
A PMAxxTM qPCR Assay Reveals That Dietary Administration of the Microalgae Tetraselmis chuii Does Not Affect Salmonella Infantis Caecal Content in Early-Treated Broiler Chickens

 , , ,
, , ,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microalgae Biomass
2.2. Bacterial Preparation
2.3. Determination of S. Infantis Load
2.3.1. Cultivation of S. Infantis and Extraction of Genomic DNA
2.3.2. Amplification of the invA Gene
2.3.3. Staining of Viable and Non-Viable Salmonella Cells Using PMAxxTM
2.4. Experimental Setup
2.5. Statistical Analyses
3. Results
3.1. Amplification of the invA Gene
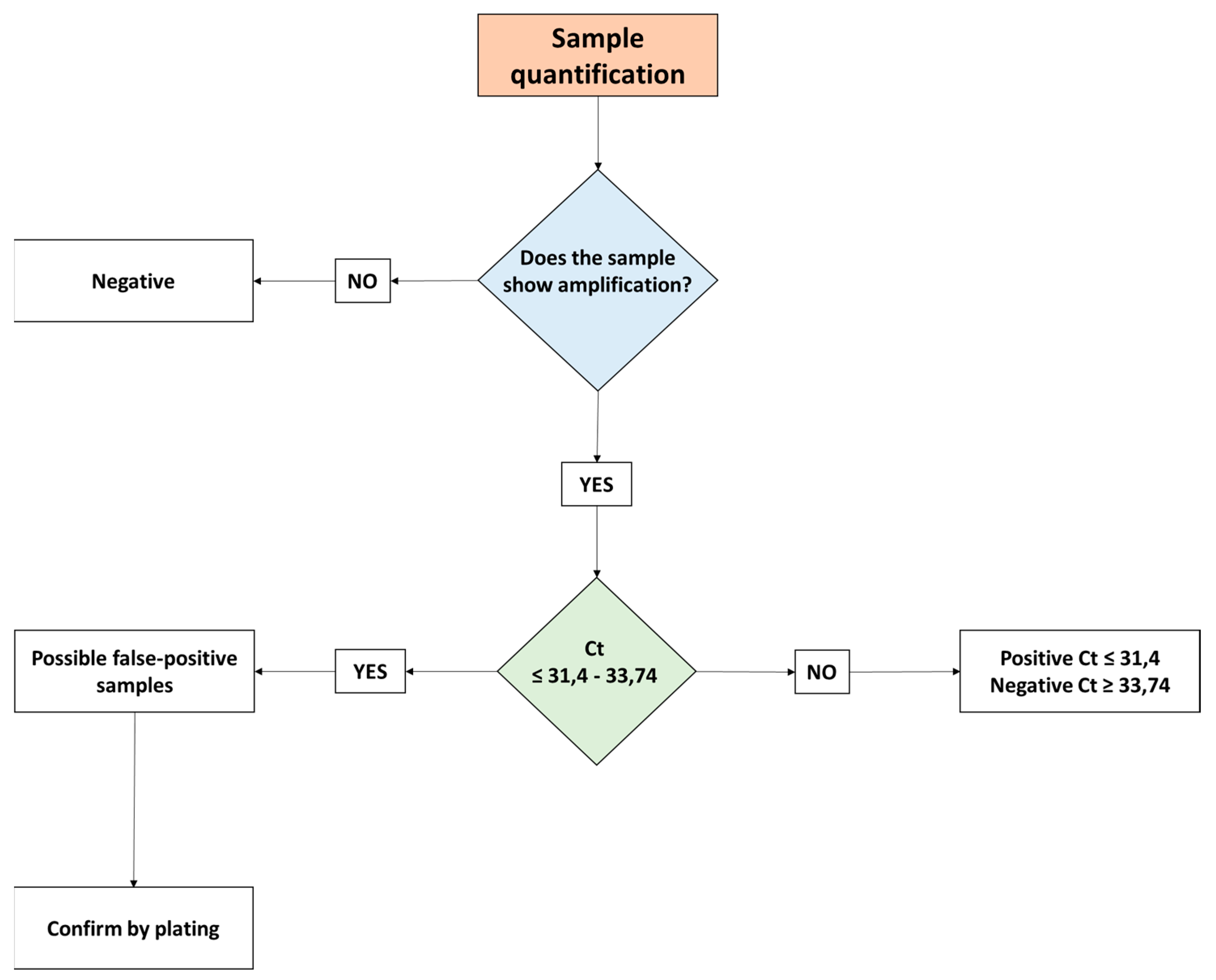
3.2. Inhibitory Effects of PMAxxTM
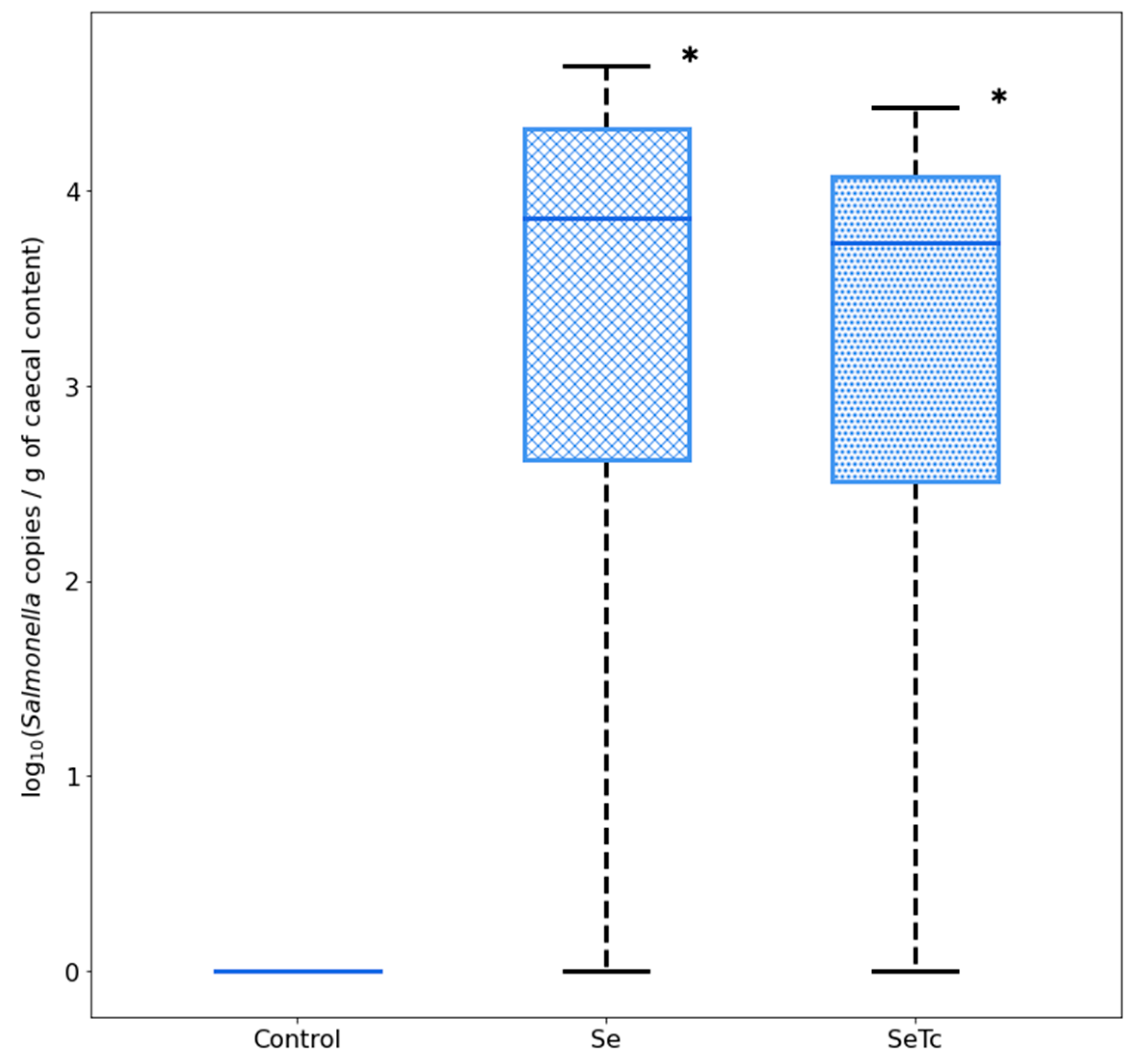
3.3. Effects of Microalgae Administration on S. Infantis Caecal Load
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Alvis Guzman, N. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Ahaduzzaman, M.; Groves, P.J.; Walkden-Brown, S.W.; Gerber, P.F. A molecular based method for rapid detection of Salmonella spp. in poultry dust samples. MethodsX 2021, 8, 101356. [Google Scholar] [CrossRef]
- Voss-Rech, D.; Vaz, C.S.L.; Alves, L.; Coldebella, A.; Leao, J.A.; Rodrigues, D.P.; Back, A. A temporal study of Salmonella enterica serotypes from broiler farms in Brazil. Poult. Sci. 2015, 94, 433–441. [Google Scholar] [CrossRef]
- Smith, S.; Seriki, A.; Ajayi, A. Typhoidal and non-typhoidal Salmonella infections in Africa. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1913–1922. [Google Scholar] [CrossRef]
- Schultz, B.M.; Melo-Gonzalez, F.; Salazar, G.A.; Porto, B.N.; Riedel, C.A.; Kalergis, A.M.; Bueno, S.M. New Insights on the Early Interaction Between Typhoid and Non-typhoid Salmonella Serovars and the Host Cells. Front. Microbiol. 2021, 12, 647044. [Google Scholar] [CrossRef]
- WHO. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiolgy Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar] [CrossRef]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Multi-country outbreak of Salmonella Enteritidis sequence type (ST)11 infections linked to poultry products in the EU/EEA and the United Kingdom. EFSA Support. Publ. 2021, 18, 1–29. [Google Scholar] [CrossRef]
