Resistance and Pathogenicity of Salmonella Thompson Isolated from Incubation End of a Poultry Farm


Abstract
:Simple Summary
Abstract
1. Introduction
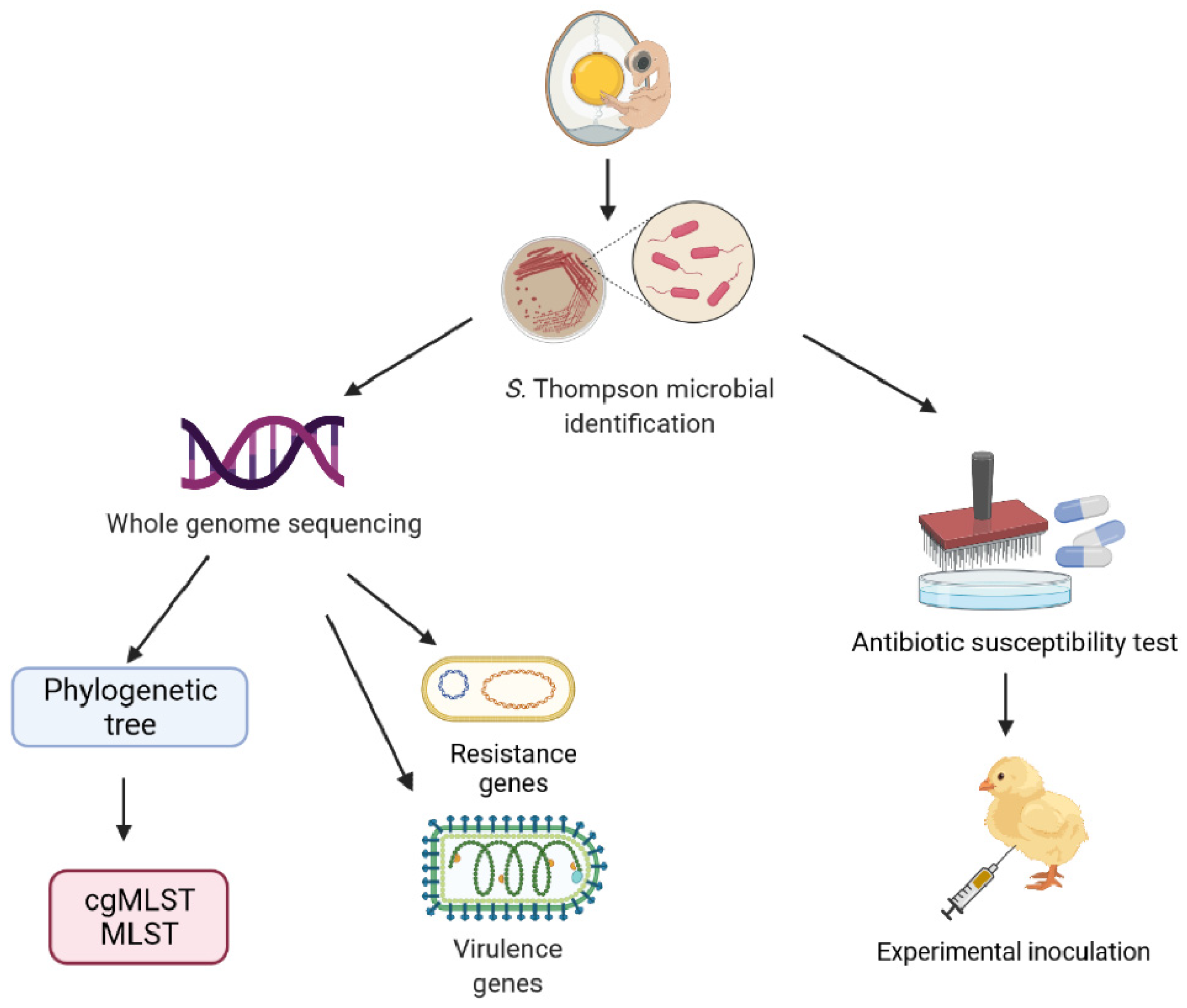
2. Materials and Methods
2.1. Strains
2.2. Microbiological Analysis
2.3. DNA Extraction and Whole Genome Sequencing
2.4. Molecular Typing and Phylogenetic Tree
2.5. Resistance Genes and Virulence Factors
2.6. Antimicrobial Susceptibility Testing
2.7. Experimental Inoculation
2.8. Sampling and Necropsy
2.9. Statistical Analysis
2.10. Ethics Statement
3. Results
3.1. Biochemical Characteristics and Serotypes
3.2. Phylogenetic Tree, MLST and cgMLST
3.3. Antimicrobial Resistance Genes and Virulence Genes
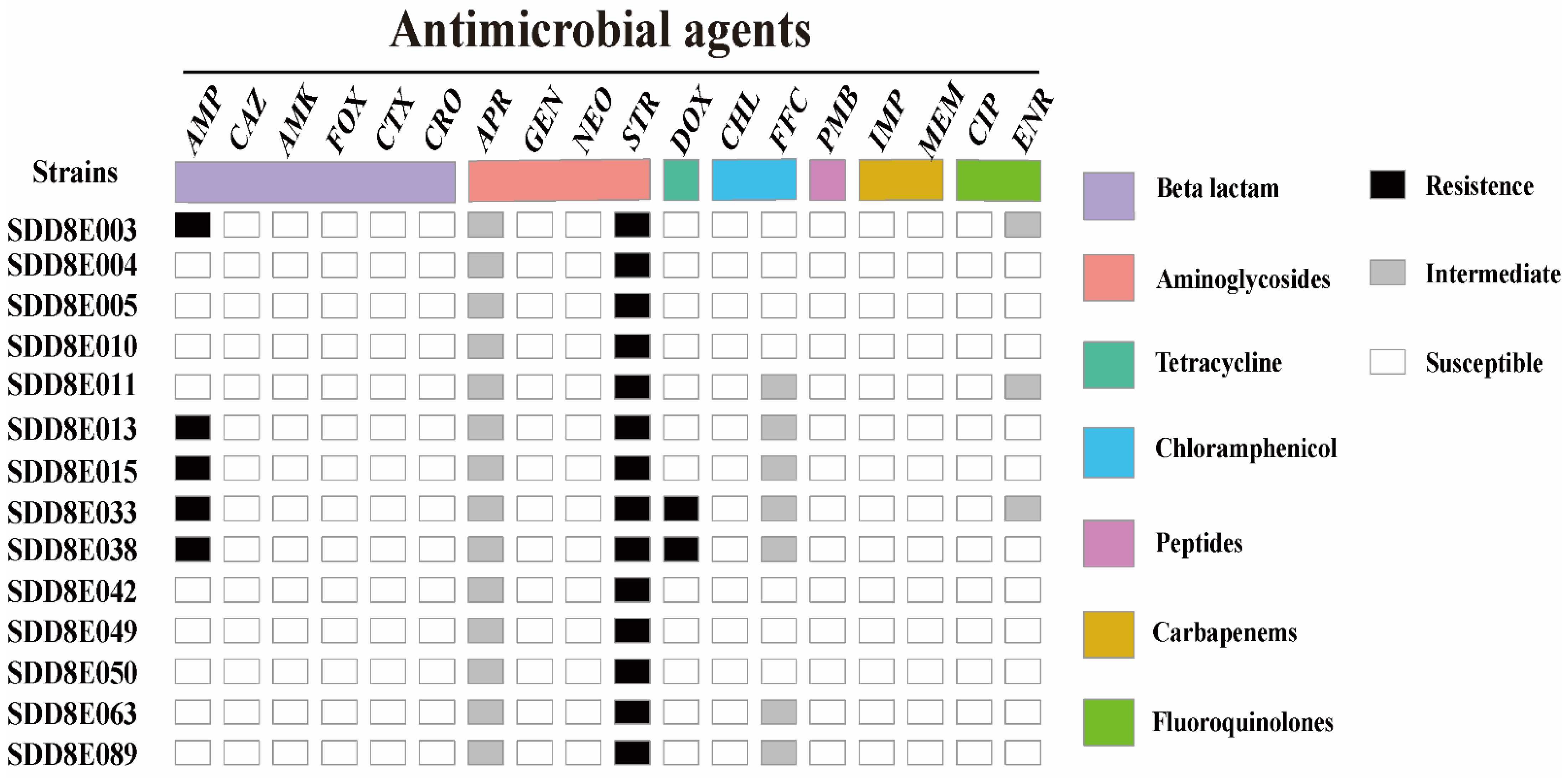
3.4. Antimicrobial Susceptibility
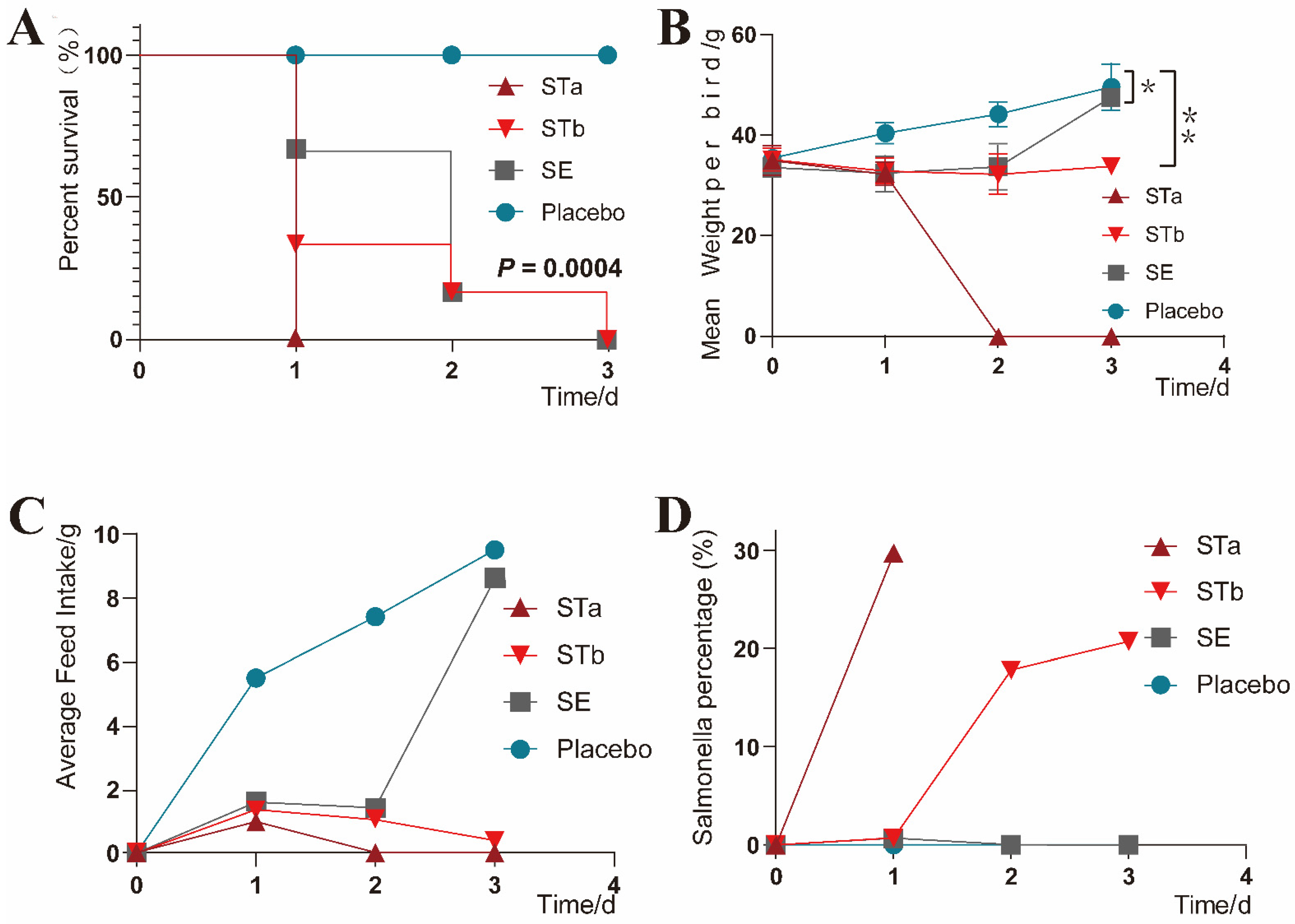
3.5. Clinical Monitoring
3.6. Necropsy and Pathological Changes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, X.; Yang, J.; Zhang, B.; Sun, S.; Chang, W. Characterization of integrons and resistance genes in Salmonella isolates from farm animals in shandong province, China. Front. Microbiol. 2017, 8, article 1300. [Google Scholar] [CrossRef] [PubMed]
- Müştak, İ.B.; Yardımcı, H. Construction and in vitro characterisation of aroA defective (aroAΔ) mutant Salmonella Infantis. Arch. Microbiol. 2019, 201, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Velge, P.; Wiedemann, A.; Rosselin, M.; Abed, N.; Boumart, Z.; Chaussé, A.M.; Grépinet, O.; Namdari, F.; Roche, S.M.; Rossignol, A.; et al. Multiplicity of Salmonella entry mechanisms, a new paradigm for Salmonella pathogenesis. MicrobiologyOpen 2012, 1, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Gorki, H.; Nicolay, N.H.; Loulmet, D.F.; Patel, N.C.; Ciuffol, G.B.; Subramanian, V.A.; Lessnau, K.D. Non-typhoid Salmonellae and prosthetic valve endocarditis: More than a rare coincidence? A review of the literature. J. Heart Valve Dis. 2009, 18, 401–410. [Google Scholar]
