
Influence of Dietary Supplementation with Boswellia serrata and Salix alba on Performance and Blood Biochemistry in Free-Range Leghorn Laying Hens

 , , , , , , ,
, , , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Study Design and Sampling
2.2. Management and Feeding
2.3. Leghorn Hens’ Performance
2.4. Mortality and Behaviour Observations
2.5. Health Checks
2.6. Serum Biochemistry
2.7. Serology for the Detection of Anti-Infectious Bronchitis Virus (IBV) Antibodies
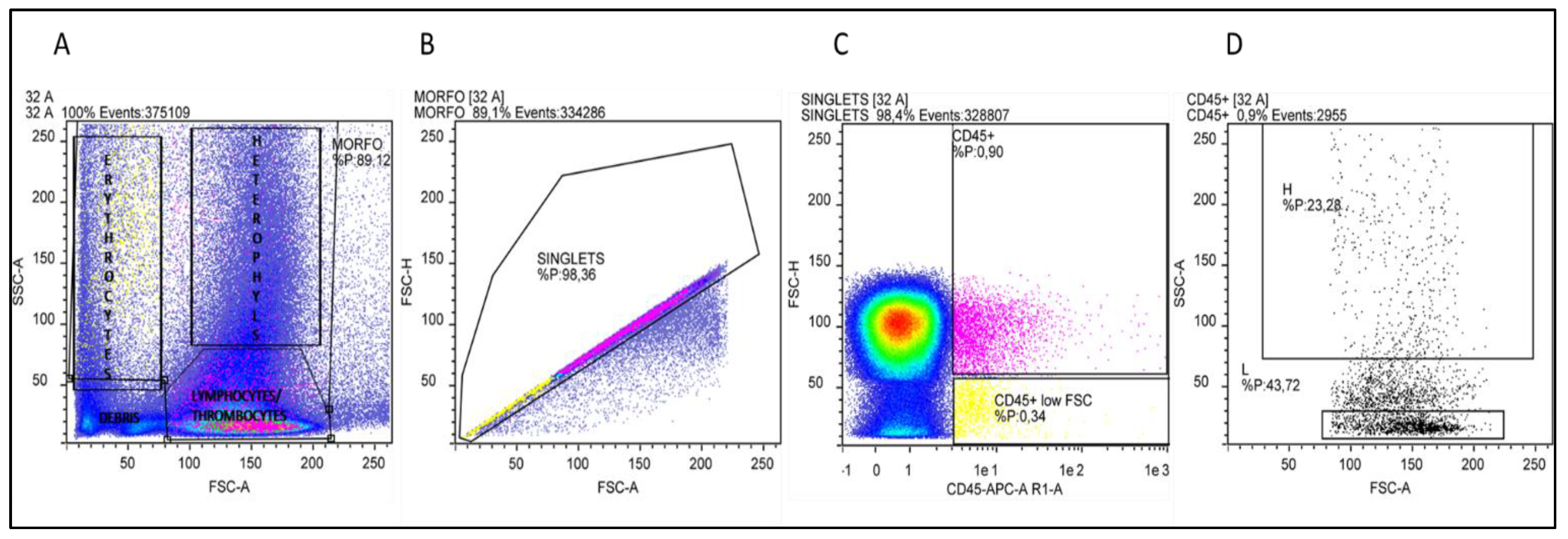
2.8. Heterophils and Lymphocytes (H/L ratio) Percentages Assessment by Flow Cytometry
2.9. IgA Determination from Cloacal Swab and Serum
2.10. Statistical Analysis
3. Results
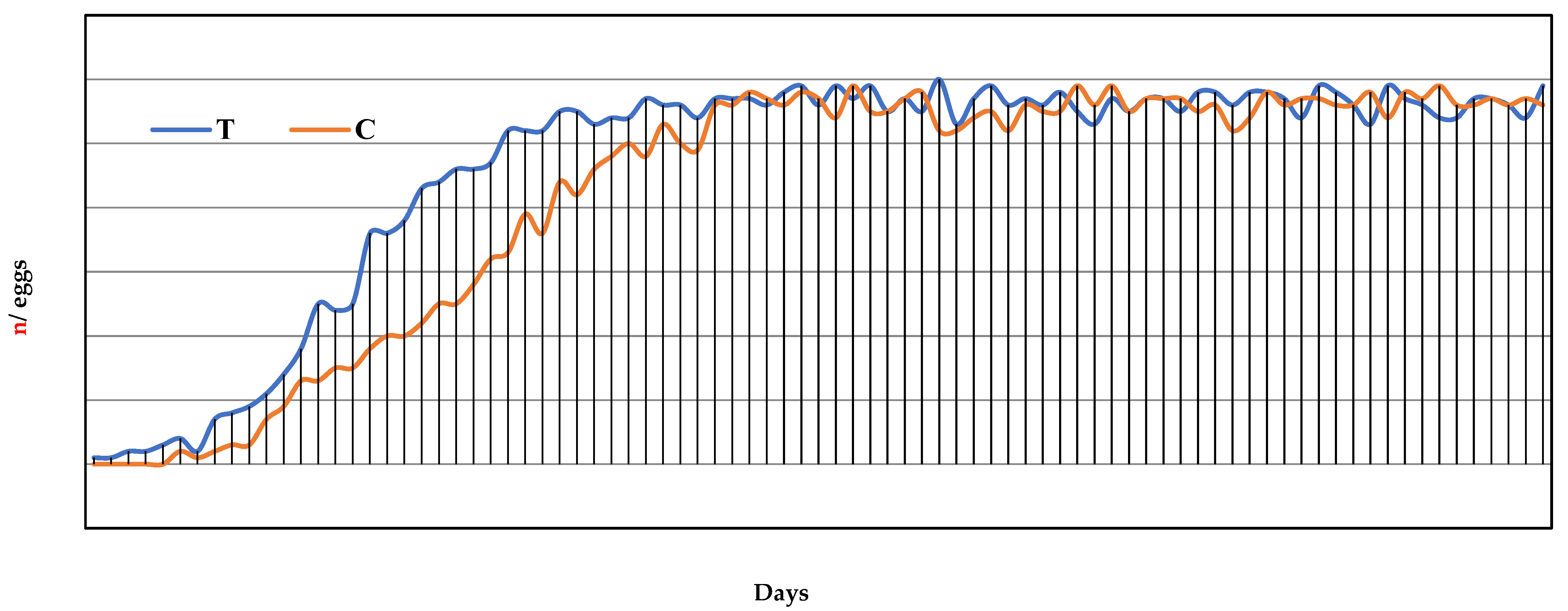
3.1. Leghorn Hens’ Performance
3.2. Mortality, Behaviour Observations and Health Checks
3.3. Serum Biochemistry
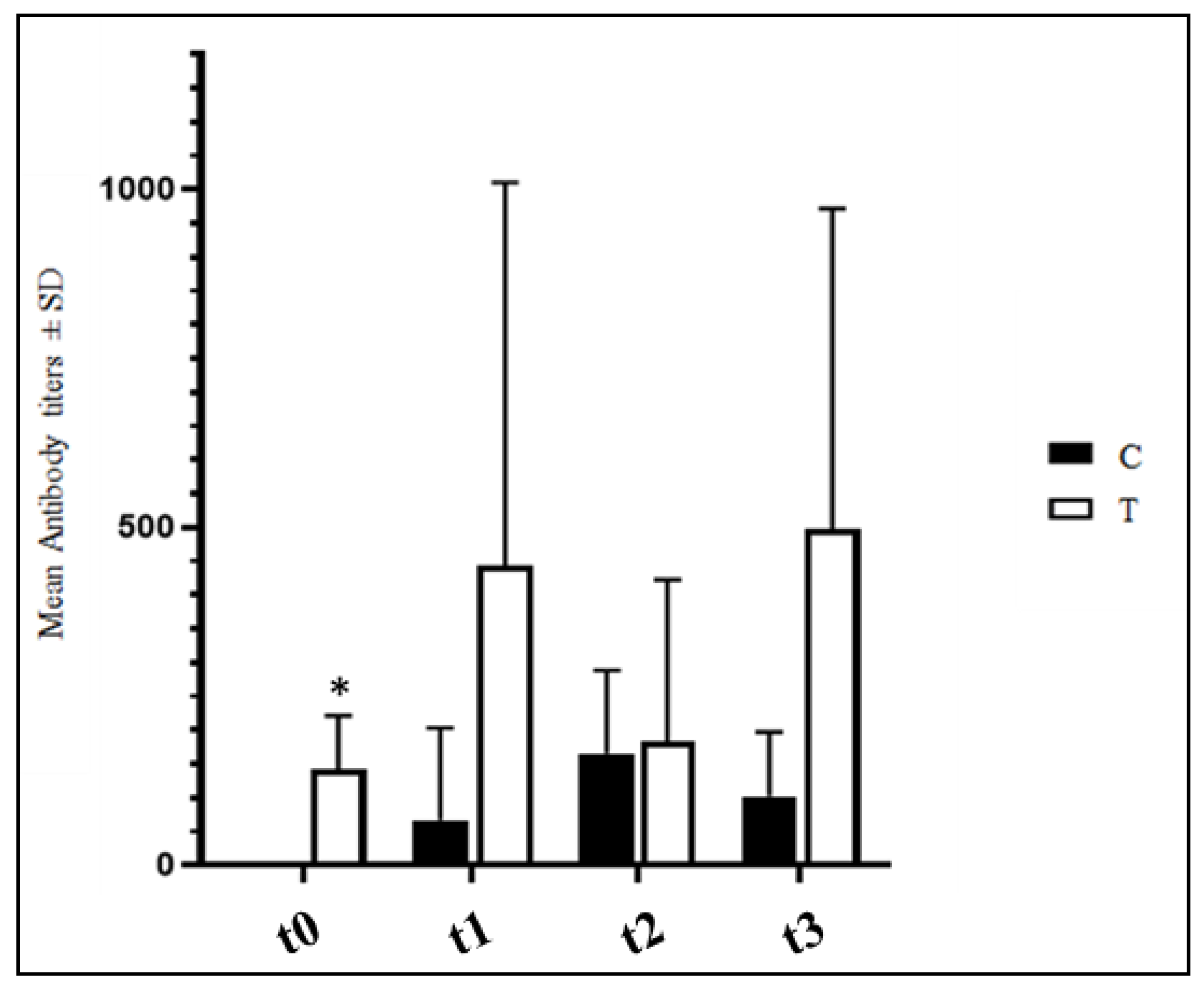
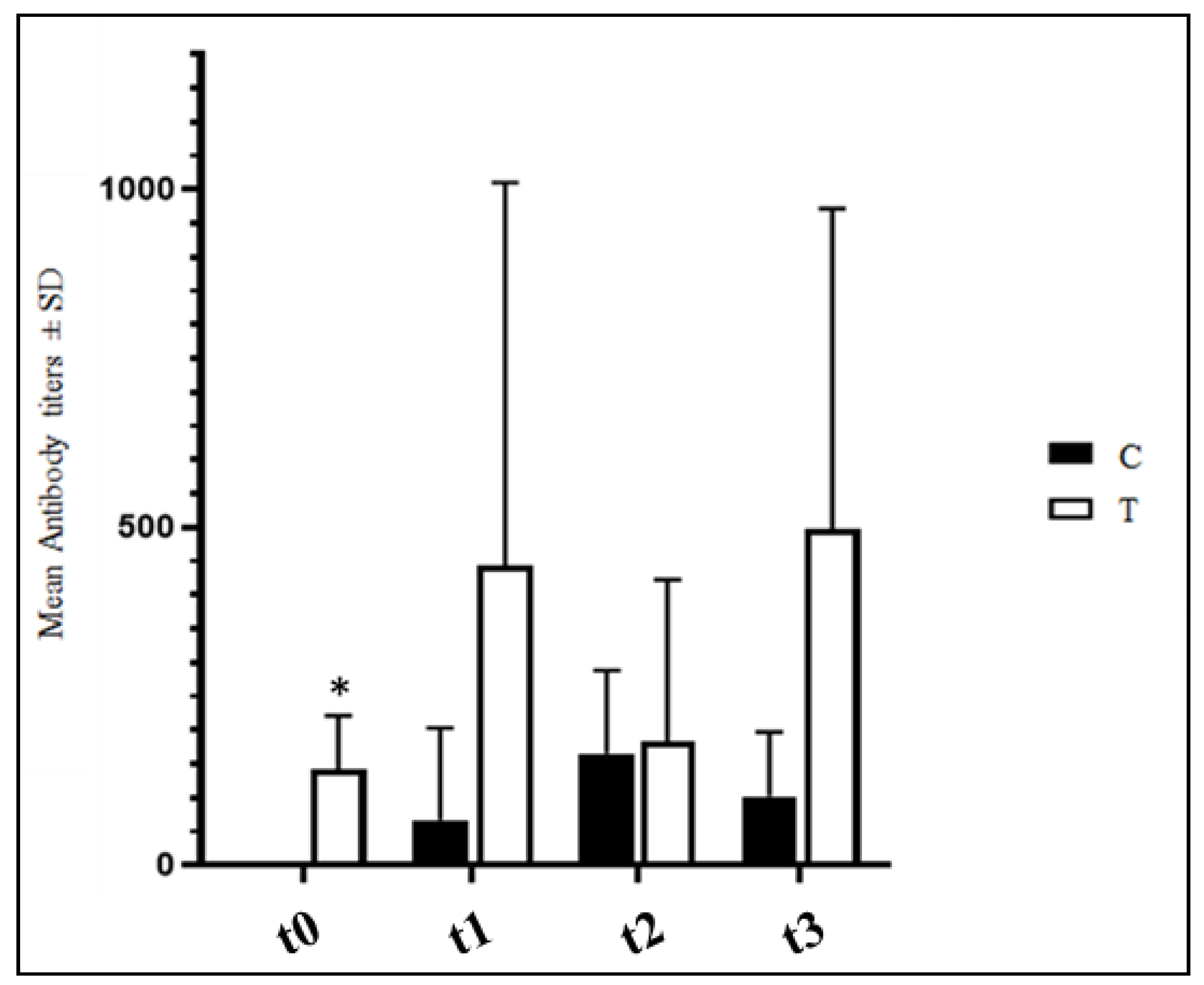