- Vinueza-Burgos, C.; Cevallos, M.; Ron-Garrido, L.; Bertrand, S.; De Zutter, L. Prevalence and diversity of Salmonella serotypes in ecuadorian broilers at slaughter age. PLoS ONE 2016, 11, e0159567. [Google Scholar] [CrossRef] [PubMed]
- Vinueza-Burgos, C.; Baquero, M.; Medina, J.; De Zutter, L. Occurrence, genotypes and antimicrobial susceptibility of Salmonella collected from the broiler production chain within an integrated poultry company. Int. J. Food Microbiol. 2019, 299, 1–7. [Google Scholar] [CrossRef]
- Nagy, T.; Szmolka, A.; Wilk, T.; Kiss, J.; Szabó, M.; Pászti, J.; Nagy, B.; Olasz, F. Comparative Genome Analysis of Hungarian and Global Strains of Salmonella Infantis. Front. Microbiol. 2020, 11, 539. [Google Scholar] [CrossRef] [PubMed]
- Medina-Santana, J.L.; Ortega-Paredes, D.; de Janon, S.; Burnett, E.; Ishida, M.; Sauders, B.; Stevens, M.; Vinueza-Burgos, C. Investigating the dynamics of Salmonella contamination in Integrated Poultry Companies. Poult. Sci. 2022, 101, 101611. [Google Scholar] [CrossRef]
- Torres-Elizalde, L.; Ortega-Paredes, D.; Loaiza, K.; Fernández-Moreira, E.; Larrea-Álvarez, M. In Silico Detection of Antimicrobial Resistance Integrons in Salmonella enterica Isolates from Countries of the Andean Community. Antibiotics 2021, 10, 1388. [Google Scholar] [CrossRef] [PubMed]
- Ricke, S.C. Strategies to improve poultry food safety, a landscape review. Annu. Rev. Anim. Biosci. 2021, 9, 379–400. [Google Scholar] [CrossRef] [PubMed]
- Schneitz, C.; Koivunen, E.; Tuunainen, P.; Valaja, J. The effects of a competitive exclusion product and two probiotics on Salmonella colonization and nutrient digestibility in broiler chickens. J. Appl. Poult. Res. 2016, 25, 396–406. [Google Scholar] [CrossRef]
- Shanmugasundaram, R.; Applegate, T.J.; Selvaraj, R.K. Effect of Bacillus subtilis and Bacillus licheniformis probiotic supplementation on cecal Salmonella load in broilers challenged with salmonella. J. Appl. Poult. Res. 2020, 29, 808–816. [Google Scholar] [CrossRef]
