
Cellular and Molecular Mechanisms of Functional Hierarchy of Pacemaker Clusters in the Sinoatrial Node: New Insights into Sick Sinus Syndrome

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Development of the SAN
2.1. Structural Changes in the SAN Development
2.2. Genetic Programing Controlling SAN Development
3. Concept of Hierarchical Pacemaker Clustering in the SAN
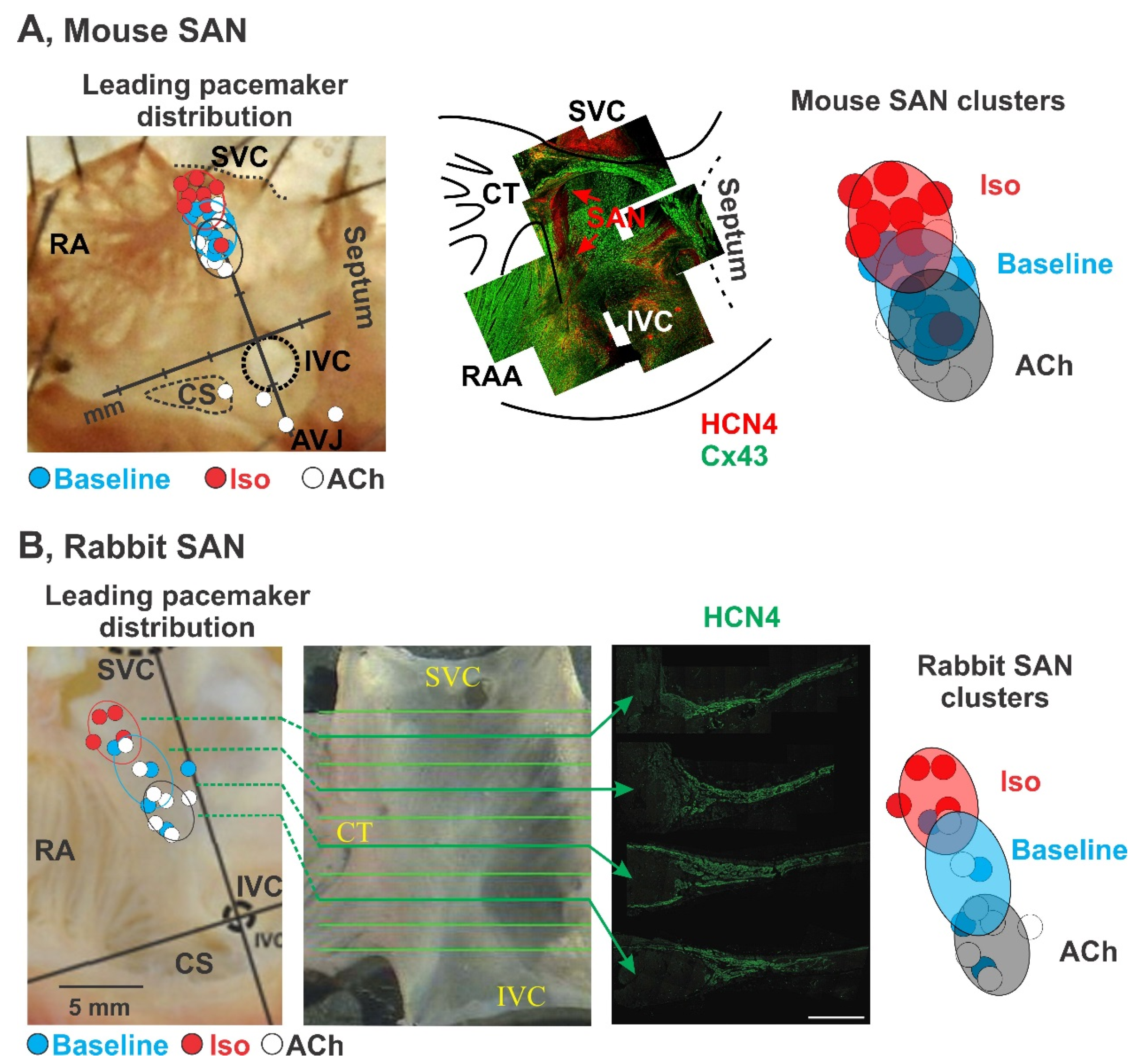
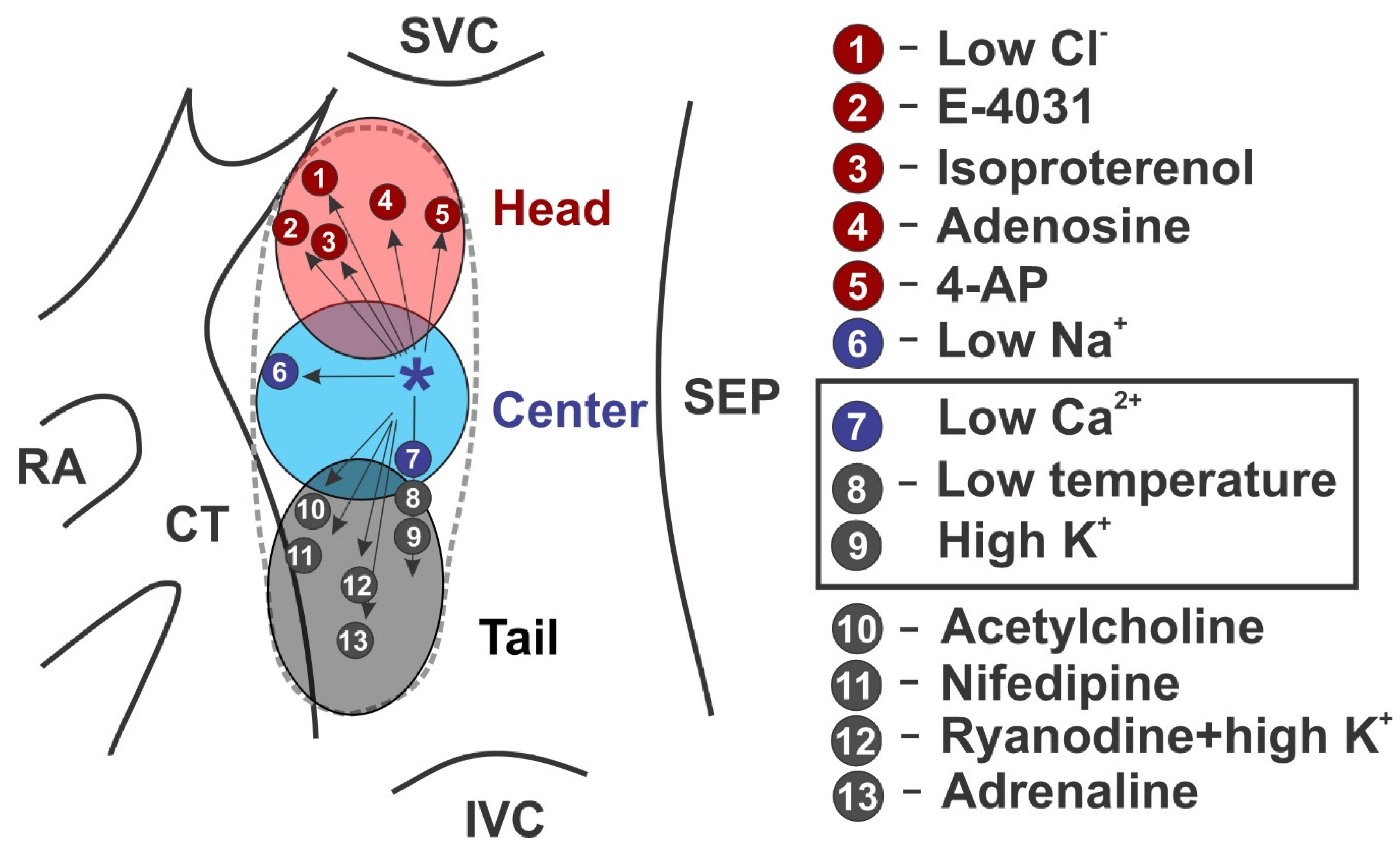
3.1. Pacemaker Shift
3.2. Dynamic Hierarchy of Cluster Pacemaking
3.3. Synchronization of the Hierarchical Pacemaker Clusters
4. Functional and Molecular Signatures of SAN Pacemaker Clusters
4.1. Fine Architecture
4.2. Electrophysiological Heterogeneity
4.3. Pacemaking Mechanisms: Voltage Versus Calcium Clocks
4.4. Autonomic Innervation
5. Hierarchical Pacemaker Clusters in Sinus Node Dysfunction
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bleeker, W.K.; Mackaay, A.J.; Masson-Pévet, M.; Bouman, L.N.; Becker, A.E. Functional and morphological organization of the rabbit sinus node. Circ. Res. 1980, 46, 11–22. [Google Scholar] [CrossRef]
- Boyett, M.R.; Dobrzynski, H.; Lancaster, M.K.; Jones, S.A.; Honjo, H.; Kodama, I. Sophisticated architecture is required for the sinoatrial node to perform its normal pacemaker function. J. Cardiovasc. Electrophysiol. 2003, 14, 104–106. [Google Scholar] [CrossRef]
- Opthof, T. The mammalian sinoatrial node. Cardiovasc. Drugs Ther. 1988, 1, 573–597. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Hansen, B.J.; Csepe, T.A.; Zhao, J.; Ignozzi, A.J.; Sul, L.V.; Zakharkin, S.O.; Kalyanasundaram, A.; Davis, J.P.; Biesiadecki, B.J.; et al. Redundant and diverse intranodal pacemakers and conduction pathways protect the human sinoatrial node from failure. Sci. Transl. Med. 2017, 9, eaam5607. [Google Scholar] [CrossRef]
- Boyett, M.; Honjo, H.; Kodama, I. The sinoatrial node, a heterogeneous pacemaker structure. Cardiovasc. Res. 2000, 47, 658–687. [Google Scholar] [CrossRef]
- Shibata, N.; Inada, S.; Mitsui, K.; Honjo, H.; Yamamoto, M.; Niwa, R.; Boyett, M.R.; Kodama, I. Pacemaker shift in the rabbit sinoatrial node in response to vagal nerve stimulation. Exp. Physiol. 2001, 86, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Petrov, V.; Lou, Q.; Osipov, G.; Efimov, I.R. Spatiotemporal control of heart rate in a rabbit heart. J. Electrocardiol. 2011, 44, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Guan, X.; Luczak, E.D.; Lang, D.; Kutschke, W.; Gao, Z.; Yang, J.; Glynn, P.; Sossalla, S.; Swaminathan, P.D.; et al. Diabetes increases mortality after myocardial infarction by oxidizing CaMKII. J. Clin. Investig. 2013, 123, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- Monfredi, O.; Tsutsui, K.; Ziman, B.D.; Stern, M.D.; Lakatta, E.G.; Maltsev, V.A. Electrophysiological heterogeneity of pacemaker cells in the rabbit intercaval region, including the SA node: Insights from recording multiple ion currents in each cell. Am. J. Physiol. Circ. Physiol. 2018, 314, H403–H414. [Google Scholar] [CrossRef]
- Yuan, X.; Ratajczyk, L.N.; Alvarado, F.; Valdivia, H.H.; Glukhov, A.V.; Lang, D. Hierarchical Pacemaker Clustering within the Rabbit Sinoatrial Node is Driven by Dynamic Interaction between the Components of the Coupled-Clock System. Biophys. J. 2020, 118, 345a. [Google Scholar] [CrossRef]
- Van Mierop, L.H. Location of pacemaker in chick embryo heart at the time of initiation of heartbeat. Am. J. Physiol. 1967, 212, 407–415. [Google Scholar] [CrossRef]
- Kamino, K. Optical approaches to ontogeny of electrical activity and related functional organization during early heart development. Physiol. Rev. 1991, 71, 53–91. [Google Scholar] [CrossRef]
- Paff, G.H.; Boucek, R.J.; Harrell, T.C. Observations on the development of the electrocardiogram. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1968, 160, 575–581. [Google Scholar] [CrossRef]