3.4. Anti-IBV Antibody Titers
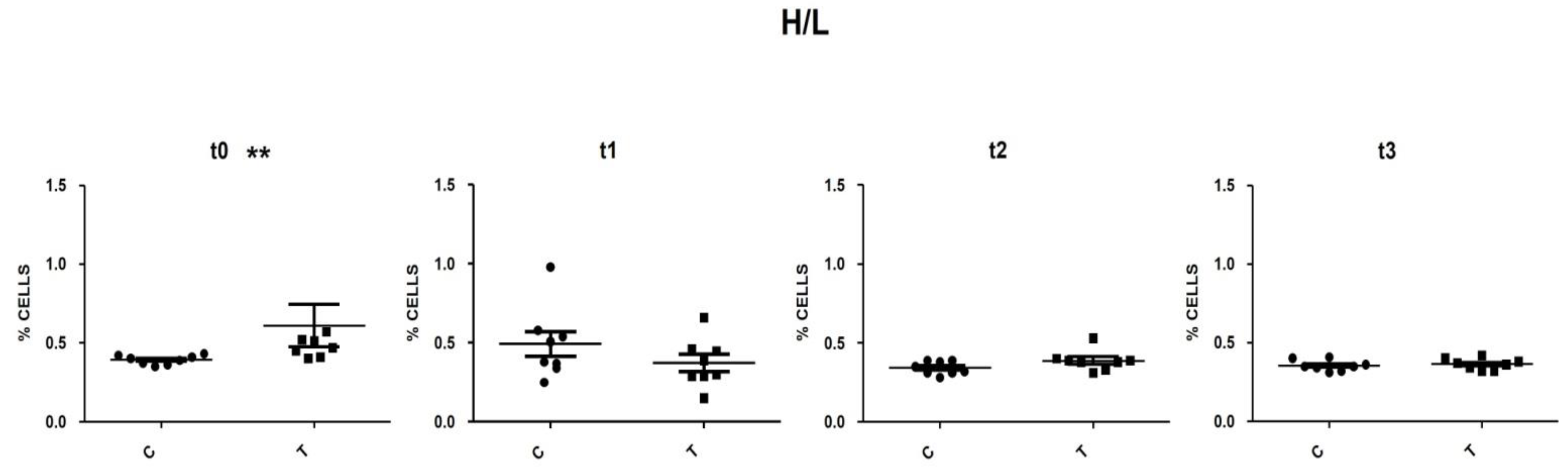
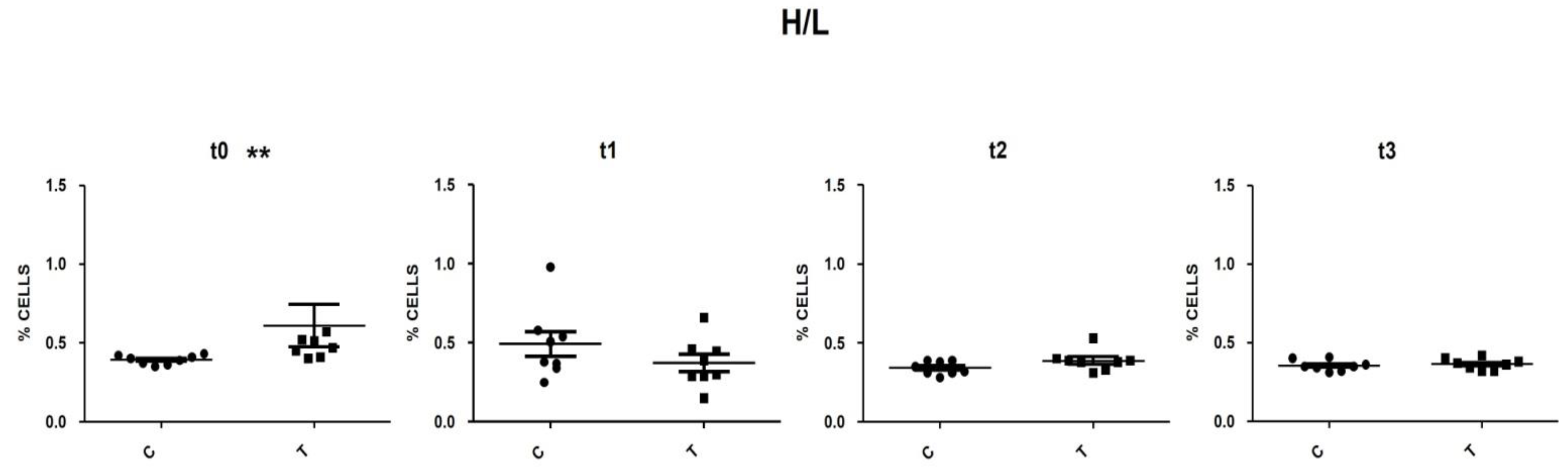
3.5. Heterophils and Lymphocytes (H/L) Ratio
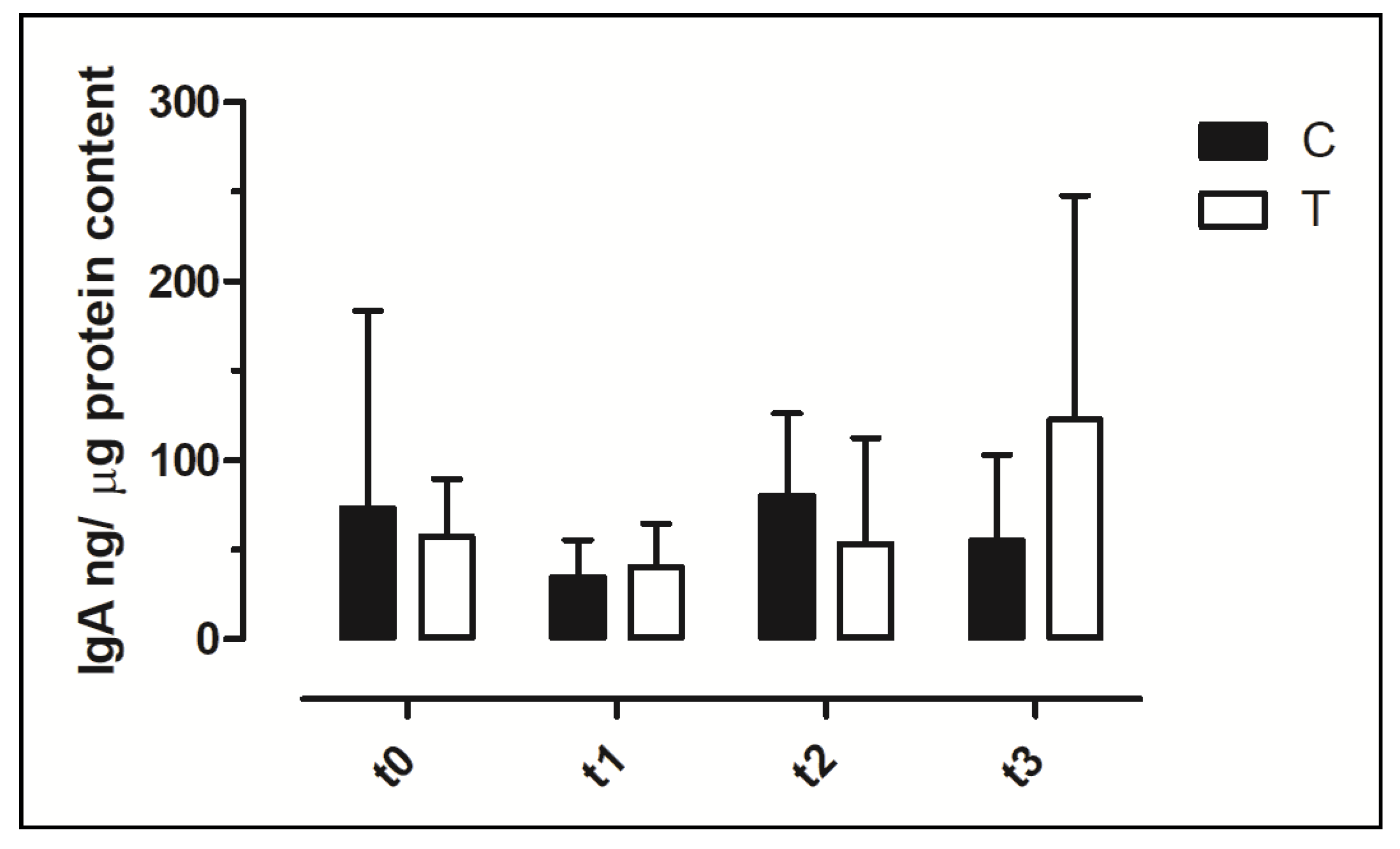
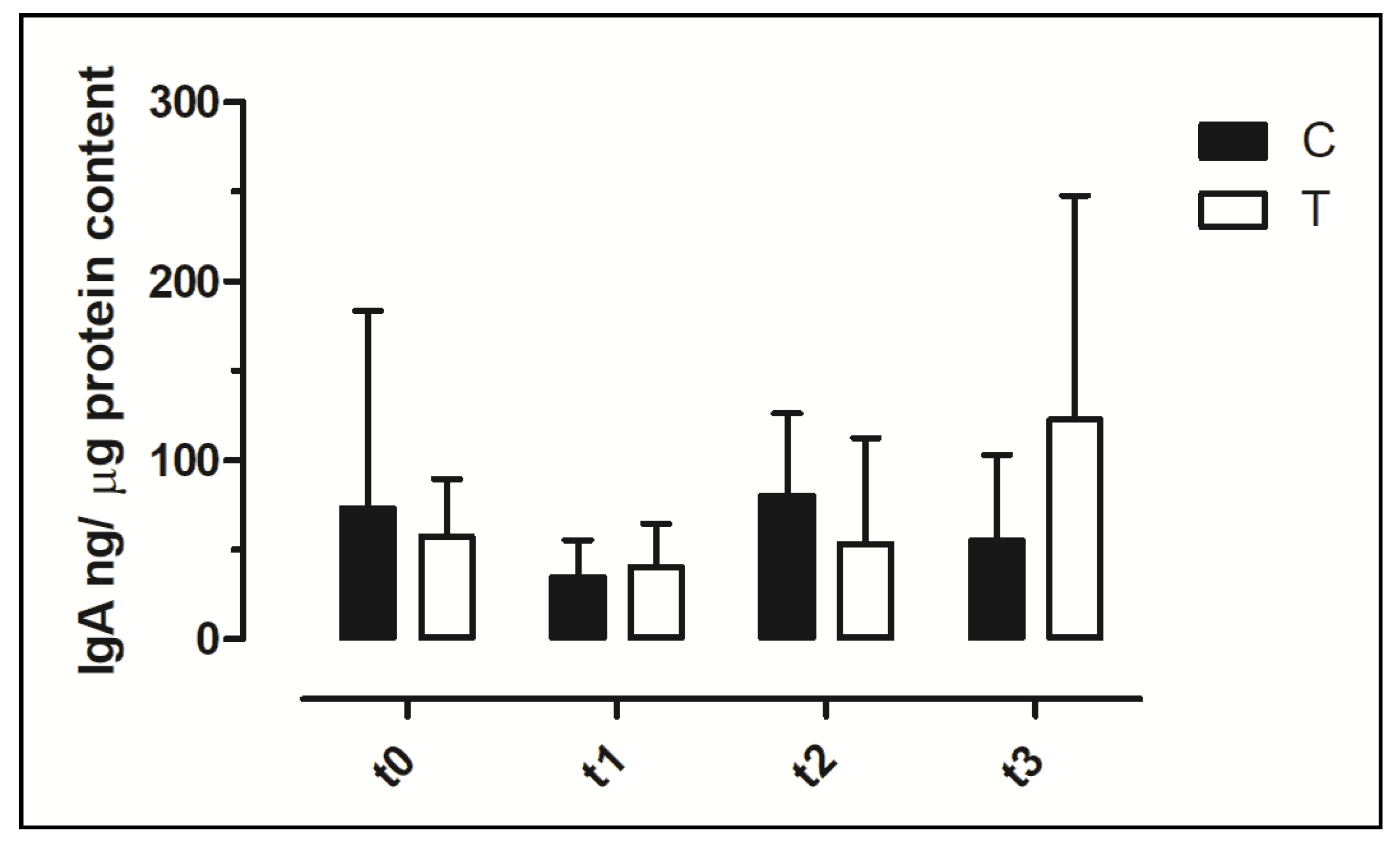
3.6. IgA Concentration in Faecal Swabs and Serum Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trovato, G.M. Behavior, nutrition and lifestyle in a comprehensive health and disease paradigm: Skills