal Physiology and Immunopathogenesis; Current Topics in Microbiology and Immunology Series; Springer: Berlin/Heidelberg, Germany, 2019; Volume 422, pp. 157–191. [Google Scholar] [CrossRef]
- Mihu, M.R.; Nosanchuk, J.D. Histoplasma Virulence and Host Responses. Int. J. Microbiol. 2012, 2012, 268123. [Google Scholar] [CrossRef]
- Kauffman, C.A.; Israel, K.S.; Smith, J.W.; White, A.C.; Schwarz, J.; Brooks, G.F. Histoplasmosis in Immunosuppressed Patients. Am. J. Med. 1978, 64, 923–932. [Google Scholar] [CrossRef]
- Caceres, D.H.; Valdes, A. Histoplasmosis and Tuberculosis Co-Occurrence in People with Advanced HIV. J. Fungi 2019, 5, 73. [Google Scholar] [CrossRef]
- Guimarães, A.J.; Nakayasu, E.S.; Sobreira, T.J.P.; Cordero, R.J.B.; Nimrichter, L.; Almeida, I.C.; Nosanchuk, J.D. Histoplasma capsulatum Heat-Shock 60 Orchestrates the Adaptation of the Fungus to Temperature Stress. PLoS ONE 2011, 6, e14660. [Google Scholar] [CrossRef]
- Hadush, B.; Michaelay, M.; Menghistu, H.T.; Abebe, N.; Genzebu, A.T.; Bitsue, H.K.; Afera, B.; Duguma, B.E.; Gugsa, G.; Ameni, G. Epidemiology of Epizootic Lymphangitis of Carthorses in Northern Ethiopia Using Conventional Diagnostic Methods and Nested Polymerase Chain Reaction. BMC Vet. Res. 2020, 16, 375. [Google Scholar] [CrossRef]
- Damasceno, L.S.; Teixeira, M.M.; Barker, B.M.; Almeida, M.A.; Muniz, M.M.; Pizzini, C.V.; Mesquita, J.R.L.; Rodríguez-Arellanes, G.; Ramírez, J.A.; Vite-Garín, T.; et al. Novel clinical and dual infection by Histoplasma capsulatum genotypes in HIV patients from Northeastern, Brazil. Sci. Rep. 2019, 9, 11789. [Google Scholar] [CrossRef]
- Taylor, M.L.; del Rocío Reyes-Montes, M.; Estrada-Bárcenas, D.A.; Zancopé-Oliveira, R.M.; Rodríguez-Arellanes, G.; Ramírez, J.A. Considerations about the Geographic Distribution of Histoplasma Species. Appl. Environ. Microbiol. 2022, 88, e0201021. [Google Scholar] [CrossRef]
- Almeida, M.A.; Almeida-Silva, F.; Guimarães, A.J.; Almeida-Paes, R.; Zancopé-Oliveira, R.M. The Occurrence of Histoplasmosis in Brazil: A Systematic Review. Int. J. Infect. Dis. 2019, 86, 147–156. [Google Scholar] [CrossRef]
- Horwath, M.C.; Fecher, R.A.; Deepe, G.S. Histoplasma capsulatum, Lung Infection and Immunity. Future Microbiol. 2015, 10, 967–975. [Google Scholar] [CrossRef]
- Ray, S.C.; Rappleye, C.A. Flying under the Radar: Histoplasma Capsulatum Avoidance of Innate Immune Recognition. Semin. Cell Dev. Biol. 2019, 89, 91–98. [Google Scholar] [CrossRef]
- Almeida, M.A.; Almeida-Paes, R.; Guimarães, A.J.; Valente, R.H.; de Almeida Soares, C.M.; Zancopé-Oliveira, R.M. Immunoproteomics Reveals Pathogen’s Antigens Involved in Homo sapiens–Histoplasma capsulatum Interaction and Specific Linear B-Cell Epitopes in Histoplasmosis. Front. Cell. Infect. Microbiol. 2020, 10, 591121. [Google Scholar] [CrossRef]
- Kischkel, B.; Boniche-Alfaro, C.; de Godoy Menezes, I.; Rossi, S.A.; Angeli, C.B.; de Almeida, S.R.; Palmisano, G.; Lopes-Bezerra, L.; Nosanchuk, J.D.; Taborda, C.P. Immunoproteomic and Immunopeptidomic Analyses of Histoplasma capsulatum Reveal Promiscuous and Conserved Epitopes among Fungi with Vaccine Potential. Front. Immunol. 2021, 12, 764501. [Google Scholar] [CrossRef]
