
Sex Differences in Circadian Clock Genes and Myocardial Infarction Susceptibility




Abstract
1. Introduction
2. Participants and Methods
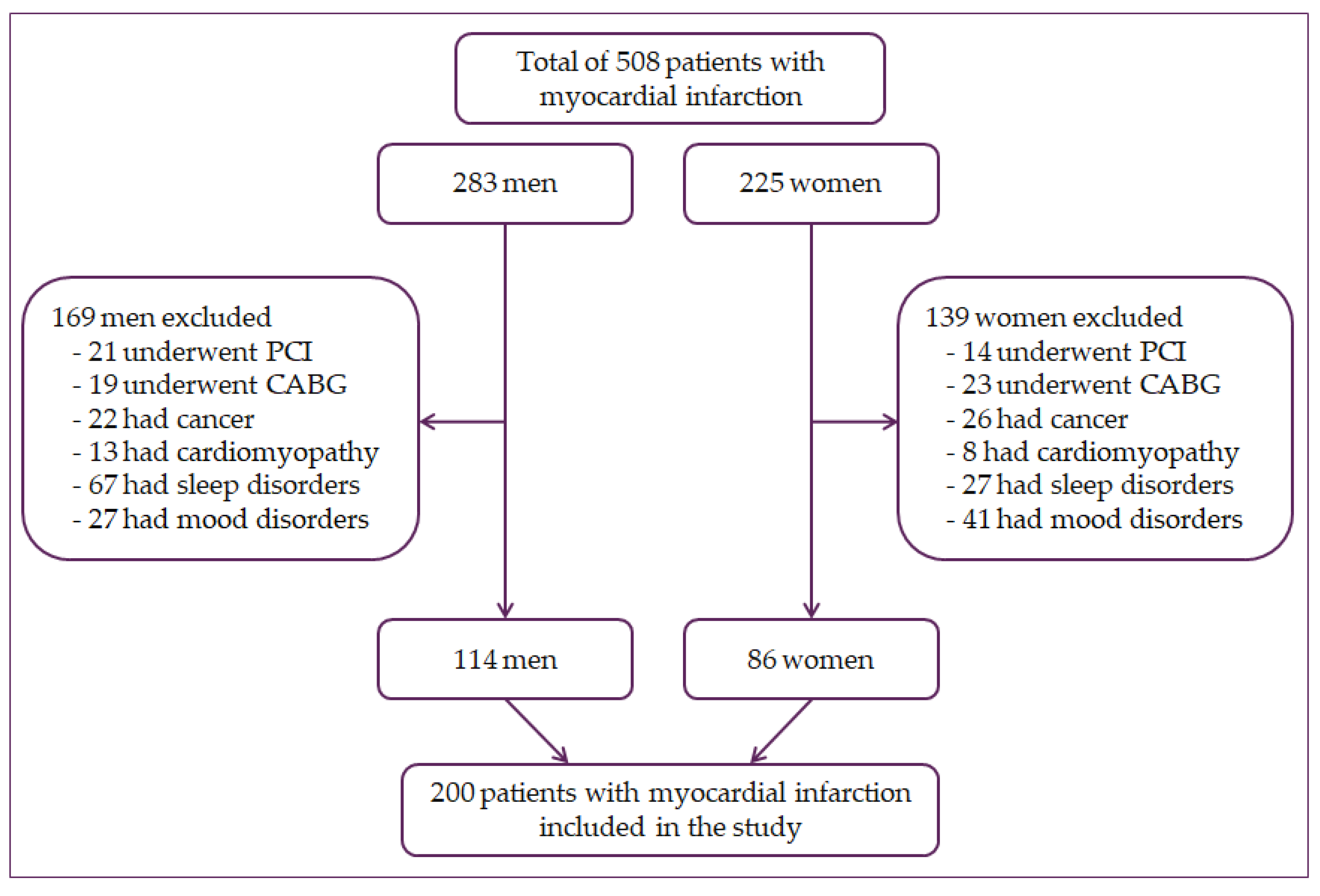
2.1. Participants
2.2. Genotyping of Single Nucleotide Polymorphisms
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hower, I.M.; Harper, S.A.; Buford, T.W. Circadian rhythms, exercise, and cardiovascular health. J. Circadian Rhythm. 2018, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Škrlec, I.; Milić, J.; Heffer, M.; Wagner, J.; Peterlin, B. Circadian clock genes and circadian phenotypes in patients with myocardial infarction. Adv. Med. Sci. 2019, 64, 224–229. [Google Scholar] [CrossRef]
- Crnko, S.; Du Pré, B.C.; Sluijter, J.P.G.; Van Laake, L.W. Circadian rhythms and the molecular clock in cardiovascular biology and disease. Nat. Rev. Cardiol. 2019, 16, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Škrlec, I.; Marić, S.; Včev, A. Myocardial infarction and circadian rhythm. In Visions of Cardiomyocyte-Fundamental Concepts of Heart Life and Disease; IntechOpen: London, UK, 2019; pp. 21–35. [Google Scholar]
- Jain, M.; Zhang, L.; Sabeh, M. Circadian rhythm and cardiovascular disorders. Chrono. Ther. 2014, 4, 27. [Google Scholar] [CrossRef]
- Škrlec, I.; Milić, J.; Cilenšek, I.; Petrovič, D.; Wagner, J.; Peterlin, B. Circadian clock genes and myocardial infarction in patients with type 2 diabetes mellitus. Gene 2019, 701, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Cain, S.W.; Chang, A.-M.; Phillips, A.J.K.; Münch, M.Y.; Gronfier, C.; Wyatt, J.K.; Dijk, D.-J.; Wright, K.P.; Czeisler, C.A. Sex difference in the near-24-hour intrinsic period of the human circadian timing system. Proc. Natl. Acad. Sci. USA 2011, 108, 15602–15608. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.B.; Shechter, A.; Boudreau, P.; Begum, E.A.; Ying-Kin, N.M.K.N. Diurnal and circadian variation of sleep and alertness in men vs. naturally cycling women. Proc. Natl. Acad. Sci. USA 2016, 113, 10980–10985. [Google Scholar] [CrossRef] [PubMed]
- Pyle, W.G.; Martino, T.A. Circadian rhythms influence cardiovascular disease differently in males and females: Role of sex and gender. Curr. Opin. Physiol. 2018, 5, 30–37. [Google Scholar] [CrossRef]
- Thosar, S.S.; Butler, M.P.; Shea, S.A. Role of the circadian system in cardiovascular disease. J. Clin. Investig. 2018, 128, 2157–2167. [Google Scholar] [CrossRef]
- Budkowska, M.; Lebiecka, A.; Marcinowska, Z.; Woźniak, J.; Jastrzębska, M.; Dołęgowska, B. The circadian rhythm of selected parameters of the hemostasis system in healthy people. Thromb. Res. 2019, 182, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Škrlec, I.; Milić, J.; Steiner, R. The impact of the circadian genes clock and arntl on myocardial infarction. J. Clin. Med. 2020, 9, 484. [Google Scholar] [CrossRef]