- Rincon-Gamboa, S.M.; Poutou-Pinales, R.A.; Carrascal-Camacho, A.K. Analysis of the assessment of antimicrobial susceptibility. Non-typhoid Salmonella in meat and meat products as model (systematic review). BMC Microbiol. 2021, 21, 223. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [Green Version]
- Rincón-Gamboa, S.M.; Poutou-Piñales, R.A.; Carrascal-Camacho, A.K. Antimicrobial resistance of Non-Typhoid Salmonella in meat and meat products. Foods 2021, 10, 1731. [Google Scholar] [CrossRef]
- Scott Quainoo, J.P.M.C. Whole-Genome sequencing of bacterial pathogens: The future of nosocomial outbreak analysis. Clin. Microbiol. Rev. 2017, 30, 1015–1063. [Google Scholar] [CrossRef] [Green Version]
- Salipante, S.J.; Sengupta, D.J.; Cummings, L.A.; Land, T.A.; Hoogestraat, D.R.; Cookson, B.T. Application of Whole-Genome sequencing for bacterial strain typing in molecular epidemiology. J. Clin. Microbiol. 2015, 53, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- Rumore, J.; Tschetter, L.; Kearney, A.; Kandar, R.; Mccormick, R.; Walker, M.; Peterson, C.; Reimer, A.; Nadon, C. Evaluation of whole-genome sequencing for outbreak detection of Verotoxigenic Escherichia coli O157:H7 from the Canadian perspective. BMC Genom. 2018, 19, 870. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, W.; Qin, T.; Liu, C.; Ren, H. Defining and evaluating a core genome multilocus sequence typing scheme for Whole-Genome Sequence-Based typing of Klebsiella pneumoniae. Front. Microbiol. 2017, 8, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.; Wu, C.; Wu, C.; Qi, J.; Wang, Y.; Wang, H.; Liu, Y.; Shen, J. Serotype distribution and antibiotic resistance of Salmonella in food-producing animals in Shandong province of China, 2009 and 2012. Int. J. Food Microbiol. 2014, 180, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qi, Y.; Deng, T.; Lu, J.; Chen, Y.; Zhang, C.; Liu, Y.; Tong, G.; Xing, Y. Isolation and identification of Salmonella spp. from dead embryos of hemp-bred chickens in Henan and analysis of drug resistance. Chin. J. Prev. Vet. Med. 2018, 40, 891–896. [Google Scholar]