and knowledge for a predictive, preventive and personalized medicine. EPMA J. 2012, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, C.; Acuti, G.; Manuali, E.; Proietti, P.C.; Pavone, S.; Trabalza-Marinucci, M.; Moscati, L.; Onofri, A.; Lorenzetti, C.; Franciosini, M.P. Effects of two different probiotics on microflora, morphology, and morphometry of gut in organic laying hens. Poult. Sci. 2016, 95, 2528–2535. [Google Scholar] [CrossRef] [PubMed]
- Dibner, J.J.; Richards, J.D. Antibiotic growth promoters in agriculture: History and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Castanon, J.I.R. History of the Use of Antibiotic as Growth Promoters in European Poultry Feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef] [PubMed]
- Al-Yasiry, A.R.M.; Kiczorowska, B.; Samolińska, W.; Kowalczuk-Vasilev, E.; Kowalczyk-Pecka, D. The effect of Boswellia serrata resin diet supplementation on production, hematological, biochemical and immunological parameters in broiler chickens. Animals 2017, 11, 1890–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council of the European Union; European Parliament. Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition (Text with EEA Relevance), CELEX1. Available online: http://op.europa.eu/en/publication-detail/-/publication/c746ceb6-1330-4b77-9a2e-5a7705b22c33 (accessed on 19 January 2022).