- Alibhai, F.J.; Reitz, C.J.; Peppler, W.T.; Basu, P.; Sheppard, P.; Choleris, E.; Bakovic, M.; Martino, T.A. Female ClockΔ19/Δ19 mice are protected from the development of age-dependent cardiomyopathy. Cardiovasc. Res. 2018, 114, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Škrlec, I.; Milic, J.; Heffer, M.; Peterlin, B.; Wagner, J. Genetic variations in circadian rhythm genes and susceptibility for myocardial infarction. Genet. Mol. Biol. 2018, 41, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Silva-Urra, J.A.; Núñez-Espinosa, C.A.; Niño-Mendez, O.A.; Gaitán-Peñas, H.; Altavilla, C.; Toro-Salinas, A.; Torrella, J.R.; Pagès, T.; Javierre, C.F.; Behn, C.; et al. Circadian and sex differences after acute high-altitude exposure: Are early acclimation responses improved by blue light. Wilderness Environ. Med. 2015, 26, 459–471. [Google Scholar] [CrossRef]
- Zhao, R.; Li, D.; Zuo, P.; Bai, R.; Zhou, Q.; Fan, J.; Li, C.; Wang, L.; Yang, X. Influences of age, gender, and circadian rhythm on deceleration capacity in subjects without evident heart diseases. Ann. Noninvasive Electrocardiol. 2014, 20, 158–166. [Google Scholar] [CrossRef]
- Eastman, C.I.; Tomaka, V.A.; Crowley, S.J. Sex and ancestry determine the free-running circadian period. J. Sleep Res. 2017, 26, 547–550. [Google Scholar] [CrossRef]
- Link, J.C.; Reue, K. Genetic basis for sex differences in obesity and lipid metabolism. Annu. Rev. Nutr. 2017, 37, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Morris, C.J.; Caputo, R.; Wang, W.; Garaulet, M.; Scheer, F.A.J.L. Sex differences in the circadian misalignment effects on energy regulation. Proc. Natl. Acad. Sci. USA 2019, 116, 23806–23812. [Google Scholar] [CrossRef]
- Arnold, A.P.; Cassis, L.A.; Eghbali, M.; Reue, K.; Sandberg, K. Sex hormones and sex chromosomes cause sex differences in the development of cardiovascular diseases. Arter. Thromb. Vasc. Biol. 2017, 37, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Čulić, V.; Eterović, D.; Mirić, D.; Rumboldt, Z.; Hozo, I. Gender differences in triggering of acute myocardial infarction. Am. J. Cardiol. 2000, 85, 753–756. [Google Scholar] [CrossRef]
- Hatcher, K.M.; Royston, S.E.; Mahoney, M.M. Modulation of circadian rhythms through estrogen receptor signaling. Eur. J. Neurosci. 2020, 51, 217–228. [Google Scholar] [CrossRef]
