
The “Dead-End Tract” and Its Role in Arrhythmogenesis

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Dead-End Tract in the Developing Cardiac Conduction System
2.1. The Developing Heart
2.2. The Developing Conduction System
2.2.1. The Ring Theory
2.2.2. Nodal Myocytes
2.2.3. The Ventricular Conduction System
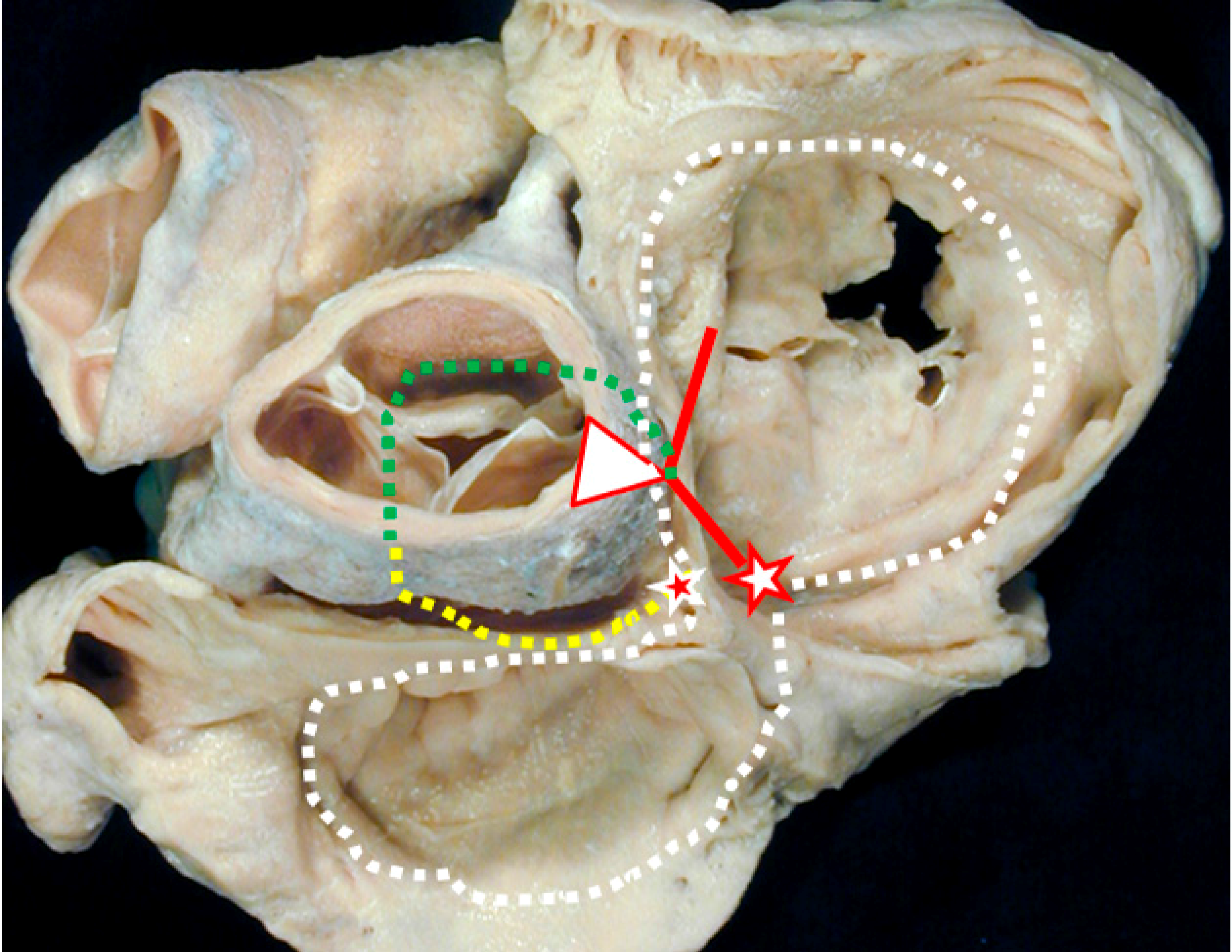
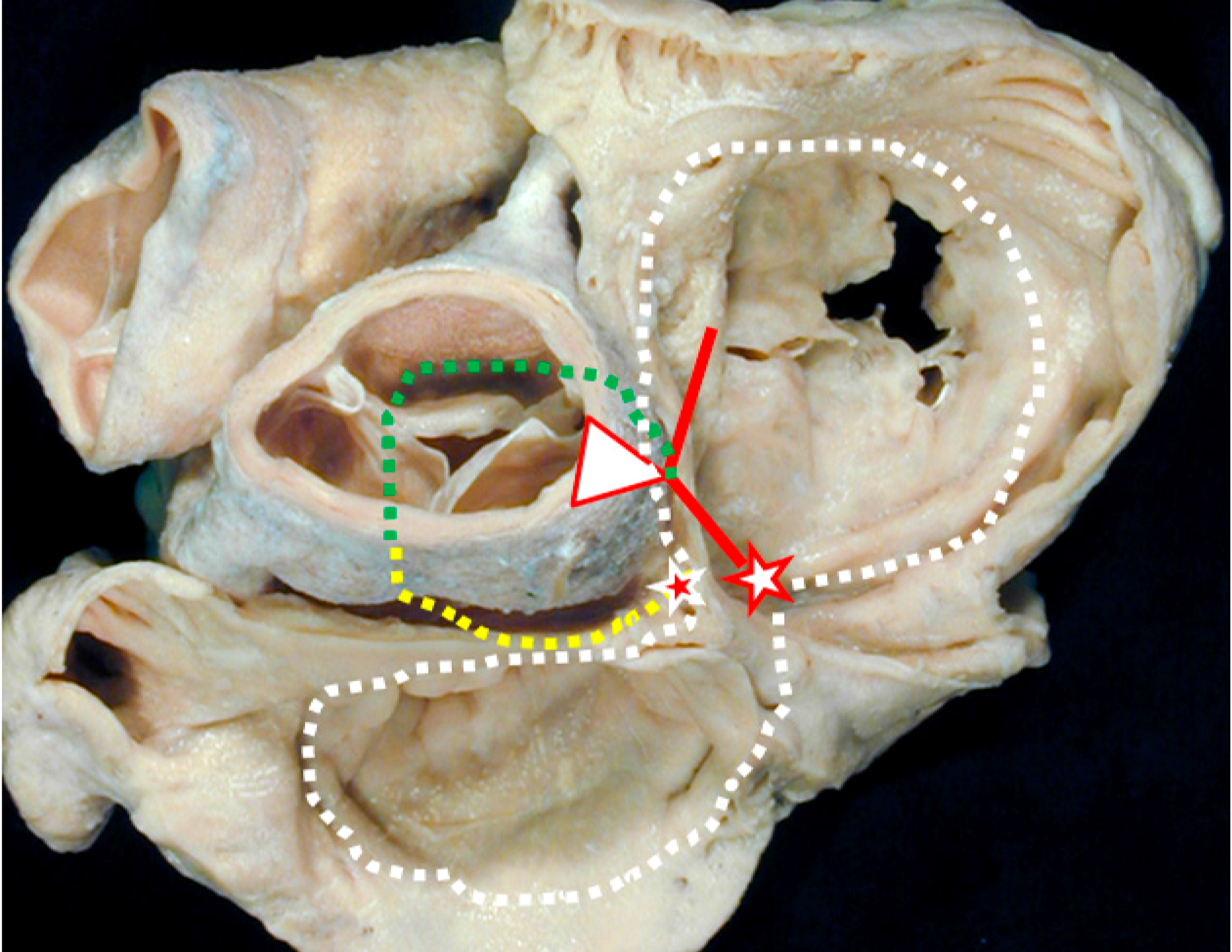
2.3. Experimental Pathologic Evidence of the Dead-End Tract during Development
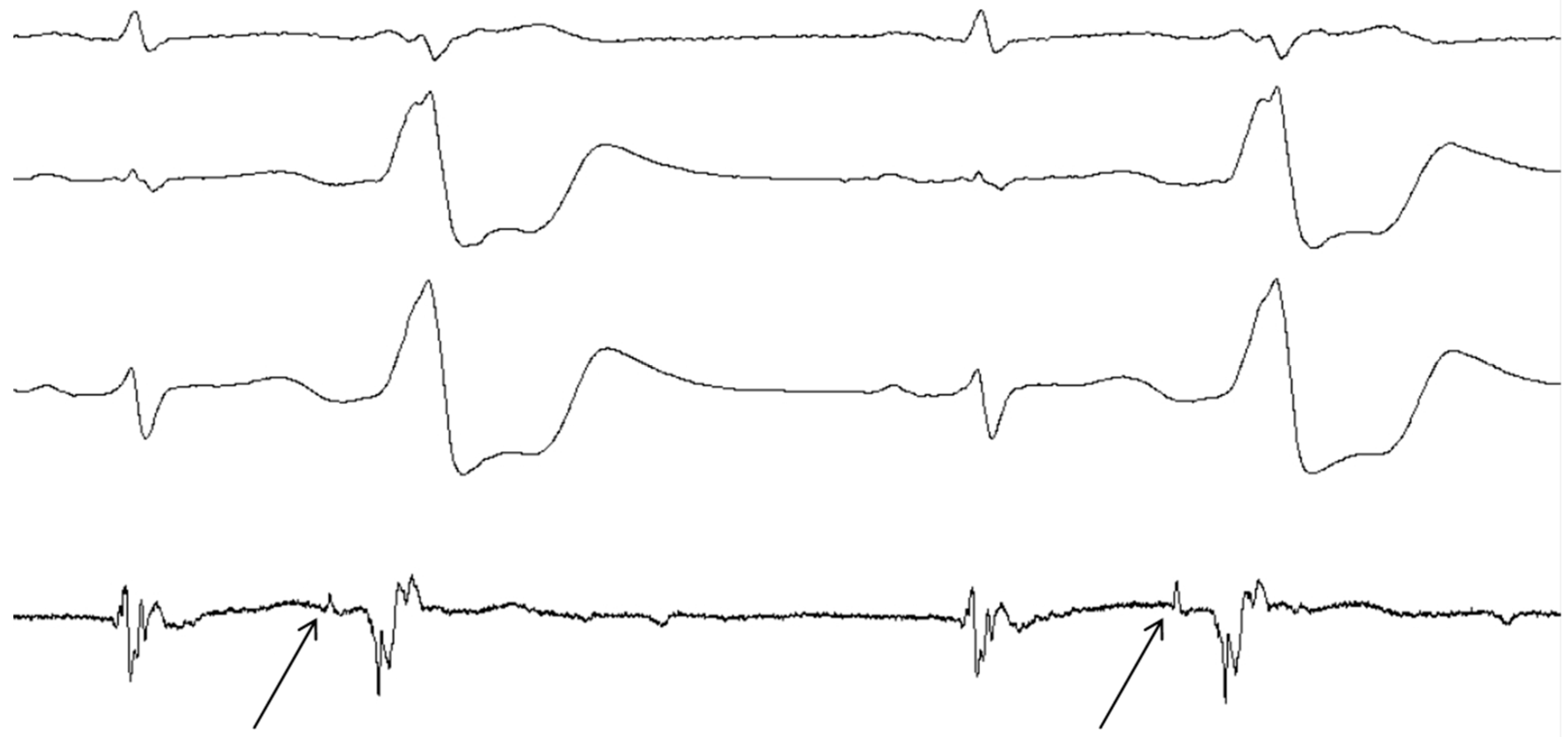
3. Clinical Evidence of the Dead-End Tract
4. Discussion
4.1. Evidence
4.2. Clinical Implications
4.3. Considerations and Limitations
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kurosawa, H.; Becker, A.E. Dead-end tract of the conduction axis. Int. J. Cardiol. 1985, 7, 13–20. [Google Scholar] [CrossRef]
- Wessels, A.; Vermeulen, J.L.; Verbeek, F.J.; Viragh, S.; Kalman, F.; Lamers, W.H.; Moorman, A.F. Spatial distribution of “tissue-specific“ antigens in the developing human heart and skeletal muscle. III. An immunohistochemical analysis of the distribution of the neural tissue antigen g1n2 in the embryonic heart; implications for the development of the atrioventricular conduction system. Anat. Rec. 1992, 232, 97–111. [Google Scholar] [PubMed]
- Jongbloed, M.R.; Mahtab, E.A.; Blom, N.A.; Schalij, M.J.; Gittenberger-de Groot, A.C. Development of the cardiac conduction system and the possible relation to predilection sites of arrhythmogenesis. Sci. World J. 2008, 8, 239–269. [Google Scholar] [CrossRef] [PubMed]
- Akca, F.; Szili-Torok, T. Challenging the dogma: Disappearance of right ventricular outflow tract premature ventricular contractions after ablation of a right free wall accessory pathway—What is the common link? Europace 2014, 16 (Suppl. 2), ii160. [Google Scholar] [CrossRef]
- Wylie, J.V., Jr.; Milliez, P.; Germano, J.J.; Richardson, A.; Ngwu, O.; Zimetbaum, P.J.; Papageorgiou, P.; Josephson, M.E. Atrioventricular nodal reentrant tachycardia associated with idiopathic ventricular tachycardia: Clinical and electrophysiologic characteristics. J. Electrocardiol. 2007, 40, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Kautzner, J.; Cihák, R.; Vancura, V.; Bytesník, J. Coincidence of idiopathic ventricular outflow tract tachycardia and atrioventricular nodal reentrant tachycardia. Europace 2003, 5, 215–220. [Google Scholar] [CrossRef]
- Hasdemir, C.; Alp, A.; Simsek, E.; Kose, N.; Aydin, M.; Payzin, S. Spontaneous atrioventricular nodal reentrant tachycardia in patients with idiopathic ventricular arrhythmias: The incidence, clinical, and electrophysiologic characteristics. J. Cardiovasc. Electrophysiol. 2013, 24, 1370–1374. [Google Scholar] [CrossRef] [PubMed]
- Topilski, I.; Glick, A.; Viskin, S.; Belhassen, B. Frequency of spontaneous and inducible atrioventricular nodal reentry tachycardia in patients with idiopathic outflow tract ventricular arrhythmias. Pacing Clin. Electrophysiol. 2006, 29, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sobotka, P.A.; Mayer, J.H.; Bauernfeind, R.A.; Kanakis, C., Jr.; Rosen, K.M. Arrhythmias documented by 24-h continuous ambulatory electrocardiographic monitoring in young women without apparent heart disease. Am. Heart J. 1981, 101, 753–759. [Google Scholar] [CrossRef]
- Brodsky, M.; Wu, D.; Denes, P.; Kanakis, C.; Rosen, K.M. Arrhythmias documented by 24 h continuous electrocardiographic monitoring in 50 male medical students without apparent heart disease. Am. J. Cardiol. 1977, 39, 390–395. [Google Scholar] [CrossRef]