- Zhu, W.-Z.; Xie, Y.; Moyes, K.W.; Gold, J.D.; Askari, B.; Laflamme, M.A. Neuregulin/ErbB Signaling Regulates Cardiac Subtype Specification in Differentiating Human Embryonic Stem Cells. Circ. Res. 2010, 107, 776–786. [Google Scholar] [CrossRef]
- Hashem, S.I.; Claycomb, W.C. Genetic isolation of stem cell-derived pacemaker-nodal cardiac myocytes. Mol. Cell. Biochem. 2013, 383, 161–171. [Google Scholar] [CrossRef]
- Zhang, H.; Lau, D.H.; Shlapakova, I.N.; Zhao, X.; Danilo, P.; Robinson, R.B.; Cohen, I.S.; Qu, D.; Xu, Z.; Rosen, M.R. Implantation of Sinoatrial Node Cells into Canine Right Ventricle: Biological Pacing Appears Limited by the Substrate; SAGE Publications: Los Angeles, CA, USA, 2011. [Google Scholar]
- Van Mierop, L.; Gessner, I.H. The morphologic development of the sinoatrial node in the mouse. Am. J. Cardiol. 1970, 25, 204–212. [Google Scholar] [CrossRef]
- Christoffels, V.M.; Smits, G.J.; Kispert, A.; Moorman, A.F.M. Development of the Pacemaker Tissues of the Heart. Circ. Res. 2010, 106, 240–254. [Google Scholar] [CrossRef]
- Dickinson, D.F.; Wilkinson, J.L.; Anderson, K.R.; Smith, A.; Ho, S.Y.; Anderson, R.H. The cardiac conduction system in situs ambiguus. Circulation 1979, 59, 879–885. [Google Scholar] [CrossRef]
- Yamamoto, M.; Dobrzynski, H.; Tellez, J.; Niwa, R.; Billeter, R.; Honjo, H.; Kodama, I.; Boyett, M.R. Extended atrial conduction system characterised by the expression of the HCN4 channel and connexin45. Cardiovasc. Res. 2006, 72, 271–281. [Google Scholar] [CrossRef]
- Liu, J.; Dobrzynski, H.; Yanni, J.; Boyett, M.R.; Lei, M. Organisation of the mouse sinoatrial node: Structure and expression of HCN channels. Cardiovasc. Res. 2007, 73, 729–738. [Google Scholar] [CrossRef]
- Brioschi, C.; Micheloni, S.; Tellez, J.O.; Pisoni, G.; Longhi, R.; Moroni, P.; Billeter, R.; Barbuti, A.F.; Dobrzynski, H.; Boyett, M.R.; et al. Distribution of the pacemaker HCN4 channel mRNA and protein in the rabbit sinoatrial node. J. Mol. Cell. Cardiol. 2009, 47, 221–227. [Google Scholar] [CrossRef]
- Später, D.; Abramczuk, M.K.; Buac, K.; Zangi, L.; Stachel, M.W.; Clarke, J.; Sahara, M.; Ludwig, A.; Chien, K.R. A HCN4+ cardiomyogenic progenitor derived from the first heart field and human pluripotent stem cells. Nat. Cell Biol. 2013, 15, 1098–1106. [Google Scholar] [CrossRef]
- Garcia-Frigola, C.; Shi, Y.; Evans, S.M. Expression of the hyperpolarization-activated cyclic nucleotide-gated cation channel HCN4 during mouse heart development. Gene Expr. Patterns 2003, 3, 777–783. [Google Scholar] [CrossRef]
- Stieber, J.; Herrmann, S.; Feil, S.; Löster, J.; Feil, R.; Biel, M.; Hofmann, F.; Ludwig, A. The hyperpolarization-activated channel HCN4 is required for the generation of pacemaker action potentials in the embryonic heart. Proc. Natl. Acad. Sci. USA 2003, 100, 15235–15240. [Google Scholar] [CrossRef]
- Gorza, L.; Schiaffino, S.; Vitadello, M. Heart conduction system: A neural crest derivative? Brain Res. 1988, 457, 360–366. [Google Scholar] [CrossRef]
- Van Kempen, M.J.; Fromaget, C.; Gros, D.; Moorman, A.F.; Lamers, W.H. Spatial distribution of connexin43, the major cardiac gap junction protein, in the developing and adult rat heart. Circ. Res. 1991, 68, 1638–1651. [Google Scholar] [CrossRef]
- Barbuti, A.F.; Robinson, R.B. Stem Cell–Derived Nodal-Like Cardiomyocytes as a Novel Pharmacologic Tool: Insights from Sinoatrial Node Development and Function. Pharmacol. Rev. 2015, 67, 368–388. [Google Scholar] [CrossRef]
- Hoogaars, W.M.; Tessari, A.; Moorman, A.F.; de Boer, P.A.; Hagoort, J.; Soufan, A.T.; Campione, M.; Christoffels, V.M. The transcriptional repressor Tbx3 delineates the developing central conduction system of the heart. Cardiovasc. Res. 2004, 62, 489–499. [Google Scholar] [CrossRef]
- Hoogaars, W.M.H.; Engel, A.; Brons, J.F.; Verkerk, A.O.; De Lange, F.J.; Wong, L.Y.E.; Bakker, M.L.; Clout, D.E.; Wakker, V.; Barnett, P.; et al. Tbx3 controls the sinoatrial node gene program and imposes pacemaker function on the atria. Genes Dev. 2007, 21, 1098–1112. [Google Scholar] [CrossRef]
