
Is Cryptococcus gattii a Primary Pathogen?

Abstract
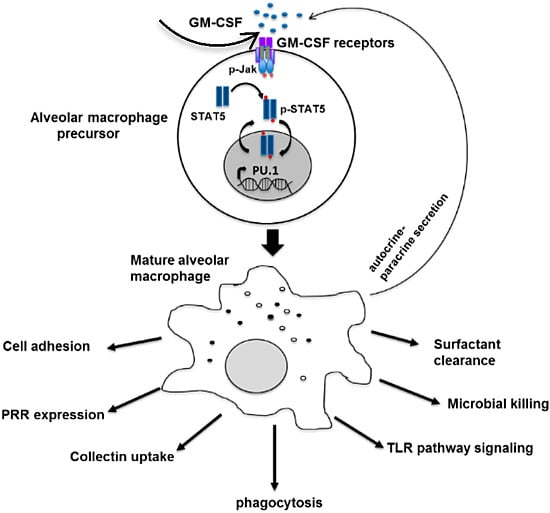
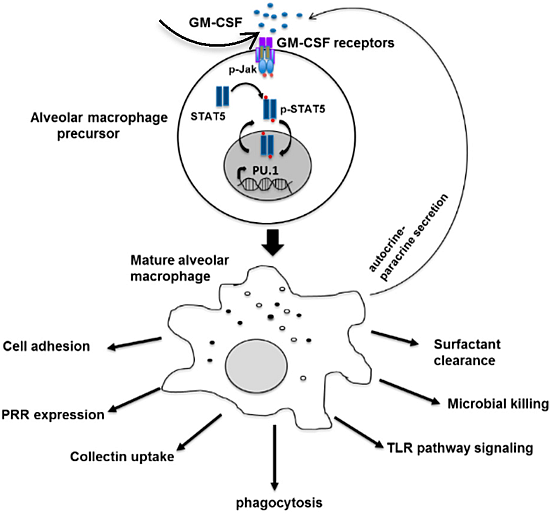
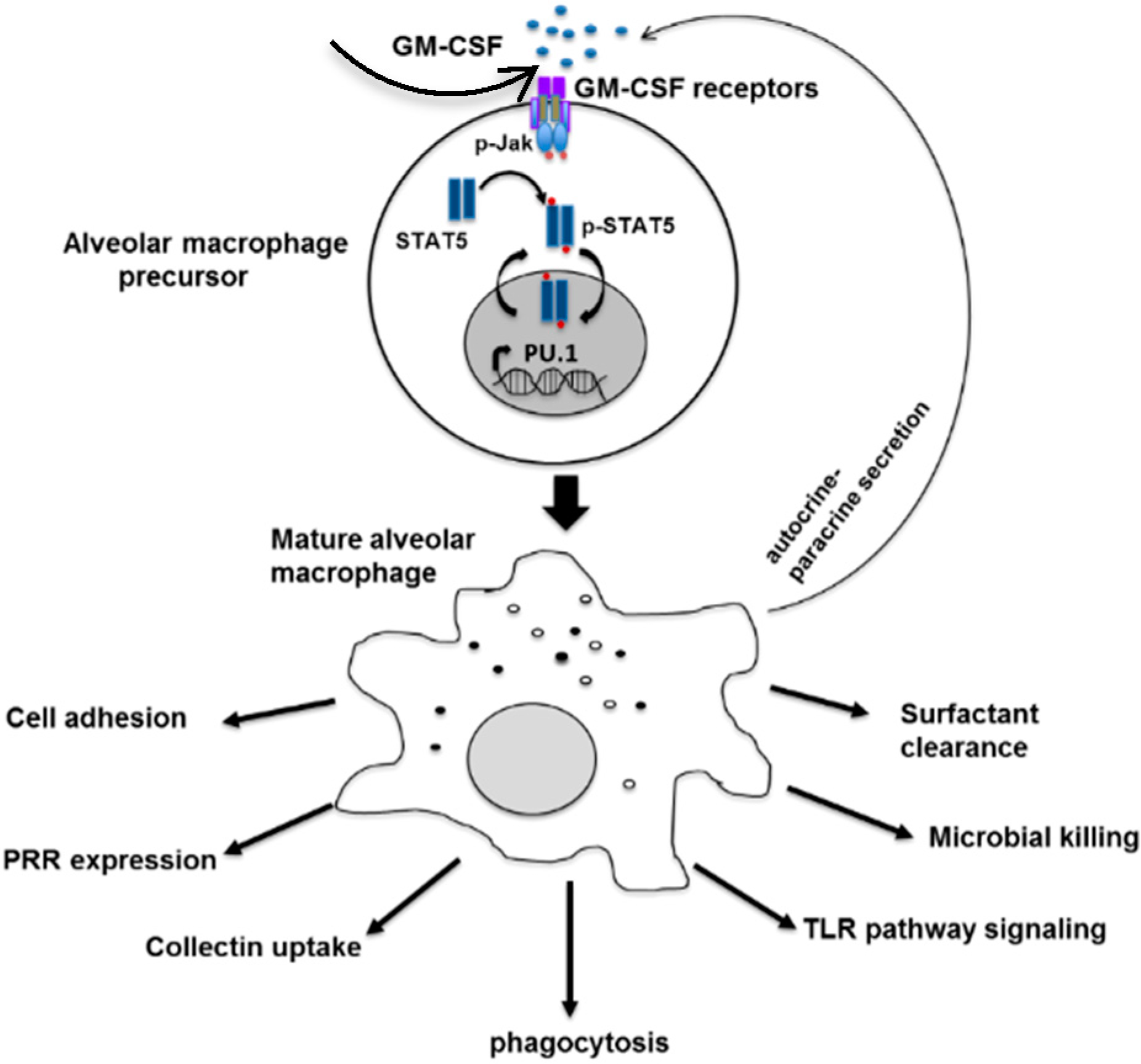
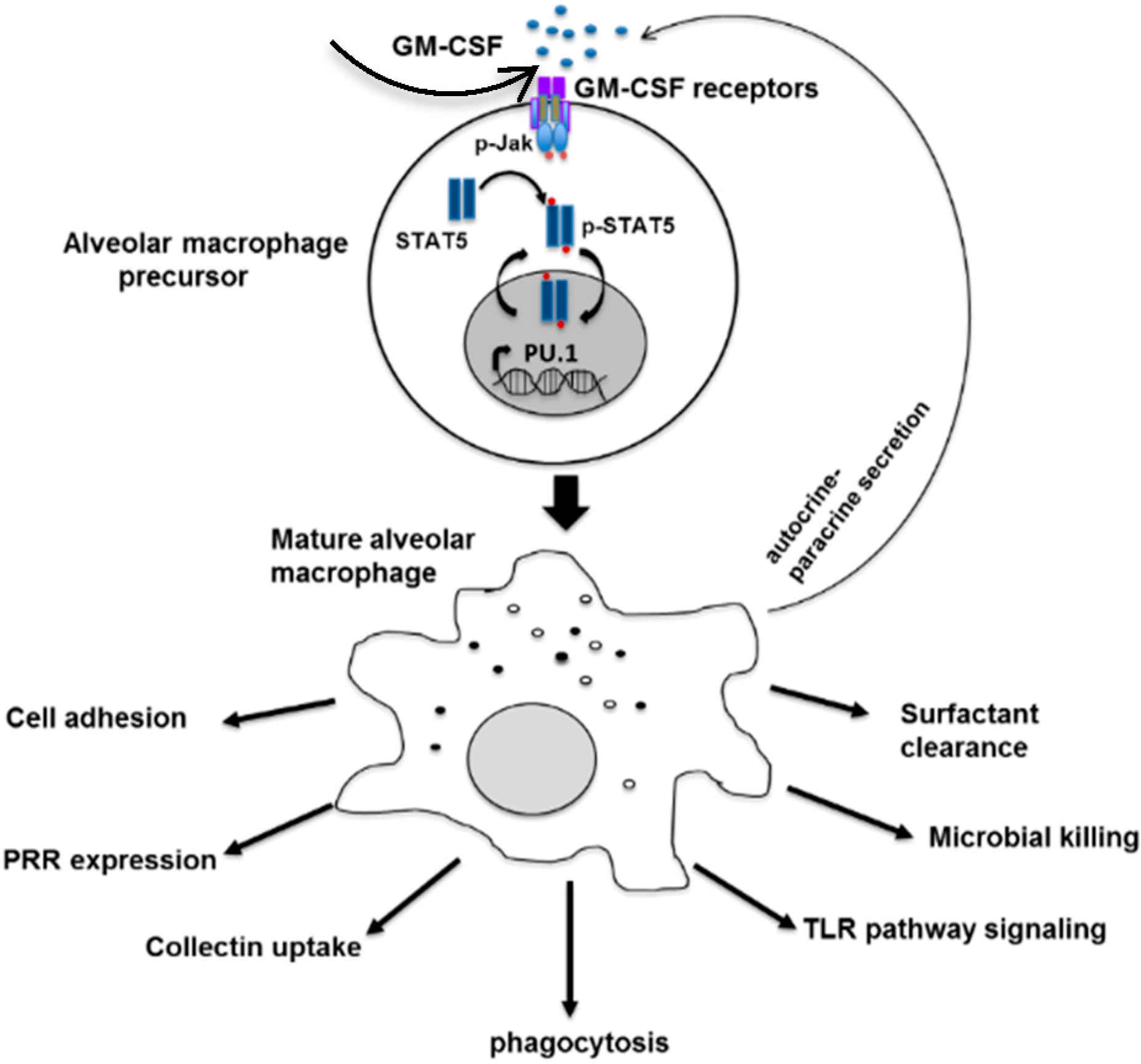
:
1. Introduction
2. How Were GM-CSF Neutralizing Antibodies Identified as a Risk Factor for C. gattii Infection?

{kind=link}
{kind=link}
| No. Patient Samples | Etiologic Agent | Anti-GM-CSF AB |
|---|---|---|
| 20 | C. neoformans, VNl, VNlll | 0 |
| 1 | C. gattii, VGl | 1 |
| 20 (Normal volunteers) | None | 1 |
| Patient | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Origin | S. CA a | S. CA | Thailand | S. CA | NA b | NJ c | NJ |
| Infection | CNS/Lung | CNS/Lung | CNS/Lung | CNS/Lung Blood, Skin | CNS | CNS/Lung | CNS/Lung |
| Species | C. neof. | C. neof. | C. neof. | C. gattii | Cryptococcus | Cryptococcus | Cryptococcus |
| Species Confirmation | N.A d | N.A | N.A | N.A | C. gattii VGlll | C. gattii VGl | C. gattii VGl |