- Ammon, H.P.T. Modulation of the immune system by Boswellia serrata extracts and boswellic acids. Phytomedicine 2010, 17, 862–867. [Google Scholar] [CrossRef]
- Azemi, M.E.; Namjoyan, F.; Khodayar, M.J.; Ahmadpour, F.; Padok, A.D.; Panahi, M. The Antioxidant Capacity and Anti-diabetic Effect of Boswellia serrata Triana and Planch Aqueous Extract in Fertile Female Diabetic Rats and the Possible Effects on Reproduction and Histological Changes in the Liver and Kidneys. Jundishapur J. Nat. Pharm. Prod. 2012, 7, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, R.M.; Fillmann, H.S.; Martins, M.I.M.; Meurer, L.; Marroni, N.P. Boswellia serrata has Beneficial Anti-Inflammatory and Antioxidant Properties in a Model of Experimental Colitis. Phytother. Res. 2014, 28, 1392–1398. [Google Scholar] [CrossRef]
- Umar, S.; Umar, K.; Sarwar, A.H.M.G.; Khan, A.; Ahmad, N.; Ahmad, S.; Katiyar, C.K.; Husain, S.A.; Khan, H.A. Boswellia serrata extract attenuates inflammatory mediators and oxidative stress in collagen induced arthritis. Phytomedicine 2014, 21, 847–856. [Google Scholar] [CrossRef]
- Al-Yasiry, A.R.M.; Kiczorowska, B.; Samolińska, W. Effect of Boswellia serrata Resin Supplementation on Basic Chemical and Mineral Element Composition in the Muscles and Liver of Broiler Chickens. Biol. Trace Elem. Res. 2017, 179, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Beghelli, D.; Isani, G.; Roncada, P.; Andreani, G.; Bistoni, O.; Bertocchi, M.; Lupidi, G.; Alunno, A. Antioxidant andEx VivoImmune System Regulatory Properties of Boswellia serrata Extracts. Oxidative Med. Cell. Longev. 2017, 2017, 7468064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertocchi, M.; Isani, G.; Medici, F.; Andreani, G.; Usca, I.T.; Roncada, P.; Forni, M.; Bernardini, C. Anti-Inflammatory Activity of Boswellia serrata Extracts: An In Vitro Study on Porcine Aortic Endothelial Cells. Oxidative Med. Cell. Longev. 2018, 2018, 2504305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiczorowska, B.; Al-Yasiry, A.; Samolińska, W.; Marek, A.; Pyzik, E. The effect of dietary supplementation of the broiler chicken diet with Boswellia serrata resin on growth performance, digestibility, and gastrointestinal characteristics, morphology, and microbiota. Livest. Sci. 2016, 191, 117–124. [Google Scholar] [CrossRef]
- Tabatabaei, S.N. Effect of Olibanum (Boswellia thurifera) as a feed additive on performance, some blood biochemical and intestinal morphology in broiler chicks. Res. Opin. Anim. Vet. Sci. 2016, 6, 130–134. [Google Scholar] [CrossRef]
- Apperson, K.D.; Cherian, G. Effect of whole flax seed and carbohydrase enzymes on gastrointestinal morphology, muscle fatty acids, and production performance in broiler chickens. Poult. Sci. 2017, 96, 1228–1234. [Google Scholar] [CrossRef]
- Chowdhury, S.; Mandal, G.P.; Patra, A.K. Different essential oils in diets of chickens: 1. Growth performance, nutrient utilisation, nitrogen excretion, carcass traits and chemical composition of meat. Anim. Feed Sci. Technol. 2018, 236, 86–97. [Google Scholar] [CrossRef]
- Kiczorowska, B.; Samolińska, W.; Al-Yasiry, A.; Zając, M. Immunomodulant feed supplement Boswellia serrata to support broiler chickens’ health and dietary and technological meat quality. Poult. Sci. 2020, 99, 1052–1061. [Google Scholar] [CrossRef]
- Csonka, C.; Sárközy, M.; Pipicz, M.; Dux, L.; Csont, T. Modulation of Hypercholesterolemia-Induced Oxidative/Nitrative Stress in the Heart. Oxidative Med. Cell. Longev. 2016, 2016, 3863726. [Google Scholar] [CrossRef]
- Khayyal, M.T.; El-Ghazaly, M.A.; Abdallah, D.M.; Okpanyi, S.N.; Kelber, O.; Weiser, D. Mechanisms Involved in the Anti-inflammatory Effect of a Standardized Willow Bark Extract. Arzneimittelforschung 2005, 55, 677–687. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Sugier, D.; Dziki, D.; Sugier, P. Bioaccessibility In Vitro of Nutraceuticals from Bark of Selected Salix Species. Sci. World J. 2014, 2014, 782763. [Google Scholar] [CrossRef] [Green Version]