- Espinoza-Lewis, R.A.; Yu, L.; He, F.; Liu, H.; Tang, R.; Shi, J.; Sun, X.; Martin, J.F.; Wang, D.; Yang, J.; et al. Shox2 is essential for the differentiation of cardiac pacemaker cells by repressing Nkx2-5. Dev. Biol. 2009, 327, 376–385. [Google Scholar] [CrossRef]
- Wiese, C.; Grieskamp, T.; Airik, R.; Mommersteeg, M.T.M.; Gardiwal, A.; Vries, C.D.G.-D.; Schuster-Gossler, K.; Moorman, A.F.M.; Kispert, A.; Christoffels, V.M. Formation of the Sinus Node Head and Differentiation of Sinus Node Myocardium Are Independently Regulated by Tbx18 and Tbx3. Circ. Res. 2009, 104, 388–397. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, Q.; Cattaneo, P.; Zhuang, S.; Gong, X.; Spann, N.J.; Jiang, C.; Cao, X.; Zhao, X.; Zhang, X.; et al. Transcription factor ISL1 is essential for pacemaker development and function. J. Clin. Investig. 2015, 125, 3256–3268. [Google Scholar] [CrossRef] [PubMed]
- Galang, G.; Mandla, R.; Ruan, H.; Jung, C.; Sinha, T.; Stone, N.R.; Wu, R.S.; Mannion, B.J.; Allu, P.K.; Chang, K.; et al. ATAC-Seq Reveals an Isl1 Enhancer That Regulates Sinoatrial Node Development and Function. Circ. Res. 2020, 127, 1502–1518. [Google Scholar] [CrossRef]
- Truex, R.C.; Smythe, M.Q.; Taylor, M.J. Reconstruction of the human sinoatrial node. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1967, 159, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Woods, W.T.; Urthaler, F.; James, T.N. Spontaneous action potentials of cells in the canine sinus node. Circ. Res. 1976, 39, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Yen, H.S.; Becker, A.E.; Gosling, J.A. The Development of the Sinoatrial Node. In The Sinus Node; Springer: Berlin/Heidelberg, Germany, 1978; pp. 166–182. [Google Scholar]
- Anderson, R.H.; Ho, S.Y. The architecture of the sinus node, the atrioventricular conduction axis, and the internodal atrial myocardium. J. Cardiovasc. Electrophysiol. 1998, 9, 1233–1248. [Google Scholar] [CrossRef] [PubMed]
- Kreitner, D. Electrophysiological study of the two main pacemaker mechanisms in the rabbit sinus node. Cardiovasc. Res. 1985, 19, 304–318. [Google Scholar] [CrossRef]
- Bouman, L.N.; Gerlings, E.D.; Biersteker, P.A.; Bonke, F.I.M. Pacemaker shift in the sino-atrial node during vagal stimulation. Pflüger Arch. 1968, 302, 255–267. [Google Scholar] [CrossRef]
- Bouman, L.; Mackaay, A.; Bleeker, W.; Becher, A. Pacemaker Shifts in the Sinus Node: Effects of Vagal Stimulation, Temperature and Reduction of Extracellular Calcium. In The Sinus Node; Springer: Berlin/Heidelberg, Germany, 1978; pp. 245–257. [Google Scholar]
- Glukhov, A.V.; Hage, L.T.; Hansen, B.J.; Pedraza-Toscano, A.; Vargas-Pinto, P.; Hamlin, R.L.; Weiss, R.; Carnes, C.A.; Billman, G.E.; Fedorov, V.V. Sinoatrial node reentry in a canine chronic left ventricular infarct model: Role of intranodal fibrosis and heterogeneity of refractoriness. Circ. Arrhythmia Electrophysiol. 2013, 6, 984–994. [Google Scholar] [CrossRef]
- Shinohara, T.; Park, H.W.; Joung, B.; Maruyama, M.; Chua, S.K.; Han, S.; Shen, M.J.; Chen, P.-S.; Lin, S.-F. Selective sinoatrial node optical mapping and the mechanism of sinus rate acceleration. Circ. J. 2011, 1111041458. [Google Scholar] [CrossRef]
- Joung, B.; Shinohara, T.; Zhang, H.; Kim, D.; Choi, E.-K.; On, Y.-K.; Piccirillo, G.; Chen, P.-S.; Lin, S.-F. Tachybradycardia in the isolated canine right atrium induced by chronic sympathetic stimulation and pacemaker current inhibition. Am. J. Physiol. Circ. Physiol. 2010, 299, H634–H642. [Google Scholar] [CrossRef]
- Ashton, J.L.; Trew, M.L.; LeGrice, I.J.; Paterson, D.J.; Paton, J.F.; Gillis, A.M.; Smaill, B.H. Shift of leading pacemaker site during reflex vagal stimulation and altered electrical source-to-sink balance. J. Physiol. 2019, 597, 3297–3313. [Google Scholar] [CrossRef]
- Schuessler, R.B.; Boineau, J.P.; Bromberg, B.I. Origin of the Sinus Impulse. J. Cardiovasc. Electrophysiol. 1996, 7, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.R.; Goldberg, J.M. P-wave morphology and atrial activation in the domestic fowl. Am. J. Physiol. 1980, 239, R483–R488. [Google Scholar] [CrossRef] [PubMed]