- Friesema, I.H.; de Jong, A.E.; Fitz, J.I.; Heck, M.E.; van den Kerkhof, J.H.; Notermans, D.W.; van Pelt, W.; Hofhuis, A. Outbreak of Salmonella Thompson in the Netherlands since July 2012. Eurosurveillance 2012, 17, 20303. [Google Scholar] [CrossRef]
- Gaulin, C.; Fiset, M.; Duchesne, C.; Ramsay, D.; Savard, N.; Urbanek, A.; Pilon, P.A.; Usongo, V.; Bekal, S. Salmonella Thompson outbreak associated with consumption of chicken shawarma and the usefulness of genome sequencing in the investigation. Can. Commun. Dis. Rep. 2017, 43, 186–192. [Google Scholar] [CrossRef]
- Eun, Y.; Jeong, H.; Kim, S.; Park, W.; Ahn, B.; Kim, D.; Kim, E.; Park, E.; Park, S.; Hwang, I.; et al. A large outbreak of Salmonella enterica serovar Thompson infections associated with chocolate cake in Busan, Korea. Epidemiol. Health 2019, 41, e2019002. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Ju, Z.; Wang, G.; Yang, J.; Wang, F.; Tang, H.; Zhao, X.; Sun, S. Prevalence and antimicrobial resistance of salmonella isolated from Dead-in-Shell chicken embryos in shandong, China. Front. Vet. Sci. 2021, 8, article 581946. [Google Scholar] [CrossRef]
- Elbediwi, M.; Tang, Y.; Shi, D.; Ramadan, H.; Xu, Y.; Xu, S.; Li, Y.; Yue, M. Genomic investigation of Antimicrobial-Resistant salmonella enterica isolates from dead chick embryos in China. Front. Microbiol. 2021, 12, article 684400. [Google Scholar] [CrossRef]
- Chisholm, S.A.; Crichton, P.B.; Knight, H.I.; Old, D.C. Molecular typing of Salmonella serotype Thompson strains isolated from human and animal sources. Epidemiol. Infect. 1999, 122, 33–39. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Ishida, Y.; Shimamoto, T. Molecular characterization of antimicrobial resistance in Salmonella isolated from animals in Japan. J. Appl. Microbiol. 2009, 106, 402–409. [Google Scholar] [CrossRef]