- Almeida, P.C.S.; Roque, B.S.; Felice, A.G.; Jaiswal, A.K.; Tiwari, S.; Azevedo, V.; Silva-Vergara, M.L.; de Castro Soares, S.; Ferreira-Paim, K.; Fonseca, F.M. Comparative Genomics of Histoplasma capsulatum and Prediction of New Vaccines and Drug Targets. J. Fungi 2023, 9, 193. [Google Scholar] [CrossRef]
- Rodrigues, T.C.V.; Jaiswal, A.K.; De Sarom, A.; Oliveira, L.D.C.; Oliveira, C.J.F.; Ghosh, P.; Tiwari, S.; Miranda, F.M.; Benevides, L.D.J.; Azevedo, V.A.D.C.; et al. Reverse Vaccinology and Subtractive Genomics Reveal New Therapeutic Targets against Mycoplasma pneumoniae: A Causative Agent of Pneumonia. R. Soc. Open Sci. 2019, 6, 190907. [Google Scholar] [CrossRef]
- Vilela Rodrigues, T.C.; Jaiswal, A.K.; Lemes, M.R.; da Silva, M.V.; Sales-Campos, H.; Alcântara, L.C.J.; de Oliviera Tosta, S.F.; Kato, R.B.; Alzahrani, K.J.; Barh, D.; et al. An Immunoinformatics-Based Designed Multi-Epitope Candidate Vaccine (Mpme-VAC/STV-1) against Mycoplasma pneumoniae. Comput. Biol. Med. 2022, 142, 105194. [Google Scholar] [CrossRef]
- Dhanda, S.K.; Mahajan, S.; Paul, S.; Yan, Z.; Kim, H.; Jespersen, M.C.; Jurtz, V.; Andreatta, M.; Greenbaum, J.A.; Marcatili, P.; et al. IEDB-AR: Immune Epitope Database—Analysis Resource in 2019. Nucleic Acids Res. 2019, 47, W502–W506. [Google Scholar] [CrossRef]
- Wang, X.; Sun, Q.; Ye, Z.; Hua, Y.; Shao, N.; Du, Y.; Zhang, Q.; Wan, C. Computational Approach for Predicting the Conserved B-Cell Epitopes of Hemagglutinin H7 Subtype Influenza Virus. Exp. Ther. Med. 2016, 12, 2439–2446. [Google Scholar] [CrossRef]
- Calis, J.J.A.; Maybeno, M.; Greenbaum, J.A.; Weiskopf, D.; De Silva, A.D.; Sette, A.; Keşmir, C.; Peters, B. Properties of MHC Class I Presented Peptides That Enhance Immunogenicity. PLoS Comput. Biol. 2013, 9, e1003266. [Google Scholar] [CrossRef]
- Dhanda, S.K.; Vaughan, K.; Schulten, V.; Grifoni, A.; Weiskopf, D.; Sidney, J.; Peters, B.; Sette, A. Development of a Novel Clustering Tool for Linear Peptide Sequences. Immunology 2018, 155, 331–345. [Google Scholar] [CrossRef]
- Bui, H.H.; Sidney, J.; Dinh, K.; Southwood, S.; Newman, M.J.; Sette, A. Predicting Population Coverage of T-Cell Epitope-Based Diagnostics and Vaccines. BMC Bioinform. 2006, 7, 153. [Google Scholar] [CrossRef]
- Reynisson, B.; Alvarez, B.; Paul, S.; Peters, B.; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: Improved Predictions of MHC Antigen Presentation by Concurrent Motif Deconvolution and Integration of MS MHC Eluted Ligand Data. Nucleic Acids Res. 2021, 48, W449–W454. [Google Scholar] [CrossRef]
- Chauhan, V.; Rungta, T.; Goyal, K.; Singh, M.P. Designing a Multi-Epitope Based Vaccine to Combat Kaposi Sarcoma Utilizing Immunoinformatics Approach. Sci. Rep. 2019, 9, 2517. [Google Scholar] [CrossRef]
- Chen, X.; Zaro, J.L.; Shen, W.C. Fusion Protein Linkers: Property, Design and Functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef]