- Glukhov, A.V.; Fedorov, V.V.; Anderson, M.E.; Mohler, P.J.; Efimov, I.R. Functional anatomy of the murine sinus node: High-resolution optical mapping of ankyrin-B heterozygous mice. Am. J. Physiol. Circ. Physiol. 2010, 299, H482–H491. [Google Scholar] [CrossRef] [PubMed]
- Glukhov, A.V.; Kalyanasundaram, A.; Lou, Q.; Hage, L.T.; Hansen, B.J.; Belevych, A.E.; Mohler, P.J.; Knollmann, B.C.; Periasamy, M.; Györke, S.; et al. Calsequestrin 2 deletion causes sinoatrial node dysfunction and atrial arrhythmias associated with altered sarcoplasmic reticulum calcium cycling and degenerative fibrosis within the mouse atrial pacemaker complex1. Eur. Heart J. 2013, 36, 686–697. [Google Scholar] [CrossRef]
- Lou, Q.; Glukhov, A.V.; Hansen, B.; Hage, L.; Vargas-Pinto, P.; Billman, G.E.; Carnes, C.A.; Fedorov, V.V. Tachy-brady arrhythmias: The critical role of adenosine-induced sinoatrial conduction block in post-tachycardia pauses. Heart Rhythm 2013, 10, 110–118. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Joung, B.; Tang, L.; Maruyama, M.; Han, S.; Chen, Z.; Stucky, M.; Jones, L.R.; Fishbein, M.C.; Weiss, J.N.; Chen, P.-S.; et al. Intracellular Calcium Dynamics and the Acceleration of Sinus Rhythm by β-Adrenergic Stimulation. Circulation 2009, 119, 788. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, V.V.; Chang, R.; Glukhov, A.V.; Kostecki, G.; Janks, D.; Schuessler, R.B.; Efimov, I.R. Complex Interactions Between the Sinoatrial Node and Atrium During Reentrant Arrhythmias in the Canine Heart. Circulation 2010, 122, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Sulkin, M.; Lou, Q.; Efimov, I.R. Optical Mapping of Action Potentials and Calcium Transients in the Mouse Heart. J. Vis. Exp. 2011, 2011, e3275. [Google Scholar] [CrossRef]
- Lang, D.; Holzem, K.; Kang, C.; Xiao, M.; Hwang, H.J.; Ewald, G.A.; Yamada, K.A.; Efimov, I.R. Arrhythmogenic remodeling of beta2 versus beta1 adrenergic signaling in the human failing heart. Circ. Arrhythm Electrophysiol. 2015, 8, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Glukhov, A.V. High-resolution Optical Mapping of the Mouse Sino-atrial Node. J. Vis. Exp. 2016, 2016, e54773. [Google Scholar] [CrossRef] [PubMed]
- Janardhan, A.H.; Lang, D.; Efimov, I.R. Optical Mapping of the Sinoatrial Node and Atrioventricular Node. Card. Mapp. 2012, 79–89. [Google Scholar] [CrossRef]
- Steinbeck, G.; Bonke, F.I.; Allessie, M.A.; Lammers, W.J. The effect of ouabain on the isolated sinus node preparation of the rabbit studied with microelectrodes. Circ. Res. 1980, 46, 406–414. [Google Scholar] [CrossRef]
- Kodama, I.; Nikmaram, M.R.; Boyett, M.R.; Suzuki, R.; Honjo, H.; Owen, J.M. Regional differences in the role of the Ca2+ and Na+ currents in pacemaker activity in the sinoatrial node. Am. J. Physiol. Circ. Physiol. 1997, 272, H2793–H2806. [Google Scholar] [CrossRef]
- Massonpevet, M.; Bleeker, W.; Besselsen, E.; Treytel, B.; Jongsma, H.; Bouman, L. Pacemaker cell types in the rabbit sinus node: A correlative ultrastructural and electrophysiological study. J. Mol. Cell. Cardiol. 1984, 16, 53–63. [Google Scholar] [CrossRef]
- Fedorov, V.V.; Glukhov, A.V.; Chang, R. Conduction barriers and pathways of the sinoatrial pacemaker complex: Their role in normal rhythm and atrial arrhythmias. Am. J. Physiol. Circ. Physiol. 2012, 302, H1773–H1783. [Google Scholar] [CrossRef]
- Fedorov, V.V.; Glukhov, A.V.; Chang, R.; Kostecki, G.; Aferol, H.; Hucker, W.J.; Wuskell, J.P.; Loew, L.M.; Schuessler, R.B.; Moazami, N.; et al. Optical Mapping of the Isolated Coronary-Perfused Human Sinus Node. J. Am. Coll. Cardiol. 2010, 56, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Fenske, S.; Hennis, K.; Rötzer, R.D.; Brox, V.F.; Becirovic, E.; Scharr, A.; Gruner, C.; Ziegler, T.; Mahlfeld, V.; Brennan, J.; et al. cAMP-dependent regulation of HCN4 controls the tonic entrainment process in sinoatrial node pacemaker cells. Nat. Commun. 2020, 11, 5555. [Google Scholar] [CrossRef]