- Omar, A.E.; Al-Khalaifah, H.S.; Mohamed, W.A.M.; Gharib, H.S.A.; Osman, A.; Al-Gabri, N.A.; Amer, S.A. Effects of Phenolic-Rich Onion (Allium cepa L.) Extract on the Growth Performance, Behavior, Intestinal Histology, Amino Acid Digestibility, Antioxidant Activity, and the Immune Status of Broiler Chickens. Front. Vet. Sci. 2020, 7, 582612. [Google Scholar] [CrossRef]
- Liu, W.C.; Zhu, Y.R.; Zhao, Z.H.; Jiang, P.; Yin, F.Q. Effects of dietary supplementation of algae-derived polysaccharides on morphology, tight junctions, antioxidant capacity and immune response of duodenum in broilers under heat stress. Animals 2021, 11, 2279. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.A.; Larrea-Álvarez, M.; Larrea-Álvarez, C.M.; Karaffová, V.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Ševčíková, Z.; Levkut, M.; Herich, R.; Revajová, V. The probiotic Lactobacillus fermentum Biocenol CCM 7514 moderates Campylobacter jejuni-induced body weight impairment by improving gut morphometry and regulating cecal cytokine abundance in broiler chickens. Animals 2021, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Karaffová, V.; Revajová, V.; Gancarčíková, S.; Ševčíková, Z.; Herich, R. Lactobacillus fermentum administration modulates cytokine expression and lymphocyte subpopulation levels in broiler chickens challenged with Campylobacter coli. Foodborne Pathog. Dis. 2020, 17, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Revajová, V.; Karaffová, V.; Koščová, J.; Nemcová, R.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Levkut, M.; et al. Effects of Lactobacillus fermentum supplementation on body weight and pro-inflammatory cytokine expression in Campylobacter jejuni-challenged chickens. Vet. Sci. 2020, 7, 121. [Google Scholar] [CrossRef]
- Coudert, E.; Baéza, E.; Berri, C. Use of algae in poultry production: A review. Worlds Poult. Sci. J. 2020, 4, 767–786. [Google Scholar] [CrossRef]