- Brooks, R.; Burgess, J.H. Idiopathic ventricular tachycardia. A review. Medicine 1988, 67, 271–294. [Google Scholar] [CrossRef] [PubMed]
- Zweytick, B.; Pignoni-Mori, P.; Zweytick, G.; Steinbach, K. Prognostic significance of right ventricular extrasystoles. Europace 2004, 6, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Pytkowski, M.; Maciag, A.; Jankowska, A.; Kowalik, I.; Kraska, A.; Farkowski, M.M.; Golicki, D.; Szwed, H. Quality of life improvement after radiofrequency catheter ablation of outflow tract ventricular arrhythmias in patients with structurally normal heart. Acta Cardiol. 2012, 67, 153–159. [Google Scholar] [PubMed]
- Chugh, S.S.; Shen, W.; Luria, D.M.; Smith, H.C. First evidence of premature ventricular complex-induced cardiomyopathy: A potentially reversible cause of heart failure. J. Cardiovasc. Electrophysiol. 2000, 11, 328–329. [Google Scholar] [CrossRef] [PubMed]
- Kanei, Y.; Friedman, M.; Ogawa, N.; Hanon, S.; Lam, P.; Schweitzer, P. Frequent premature ventricular complexes originating from the right ventricular outflow tract are associated with left ventricular dysfunction. Ann. Noninvasive Electrocardiol. 2008, 13, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Yong, Y.; Cao, K.; Chen, S.; Xu, D. Evaluation of global and regional left ventricular systolic function in patients with frequent isolated premature ventricular complexes from the right ventricular outflow tract. Clin. Med. J. 2012, 125, 214–220. [Google Scholar]
- Plageman, T.F., Jr.; Yutzey, K.E. T-box genes and heart development: Putting the “t“ in heart. Dev. Dyn. 2005, 232, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Boogerd, C.J.; Moorman, A.F.; Barnett, P. Protein interactions at the heart of cardiac chamber formation. Ann. Anat. 2009, 191, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.G.; Brown, N.A.; Buckingham, M.E. The arterial pole of the mouse heart forms from fgf10-expressing cells in pharyngeal mesoderm. Dev. Cell 2001, 1, 435–440. [Google Scholar] [CrossRef]
- Mjaatvedt, C.H.; Nakaoka, T.; Moreno-Rodriguez, R.; Norris, R.A.; Kern, M.J.; Eisenberg, C.A.; Turner, D.; Markwald, R.R. The outflow tract of the heart is recruited from a novel heart-forming field. Dev. Biol. 2001, 238, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Waldo, K.L.; Kumiski, D.H.; Wallis, K.T.; Stadt, H.A.; Hutson, M.R.; Platt, D.H.; Kirby, M.L. Conotruncal myocardium arises from a secondary heart field. Development 2001, 128, 3179–3188. [Google Scholar] [PubMed]
- Cai, C.L.; Liang, X.; Shi, Y.; Chu, P.H.; Pfaff, S.L.; Chen, J.; Evans, S. Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev. Cell 2003, 5, 877–889. [Google Scholar] [CrossRef]
- Christoffels, V.M.; Habets, P.E.; Franco, D.; Campione, M.; de Jong, F.; Lamers, W.H.; Bao, Z.Z.; Palmer, S.; Biben, C.; Harvey, R.P.; et al. Chamber formation and morphogenesis in the developing mammalian heart. Dev. Biol. 2000, 223, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Campione, M.; Kelly, R.; Zammit, P.S.; Buckingham, M.; Lamers, W.H.; Moorman, A.F. Multiple transcriptional domains, with distinct left and right components, in the atrial chambers of the developing heart. Circ. Res. 2000, 87, 984–991. [Google Scholar] [CrossRef] [PubMed]
- DeRuiter, M.C.; Gittenberger-De Groot, A.C.; Wenink, A.C.; Poelmann, R.E.; Mentink, M.M. In normal development pulmonary veins are connected to the sinus venosus segment in the left atrium. Anat. Rec. 1995, 243, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger-de Groot, A.C.; Bartelings, M.M.; Deruiter, M.C.; Poelmann, R.E. Basics of cardiac development for the understanding of congenital heart malformations. Pediatr. Res. 2005, 57, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Moorman, A.F.; Christoffels, V.M. Cardiac chamber formation: Development, genes, and evolution. Physiol. Rev. 2003, 83, 1223–1267. [Google Scholar] [CrossRef] [PubMed]