- Anderson, S.T.; Fitzgerald, G.A. Sexual dimorphism in body clocks. Science 2020, 369, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Škrlec, I.; Milić, J.; Heffer, M.; Steiner, R.; Peterlin, B.; Wagner, J. Association of circadian rhythm with myocardial infarction. Acta Clin. Croat. 2018, 57, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Douma, L.G.; Solocinski, K.; Holzworth, M.R.; Crislip, G.R.; Masten, S.H.; Miller, A.H.; Cheng, K.-Y.; Lynch, I.J.; Cain, B.D.; Wingo, C.S.; et al. Female C57BL/6J mice lacking the circadian clock protein PER1 are protected from nondipping hypertension. Am. J. Physiol. Integr. Comp. Physiol. 2019, 316, R50–R58. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y.; Bogdan, A. Circadian and seasonal variations of physiological and biochemical determinants of acute myocardial infarction. Biol. Rhythm. Res. 2007, 38, 169–179. [Google Scholar] [CrossRef]
- Campbell, D.J. Why do men and women differ in their risk of myocardial infarction. Eur. Heart J. 2008, 29, 835–836. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Breet, N.J.; Sluman, M.A.; Van Berkel, M.A.J.P.J.; Van Werkum, J.W.; Bouman, H.J.; Harmsze, A.M.; Kelder, J.C.; Zijlstra, F.; Hackeng, C.M.; Berg, J.M.T. Effect of gender difference on platelet reactivity. Neth. Hear. J. 2011, 19, 451–457. [Google Scholar] [CrossRef]
- Thygesen, K.; Alpert, J.S.; Jaffe, A.S.; Simoons, M.L.; Chaitman, B.R.; White, H.D. Third universal definition of myocardial infarction. Nat. Rev. Cardiol. 2012, 9, 620–633. [Google Scholar] [CrossRef]
- Thygesen, K.; Alpert, J.S.; Jaffe, A.S.; Chaitman, B.R.; Bax, J.J.; Morrow, D.A.; White, H.D. ESC scientific document group fourth universal definition of myocardial infarction. Eur. Heart J. 2019, 40, 237–269. [Google Scholar] [CrossRef]
- Garaulet, M.; Madrid, J.A. Chronobiology, genetics and metabolic syndrome. Curr. Opin. Lipidol. 2009, 20, 127–134. [Google Scholar] [CrossRef]
- Leu, H.-B.; Chung, C.-M.; Lin, S.-J.; Chiang, K.-M.; Yang, H.-C.; Ho, H.-Y.; Ting, C.-T.; Lin, T.-H.; Sheu, S.-H.; Tsai, W.-C.; et al. Association of circadian genes with diurnal blood pressure changes and non-dipper essential hypertension: A genetic association with young-onset hypertension. Hypertens. Res. 2015, 38, 155–162. [Google Scholar] [CrossRef]
- Englund, A.; Kovanen, L.; Saarikoski, S.T.; Haukka, J.; Reunanen, A.; Aromaa, A.; Lönnqvist, J.; Partonen, T. NPAS2 and PER2 are linked to risk factors of the metabolic syndrome. J. Circadian Rhythm. 2009, 7, 5. [Google Scholar] [CrossRef]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed]
- Moshki, M.; Zareie, M.; Hashemizadeh, H. Sex differences in acute myocardial infarction. Nurs. Midwifery Stud. 2015, 4, 22395. [Google Scholar] [CrossRef] [PubMed]