- Šefcová, M.A.; Santacruz, F.; Larrea-Álvarez, C.M.; Vinueza-Burgos, C.; Ortega-Paredes, D.; Molina-Cuasapaz, G.; Rodríguez, J.; Calero-Cáceres, W.; Revajová, V.; Fernández-Moreira, E.; et al. Administration of Dietary Microalgae Ameliorates Intestinal Parameters, Improves Body Weight, and Reduces Thawing Loss of Fillets in Broiler Chickens: A Pilot Study. Animals 2021, 11, 3601. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; Costa de Morais, R.M.S.; Bernando de Morais, A.M.M. Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Bernaerts, T.M.; Gheysen, L.; Kyomugasho, C.; Kermani, Z.J.; Vandionant, S.; Foubert, I.; Hendrickx, M.E.; Van Loey, A.M. Comparison of microalgal biomasses as functional food ingredients: Focus on the composition of cell wall related polysaccharides. Algal Res. 2018, 32, 150–161. [Google Scholar] [CrossRef]
- Sardari, R.R.; Nordberg Karlsson, E. Marine poly-and oligosaccharides as prebiotics. J. Agric. Food Chem. 2018, 66, 11544–11549. [Google Scholar] [CrossRef] [PubMed]
- Kulshreshtha, G.; Rathgeber, B.; MacIsaac, J.; Boulianne, M.; Brigitte, L.; Stratton, G.; Thomas, N.A.; Critchley, A.T.; Hafting, J.; Prithiviraj, B. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, reduce Salmonella Enteritidis in laying hens. Front. Microbiol. 2017, 8, 567. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Borza, T.; Rathgeber, B.; Stratton, G.S.; Thomas, N.A.; Critchley, A.; Hafting, J.; Prithiviraj, B. Red seaweeds Sarcodiotheca gaudichaudii and Chondrus crispus down regulate virulence factors of Salmonella enteritidis and induce immune responses in Caenorhabditis elegans. Front. Microbiol. 2016, 7, 421. [Google Scholar] [CrossRef]
- Lu, L.; Wang, J.; Yang, G.; Zhu, B.; Pan, K. Heterotrophic growth and nutrient productivities of Tetraselmis chuii using glucose as a carbon source under different C/N ratios. J. Appl. Phycol. 2017, 29, 15–21. [Google Scholar] [CrossRef]
- Isfahani, N.H.; Rahimi, S.; Rasaee, M.J.; Torshizi, M.A.K.; Salehi, T.Z.; Grimes, J.L. The effect of capsulated and noncapsulated egg-yolk–specific antibody to reduce colonization in the intestine of Salmonella enterica ssp. enterica serovar Infantis–challenged broiler chickens. Poult. Sci. 2020, 99, 1387–1394. [Google Scholar] [CrossRef]