- De Boer, B.A.; van den Berg, G.; de Boer, P.A.; Moorman, A.F.; Ruijter, J.M. Growth of the developing mouse heart: An interactive qualitative and quantitative 3D atlas. Dev. Biol. 2012, 368, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Lavine, K.J.; Yu, K.; White, A.C.; Zhang, X.; Smith, C.; Partanen, J.; Ornitz, D.M. Endocardial and epicardial derived FGF signals regulate myocardial proliferation and differentiation in vivo. Dev. Cell 2005, 8, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Ieda, M.; Tsuchihashi, T.; Ivey, K.N.; Ross, R.S.; Hong, T.T.; Shaw, R.M.; Srivastava, D. Cardiac fibroblasts regulate myocardial proliferation through beta1 integrin signaling. Dev. Cell 2009, 16, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Wenink, A.C. Development of the human cardiac conducting system. J. Anat. 1976, 121, 617–631. [Google Scholar] [PubMed]
- Lamers, W.H.; Wessels, A.; Verbeek, F.J.; Moorman, A.F.; Viragh, S.; Wenink, A.C.; Gittenberger-de Groot, A.C.; Anderson, R.H. New findings concerning ventricular septation in the human heart. Implications for maldevelopment. Circulation 1992, 86, 1194–1205. [Google Scholar] [CrossRef] [PubMed]
- Kamino, K. Optical approaches to ontogeny of electrical activity and related functional organization during early heart development. Physiol. Rev. 1991, 71, 53–91. [Google Scholar] [PubMed]
- Lamers, W.H.; De Jong, F.; De Groot, I.J.; Moorman, A.F. The development of the avian conduction system, a review. Eur. J. Morphol. 1991, 29, 233–253. [Google Scholar] [PubMed]
- Bakker, M.L.; Boukens, B.J.; Mommersteeg, M.T.; Brons, J.F.; Wakker, V.; Moorman, A.F.; Christoffels, V.M. Transcription factor tbx3 is required for the specification of the atrioventricular conduction system. Circ. Res. 2008, 102, 1340–1349. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Taylor, I.M. Development of atrioventricular specialized tissue in human heart. Brit. Heart J. 1972, 34, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Moorman, A.F.; de Jong, F.; Denyn, M.M.; Lamers, W.H. Development of the cardiac conduction system. Circ. Res. 1998, 82, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Van Mierop, L.H. Location of pacemaker in chick embryo heart at the time of initiation of heartbeat. Am. J. Physiol. 1967, 212, 407–415. [Google Scholar] [PubMed]
- Paff, G.H.; Boucek, R.J.; Harrell, T.C. Observations on the development of the electrocardiogram. Anat. Rec. 1968, 160, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Arraez-Aybar, L.A.; Turrero-Nogues, A.; Marantos-Gamarra, D.G. Embryonic cardiac morphometry in carnegie stages 15–23, from the complutense university of madrid institute of embryology human embryo collection. Cells Tissues Organs 2008, 187, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Oostra, R.J.; Steding, G.; Lamers, W.H.; Moorman, A.F.M. Steding's and Viragh's Scanning Electron Microscopy Atlas of the Developing Human Heart; Springer Science & Business Media: New York, NY, USA, 2007. [Google Scholar]
- O’Rahilly, R.; Müller, F. Developmental Stages in Human Embryos; Carnegie Institution of Washington: Washington, DC, USA, 1987. [Google Scholar]
- Viragh, S.; Challice, C.E. The development of the conduction system in the mouse embryo heart. Dev. Biol. 1980, 80, 28–45. [Google Scholar] [CrossRef]