- Makino, S.O.U.O.; Kurazono, H.; Chongsanguam, M.; Hayashi, H.; Cheun, H.; Suzuki, S.; Shirahata, T. Establishment of the PCR system specific to Salmonella spp. And its application for the inspection of food and fecal samples. J. Vet. Med. Sci. 1999, 61, 1245–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Zhou, X.; Chai, W.; Paudyal, N.; Li, S.; Zhou, X.; Zhou, K.; Wu, Q.; Wu, B.; Li, G.; et al. Diversified sources for human infections by Salmonella enterica serovar newport. Transbound. Emerg. Dis. 2019, 66, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Ed-Dra, A.; Pan, H.; Dong, C.; Jia, C.; Yue, M. Genomic investigation of Salmonella isolates recovered from a pig slaughtering process in hangzhou, China. Front. Microbiol. 2021, 12, article 704636. [Google Scholar] [CrossRef]
- Mei-Han, Q.W.; Baiyintala, W. The whole-genome sequence analysis of Morchella sextelata. Sci. Rep. 2019, 9, 15376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to Single-Cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Joensen, K.G.; Zankari, E.; Ahrenfeldt, J.; Lukjancenko, O.; Kaas, R.S.; Roer, L.; Leekitcharoenphon, P.; Saputra, D.; Cosentino, S.; et al. The CGE Tool Box; Springer International Publishing: Cham, Switzerland, 2017; pp. 65–90. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, L. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2004, 33, D325–D328. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Stephens, J.F.; Anderson, E.L. Growth of Salmonella in chickens’ yolk sacs and its relationship to pathogenicity. Appl. Microbiol. 1967, 15, 1468–1472. [Google Scholar] [CrossRef]
- Dewei, C.; Huade, W.; Qiyu, G. Isolation and Identification of Salmonella Thompson from Chicks. Chin. J. Vet. Med. 1991, 2, 9–10. [Google Scholar]