| Patients | Ethnicity | Etiologic Agent | Anti-GM-CSF Ab | Mol. Types |
|---|---|---|---|---|
| 1 | Caucasian | C. gattii | + | VGl |
| 2 | Caucasian | C. gattii | + | VGl |
| 3 | Caucasian | C. gattii | + | VGl |
| 4 | Aborigine | C. gattii | + | VGl |
| 5 | Caucasian | C. gattii | − | VGl |
| 6 | Asian | C. neoformans | − | NA |
| 7 | Asian | C. gattii | + | VGl |
| 8 | Caucasian | C. gattii | + | VGll |
| 9 | Asian | C. gattii | − | VGl |
3. GM-CSF Neutralizing Antibody and Susceptibility to Cryptococcal Infection
4. Could C. gattii Infection Have Induced Production of GM-CSF Neutralizing Antibodies?
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kavanaugh, L.A.; Fraser, J.A.; Dietrich, F.S. Recent evolution of the human pathogen Cryptococcus neoformans by intervarietal transfer of a 14-gene fragment. Mol. Biol. Evol. 2006, 23, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Bennett, J.E. Medical Mycology; Lea & Febiger: Philadelphia, PA, USA, 1992. [Google Scholar]
- Casadevall, A.; Perfect, J.R. Cryptococcus neoformans; ASM Press: Washington, DC, USA, 1998. [Google Scholar]
- Dromer, F.; Mathoulin, S.; Dupont, B.; Laporte, A. Epidemiology of cryptococcosis in France: A 9-year survey (1985–1993). French Cryptococcosis Study Group. Clin. Infect. Dis. 1996, 23, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Varma, A.; Diaz, M.R.; Litvintseva, A.P.; Wollenberg, K.K.; Kwon-Chung, K.J. Cryptococcus neoformans strains and infection in apparently immunocompetent patients, China. Emerg. Infect. Dis. 2008, 14, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Ngamskulrungroj, P.; Varma, A.; Sionov, E.; Hwang, S.M.; Carriconde, F.; Meyer, W.; Litvintseva, A.P.; Lee, W.G.; Shin, J.H.; et al. Prevalence of the VNIc genotype of Cryptococcus neoformans in non-HIV-associated cryptococcosis in the Republic of Korea. FEMS Yeast Res. 2010, 10, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Mihara, T.; Izumikawa, K.; Kakeya, H.; Ngamskulrungroj, P.; Umeyama, T.; Takazono, T.; Tashiro, M.; Nakamura, S.; Imamura, Y.; Miyazaki, T.; et al. Multilocus sequence typing of Cryptococcus neoformans in non-HIV associated cryptococcosis in Nagasaki, Japan. Med. Mycol. 2013, 51, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.; McCarthy, K.M.; Gould, S.; Fan, K.; Arthington-Skaggs, B.; Iqbal, N.; Stamey, K.; Hajjeh, R.A.; Brandt, M.E. Cryptococcus gattii infection: Characteristics and epidemiology of cases identified in a South African province with high HIV seroprevalence, 2002–2004. Clin. Infect. Dis. 2006, 43, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Lizarazo, J.; Escandon, P.; Agudelo, C.I.; Firacative, C.; Meyer, W.; Castaneda, E. Retrospective study of the epidemiology and clinical manifestations of Cryptococcus gattii infections in Colombia from 1997–2011. PLoS Negl. Trop. Dis. 2014, 8, e3272. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, S.; Dyavaiah, M.; Larsen, R.A.; Chaturvedi, V. Cryptococcus gattii in AIDS patients, southern California. Emerg. Infect. Dis. 2005, 11, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Speed, B.; Dunt, D. Clinical and host differences between infections with the two varieties of Cryptococcus neoformans. Clin. Infect. Dis. 1995, 21, 28–34, discussion 35–26. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, T.C.; Chen, S.C.A.; Phillips, P.; Marr, K.A. Clinical perspectives on Cryptococcus neoformans and Cryptococcus gattii: Implications for diagnosis and management. In Cryptococcus: From Human Pathogen to Model Yeast; Heitman, J., Kozel, T.R., Kwon-Chung, K.J., Perfect, J.R., Casadevall, A., Eds.; ASM Press: Washington, DC, USA, 2011; pp. 595–606. [Google Scholar]