- Jalife, J. Mutual entrainment and electrical coupling as mechanisms for synchronous firing of rabbit sino-atrial pace-maker cells. J. Physiol. 1984, 356, 221–243. [Google Scholar] [CrossRef]
- Michaels, D.C.; Matyas, E.P.; Jalife, J. Dynamic interactions and mutual synchronization of sinoatrial node pacemaker cells. A mathematical model. Circ. Res. 1986, 58, 706–720. [Google Scholar] [CrossRef] [PubMed]
- Honjo, H.; Boyett, M.R.; Kodama, I.; Toyama, J. Correlation between electrical activity and the size of rabbit sino-atrial node cells. J. Physiol. 1996, 496, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Oosthoek, P.W.; Virágh, S.; Mayen, A.E.; Van Kempen, M.J.; Lamers, W.H.; Moorman, A.F. Immunohistochemical delineation of the conduction system. I: The sinoatrial node. Circ. Res. 1993, 73, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Masson-Pévet, M.; Bleeker, W.K.; Gros, D. The plasma membrane of leading pacemaker cells in the rabbit sinus node. A qualitative and quantitative ultrastructural analysis. Circ. Res. 1979, 45, 621–629. [Google Scholar] [CrossRef]
- James, T.N.; Sherf, L.; Fine, G.; Morales, A.R. Comparative Ultrastructure of the Sinus Node in Man and Dog. Circulation 1966, 34, 139–163. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Glukhov, A.V. Functional Microdomains in Heart’s Pacemaker: A Step beyond Classical Electrophysiology and Remodeling. Front. Physiol. 2018, 9, 1686. [Google Scholar] [CrossRef] [PubMed]
- Musa, H.; Lei, M.; Honjo, H.; Jones, S.A.; Dobrzynski, H.; Lancaster, M.K.; Takagishi, Y.; Henderson, Z.; Kodama, I.; Boyett, M.R. Heterogeneous Expression of Ca2+Handling Proteins in Rabbit Sinoatrial Node. J. Histochem. Cytochem. 2002, 50, 311–324. [Google Scholar] [CrossRef]
- Boyett, M.R.; Honjo, H.; Yamamoto, M.; Nikmaram, M.R.; Niwa, R.; Kodama, I. Downward gradient in action potential duration along conduction path in and around the sinoatrial node. Am. J. Physiol. Circ. Physiol. 1999, 276, H686–H698. [Google Scholar] [CrossRef]
- Kodama, I.; Boyett, M.R.; Nikmaram, M.R.; Yamamoto, M.; Honjo, H.; Niwa, R. Regional differences in effects of E-4031 within the sinoatrial node. Am. J. Physiol. 1999, 276, H793–H802. [Google Scholar] [CrossRef]
- Glukhov, A.V.; Balycheva, M.; Sanchez-Alonso, J.L.; Ilkan, Z.; Alvarez-Laviada, A.; Bhogal, N.; Diakonov, I.; Schobesberger, S.; Sikkel, M.B.; Bhargava, A.; et al. Direct Evidence for Microdomain-Specific Localization and Remodeling of Functional L-Type Calcium Channels in Rat and Human Atrial Myocytes. Circulation 2015, 132, 2372–2384. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Sato, D.; Jiang, Y.; Ginsburg, K.S.; Ripplinger, C.M.; Bers, D.M. Calcium-Dependent Arrhythmogenic Foci Created by Weakly Coupled Myocytes in the Failing Heart. Circ. Res. 2017, 121, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, T.M.; Brochet, D.X.; Sirenko, S.; Li, Y.; Spurgeon, H.; Lakatta, E.G. Sarcoplasmic Reticulum Ca2+ Pumping Kinetics Regulates Timing of Local Ca2+ Releases and Spontaneous Beating Rate of Rabbit Sinoatrial Node Pacemaker Cells. Circ. Res. 2010, 107, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Maltsev, A.V.; Monfredi, O.; Maltseva, L.A.; Wirth, A.; Florio, M.C.; Tsutsui, K.; Riordon, D.R.; Parsons, S.P.; Tagirova, S.; et al. Heterogeneity of calcium clock functions in dormant, dysrhythmically and rhythmically firing single pacemaker cells isolated from SA node. Cell Calcium 2018, 74, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, K.; Monfredi, O.J.; Sirenko-Tagirova, S.G.; Maltseva, L.A.; Bychkov, R.; Kim, M.S.; Ziman, B.D.; Tarasov, K.V.; Tarasova, Y.S.; Zhang, J.; et al. A coupled-clock system drives the automaticity of human sinoatrial nodal pacemaker cells. Sci. Signal. 2018, 11, eaap7608. [Google Scholar] [CrossRef]
- Abramochkin, D.V.; Kuzmin, V.S.; Sukhova, G.S.; Rosenshtraukh, L.V. Modulation of rabbit sinoatrial node activation sequence by acetylcholine and isoproterenol investigated with optical mapping technique. Acta Physiol. 2009, 196, 385–394. [Google Scholar] [CrossRef]