- De Groot, I.J.M.; Wessels, A.; Virágh, S.; Lamers, W.H.; Moorman, A.F. The relation between isomyosin heavy chain expression pattern and the architecture of sinoatrial nodes in chicken, rat and human embryos. In Sarcomeric and Non-Sarcomeric Muscles: Basic and Applied Research Prospects for the 90s; Carraro, U., Ed.; Unipress: Padova, Italy, 1988; pp. 305–310. [Google Scholar]
- Hoff, E.C.; Kramer, T.C.; DuBois, D.; Patten, B.M. The development of the electrocardiogram of the embryonic heart. Am. Heart J. 1939, 17, 470–488. [Google Scholar] [CrossRef]
- Sylva, M.; van den Hoff, M.J.; Moorman, A.F. Development of the human heart. Am. J. Med. Genet. A 2014, 164a, 1347–1371. [Google Scholar] [CrossRef] [PubMed]
- De Jong, F.; Opthof, T.; Wilde, A.A.; Janse, M.J.; Charles, R.; Lamers, W.H.; Moorman, A.F. Persisting zones of slow impulse conduction in developing chicken hearts. Circ. Res. 1992, 71, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Arguello, C.; Alanis, J.; Pantoja, O.; Valenzuela, B. Electrophysiological and ultrastructural study of the atrioventricular canal during the development of the chick embryo. J. Mol. Cell. Cardiol. 1986, 18, 499–510. [Google Scholar] [CrossRef]
- Lieberman, M.; Paes de Carvalho, A. The electrophysiological organization of the embryonic chick heart. J. Gen. Physiol. 1965, 49, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, M.; de Paes Carvalho, A. The spread of excitation in the embryonic chick heart. J. Gen. Physiol. 1965, 49, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Paff, G.H.; Boucek, R.J.; Klopfenstein, H.S. Experimental heart-block in the chick embryo. Anat. Rec. 1964, 149, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Patten, B.M. The development of the sinoventricular conduction system. Med. Bull. 1956, 22, 1–21. [Google Scholar]
- Vassall-Adams, P.R. The development of the atrioventricular bundle and its branches in the avian heart. J. Anat. 1982, 134, 169–183. [Google Scholar] [PubMed]
- Sanders, E.; de Groot, I.J.; Geerts, W.J.; de Jong, F.; van Horssen, A.A.; Los, J.A.; Moorman, A.F. The local expression of adult chicken heart myosins during development. Ii. Ventricular conducting tissue. Anat. Embryol. 1986, 174, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Viragh, S.; Porte, A. The fine structure of the conducting system of the monkey heart (macaca mulatta). I. The sino-atrial node and the internodal connections. Z Zellforsch. Mikrosk. Anat. 1973, 145, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Viragh, S.; Challice, C.E. The development of the conduction system in the mouse embryo heart. II. Histogenesis of the atrioventricular node and bundle. Dev. Biol. 1977, 56, 397–411. [Google Scholar] [CrossRef]
- De Mello, W.C. Intercellular communication in cardiac muscle. Circ. Res. 1982, 51, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Van Kempen, M.J.; Fromaget, C.; Gros, D.; Moorman, A.F.; Lamers, W.H. Spatial distribution of connexin43, the major cardiac gap junction protein, in the developing and adult rat heart. Circ. Res. 1991, 68, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Gros, D.; Mocquard, J.P.; Challice, C.E.; Schrevel, J. Formation and growth of gap junctions in mouse myocardium during ontogenesis: A freeze-cleave study. J. Cell Sci. 1978, 30, 45–61. [Google Scholar] [PubMed]
- Fromaget, C.; el Aoumari, A.; Gros, D. Distribution pattern of connexin 43, a gap junctional protein, during the differentiation of mouse heart myocytes. Differentiation 1992, 51, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Arguello, C.; Alanis, J.; Valenzuela, B. The early development of the atrioventricular node and bundle of his in the embryonic chick heart. An electrophysiological and morphological study. Development 1988, 102, 623–637. [Google Scholar] [PubMed]