- Chen, S.C.; Slavin, M.A.; Heath, C.H.; Playford, E.G.; Byth, K.; Marriott, D.; Kidd, S.E.; Bak, N.; Currie, B.; Hajkowicz, K.; et al. Clinical manifestations of Cryptococcus gattii infection: Determinants of neurological sequelae and death. Clin. Infect. Dis. 2012, 55, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Seaton, R.A.; Naraqi, S.; Wembri, J.P.; Warrell, D.A. Predictors of outcome in Cryptococcus neoformans var. gattii meningitis. QJM 1996, 89, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.H.; Sorrell, T.C.; Allworth, A.M.; Heath, C.H.; McGregor, A.R.; Papanaoum, K.; Richards, M.J.; Gottlieb, T. Cryptococcal disease of the CNS in immunocompetent hosts: Influence of cryptococcal variety on clinical manifestations and outcome. Clin. Infect. Dis. 1995, 20, 611–616. [Google Scholar] [CrossRef] [PubMed]
- MacDougall, L.; Fyfe, M.; Romney, M.; Starr, M.; Galanis, E. Risk factors for Cryptococcus gattii infection, British Columbia, Canada. Emerg. Infect. Dis. 2011, 17, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Harrison, T.S. HIV-associated cryptococcal meningitis. AIDS 2007, 21, 2119–2129. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Casazza, J.P.; Stone, H.H.; Meintjes, G.; Lawn, S.D.; Levitz, S.M.; Harrison, T.S.; Koup, R.A. The phenotype of the Cryptococcus-specific CD4+ memory T-cell response is associated with disease severity and outcome in HIV-associated cryptococcal meningitis. J. Infect. Dis. 2013, 207, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Polacheck, I.; Bennett, J.E. Improved diagnostic medium for separation of Cryptococcus neoformans var. neoformans (serotypes A and D) and Cryptococcus neoformans var. gattii (serotypes B and C). J. Clin. Microbiol. 1982, 15, 535–537. [Google Scholar]
- Kwon-Chung, K.J.; Fraser, J.A.; Doering, T.L.; Wang, Z.; Janbon, G.; Idnurm, A.; Bahn, Y.S. Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of cryptococcosis. In Human Fungal Pathogens; Casadevall, A., Mitchell, A.P., Berman, J., Kwon-Chung, K.J., Perfect, J.R., Heitman, J., Eds.; Cold Spring Harbor Press: New York, NY, USA, 2015; pp. 385–411. [Google Scholar]
- Chang, Y.C. Differences between Cryptococcus neoformans and Cryptococcus gattii in the molecular mechanisms governing utilization of D-amino acids as the sole nitrogen source. PLoS ONE 2015, e0131865. [Google Scholar] [CrossRef] [PubMed]
- Ngamskulrungroj, P.; Chang, Y.; Sionov, E.; Kwon-Chung, K.J. The primary target organ of Cryptococcus gattii is different from that of Cryptococcus neoformans in a murine model. MBio 2012, 3, e103–e112. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, T.C. Cryptococcus neoformans variety gattii. Med. Mycol. 2001, 39, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Macdougall, L.; Kidd, S.; Morshed, M. Epidemiology of Cryptococcus gattii, British Columbia, Canada, 1999–2007. Emerg. Infect. Dis. 2010, 16, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Rosen, L.B.; Freeman, A.F.; Yang, L.M.; Jutivorakool, K.; Olivier, K.N.; Angkasekwinai, N.; Suputtamongkol, Y.; Bennett, J.E.; Pyrgos, V.; Williamson, P.R.; et al. Anti-GM-CSF autoantibodies in patients with cryptococcal meningitis. J. Immunol. 2013, 190, 3959–3966. [Google Scholar] [CrossRef] [PubMed]
- Saijo, T.; Chen, J.; Chen, S.C.; Rosen, L.B.; Yi, J.; Sorrell, T.C.; Bennett, J.E.; Holland, S.M.; Browne, S.K.; Kwon-Chung, K.J. Anti-granulocyte-macrophage colony-stimulating factor autoantibodies are a risk factor for central nervous system infection by Cryptococcus gattii in otherwise immunocompetent patients. MBio 2014, 5, e912–e914. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.; Gilgado, F.; Ngamskulrungroj, P.; Trilles, L.; Hagen, F.; Castaneda, E.; Boekhout, T. Molecular typing of the Cryptococcus neoformans/Cryptococcus gattii species complex. In Cryptococcus: From Human Pathogen to Model Yeast; ASM Press: Washington, DC, USA, 2011; pp. 327–357. [Google Scholar]
- Kwon-Chung, K.J.; Bennett, J.E. Epidemiologic differences between the two varieties of Cryptococcus neoformans. Am. J. Epidemiol. 1984, 120, 123–130. [Google Scholar] [PubMed]
- Kwon-Chung, K.J.; Bennett, J.E. High prevalence of Cryptococcus neoformans var. gattii in tropical and subtropical regions. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1984, 257, 213–218. [Google Scholar] [PubMed]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H.; et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [PubMed]