- Kumar, P.; Lata, S.; Shankar, U.N.; Akif, M. Immunoinformatics-Based Designing of a Multi-Epitope Chimeric Vaccine from Multi-Domain Outer Surface Antigens of Leptospira. Front. Immunol. 2021, 12, 735373. [Google Scholar] [CrossRef]
- Parra, M.; Pickett, T.; Delogu, G.; Dheenadhayalan, V.; Debrie, A.S.; Locht, C.; Brennan, M.J. The Mycobacterial Heparin-Binding Hemagglutinin Is a Protective Antigen in the Mouse Aerosol Challenge Model of Tuberculosis. Infect. Immun. 2004, 72, 6799–6805. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Springer Protocols Handbooks Series; Springer: Berlin/Heidelberg, Germany, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Dimitrov, I.; Flower, D.R.; Doytchinova, I. AllerTOP—A server for in silico prediction of allergens. BMC Bioinform. 2013, 14, S4. [Google Scholar] [CrossRef]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Raghava, G.P.S. In Silico Approach for Predicting Toxicity of Peptides and Proteins. PLoS ONE 2013, 8, e73957. [Google Scholar] [CrossRef]
- Larsen, M.V.; Lundegaard, C.; Lamberth, K.; Buus, S.; Lund, O.; Nielsen, M. Large-Scale Validation of Methods for Cytotoxic T-Lymphocyte Epitope Prediction. BMC Bioinform. 2007, 8, 424. [Google Scholar] [CrossRef]
- Mcguffin, L.J.; Bryson, K.; Jones, D.T. The PSIPRED protein structure prediction server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Macarthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A Program to Check the Stereochemicai Quality of Protein Structures. J. Appl. Cryst. 1983, 26, 283–291. [Google Scholar] [CrossRef]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein Structure Refinement Driven by Side-Chain Repacking. Nucleic Acids Res. 2013, 41, W384–W388. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, D.; Zhou, P.; Li, B.; Huang, S.Y. HDOCK: A Web Server for Protein-Protein and Protein-DNA/RNA Docking Based on a Hybrid Strategy. Nucleic Acids Res. 2017, 45, W365–W373. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand-Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Weng, G.; Wang, E.; Wang, Z.; Liu, H.; Zhu, F.; Li, D.; Hou, T. HawkDock: A Web Server to Predict and Analyze the Protein-Protein Complex Based on Computational Docking and MM/GBSA. Nucleic Acids Res. 2019, 47, W322–W330. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Ponomarenko, J.; Bui, H.H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A New Structure-Based Tool for the Prediction of Antibody Epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef]
- Rapin, N.; Lund, O.; Bernaschi, M.; Castiglione, F. Computational Immunology Meets Bioinformatics: The Use of Prediction Tools for Molecular Binding in the Simulation of the Immune System. PLoS ONE 2010, 5, e9862. [Google Scholar] [CrossRef]
- Dhall, A.; Patiyal, S.; Choudhury, S.; Jain, S.; Narang, K.; Raghava, G.P.S. TNFepitope: A Webserver for the Prediction of TNF-α Inducing Epitopes. Comput. Biol. Med. 2023, 160, 106929. [Google Scholar] [CrossRef]
- Dhanda, S.K.; Vir, P.; Raghava, G.P. Designing of Interferon-Gamma Inducing MHC Class-II Binders. Biol. Direct 2013, 8, 30. [Google Scholar] [CrossRef]