- Forman, D.E.; Cittadini, A.; Azhar, G.; Douglas, P.S.; Wei, J.Y. Cardiac morphology and function in senescent rats: Gender-related differences. J. Am. Coll. Cardiol. 1997, 30, 1872–1877. [Google Scholar] [CrossRef]
- Rhoads, M.K.; Balagee, V.; Thomas, S.J. Circadian regulation of blood pressure: Of mice and men. Curr. Hypertens. Rep. 2020, 22, 1–9. [Google Scholar] [CrossRef]
- Douma, L.G.; Gumz, M.L. Circadian clock-mediated regulation of blood pressure. Free. Radic. Biol. Med. 2018, 119, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.; Diaz, A.N.; Gumz, M.L. Clock genes in hypertension. Blood Press. Monit. 2014, 19, 249–254. [Google Scholar] [CrossRef]
- Mikulska, A.; Grzelak, T.; Pelczyńska, M.; Bogdański, P.; Czyżewska, K. Assessment of selected clock proteins (clock and cry1) and their relationship with biochemical, anthropometric, and lifestyle parameters in hypertensive patients. Biomolecules 2021, 11, 517. [Google Scholar] [CrossRef]
- Scott, E.M. Circadian clocks, obesity and cardiometabolic function. Diabetes Obes. Metab. 2015, 17, 84–89. [Google Scholar] [CrossRef]
- Bray, M.S.; Shaw, C.A.; Moore, M.W.S.; Garcia, R.A.P.; Zanquetta, M.M.; Durgan, D.J.; Jeong, W.J.; Tsai, J.-Y.; Bugger, H.; Zhang, D.; et al. Disruption of the circadian clock within the cardiomyocyte influences myocardial contractile function, metabolism, and gene expression. Am. J. Physiol. Circ. Physiol. 2008, 294, H1036–H1047. [Google Scholar] [CrossRef] [PubMed]
- Dibner, C.; Schibler, U. Circadian timing of metabolism in animal models and humans. J. Intern. Med. 2015, 277, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Gemma, C.; Gianotti, T.F.; Burgueño, A.; Castaño, G.; Pirola, C.J. Genetic variants of clock transcription factor are associated with individual susceptibility to obesity. Am. J. Clin. Nutr. 2008, 87, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Kovanen, L.; Donner, K.; Kaunisto, M.; Partonen, T. CRY1, CRY2 and PRKCDBP genetic variants in metabolic syndrome. Hypertens. Res. 2014, 38, 186–192. [Google Scholar] [CrossRef]
- Škrlec, I. Circadian rhythm and myocardial infarction. Med. Flum. 2019, 55, 32–42. [Google Scholar] [CrossRef][Green Version]
- Calabrò, P.; Niccoli, G.; Gragnano, F.; Grove, E.L.; Vergallo, R.; Mikhailidis, D.P.; Patti, G.; Spaccarotella, C.; Katsiki, N.; Masiero, G.; et al. Are we ready for a gender-specific approach in interventional cardiology. Int. J. Cardiol. 2019, 286, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Gul, B.; Kozuma, K.; Haimi, I.; Zhao, W.; Simonton, C.; Ying, S.-W.; Buda, A.; Mehta, S.; Baumbach, A.; Lansky, A. Sex disparities in acute myocardial infarction care and outcomes. Catheter. Cardiovasc. Interv. 2018, 92, E341–E347. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Variable | Women | Men | p-Value |