- Li, N.; Csepe, T.A.; Hansen, B.J.; Dobrzynski, H.; Higgins, R.S.; Kilic, A.; Mohler, P.J.; Janssen, P.M.; Rosen, M.R.; Biesiadecki, B.J.; et al. Molecular Mapping of Sinoatrial Node HCN Channel Expression in the Human Heart. Circ. Arrhythmia Electrophysiol. 2015, 8, 1219–1227. [Google Scholar] [CrossRef]
- DiFrancesco, D. Serious workings of the funny current. Prog. Biophys. Mol. Biol. 2006, 90, 13–25. [Google Scholar] [CrossRef]
- Vinogradova, T.M.; Lyashkov, A.E.; Zhu, W.; Ruknudin, A.M.; Sirenko, S.; Yang, D.; Deo, S.; Barlow, M.; Johnson, S.; Caffrey, J.L.; et al. High Basal Protein Kinase A–Dependent Phosphorylation Drives Rhythmic Internal Ca2+ Store Oscillations and Spontaneous Beating of Cardiac Pacemaker Cells. Circ. Res. 2006, 98, 505–514. [Google Scholar] [CrossRef]
- Li, Y.; Sirenko, S.; Riordon, D.R.; Yang, D.; Spurgeon, H.; Lakatta, E.G.; Vinogradova, T.M. CaMKII-dependent phosphorylation regulates basal cardiac pacemaker function via modulation of local Ca2+ releases. Am. J. Physiol.-Heart Circ. Physiol. 2016, 311, H532–H544. [Google Scholar] [CrossRef]
- Liu, J.; Sirenko, S.; Juhaszova, M.; Ziman, B.; Shetty, V.; Rain, S.; Shukla, S.; Spurgeon, H.A.; Vinogradova, T.M.; Maltsev, V.A.; et al. A full range of mouse sinoatrial node AP firing rates requires protein kinase A-dependent calcium signaling. J. Mol. Cell. Cardiol. 2011, 51, 730–739. [Google Scholar] [CrossRef][Green Version]
- Boink, G.J.; Nearing, B.D.; Shlapakova, I.N.; Duan, L.; Kryukova, Y.; Bobkov, Y.; Tan, H.L.; Cohen, I.S.; Danilo, J.P.; Robinson, R.B.; et al. Ca2+ -Stimulated Adenylyl Cyclase AC1 Generates Efficient Biological Pacing as Single Gene Therapy and in Combination With HCN2. Circulation 2012, 126, 528–536. [Google Scholar] [CrossRef]
- Brandenburg, S.; Kohl, T.; Williams, G.S.; Gusev, K.; Wagner, E.; Rog-Zielinska, E.A.; Hebisch, E.; Dura, M.; Didié, M.; Gotthardt, M.; et al. Axial tubule junctions control rapid calcium signaling in atria. J. Clin. Investig. 2016, 126, 3999–4015. [Google Scholar] [CrossRef]
- Brandenburg, S.; Pawlowitz, J.; Fakuade, F.E.; Kownatzki-Danger, D.; Kohl, T.; Mitronova, G.Y.; Scardigli, M.; Neef, J.; Schmidt, C.; Wiedmann, F.; et al. Axial Tubule Junctions Activate Atrial Ca2+ Release Across Species. Front. Physiol. 2018, 9, 1227. [Google Scholar] [CrossRef]
- Ayettey, A.S.; Navaratnam, V. The T-tubule system in the specialized and general myocardium of the rat. J. Anat. 1978, 127, 125–140. [Google Scholar]
- Vinogradova, T.M.; Fedorov, V.V.; Yuzyuk, T.N.; Zaitsev, A.V.; Rosenshtraukh, L.V. Local Cholinergic Suppression of Pacemaker Activity in the Rabbit Sinoatrial Node. J. Cardiovasc. Pharmacol. 1998, 32, 413–424. [Google Scholar] [CrossRef]
- Fedorov, V.V.; Hucker, W.J.; Dobrzynski, H.; Rosenshtraukh, L.V.; Efimov, I.R. Postganglionic nerve stimulation induces temporal inhibition of excitability in rabbit sinoatrial node. Am. J. Physiol. Circ. Physiol. 2006, 291, H612–H623. [Google Scholar] [CrossRef]
- Lipsius, S.L.; Vassalle, M. Dual excitatory channels in the sinus node. J. Mol. Cell. Cardiol. 1978, 10, 753–767. [Google Scholar] [CrossRef]
- Lyashkov, A.E.; Juhaszova, M.; Dobrzynski, H.; Vinogradova, T.M.; Maltsev, V.A.; Juhasz, O.; Spurgeon, H.A.; Sollott, S.J.; Lakatta, E.G. Calcium Cycling Protein Density and Functional Importance to Automaticity of Isolated Sinoatrial Nodal Cells Are Independent of Cell Size. Circ. Res. 2007, 100, 1723–1731. [Google Scholar] [CrossRef]
- Keith, A.; MacKenzie, I. Recent researches on the anatomy of the heart. Lancet 1910, 175, 101–103. [Google Scholar] [CrossRef]
- Anderson, R.H. The disposition, morphology and innervation of cardiac specialized tissue in the guinea-pig. J. Anat. 1972, 111, 453–468. [Google Scholar]
- Wake, E.; Brack, K. Characterization of the intrinsic cardiac nervous system. Auton. Neurosci. 2016, 199, 3–16. [Google Scholar] [CrossRef]