- Gasson, J.C. Molecular physiology of granulocyte-macrophage colony-stimulating factor. Blood 1991, 77, 1131–1145. [Google Scholar] [PubMed]
- Stanley, E.; Lieschke, G.J.; Grail, D.; Metcalf, D.; Hodgson, G.; Gall, J.A.; Maher, D.W.; Cebon, J.; Sinickas, V.; Dunn, A.R. Granulocyte/macrophage colony-stimulating factor-deficient mice show no major perturbation of hematopoiesis but develop a characteristic pulmonary pathology. Proc. Natl Acad. Sci. USA 1994, 91, 5592–5596. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, D. Control of granulocytes and macrophages: Molecular, cellular, and clinical aspects. Science 1991, 254, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Akagawa, K.S.; Kamoshita, K.; Tokunaga, T. Effects of granulocyte-macrophage colony-stimulating factor and colony-stimulating factor-1 on the proliferation and differentiation of murine alveolar macrophages. J. Immunol. 1988, 141, 3383–3390. [Google Scholar] [PubMed]
- Chen, B.D.; Mueller, M.; Chou, T.H. Role of granulocyte/macrophage colony-stimulating factor in the regulation of murine alveolar macrophage proliferation and differentiation. J. Immunol. 1988, 141, 139–144. [Google Scholar] [PubMed]
- Uchida, K.; Beck, D.C.; Yamamoto, T.; Berclaz, P.Y.; Abe, S.; Staudt, M.K.; Carey, B.C.; Filippi, M.D.; Wert, S.E.; Denson, L.A.; et al. GM-CSF autoantibodies and neutrophil dysfunction in pulmonary alveolar proteinosis. N. Engl. J. Med. 2007, 356, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Van de Laar, L.; Coffer, P.J.; Woltman, A.M. Regulation of dendritic cell development by GM-CSF: Molecular control and implications for immune homeostasis and therapy. Blood 2012, 119, 3383–3393. [Google Scholar]
- Bonfield, T.L.; Raychaudhuri, B.; Malur, A.; Abraham, S.; Trapnell, B.C.; Kavuru, M.S.; Thomassen, M.J. PU.1 regulation of human alveolar macrophage differentiation requires granulocyte-macrophage colony-stimulating factor. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, L1132–L1136. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Berclaz, P.Y.; Chroneos, Z.C.; Yoshida, M.; Whitsett, J.A.; Trapnell, B.C. GM-CSF regulates alveolar macrophage differentiation and innate immunity in the lung through PU.1. Immunity 2001, 15, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Collins, H.L.; Bancroft, G.J. Cytokine enhancement of complement-dependent phagocytosis by macrophages: Synergy of tumor necrosis factor-alpha and granulocyte-macrophage colony-stimulating factor for phagocytosis of Cryptococcus neoformans. Eur. J. Immunol. 1992, 22, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.H.; Olszewski, M.A.; McDonald, R.A.; Wells, J.C.; Paine, R.; Huffnagle, G.B.; Toews, G.B. Role of granulocyte macrophage colony-stimulating factor in host defense against pulmonary Cryptococcus neoformans infection during murine allergic bronchopulmonary mycosis. Am. J. Pathol. 2007, 170, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Chiller, T.; Farrokhshad, K.; Brummer, E.; Stevens, D.A. Effect of granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor on polymorphonuclear neutrophils, monocytes or monocyte-derived macrophages combined with voriconazole against Cryptococcus neoformans. Med. Mycol. 2002, 40, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Tascini, C.; Vecchiarelli, A.; Preziosi, R.; Francisci, D.; Bistoni, F.; Baldelli, F. Granulocyte-macrophage colony-stimulating factor and fluconazole enhance anti-cryptococcal activity of monocytes from AIDS patients. AIDS 1999, 13, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Seymour, J.F.; Presneill, J.J. Pulmonary alveolar proteinosis: Progress in the first 44 years. Am. J. Respir. Crit. Care Med. 2002, 166, 215–235. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Katyal, S.L.; Bedrossian, C.W.; Rogers, R.M. Pulmonary alveolar proteinosis. Staining for surfactant apoprotein in alveolar proteinosis and in conditions simulating it. Chest 1983, 83, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, B.C.; Whitsett, J.A. GM-CSF regulates pulmonary surfactant homeostasis and alveolar macrophage-mediated innate host defense. Annu. Rev. Physiol. 2002, 64, 775–802. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, B.C.; Carey, B.C.; Uchida, K.; Suzuki, T. Pulmonary alveolar proteinosis, a primary immunodeficiency of impaired GM-CSF stimulation of macrophages. Curr. Opin. Immunol. 2009, 21, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.M.; Macleod, W.M. Pulmonary alveolar proteinosis. Br. J. Dis. Chest 1969, 63, 13–28. [Google Scholar] [CrossRef]