- Gupta, S.; Mittal, P.; Madhu, M.K.; Sharma, V.K. IL17eScan: A Tool for the Identification of Peptides Inducing IL-17 Response. Front. Immunol. 2017, 8, 1430. [Google Scholar] [CrossRef]
- Dhall, A.; Patiyal, S.; Sharma, N.; Usmani, S.S.; Raghava, G.P.S. Computer-Aided Prediction and Design of IL-6 Inducing Peptides: IL-6 Plays a Crucial Role in COVID-19. Brief. Bioinform. 2021, 22, 936–945. [Google Scholar] [CrossRef]
- Grote, A.; Hiller, K.; Scheer, M.; Münch, R.; Nörtemann, B.; Hempel, D.C.; Jahn, D. JCat: A Novel Tool to Adapt Codon Usage of a Target Gene to Its Potential Expression Host. Nucleic Acids Res. 2005, 33, W526–W531. [Google Scholar] [CrossRef]
- Linder, K.A.; Kauffman, C.A. Histoplasmosis: Epidemiology, Diagnosis, and Clinical Manifestations. Curr. Fungal Infect. Rep. 2019, 13, 120–128. [Google Scholar] [CrossRef]
- Cain, J.A.; Deepe, G.S. Evolution of the Primary Immune Response to Histoplasma capsulatum in Murine Lung. Infect. Immun. 1998, 66, 1473–1481. [Google Scholar] [CrossRef]
- Shi, L.; Albuquerque, P.C.; Lazar-Molnar, E.; Wang, X.; Santambrogio, L.; Gácser, A.; Nosanchuk, J.D. A Monoclonal Antibody to Histoplasma capsulatum Alters the Intracellular Fate of the Fungus in Murine Macrophages. Eukaryot. Cell 2008, 7, 1109–1117. [Google Scholar] [CrossRef]
- Wheat, J.; French, M.L.; Kohler, R.B.; Zimmerman, S.E.; Smith, W.R.; Norton, J.A.; Eitzen, H.E. The Diagnostic Laboratory Tests for Histoplasmosis Analysis of Experience in a Large Urban Outbreak. Ann. Inter. Med. 1982, 97, 680–685. [Google Scholar] [CrossRef]
- Deepe, G.S. Preventive and Therapeutic Vaccines for Fungal Infections: From Concept to Implementation. Expert Rev. Vaccines 2004, 3, 701–709. [Google Scholar] [CrossRef]
- Bian, Y.; Walter, D.L.; Zhang, C. Efficiency of Interferon-γ in Activating Dendritic Cells and Its Potential Synergy with Toll-like Receptor Agonists. Viruses 2023, 15, 1198. [Google Scholar] [CrossRef]
- Wood, K.L.; Hage, C.A.; Knox, K.S.; Kleiman, M.B.; Sannuti, A.; Day, R.B.; Wheat, L.J.; Twigg, H.L. Histoplasmosis after Treatment with Anti-Tumor Necrosis Factor-α Therapy. Am. J. Respir. Crit. Care Med. 2003, 167, 1279–1282. [Google Scholar] [CrossRef]
- Kleinschek, M.A.; Muller, U.; Brodie, S.J.; Stenzel, W.; Kohler, G.; Blumenschein, W.M.; Straubinger, R.K.; Mcclanahan, T.; Kastelein, R.A.; Alber, G. IL-23 enhances the inflammatory cell response in Cryptococcus neoformans infection and induces a cytokine pattern distinct from IL-12. J. Immunol. 2006, 176, 1098–1106. [Google Scholar] [CrossRef]
- Huang, W.; Na, L.; Fidel, P.L.; Schwarzenberger, P. Requirement of Interleukin-17A for Systemic Anti-Candida Albicans Host Defense in Mice. J. Infect. Dis. 2004, 190, 624–631. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Zancopé-Oliveira, R.M.; Hamilton, A.J.; Guimarães, A.J. Antibody therapy for histoplasmosis. Front. Microbiol. 2012, 3, 21. [Google Scholar] [CrossRef]
- Brömel, C.; Sykes, J.E. Histoplasmosis in Dogs and Cats. Clin. Tech. Small Anim. Pract. 2005, 20, 227–232. [Google Scholar] [CrossRef]