- Lazzara, R.; Scherlag, B.J.; Robinson, M.J.; Samet, P. Selective In Situ Parasympathetic Control of the Canine Sinoatrial and Atrioventricular Nodes. Circ. Res. 1973, 32, 393–401. [Google Scholar] [CrossRef]
- Lyashkov, A.E.; Vinogradova, T.M.; Zahanich, I.; Li, Y.; Younes, A.; Nuss, H.B.; Spurgeon, H.A.; Maltsev, V.A.; Lakatta, E.G. Cholinergic receptor signaling modulates spontaneous firing of sinoatrial nodal cells via integrated effects on PKA-dependent Ca2+ cycling and I(KACh). Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H949–H959. [Google Scholar] [CrossRef]
- Roberts, L.A.; Slocum, G.R.; Riley, D.A. Morphological study of the innervation pattern of the rabbit sinoatrial node. Am. J. Anat. 1989, 185, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Beau, S.L.; Hand, D.E.; Schuessler, R.B.; Bromberg, B.I.; Kwon, B.; Boineau, J.P.; Saffitz, J.E. Relative densities of muscarinic cholinergic and beta-adrenergic receptors in the canine sinoatrial node and their relation to sites of pacemaker activity. Circ. Res. 1995, 77, 957–963. [Google Scholar] [CrossRef]
- Kurogouchi, F.; Nakane, T.; Furukawa, Y.; Hirose, M.; Inada, Y.; Chiba, S. Heterogeneous Distribution Of β-Adrenoceptors and Muscarinic Receptors In The Sinoatrial Node And Right Atrium Of The Dog. Clin. Exp. Pharmacol. Physiol. 2002, 29, 666–672. [Google Scholar] [CrossRef]
- Sutyagin, P.V.; Kalinina, E.E.; Pylaev, A.S. Relative Distribution Densities of Cholinergic and Adrenoceptor Structures in the Central Part of the Sinoatrial Node in Rat Heart. Bull. Exp. Biol. Med. 2005, 140, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Uijtdehaage, S.H.J.; Thayer, J.F. Accentuated antagonism in the control of human heart rate. Clin. Auton. Res. 2000, 10, 107–110. [Google Scholar] [CrossRef]
- Stramba-Badiale, M.; Vanoli, E.; De Ferrari, G.M.; Cerati, D.; Foreman, R.D.; Schwartz, P.J. Sympathetic-parasympathetic interaction and accentuated antagonism in conscious dogs. Am. J. Physiol. 1991, 260, 335–340. [Google Scholar] [CrossRef]
- van der Werf, C.; Nederend, I.; Hofman, N.; van Geloven, N.; Ebink, C.; Frohn-Mulder, I.M.; Aling, A.M.W.; Bosker, H.A.; Bracke, F.A.; can den Heuvel, F.; et al. Familial evaluation in catecholaminergic polymorphic ventricular tachycardia: Disease penetrance and expression in cardiac ryanodine receptor mutation-carrying relatives. Circ. Arrhythmia Electrophysiol. 2012, 5, 748–756. [Google Scholar] [CrossRef]
- Neco, P.; Torrente, A.G.; Mesirca, P.; Zorio, E.; Liu, N.; Priori, S.G.; Napolitano, C.; Richard, S.; Benitah, J.-P.; Mangoni, M.E.; et al. Paradoxical Effect of Increased Diastolic Ca2+ Release and Decreased Sinoatrial Node Activity in a Mouse Model of Catecholaminergic Polymorphic Ventricular Tachycardia. Circulation 2012, 126, 392–401. [Google Scholar] [CrossRef]
- Dobrzynski, H.; Li, J.; Tellez, J.; Greener, I.; Nikolski, V.; Wright, S.; Parson, S.; Jones, S.; Lancaster, M.; Yamamoto, M.; et al. Computer Three-Dimensional Reconstruction of the Sinoatrial Node. Circulation 2005, 111, 846–854. [Google Scholar] [CrossRef]
- Lange, G. Action of Driving Stimuli from Intrinsic and Extrinsic Sources on in Situ Cardiac Pacemaker Tissues. Circ. Res. 1965, 17, 449–459. [Google Scholar] [CrossRef]
- Hoffman, B.; Cranefield, P.F. Electrophysiology of the Heart; McGraw-Hill: New York, NY, USA, 1960. [Google Scholar]
- Boineau, J.P.; Canavan, T.E.; Schuessler, R.B.; Cain, M.E.; Corr, P.B.; Cox, J.L. Demonstration of a widely distributed atrial pacemaker complex in the human heart. Circulation 1988, 77, 1221–1237. [Google Scholar] [CrossRef]
- Boineau, J.P.; Schuessler, R.B.; Mooney, C.R.; Wylds, A.C.; Miller, C.B.; Hudson, R.D.; Borremans, J.M.; Brockus, C.W. Multicentric origin of the atrial depolarization wave: The pacemaker complex. Relation to dynamics of atrial conduction, P-wave changes and heart rate control. Circulation 1978, 58, 1036–1048. [Google Scholar] [CrossRef]