- Awang, M.S.; Bustami, Y.; Hamzah, H.H.; Zambry, N.S.; Najib, M.A.; Khalid, M.F.; Aziah, I.; Abd Manaf, A. Advancement in Salmonella Detection Methods: From Conventional to Electrochemical-Based Sensing Detection. Biosensors 2021, 11, 346. [Google Scholar] [CrossRef]
- Paniel, N.; Noguer, T. Detection of Salmonella in Food Matrices, from Conventional Methods to Recent Aptamer-Sensing Technologies. Foods 2019, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Connolly, C. Use of Viability qPCR for Quantification of Salmonella Typhimurium and Listeria Monocytogenes in Food Safety Challenge Studies; Pennsylvania State University: State College, PA, USA, 2021. [Google Scholar]
- Xiao, L.; Zhang, Z.; Sun, X.; Pan, Y.; Zhao, Y. Development of a quantitative real-time PCR assay for viable Salmonella spp. without enrichment. Food Control. 2015, 57, 185–189. [Google Scholar] [CrossRef]
- Zhang, J.; Khan, S.; Chousalkar, K.K. Development of PMAxxTM-Based qPCR for the Quantification of Viable and Non-viable Load of Salmonella from Poultry Environment. Front. Microbiol. 2020, 11, 581201. [Google Scholar] [CrossRef] [PubMed]
- Geneious Prime. Available online: https://www.geneious.com/ (accessed on 31 August 2021).
- Rahn, K.; De Grandis, S.A.; Clarke, R.C.; McEwen, S.A.; Galan, J.E.; Ginocchio, C.; Curtiss, R.; Gyles, C.L. Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell. Probes. 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Strain-Specific Bacterial Viability PCR Kits. Biotium. Available online: https://biotium.com/wp-content/uploads/2016/12/PI-Strain-Specific-Bacterial-Viability-PCR-Kits.pdf (accessed on 17 August 2021).
- Lazic, S.E.; Clarke-Williams, C.J.; Munafò, M.R. What exactly is ‘N’ in cell culture and animal experiments? PLoS Biol. 2018, 16, e2005282. [Google Scholar] [CrossRef]
- Cobb500 Broiler Performance and Nutrition Supplement. Available online: https://www.cobb-vantress.com/resource/featured?q=nutrition (accessed on 3 September 2021).