|---|---|---|---|
| Number (%) | 86 (43%) | 114 (57%) | - |
| Age (years) | 69 ± 12 | 64 ± 12 | 0.002 * |
| Smoking | 16 (18.6%) | 25 (21.9%) | <0.001 † |
| Hypertension | 50 (58.1%) | 57 (50%) | 0.25 † |
| Dyslipidemia | 17 (19.8%) | 9 (7.9) | 0.013 † |
| Type 2 diabetes mellitus | 18 (20.9%) | 26 (22.8%) | 0.75 † |
| Positive family history of CVD | 29 (25.4%) | 18 (20.9%) | 0.18 † |
| History of former CVD | 82 (71.9%) | 61 (70.9%) | 0.33 † |
| Systolic blood pressure (mm Hg) | 127.55 ± 18.21 | 126.14 ± 13.89 | 0.97 |
| Diastolic blood pressure (mm Hg) | 75.77 ± 10.86 | 77.27 ± 9.25 | 0.46 |
| BMI (kg/m2) | 29.31 ± 4.19 | 28.30 ± 5.07 | 0.09 |
| Gene | Codominant Model | Dominant Model | Recessive Model | ||||||
|---|---|---|---|---|---|---|---|---|---|
| p | OR | 95% CI | p | OR | 95% CI | p | OR | 95% CI | |
| ARNTL | |||||||||
| rs3789327 | 0.18 | 0.56 | 0.29–1.07 | 0.06 | 0.56 | 0.30–1.04 | 0.56 | 0.80 | 0.37–1.70 |
| rs4757144 | 0.48 | 1.28 | 0.69–2.39 | 0.67 | 1.13 | 0.63–2.02 | 0.36 | 0.69 | 0.31–1.52 |
| rs12363415 | 0.77 | 1.05 | 0.56–1.99 | 0.98 | 1.01 | 0.54–1.88 | 0.48 | 0.43 | 0.04–4.89 |
| CLOCK | |||||||||
| rs11932595 | 0.08 | 0.93 | 0.50–1.72 | 0.67 | 1.14 | 0.63–2.05 | 0.03 | 2.66 | 1.07–6.66 |
| rs6811520 | 0.45 | 0.78 | 0.42–1.46 | 0.28 | 0.72 | 0.40–1.31 | 0.33 | 0.68 | 0.31–1.47 |
| rs13124436 | 0.95 | 0.97 | 0.53–1.77 | 0.99 | 1.00 | 0.57–1.77 | 0.76 | 1.15 | 0.46–2.87 |
| CRY2 | |||||||||
| rs2292912 | 0.99 | 0.97 | 0.54–1.76 | 0.94 | 0.98 | 0.55–1.75 | 0.96 | 1.05 | 0.17–6.60 |
| rs10838524 | 0.30 | 1.46 | 0.75–1.97 | 0.46 | 1.26 | 0.68–2.36 | 0.28 | 0.67 | 0.33–1.38 |
| PER2 | |||||||||
| rs35333999 | 0.89 | 1.24 | 0.42–3.72 | 0.79 | 1.15 | 0.41–3.21 | 0.77 | 0.66 | 0.04–10.76 |
| rs934945 | 0.43 | 1.23 | 0.64–2.34 | 0.79 | 1.09 | 0.59–2.01 | 0.25 | 0.39 | 0.07–2.11 |
| Risk Factor | OR (95% CI) | p Value |
|---|---|---|
| Age | 0.96 (0.94–0.99) | 0.017 |
| Smoking | 2.24 (1.45–3.47) | <0.001 |
| Hypertension | 1.06 (0.53–1.44) | 0.86 |
| Dyslipidemia | 2.36 (0.85–6.56) | 0.10 |
| Type 2 diabetes mellitus | 0.85 (0.37–1.95) | 0.71 |
| Positive family history of CVD | 1.20 (0.69–2.09) | 0.51 |
| History of former CVD | 0.92 (0.59–1.44) | 0.72 |
| Systolic blood pressure (mm Hg) | 0.97 (0.94–1.00) | 0.07 |
| Diastolic blood pressure (mm Hg) | 1.05 (1.01–1.11) | 0.045 |
| BMI (kg/m2) | 0.96 (0.89–1.03) | 0.27 |