- Katz, A.M. Physiology of the Heart; Raven Press: New York, NY, USA, 1977. [Google Scholar]
- Oosthoek, P.W.; Viragh, S.; Mayen, A.E.; van Kempen, M.J.; Lamers, W.H.; Moorman, A.F. Immunohistochemical delineation of the conduction system. I: The sinoatrial node. Circ. Res. 1993, 73, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Oosthoek, P.W.; Viragh, S.; Lamers, W.H.; Moorman, A.F. Immunohistochemical delineation of the conduction system. Ii: The atrioventricular node and purkinje fibers. Circ. Res. 1993, 73, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Birchmeier, C. Multiple essential functions of neuregulin in development. Nature 1995, 378, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.F.; Simon, H.; Chen, H.; Bates, B.; Hung, M.C.; Hauser, C. Requirement for neuregulin receptor erbb2 in neural and cardiac development. Nature 1995, 378, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, M.; Casagranda, F.; Orioli, D.; Simon, H.; Lai, C.; Klein, R.; Lemke, G. Aberrant neural and cardiac development in mice lacking the erbb4 neuregulin receptor. Nature 1995, 378, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Marchionni, M.A. Cell-cell signalling. Neu tack on neuregulin. Nature 1995, 378, 334–335. [Google Scholar] [CrossRef] [PubMed]
- Dyson, E.; Sucov, H.M.; Kubalak, S.W.; Schmid-Schonbein, G.W.; DeLano, F.A.; Evans, R.M.; Ross, J., Jr.; Chien, K.R. Atrial-like phenotype is associated with embryonic ventricular failure in retinoid X receptor alpha -/- mice. Proc. Natl. Acad. Sci. USA 1995, 92, 7386–7390. [Google Scholar] [CrossRef] [PubMed]
- Kastner, P.; Grondona, J.M.; Mark, M.; Gansmuller, A.; LeMeur, M.; Decimo, D.; Vonesch, J.L.; Dolle, P.; Chambon, P. Genetic analysis of rxr alpha developmental function: Convergence of rxr and rar signaling pathways in heart and eye morphogenesis. Cell 1994, 78, 987–1003. [Google Scholar] [CrossRef]
- Sucov, H.M.; Dyson, E.; Gumeringer, C.L.; Price, J.; Chien, K.R.; Evans, R.M. RXR alpha mutant mice establish a genetic basis for vitamin a signaling in heart morphogenesis. Genes Dev. 1994, 8, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Robb, J.S.; Petri, R. Expansions of the atrio-ventricular system in the atria. In The Specialized Tissue of the Heart; Elsevier: Amsterdam, the Netherlands, 1961; pp. 1–18. [Google Scholar]
- Thompson, R.P.; Lindroth, J.R.; Wong, Y.M.M. Regional differences in DNA-synthetic activity in the preseptation myocardium of the chick. In Developmental Cardioliology: Morphogenesis and Function; Futura Publishing Co: Mount Kisco, NY, USA, 1990; pp. 219–234. [Google Scholar]
- Gourdie, R.G.; Mima, T.; Thompson, R.P.; Mikawa, T. Terminal diversification of the myocyte lineage generates purkinje fibers of the cardiac conduction system. Development 1995, 121, 1423–1431. [Google Scholar] [PubMed]
- Mikawa, T.; Borisov, A.; Brown, A.M.; Fischman, D.A. Clonal analysis of cardiac morphogenesis in the chicken embryo using a replication-defective retrovirus: I. Formation of the ventricular myocardium. Dev. Dyn. 1992, 193, 11–23. [Google Scholar] [CrossRef] [PubMed]
- De Haan, R.L. Differentiation of the atrioventricular conducting system of the heart. Circulation 1961, 24, 458–470. [Google Scholar]