- Ameni, G.; Messele Kebede, A.; Zewude, A.; Girma Abdulla, M.; Asfaw, R.; Gobena, M.M.; Kyalo, M.; Stomeo, F.; Gumi, B.; Sori, T. Equine Histoplasmosis in Ethiopia: Phylogenetic Analysis by Sequencing of the Internal Transcribed Spacer Region of RRNA Genes. Front. Cell. Infect. Microbiol. 2022, 12, 789157. [Google Scholar] [CrossRef]
- Ghaemi, M.; Ahmadi, N.; Sharifiyazdi, H.; Ghane, M.; Golvajooei, M.S. First Report of Ocular Histoplasmosis in a Horse from Iran: Molecular, Clinical and Pathological Findings. Vet. Res. Forum 2022, 13, 455–459. [Google Scholar] [CrossRef]
- Cabañes, F.J. Epizootic Lymphangitis: A Neglected Disease of Working Equids. Rev. Iberoam. Micol. 2022, 39, 4–5. [Google Scholar] [CrossRef]
- Oli, A.N.; Obialor, W.O.; Ifeanyichukwu, M.O.; Odimegwu, D.C.; Okoyeh, J.N.; Emechebe, G.O.; Adejumo, S.A.; Ibeanu, G.C. Immunoinformatics and Vaccine Development: An Overview. Immunotargets Ther. 2020, 9, 13–30. [Google Scholar] [CrossRef]
- Azar, M.M.; Loyd, J.L.; Relich, R.F.; Wheat, L.J.; Hage, C.A. Current Concepts in the Epidemiology, Diagnosis, and Management of Histoplasmosis Syndromes. Semin. Respir. Crit. Care Med. 2020, 41, 13–30. [Google Scholar] [CrossRef]
- Falci, D.R.; Dalla Lana, D.F.; Pasqualotto, A.C. The Era of Histoplasmosis in Brazilian Endemic Mycoses. Lancet Reg. Health Am. 2021, 3, 100037. [Google Scholar] [CrossRef]
- Baker, J.; Kosmidis, C.; Rozaliyani, A.; Wahyuningsih, R.; Denning, D.W. Chronic Pulmonary Histoplasmosis—A Scoping Literature Review. Open Forum Infect. Dis. 2020, 7, ofaa119. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, P.H.; Tiwari, S.; Felice, A.G.; Jaiswal, A.K.; Aburjaile, F.F.; Azevedo, V.; Silva-Vergara, M.L.; Ferreira-Paim, K.; Soares, S.d.C.; Fonseca, F.M. Design of a Multi-Epitope Vaccine against Histoplasma capsulatum through Immunoinformatics Approaches. J. Fungi 2024, 10, 43. https://doi.org/10.3390/jof10010043
Marques PH, Tiwari S, Felice AG, Jaiswal AK, Aburjaile FF, Azevedo V, Silva-Vergara ML, Ferreira-Paim K, Soares SdC, Fonseca FM. Design of a Multi-Epitope Vaccine against Histoplasma capsulatum through Immunoinformatics Approaches. Journal of Fungi. 2024; 10(1):43. https://doi.org/10.3390/jof10010043
Chicago/Turabian StyleMarques, Pedro Henrique, Sandeep Tiwari, Andrei Giacchetto Felice, Arun Kumar Jaiswal, Flávia Figueira Aburjaile, Vasco Azevedo, Mario León Silva-Vergara, Kennio Ferreira-Paim, Siomar de Castro Soares, and Fernanda Machado Fonseca. 2024. "Design of a Multi-Epitope Vaccine against Histoplasma capsulatum through Immunoinformatics Approaches" Journal of Fungi 10, no. 1: 43. https://doi.org/10.3390/jof10010043
APA StyleMarques, P. H., Tiwari, S., Felice, A. G., Jaiswal, A. K., Aburjaile, F. F., Azevedo, V., Silva-Vergara, M. L., Ferreira-Paim, K., Soares, S. d. C., & Fonseca, F. M. (2024). Design of a Multi-Epitope Vaccine against Histoplasma capsulatum through Immunoinformatics Approaches. Journal of Fungi, 10(1), 43. https://doi.org/10.3390/jof10010043