| Gene | SNP | Minor Allele | MAF * Women | MAF * Men | p-Value | q Value | Genotype | Genotype Frequency, N (%) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Women | Men | p Value | Χ2 | q Value | ||||||||
| ARNTL | rs3789327 | C | 0.46 | 0.38 | 0.118 | 0.125 | TT | 23 (26.7%) | 44 (38.5%) | 0.210 | 3.11 | 0.216 |
| TC | 47 (54.6%) | 53 (46.4%) | ||||||||||
| CC | 16 (18.6%) | 17 (14.9%) | ||||||||||
| rs4757144 | G | 0.38 | 0.37 | 0.823 | 0.835 | AA | 35 (40.6%) | 44 (38.5%) | 0.578 | 1.09 | 0.577 | |
| AG | 36 (41.8%) | 55 (48.2%) | ||||||||||
| GG | 15 (17.4%) | 15 (13.1%) | ||||||||||
| rs12363415 | G | 0.16 | 0.15 | 0.828 | 0.888 | AA | 61 (70.9%) | 81 (71%) | 0.698 | 0.72 | 0.757 | |
| AG | 23 (26.7%) | 32 (28%) | ||||||||||
| GG | 2 (2.3%) | 1 (0.8%) | ||||||||||
| CLOCK | rs11932595 | G | 0.35 | 0.41 | 0.214 | 0.255 | AA | 32 (37.2%) | 41 (35.9%) | 0.103 | 4.53 | 0.102 |
| AG | 47 (54.6%) | 52 (45.6%) | ||||||||||
| GG | 7 (8.1%) | 21 (18.4%) | ||||||||||
| rs6811520 | T | 0.42 | 0.36 | 0.220 | 0.254 | CC | 29 (33.7%) | 46 (40.3%) | 0.465 | 1.53 | 0.486 | |
| CT | 41 (47.6%) | 53 (46.4%) | ||||||||||
| TT | 16 (18.6%) | 15 (13.1%) | ||||||||||
| rs13124436 | A | 0.32 | 0.33 | 0.846 | 0.914 | AA | 9 (10.4%) | 13 (11.4%) | 0.976 | 0.05 | 1 | |
| AG | 37 (43%) | 49 (42.9%) | ||||||||||
| GG | 40 (46.5%) | 52 (45.6%) | ||||||||||
| CRY2 | rs2292912 | G | 0.20 | 0.20 | 1 | 1 | CC | 53 (61.6%) | 71 (62.2%) | 0.982 | 0.03 | 0.999 |
| CG | 31 (36%) | 40 (35%) | ||||||||||
| GG | 2 (2.3%) | 3 (2.6%) | ||||||||||
| rs10838524 | A | 0.45 | 0.45 | 1 | 1 | GG | 27 (31.3%) | 30 (26.3%) | 0.327 | 2.23 | 0.317 | |
| GA | 40 (46.5%) | 65 (57%) | ||||||||||
| AA | 19 (22%) | 19 (16.6%) | ||||||||||
| PER2 | rs35333999 | T | 0.05 | 0.05 | 1 | 1 | CC | 79 (91.8%) | 104 (91.2%) | 0.952 | 0.09 | 0.999 |
| CT | 6 (6.9%) | 9 (7.8%) | ||||||||||
| TT | 1 (1.1%) | 1 (0.8%) | ||||||||||
| rs934945 | T | 0.18 | 0.16 | 0.722 | 0.789 | CC | 60 (69.7%) | 78 (68.4%) | 0.241 | 2.84 | 0.247 | |
| CT | 21 (24.4%) | 34 (29.8%) | ||||||||||
| TT | 5 (5.8%) | 2 (1.7%) | ||||||||||
| Gene | Age * | Smoking † | Hypertension † | Dyslipidemia † | Type 2 Diabetes Mellitus † | Positive Family History of CVD † | History of Former CVD † | SBP * | DBP * | BMI * | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Women | Men | Women | Men | Women | Men | Women | Men | Women | Men | Women | Men | Women | Men | Women | Men | Women | Men | Women | Men | |
| ARNTL | ||||||||||||||||||||
| rs3789327 | 0.425 | 0.391 | 0.011 | 0.895 | 0.118 | 0.406 | 0.233 | 0.314 | 0.182 | 0.339 | 0.181 | 0.088 | 0.071 | 0.034 | 0.330 | 0.209 | 0.348 | 0.475 | 0.607 | 0.183 |