- Kang, H.K.; Salim, H.M.; Akter, N.; Kim, D.W.; Kim, J.H.; Bang, H.T.; Kim, M.J.; Na, J.C.; Hwangbo, J.; Choi, H.C.; et al. Effect of various forms of dietary Chlorella supplementation on growth performance, immune characteristics, and intestinal microflora population of broiler chickens. J. Appl. Poult. Res. 2013, 22, 100–108. [Google Scholar] [CrossRef]
- He, T.; Zhu, Y.H.; Yu, J.; Xia, B.; Liu, X.; Yang, G.Y.; Su, J.H.; Guo, L.; Wang, M.L.; Wang, J.F. Lactobacillus johnsonii L531 reduces pathogen load and helps maintain short-chain fatty acid levels in the intestines of pigs challenged with Salmonella enterica Infantis. Vet. Microbiol. 2019, 230, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Torok, V.A.; Ophel-Keller, K.; Loo, M.; Hughes, R.J. Application of methods for identifying broiler chicken gut bacterial species linked with increased energy metabolism. Appl. Environ. Microbiol. 2008, 74, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.K.; Bradshaw, R.; Nyarko, E.; Handy, E.T.; East, C.; Millner, P.D.; Bergholz, T.M.; Sharma, M. Salmonella enterica in soils amended with heat-treated poultry pellets survived longer than bacteria in unamended soils and more readily transferred to and persisted on spinach. Appl. Environ. Microbiol. 2019, 85, e00334-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dilution | Ct1x | Ct2x | Ct3x | Average Ct | SD | CV (%) |
|---|---|---|---|---|---|---|
| −1 | 16.93 | 16.94 | 17.07 | 16.98 | 0.06 | 0.38 |
| −2 | 20.19 | 20.49 | 20.88 | 20.52 | 0.28 | 1.38 |
| −3 | 23.97 | 24.26 | 24.25 | 24.16 | 0.13 | 1.54 |
| −4 | 28.83 | 28.81 | 28.88 | 28.84 | 0.03 | 0.10 |
| −5 | 32.56 | 32.46 | 32.62 | 32.55 | 0.07 | 0.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corrales-Martinez, J.; Ortega-Paredes, D.; Šefcová, M.A.; Larrea-Álvarez, C.M.; de Janon, S.; Medina-Santana, J.; Molina-Cuasapaz, G.; Vinueza-Burgos, C.; Revajová, V.; Larrea-Álvarez, M.; et al. A PMAxxTM qPCR Assay Reveals That Dietary Administration of the Microalgae Tetraselmis chuii Does Not Affect Salmonella Infantis Caecal Content in Early-Treated Broiler Chickens. Vet. Sci. 2022, 9, 487. https://doi.org/10.3390/vetsci9090487
Corrales-Martinez J, Ortega-Paredes D, Šefcová MA, Larrea-Álvarez CM, de Janon S, Medina-Santana J, Molina-Cuasapaz G, Vinueza-Burgos C, Revajová V, Larrea-Álvarez M, et al. A PMAxxTM qPCR Assay Reveals That Dietary Administration of the Microalgae Tetraselmis chuii Does Not Affect Salmonella Infantis Caecal Content in Early-Treated Broiler Chickens. Veterinary Sciences. 2022; 9(9):487. https://doi.org/10.3390/vetsci9090487
Chicago/Turabian StyleCorrales-Martinez, Joselyn, David Ortega-Paredes, Miroslava Anna Šefcová, César Marcelo Larrea-Álvarez, Sofía de Janon, José Medina-Santana, Gabriel Molina-Cuasapaz, Christian Vinueza-Burgos, Viera Revajová, Marco Larrea-Álvarez, and et al. 2022. "A PMAxxTM qPCR Assay Reveals That Dietary Administration of the Microalgae Tetraselmis chuii Does Not Affect Salmonella Infantis Caecal Content in Early-Treated Broiler Chickens" Veterinary Sciences 9, no. 9: 487. https://doi.org/10.3390/vetsci9090487
APA StyleCorrales-Martinez, J., Ortega-Paredes, D., Šefcová, M. A., Larrea-Álvarez, C. M., de Janon, S., Medina-Santana, J., Molina-Cuasapaz, G., Vinueza-Burgos, C., Revajová, V., Larrea-Álvarez, M., & Calero-Cáceres, W. (2022). A PMAxxTM qPCR Assay Reveals That Dietary Administration of the Microalgae Tetraselmis chuii Does Not Affect Salmonella Infantis Caecal Content in Early-Treated Broiler Chickens. Veterinary Sciences, 9(9), 487. https://doi.org/10.3390/vetsci9090487