- Chuck, E.T.; Freeman, D.M.; Watanabe, M.; Rosenbaum, D.S. Changing activation sequence in the embryonic chick heart. Implications for the development of the his-purkinje system. Circ. Res. 1997, 81, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Thompson, R.P.; Terracio, L.; Borg, T.K. Developmental anatomy of hnk-1 immunoreactivity in the embryonic rat heart: Co-distribution with early conduction tissue. Anat. Embryol. 1993, 187, 445–460. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Jing, Y.; Wagenaar, G.T.M.; Lamers, W.H.; Moorman, A.F. The trabecular component of the embryonic ventricle. In The Developing Heart; Ost'ádal, B., Nagano, M., Takeda, N., Dhalla, N.S., Eds.; Lippincott Raven: New York, NY, USA, 1996; pp. 51–60. [Google Scholar]
- Robb, J.S. Comparative Basic Cardiology; Grune & Stratton: New York, NY, USA, 1965. [Google Scholar]
- Wessels, A.; Markman, M.W.; Vermeulen, J.L.; Anderson, R.H.; Moorman, A.F.; Lamers, W.H. The development of the atrioventricular junction in the human heart. Circ. Res. 1996, 78, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Timm, M.; Fallah-Najmabadi, H. Cardiac expression of polysialylated ncam in the chicken embryo: Correlation with the ventricular conduction system. Dev. Dyn. 1992, 194, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Viragh, S.; Challice, C.E. The impulse generation and conduction system of the heart. Ultrastruct. Mamm. Heart 1973, 6, 43–89. [Google Scholar]
- Lu, Y.; James, T.N.; Yamamoto, S.; Terasaki, F. Cardiac conduction system in the chicken: Gross anatomy plus light and electron microscopy. Anat. Rec. 1993, 236, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Davies, M.J.; Becker, A.E. Atrioventricular ring specialized tissue in the normal heart. Eur. J. Cardiol. 1974, 2, 219–230. [Google Scholar]
- Kurosawa, H.; Becker, A.E. Modification of the precise relationship of the atrioventricular conduction bundle to the margins of the ventricular septal defects by the trabecula septomarginalis. J. Thorac. Cardiovasc. Surg. 1984, 87, 605–615. [Google Scholar] [PubMed]
- Anderson, R.H. The disposition, morphology and innervation of cardiac specialized tissue in the guinea-pig. J. Anat. 1972, 111, 453–468. [Google Scholar] [PubMed]
- McGuire, M.A.; de Bakker, J.M.; Vermeulen, J.T.; Opthof, T.; Becker, A.E.; Janse, M.J. Origin and significance of double potentials near the atrioventricular node. Correlation of extracellular potentials, intracellular potentials, and histology. Circulation 1994, 89, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.A.; de Bakker, J.M.; Vermeulen, J.T.; Moorman, A.F.; Loh, P.; Thibault, B.; Vermeulen, J.L.; Becker, A.E.; Janse, M.J. Atrioventricular junctional tissue. Discrepancy between histological and electrophysiological characteristics. Circulation 1996, 94, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Anselme, F. Association of idiopathic rvot vt and avnrt: Anything else than chance? Europace 2003, 5, 221–223. [Google Scholar] [CrossRef]
- Anderson, R.H.; Boyett, M.R.; Dobrzynski, H.; Moorman, A.F. The anatomy of the conduction system: Implications for the clinical cardiologist. J. Cardiovasc. Transl. Res. 2013, 6, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Yanni, J.; Boyett, M.R.; Anderson, R.H.; Dobrzynski, H. The extent of the specialized atrioventricular ring tissue. Heart Rhythm. 2009, 6, 672–680. [Google Scholar]