- Newberry, E.A.; Jardini, T.M.; Rubio, I.; Roberts, P.D.; Babu, B.; Koike, S.T.; Bouzar, H.; Goss, E.M.; Jones, J.B.; Bull, C.T.; et al. Angular leaf spot of cucurbits is associated with genetically diverse pseudomonas syringae strains. Plant Dis. 2016, 100, 1397–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richert, K.; Brambilla, E.; Stackebrandt, E. The phylogenetic significance of peptidoglycan types: Molecular analysis of the genera Microbacterium and Aureobacterium based upon sequence comparison of gyrB, rpoB, recA and ppk and 16SrRNA genes. Syst. Appl. Microbiol. 2007, 30, 102–108. [Google Scholar] [CrossRef]
- Hata, H.; Natori, T.; Mizuno, T.; Kanazawa, I.; Eldesouky, I.; Hayashi, M.; Miyata, M.; Fukunaga, H.; Ohji, S.; Hosoyama, A.; et al. Phylogenetics of family Enterobacteriaceae and proposal to reclassify Escherichia hermannii and Salmonella subterranea as Atlantibacter hermannii and Atlantibacter subterranea gen. Nov., Comb. Nov. Microbiol. Immunol. 2016, 60, 303–311. [Google Scholar] [CrossRef]
- Zhujun, Y.M.Z.W. Whole genome sequence-based typing methods for Neisseria meningitidis. Acta Microbiol. Sin. 2021, 61, 1910–1919. [Google Scholar]
- Tyson, G.H.; Mcdermott, P.F.; Li, C.; Chen, Y.; Tadesse, D.A.; Mukherjee, S.; Bodeis-Jones, S.; Kabera, C.; Gaines, S.A.; Loneragan, G.H.; et al. WGS accurately predicts antimicrobial resistance in Escherichia coli. J. Antimicrob. Chemoth. 2015, 70, 2763–2769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Tyson, G.H.; Chen, Y.; Li, C.; Mukherjee, S.; Young, S.; Lam, C.; Folster, J.P.; Whichard, J.M.; Mcdermott, P.F. Whole-Genome sequencing analysis accurately predicts antimicrobial resistance phenotypes in Campylobacter spp. Appl. Environ. Microb. 2016, 82, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Salipante, S.J. Determining the limits of the evolutionary potential of an antibiotic resistance gene. Mol. Biol. Evol. 2003, 20, 653–659. [Google Scholar] [CrossRef]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M.; Turner, M.S. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. mBio 2019, 10, e819–e853. [Google Scholar] [CrossRef] [Green Version]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of mobile colistin resistance genes mcr-1 to mcr-9. J. Antimicrob. Chemoth. 2020, 75, 3087–3095. [Google Scholar] [CrossRef]
- Yang, Q.; Pogue, J.M.; Li, Z.; Nation, R.L.; Kaye, K.S.; Li, J. Agents of Last Resort an Update on Polymyxin Resistance. Infect. Dis. Clin. N. Am. 2020, 34, 723–750. [Google Scholar] [CrossRef]
- Johnson, R.; Mylona, E.; Frankel, G. Typhoidal Salmonella: Distinctive virulence factors and pathogenesis. Cell. Microbiol. 2018, 20, e12939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimi, T.; Zamirnasta, M.; Sani, M.A.; Soltan Dallal, M.M.; Nasser, A. Molecular mechanisms of Salmonella effector proteins a comprehensive review. Infect. Drug Resist. 2020, 13, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Qazi, I.H.; Wang, L.; Zhou, G.; Han, H. Salmonella virulence and immune escape. Microorganisms 2020, 8, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuche, F.J.; Sow, O.; Simon, R.; Tennant, S.M. Salmonella serogroup c: Current status of vaccines and why they are needed. Clin. Vaccine Immunol. 2016, 23, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, F.D.O.; Freitas Neto, O.C.D.; Batista, D.F.A.; Almeida, A.M.D.; Rubio, M.D.S.; Alves, L.B.R.; Vasconcelos, R.D.O.; Barrow, P.A.; Berchieri Junior, A. Contribution of flagella and motility to gut colonisation and pathogenicity of Salmonella Enteritidis in the chicken. Braz. J. Microbiol. 2017, 48, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Tesarova, M.P.; Skoupa, M.; Foltyn, M.; Tvrdon, Z.; Lichovnikova, M. Research Note: Effects of preincubation and higher initiating incubation temperature of long-term stored hatching eggs on hatchability and day-old chick and yolk sac weight. Poultry Sci. 2021, 100, 101293. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Abbreviation | Concentration Range (μg/mL) | |