| rs4757144 | 0.264 | 0.111 | 0.718 | 0.182 | 0.187 | 0.383 | 0.199 | 0.477 | 0.642 | 0.184 | 0.057 | 0.172 | 0.038 | 0.527 | 0.696 | 0.729 | 0.297 | 0.164 | 0.106 | 0.609 |
| rs12363415 | 0.634 | 0.587 | 0.737 | 0.595 | 0.170 | 0.566 | 0.094 | 0.875 | 0.003 | 0.848 | 0.152 | 0.992 | 0.122 | 0.443 | 0.931 | 0.831 | 0.318 | 0.927 | 0.523 | 0.774 |
| CLOCK | ||||||||||||||||||||
| rs11932595 | 0.190 | 0.567 | 0.426 | 0.937 | 0.269 | 0.617 | 0.173 | 0.060 | 0.798 | 0.224 | 0.276 | 0.702 | 0.693 | 0.724 | 0.005 | 0.157 | 0.006 | 0.275 | 0.996 | 0.024 |
| rs6811520 | 0.439 | 0.169 | 0.304 | 0.691 | 0.687 | 0.560 | 0.399 | 0.431 | 0.378 | 0.693 | 0.389 | 0.463 | 0.313 | 0.501 | 0.499 | 0.327 | 0.592 | 0.161 | 0.334 | 0.219 |
| rs13124436 | 0.891 | 0.561 | 0.651 | 0.924 | 0.215 | 0.527 | 0.772 | 0.514 | 0.980 | 0.168 | 0.355 | 0.849 | 0.443 | 0.241 | 0.895 | 0.856 | 0.869 | 0.983 | 0.191 | 0.546 |
| CRY2 | ||||||||||||||||||||
| rs2292912 | 0.818 | 0.366 | 0.811 | 0.562 | 0.169 | 0.178 | 0.768 | 0.020 | 0.188 | 0.908 | 0.840 | 0.579 | 0.139 | 0.303 | 0.749 | 0.503 | 0.750 | 0.515 | 0.531 | 0.834 |
| rs10838524 | 0.135 | 0.544 | 0.215 | 0.776 | 0.338 | 0.138 | 0.288 | 0.175 | 0.606 | 0.224 | 0.879 | 0.575 | 0.095 | 0.857 | 0.202 | 0.814 | 0.500 | 0.844 | 0.274 | 0.080 |
| PER2 | ||||||||||||||||||||
| rs35333999 | 0.194 | 0.035 | 0.101 | 0.632 | 0.624 | 0.574 | 0.391 | 0.896 | 0.365 | 0.860 | 0.274 | 0.029 | 0.713 | 0.525 | 0.607 | 0.149 | 0.368 | 0.511 | 0.423 | 0.217 |
| rs934945 | 0.457 | 0.096 | 0.529 | 0.107 | 0.146 | 0.034 | 0.499 | <0.001 | 0.933 | 0.162 | 0.819 | 0.785 | 0.129 | 0.004 | 0.305 | 0.195 | 0.264 | 0.668 | 0.345 | 0.449 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Škrlec, I.; Talapko, J.; Juzbašić, M.; Steiner, R. Sex Differences in Circadian Clock Genes and Myocardial Infarction Susceptibility. J. Cardiovasc. Dev. Dis. 2021, 8, 53. https://doi.org/10.3390/jcdd8050053
Škrlec I, Talapko J, Juzbašić M, Steiner R. Sex Differences in Circadian Clock Genes and Myocardial Infarction Susceptibility. Journal of Cardiovascular Development and Disease. 2021; 8(5):53. https://doi.org/10.3390/jcdd8050053
Chicago/Turabian StyleŠkrlec, Ivana, Jasminka Talapko, Martina Juzbašić, and Robert Steiner. 2021. "Sex Differences in Circadian Clock Genes and Myocardial Infarction Susceptibility" Journal of Cardiovascular Development and Disease 8, no. 5: 53. https://doi.org/10.3390/jcdd8050053
APA StyleŠkrlec, I., Talapko, J., Juzbašić, M., & Steiner, R. (2021). Sex Differences in Circadian Clock Genes and Myocardial Infarction Susceptibility. Journal of Cardiovascular Development and Disease, 8(5), 53. https://doi.org/10.3390/jcdd8050053