- Sharifov, O.F.; Fedorov, V.V.; Beloshapko, G.G.; Glukhov, A.V.; Yushmanova, A.V.; Rosenshtraukh, L.V. Roles of adrenergic and cholinergic stimulation in spontaneous atrial fibrillation in dogs. J. Am. Coll. Cardiol. 2004, 43, 483–490. [Google Scholar] [CrossRef]
- Fedorov, V.V.; Trifonova, O.P.; Glukhov, A.V.; Rosen, M.R.; Rosenshtraukh, L.V. The role of mechano-electrical feedback in the cholinergic atrial fibrillation initiation. In Mechanosensitivity in Cells and Tissues; Academia Publishing House Ltd.: New York, NY, USA, 2005. [Google Scholar]
- Gaskell, W.H. On the Innervation of the Heart, with especial reference to the Heart of the Tortoise. J. Physiol. 1883, 4, 43–230. [Google Scholar] [CrossRef]
- Soattin, L.; Borbas, Z.; Caldwell, J.; Prendergast, B.; Vohra, A.; Saeed, Y.; Hoschtitzky, A.; Yanni, J.; Atkinson, A.; Logantha, S.J.; et al. Structural and Functional Properties of Subsidiary Atrial Pacemakers in a Goat Model of Sinus Node Disease. Front. Physiol. 2021, 12, 592229. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; D’Souza, A.; Dobrzynski, H.; Lei, M.; Choudhury, M.; Billeter, R.; Kryukova, Y.; Robinson, R.B.; Kingston, P.A.; Boyett, M.R. Characterization of a right atrial subsidiary pacemaker and acceleration of the pacing rate by HCN over-expression. Cardiovasc. Res. 2013, 100, 160–169. [Google Scholar] [CrossRef]
- Gray, A.L.; Johnson, T.A.; Ardell, J.L.; Massari, V.J. Parasympathetic control of the heart. II. A novel interganglionic intrinsic cardiac circuit mediates neural control of heart rate. J. Appl. Physiol. 2004, 96, 2273–2278. [Google Scholar] [CrossRef]
- Choudhury, M.; Black, N.; Alghamdi, A.; D’Souza, A.; Wang, R.; Yanni, J.; Dobrzynski, H.; Kingston, P.A.; Zhang, H.; Boyett, M.R.; et al. TBX18 overexpression enhances pacemaker function in a rat subsidiary atrial pacemaker model of sick sinus syndrome. J. Physiol. 2018, 596, 6141–6155. [Google Scholar] [CrossRef] [PubMed]
- Rozanski, G.J.; Lipsius, S.L.; Randall, W.C.; Jones, S.B. Alterations in subsidiary pacemaker function after prolonged subsidiary pacemaker dominance in the canine right atrium. J. Am. Coll. Cardiol. 1984, 4, 535–542. [Google Scholar] [CrossRef]
- Kalman, J.M.; Olgin, J.E.; Karch, M.R.; Hamdan, M.; Lee, R.J.; Lesh, M.D. “Cristal tachycardias”: Origin of right atrial tachycardias from the crista terminalis identified by intracardiac echocardiography. J. Am. Coll. Cardiol. 1998, 31, 451–459. [Google Scholar] [CrossRef]
- Kistler, P.; Fynn, S.; Haqqani, H. Focal Atrial Tachycardia From the Ostium of the Coronary Sinus. Electrocardiographic and Electrophysiological Characterization and Radiofrequency Ablation. ACC Curr. J. Rev. 2005, 14, 55. [Google Scholar] [CrossRef]
- Marrouche, N.F.; SippensGroenewegen, A.; Yang, Y.; Dibs, S.; Scheinman, M.M. Clinical and electrophysiologic characteristics of left septal atrial tachycardia. J. Am. Coll. Cardiol. 2002, 40, 1133–1139. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, D.; Glukhov, A.V. Cellular and Molecular Mechanisms of Functional Hierarchy of Pacemaker Clusters in the Sinoatrial Node: New Insights into Sick Sinus Syndrome. J. Cardiovasc. Dev. Dis. 2021, 8, 43. https://doi.org/10.3390/jcdd8040043
Lang D, Glukhov AV. Cellular and Molecular Mechanisms of Functional Hierarchy of Pacemaker Clusters in the Sinoatrial Node: New Insights into Sick Sinus Syndrome. Journal of Cardiovascular Development and Disease. 2021; 8(4):43. https://doi.org/10.3390/jcdd8040043
Chicago/Turabian StyleLang, Di, and Alexey V. Glukhov. 2021. "Cellular and Molecular Mechanisms of Functional Hierarchy of Pacemaker Clusters in the Sinoatrial Node: New Insights into Sick Sinus Syndrome" Journal of Cardiovascular Development and Disease 8, no. 4: 43. https://doi.org/10.3390/jcdd8040043
APA StyleLang, D., & Glukhov, A. V. (2021). Cellular and Molecular Mechanisms of Functional Hierarchy of Pacemaker Clusters in the Sinoatrial Node: New Insights into Sick Sinus Syndrome. Journal of Cardiovascular Development and Disease, 8(4), 43. https://doi.org/10.3390/jcdd8040043