|---|---|---|---|
| β-lactam | Ampicillin | AMP | 0.5~256 |
| Ceftazidine | CAZ | 0.25~128 | |
| Ceftriaxone | CRO | 0.06~32 | |
| Cefotaxime | CTX | 0.06~32 | |
| Cefoxitin | (FOX | 0.5~256 | |
| Aminoglycosides | Amikacin | AMK | 1~512 |
| Apramycin | APR | 0.25~128 | |
| Gentamicin | GEN | 0.25~128 | |
| Neomycin | NEO | 1~512 | |
| Streptomycin | STR | 1~512 | |
| Tetracycline | Doxycycline | DOX | 0.25~128 |
| Chloramphenicol | Chloramphenicol | CHL | 1~512 |
| Florfenicol | FFC | 0.25~128 | |
| Polypeptide | Polymyxin B | PMB | 0.06~32 |
| Carbapenems | Imipenem | IMP | 0.06~32 |
| Meropenem | MEM | 0.06~32 | |
| Fluoroquinolones | Ciprofloxacin | CIP | 0.015~8 |
| Enrofloxacin | ENR | 0.015~8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Liu, J.; Chen, C.; Wang, Y.; Chen, X.; Li, X.; Xu, F. Resistance and Pathogenicity of Salmonella Thompson Isolated from Incubation End of a Poultry Farm. Vet. Sci. 2022, 9, 349. https://doi.org/10.3390/vetsci9070349
Zhang J, Liu J, Chen C, Wang Y, Chen X, Li X, Xu F. Resistance and Pathogenicity of Salmonella Thompson Isolated from Incubation End of a Poultry Farm. Veterinary Sciences. 2022; 9(7):349. https://doi.org/10.3390/vetsci9070349
Chicago/Turabian StyleZhang, Jingju, Jing Liu, Chen Chen, Yufeng Wang, Xiaojie Chen, Xiubo Li, and Fei Xu. 2022. "Resistance and Pathogenicity of Salmonella Thompson Isolated from Incubation End of a Poultry Farm" Veterinary Sciences 9, no. 7: 349. https://doi.org/10.3390/vetsci9070349
APA StyleZhang, J., Liu, J., Chen, C., Wang, Y., Chen, X., Li, X., & Xu, F. (2022). Resistance and Pathogenicity of Salmonella Thompson Isolated from Incubation End of a Poultry Farm. Veterinary Sciences, 9(7), 349. https://doi.org/10.3390/vetsci9070349