- Rosen, S.H.; Castleman, B.; Liebow, A.A. Pulmonary alveolar proteinosis. N. Engl. J. Med. 1958, 258, 1123–1142. [Google Scholar] [CrossRef] [PubMed]
- Carey, B.; Trapnell, B.C. The molecular basis of pulmonary alveolar proteinosis. Clin. Immunol. 2010, 135, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, B.C.; Whitsett, J.A.; Nakata, K. Pulmonary alveolar proteinosis. N. Engl. J. Med. 2003, 349, 2527–2539. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Watanabe, J.; Kitamura, T.; Yamada, Y.; Kanegasaki, S.; Nakata, K. Lungs of patients with idiopathic pulmonary alveolar proteinosis express a factor which neutralizes granulocyte-macrophage colony stimulating factor. FEBS Lett. 1999, 442, 246–250. [Google Scholar] [CrossRef]
- Kitamura, T.; Tanaka, N.; Watanabe, J.; Uchida; Kanegasaki, S.; Yamada, Y.; Nakata, K. Idiopathic pulmonary alveolar proteinosis as an autoimmune disease with neutralizing antibody against granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 1999, 190, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.L.; Hansell, D.; Lawson, P.R.; Reid, K.B.; Morgan, C. Pulmonary alveolar proteinosis: Clinical aspects and current concepts on pathogenesis. Thorax 2000, 55, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Thomson, C.A.; Allan, L.L.; Jackson, L.M.; Olson, M.; Hercus, T.R.; Nero, T.L.; Turner, A.; Parker, M.W.; Lopez, A.L.; et al. Characterization of pathogenic human monoclonal autoantibodies against GM-CSF. Proc. Natl. Acad. Sci. USA 2013, 110, 7832–7837. [Google Scholar] [CrossRef] [PubMed]
- Kavuru, M.S.; Sullivan, E.J.; Piccin, R.; Thomassen, M.J.; Stoller, J.K. Exogenous granulocyte-macrophage colony-stimulating factor administration for pulmonary alveolar proteinosis. Am. J. Respir. Crit. Care Med. 2000, 161, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Seymour, J.F.; Presneill, J.J.; Schoch, O.D.; Downie, G.H.; Moore, P.E.; Doyle, I.R.; Vincent, J.M.; Nakata, K.; Kitamura, T.; Langton, D.; et al. Therapeutic efficacy of granulocyte-macrophage colony-stimulating factor in patients with idiopathic acquired alveolar proteinosis. Am. J. Respir. Crit. Care Med. 2001, 163, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Prakash, U.B.S.; Barham, S.S.; Carpenter, H.A.; Dines, D.E.; Marsh, H.M. Pulmonary alveolar phospholipoproteinosis: Experience with 34 cases and a review. Mayo Clin. Proc. 1987, 62, 499–518. [Google Scholar] [CrossRef]
- Bedrossian, C.W.; Luna, M.A.; Conklin, R.H.; Miller, W.C. Alveolar proteinosis as a consequence of immunosuppression. A hypothesis based on clinical and pathologic observations. Hum. Pathol. 1980, 11, 527–535. [Google Scholar] [PubMed]
- Kohl, K.H.; Hof, H.; Schrettenbrunner, A.; Seeliger, H.P.; Kwon-Chung, K.J. Cryptococcus neoformans var. gattii in Europe. Lancet 1985, 1, 1515. [Google Scholar] [CrossRef]
- Ellis, D.H.; Pfeiffer, T.J. Natural habitat of Cryptococcus neoformans var. gattii. J. Clin. Microbiol. 1990, 28, 1642–1644. [Google Scholar] [PubMed]
- Linell, F.; Magnusson, B.; Norden, A. Cryptococcosis; review and report of a case. Acta Derm. Venereol. 1953, 33, 103–122. [Google Scholar] [PubMed]
- Sunderland, W.A.; Campbell, R.A.; Edwards, M.J. Pulmonary alveolar proteinosis and pulmonary cryptococcosis in an adolescent boy. J. Pediatr. 1972, 80, 450–456. [Google Scholar] [CrossRef]