- Hai, J.J.; Chahal, A.A.; Friedman, P.A.; Vaidya, V.R.; Syed, F.F.; DeSimone, C.V.; Nanda, S.; Brady, P.A.; Madhavan, M.; Cha, Y.M.; et al. Electrophysiologic characteristics of ventricular arrhythmias arising from the aortic mitral continuity-potential role of the conduction system. J. Cardiovasc. Electrophysiol. 2015, 26, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, F.; Fotuhi, P.; Ho, S.Y.; Hebe, J.; Volkmer, M.; Goya, M.; Burns, M.; Antz, M.; Ernst, S.; Cappato, R.; et al. Repetitive monomorphic ventricular tachycardia originating from the aortic sinus cusp: Electrocardiographic characterization for guiding catheter ablation. J. Am. Coll. Cardiol. 2002, 39, 500–508. [Google Scholar] [CrossRef]
- Tada, H.; Naito, S.; Ito, S.; Kurosaki, K.; Ueda, M.; Shinbo, G.; Hoshizaki, H.; Oshima, S.; Taniguchi, K.; Nogami, A. Significance of two potentials for predicting successful catheter ablation from the left sinus of valsalva for left ventricular epicardial tachycardia. Pacing Clin. Electrophysiol. 2004, 27, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, K.; Yamauchi, Y.; Takahashi, A.; Yokoyama, Y.; Sekiguchi, Y.; Watanabe, J.; Iesaka, Y.; Shirato, K.; Aonuma, K. Idiopathic left ventricular tachycardia originating from the mitral annulus. J. Cardiovasc. Electrophysiol. 2005, 16, 1029–1036. [Google Scholar] [CrossRef] [PubMed]
- Tada, H.; Ito, S.; Naito, S.; Kurosaki, K.; Kubota, S.; Sugiyasu, A.; Tsuchiya, T.; Miyaji, K.; Yamada, M.; Kutsumi, Y.; et al. Idiopathic ventricular arrhythmia arising from the mitral annulus: A distinct subgroup of idiopathic ventricular arrhythmias. J. Am. Coll. Cardiol. 2005, 45, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Murakami, Y.; Yoshida, N.; Okada, T.; Shimizu, T.; Toyama, J.; Yoshida, Y.; Tsuboi, N.; Muto, M.; Inden, Y.; et al. Preferential conduction across the ventricular outflow septum in ventricular arrhythmias originating from the aortic sinus cusp. J. Am. Coll. Cardiol. 2007, 50, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Platonov, M.; McElderry, H.T.; Kay, G.N. Left ventricular outflow tract tachycardia with preferential conduction and multiple exits. Circ. Arrhythm. Electrophysiol. 2008, 1, 140–142. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, K.; Fukuda, K.; Wakayama, Y.; Sugai, Y.; Hirose, M.; Yamaguchi, N.; Takase, K.; Yamauchi, Y.; Takahashi, A.; Aonuma, K.; et al. Electrocardiographic characteristics of the variants of idiopathic left ventricular outflow tract ventricular tachyarrhythmias. J. Cardiovasc. Electrophysiol. 2008, 19, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; McElderry, H.T.; Doppalapudi, H.; Murakami, Y.; Yoshida, Y.; Yoshida, N.; Okada, T.; Tsuboi, N.; Inden, Y.; Murohara, T.; et al. Idiopathic ventricular arrhythmias originating from the aortic root prevalence, electrocardiographic and electrophysiologic characteristics, and results of radiofrequency catheter ablation. J. Am. Coll. Cardiol. 2008, 52, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Litovsky, S.H.; Kay, G.N. The left ventricular ostium: An anatomic concept relevant to idiopathic ventricular arrhythmias. Circ. Arrhythm. Electrophysiol. 2008, 1, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; McElderry, H.T.; Doppalapudi, H.; Okada, T.; Murakami, Y.; Yoshida, Y.; Yoshida, N.; Inden, Y.; Murohara, T.; Plumb, V.J.; et al. Idiopathic ventricular arrhythmias originating from the left ventricular summit: Anatomic concepts relevant to ablation. Circ. Arrhythm. Electrophysiol. 2010, 3, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Yoshida, Y.; Inden, Y.; Murohara, T.; Kay, G.N. Sequential ventricular prepotentials recorded within the left coronary cusp of the aorta during idiopathic pvcs: What is the mechanism? Pacing Clin. Electrophysiol. 