- McCook, T.A.; Kirks, D.R.; Merten, D.F.; Osborne, D.R.; Spock, A.; Pratt, P.C. Pulmonary alveolar proteinosis in children. Am. J. Roentgenol. 1981, 137, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Chew, G.T.; Robinson, B.W. Pulmonary and meningeal cryptococcosis in pulmonary alveolar proteinosis. Aust. N. Z. J. Med. 1999, 29, 843–844. [Google Scholar] [CrossRef] [PubMed]
- Bjorkholm, B.; Elgefors, B. Cerebellar aspergilloma. Scand. J. Infect. Dis. 1986, 18, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Von Egidy, H.; Bassler, R.; Tilling, W. Contribution to the alveolar proteinosis of the lungs. Beitr. Klin. Erforsch. Tuberk. Lungenkr. 1967, 134, 365–380. [Google Scholar]
- Groniowski, J.; Walski, M.; Szymanska, D. Electron microscopic observations on pulmonary alveolar lipoproteinosis. Ann. Med. Sect. Pol. Acad. Sci. 1974, 19, 109–110. [Google Scholar] [PubMed]
- Kellar, S.L.; Harshfield, D.L.; Grigg, K.G. Radiological case of the month. Pulmonary alveolar proteinosis. J. Ark. Med. Soc. 1995, 92, 307–308. [Google Scholar] [PubMed]
- Herger, P.C. A case study: Anesthetic considerations for pulmonary lavage. AANA J. 1975, 43, 398–400. [Google Scholar] [PubMed]
- Hartung, M.; Salfelder, K. Pulmonary alveolar proteinosis and histoplasmosis: Report of three cases. Virchows Arch. A Pathol. Anat. Histol. 1975, 368, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Deepe, G.S., Jr.; Gibbons, R.; Woodward, E. Neutralization of endogenous granulocyte-macrophage colony-stimulating factor subverts the protective immune response to Histoplasma capsulatum. J. Immunol. 1999, 163, 4985–4993. [Google Scholar] [PubMed]
- Subramanian Vignesh, K.; Landero Figueroa, J.A.; Porollo, A.; Caruso, J.A.; Deepe, G.S., Jr. Zinc sequestration: Arming phagocyte defense against fungal attack. PLoS Pathog. 2013, 9, e1003815. [Google Scholar]
- Subramanian Vignesh, K.; Landero Figueroa, J.A.; Porollo, A.; Caruso, J.A.; Deepe, G.S., Jr. Granulocyte macrophage-colony stimulating factor induced Zn sequestration enhances macrophage superoxide and limits intracellular pathogen survival. Immunity 2013, 39, 697–710. [Google Scholar]
- Brummer, E.; Kamberi, M.; Stevens, D.A. Regulation by granulocyte-macrophage colony-stimulating factor and/or steroids given in vivo of proinflammatory cytokine and chemokine production by bronchoalveolar macrophages in response to Aspergillus conidia. J. Infect. Dis. 2003, 187, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.; Galanis, E.; MacDougall, L.; Chong, M.Y.; Balshaw, R.; Cook, V.J.; Bowie, W.; Steiner, T.; Hoang, L.; Morshed, M.; et al. Longitudinal clinical findings and outcome among patients with Cryptococcus gattii infection in British Columbia. Clin. Infect. Dis. 2015, 60, 1368–1376. [Google Scholar] [PubMed]
- Lipovsky, M.M.; Juliana, A.E.; Gekker, G.; Hu, S.; Hoepelman, A.I.; Peterson, P.K. Effect of cytokines on anticryptococcal activity of human microglial cells. Clin. Diagn. Lab. Immunol. 1998, 5, 410–411. [Google Scholar] [PubMed]
- De Sanctis, P.N. Pulmonary alveolar proteinosis. A review of the findings and theories to date, with a digression on Pneumocystis carinii pneumonia. BMQ 1962, 13, 19–35. [Google Scholar]
- Plenk, H.P.; Swift, S.A.; Chambers, W.L.; Peltzer, W.E. Pulmonary alveolar proteinosis-a new disease? Radiology 1960, 74, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Bergman, F.; Linell, F. Cryptococcosis as a cause of pulmonary alveolar proteinosis. Acta Pathol. Microbiol. Scand. 1961, 53, 217–224. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon-Chung, K.J.; Saijo, T. Is Cryptococcus gattii a Primary Pathogen? J. Fungi 2015, 1, 154-167. https://doi.org/10.3390/jof1020154
Kwon-Chung KJ, Saijo T. Is Cryptococcus gattii a Primary Pathogen? Journal of Fungi. 2015; 1(2):154-167. https://doi.org/10.3390/jof1020154
Chicago/Turabian StyleKwon-Chung, Kyung J., and Tomomi Saijo. 2015. "Is Cryptococcus gattii a Primary Pathogen?" Journal of Fungi 1, no. 2: 154-167. https://doi.org/10.3390/jof1020154
APA StyleKwon-Chung, K. J., & Saijo, T. (2015). Is Cryptococcus gattii a Primary Pathogen? Journal of Fungi, 1(2), 154-167. https://doi.org/10.3390/jof1020154