2011, 34, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Furushima, H.; Chinushi, M.; Iijima, K.; Izumi, D.; Hosaka, Y.; Aizawa, Y. Relationship between electroanatomical voltage mapping characteristics and breakout site of ventricular activation in idiopathic ventricular tachyarrhythmia originating from the right ventricular outflow tract septum. J. Interv. Card. Electrophysiol. 2012, 33, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, H.; Yamauchi, Y.; Iesaka, Y.; Yagishita, A.; Sasaki, T.; Higuchi, K.; Kawabata, M.; Sugiyama, K.; Tanaka, Y.; Kusa, S.; et al. Discrete prepotential as an indicator of successful ablation in patients with coronary cusp ventricular arrhythmia. Circ. Arrhythm. Electrophysiol. 2013, 6, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Shirai, Y.; Goya, M.; Isobe, M.; Hirao, K. Preferential pathway pacing within the aortic sinus of valsalva: Strong evidence for the existence of preferential conduction with different exit sites traversing the ventricular septum. J. Cardiovasc. Electrophysiol. 2015, 26, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Cuello, C.; Huang, S.K.; Wagshal, A.B.; Pires, L.A.; Mittleman, R.S.; Bonavita, G.J. Radiofrequency catheter ablation of the atrioventricular junction by a supravalvular noncoronary aortic cusp approach. Pacing Clin. Electrophysiol. 1994, 17, 1182–1185. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H. Clinical anatomy of the aortic root. Heart 2000, 84, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.M.; Smeets, J.L.; Timmermans, C.; Wellens, H.J. Predictors for successful ablation of right- and left-sided idiopathic ventricular tachycardia. Am. J. Cardiol. 1997, 79, 309–314. [Google Scholar] [CrossRef]
- Kennedy, H.L.; Whitlock, J.A.; Sprague, M.K.; Kennedy, L.J.; Buckingham, T.A.; Goldberg, R.J. Long-term follow-up of asymptomatic healthy subjects with frequent and complex ventricular ectopy. N. Engl. J. Med. 1985, 312, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Gaita, F.; Giustetto, C.; Di Donna, P.; Richiardi, E.; Libero, L.; Brusin, M.C.; Molinari, G.; Trevi, G. Long-term follow-up of right ventricular monomorphic extrasystoles. J. Am. Coll. Cardiol. 2001, 38, 364–370. [Google Scholar] [CrossRef]
- Bogun, F.; Crawford, T.; Reich, S.; Koelling, T.M.; Armstrong, W.; Good, E.; Jongnarangsin, K.; Marine, J.E.; Chugh, A.; Pelosi, F.; et al. Radiofrequency ablation of frequent, idiopathic premature ventricular complexes: Comparison with a control group without intervention. Heart Rhythm. 2007, 4, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Yarlagadda, R.K.; Iwai, S.; Stein, K.M.; Markowitz, S.M.; Shah, B.K.; Cheung, J.W.; Tan, V.; Lerman, B.B.; Mittal, S. Reversal of cardiomyopathy in patients with repetitive monomorphic ventricular ectopy originating from the right ventricular outflow tract. Circulation 2005, 112, 1092–1097. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Vries, L.; Hendriks, A.; Szili-Torok, T. The “Dead-End Tract” and Its Role in Arrhythmogenesis. J. Cardiovasc. Dev. Dis. 2016, 3, 11. https://doi.org/10.3390/jcdd3020011
De Vries L, Hendriks A, Szili-Torok T. The “Dead-End Tract” and Its Role in Arrhythmogenesis. Journal of Cardiovascular Development and Disease. 2016; 3(2):11. https://doi.org/10.3390/jcdd3020011
Chicago/Turabian StyleDe Vries, Lennart, Astrid Hendriks, and Tamas Szili-Torok. 2016. "The “Dead-End Tract” and Its Role in Arrhythmogenesis" Journal of Cardiovascular Development and Disease 3, no. 2: 11. https://doi.org/10.3390/jcdd3020011
APA StyleDe Vries, L., Hendriks, A., & Szili-Torok, T. (2016). The “Dead-End Tract” and Its Role in Arrhythmogenesis. Journal of Cardiovascular Development and Disease, 3(2), 11. https://doi.org/10.3390/jcdd3020011