- Maistro, E.L.; Terrazzas, P.M.; Perazzo, F.F.; Gaivao, I.O.D.M.; Sawaya, A.C.H.F.; Rosa, P.C.P. Salix alba (white willow) medicinal plant presents genotoxic effects in human cultured leukocytes. J. Toxicol. Environ. Health Part A 2019, 82, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Shara, M.; Stohs, S.J. Efficacy and Safety of White Willow Bark (Salix alba) Extracts. Phytother. Res. 2015, 29, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Karimi, I.; Modaresi, M.; Cheshmekaboodi, F.; Miraghaee, S.S. The Effects of Aromatic Water of Salix aegyptiaca L. and its Major Component, 1,4-Dimethoxybenzene, on Lipid and Lipoprotein Profiles and Ethology of Normolipidemic Rabbits. Int. J. Clin. Toxicol. 2015, 2, 55–63. [Google Scholar] [CrossRef]
- Zabihi, N.A.; Mahmoudabady, M.; Soukhtanloo, M.; Hayatdavoudi, P.; Beheshti, F.; Niazmand, S. Salix alba attenuated oxidative stress in the heart and kidney of hypercholesterolemic rabbits. Avicenna J. Phytomed 2018, 8, 63–72. [Google Scholar]
- Poźniak, B.; Świtała, M.; Bobrek, K.; Graczyk, S.; Dzimira, S. Adverse effects associated with high-dose acetylsalicylic acid and sodium salicylate treatment in broilers. Br. Poult. Sci. 2012, 53, 777–783. [Google Scholar] [CrossRef]
- Seliger, C.; Schaerer, B.; Kohn, M.; Pendl, H.; Weigend, S.; Kaspers, B.; Härtle, S. A rapid high-precision flow cytometry based technique for total white blood cell counting in chickens. Vet. Immunol. Immunopathol. 2012, 145, 86–99. [Google Scholar] [CrossRef]
- Naghizadeh, M.; Larsen, F.T.; Wattrang, E.; Norup, L.R.; Dalgaard, T.S. Rapid whole blood assay using flow cytometry for measuring phagocytic activity of chicken leukocytes. Vet. Immunol. Immunopathol. 2019, 207, 53–61. [Google Scholar] [CrossRef]
- Merino-Guzmán, R.; Latorre, J.D.; Delgado, R.; Hernandez-Velasco, X.; Wolfenden, A.D.; Teague, K.D.; Graham, L.E.; Mahaffey, B.D.; Baxter, M.F.A.; Hargis, B.M.; et al. Comparison of total immunoglobulin A levels in different samples in Leghorn and broiler chickens. Asian Pac. J. Trop. Biomed. 2017, 7, 116–120. [Google Scholar] [CrossRef]
- Board, M.M.; Crespo, R.; Shah, D.H.; Faux, C.M. Biochemical Reference Intervals for Backyard Hens. J. Avian Med. Surg. 2018, 32, 301–306. [Google Scholar] [CrossRef]
- Tang, S.G.H.; Sieo, C.C.; Ramasamy, K.; Saad, W.Z.; Wong, H.K.; Ho, Y.W. Performance, biochemical and haematological responses, and relative organ weights of laying hens fed diets supplemented with prebiotic, probiotic and synbiotic. BMC Vet. Res. 2017, 13, 248. [Google Scholar] [CrossRef] [Green Version]
- Tamminen, L.-M.; Emanuelson, U.; Blanco-Penedo, I. Systematic Review of Phytotherapeutic Treatments for Different Farm Animals Under European Conditions. Front. Vet. Sci. 2018, 5, 140. [Google Scholar] [CrossRef] [PubMed]
- Al-Yasiry, A.R.M.; Kiczorowska, B.; Samolińska, W. Nutritional value and content of mineral elements in the meat of broiler chickens fed Boswellia serrata supplemented diets. J. Elem. 2017, 22, 1027–1037. [Google Scholar] [CrossRef]
- Khan, A.; Al-Harrasi, A.; Rehman, N.U.; Sarwar, R.; Ahmad, T.; Ghaffar, R.; Khan, H.; Al-Amri, I.; Csuk, R.; Al-Rawahi, A. Loading AKBA on surface of silver nanoparticles to improve their sedative-hypnotic and anti-inflammatory efficacies. Nanomedicine 2019, 14, 2783–2798. [Google Scholar] [CrossRef] [PubMed]
- Moussaieff, A.; Rimmerman, N.; Bregman, T.; Straiker, A.; Felder, C.C.; Shoham, S.; Kashman, Y.; Huang, S.M.; Lee, H.; Shohami, E.; et al. Incensole acetate, an incense component, elicits psychoactivity by activating TRPV3 channels in the brain. FASEB J. 2008, 22, 3024–3034. [Google Scholar] [CrossRef] [PubMed]
- Okano, S.; Honda, Y.; Kodama, T.; Kimura, M. The Effects of Frankincense Essential Oil on Stress in Rats. J. Oleo Sci. 2019, 68, 1003–1009. [Google Scholar] [CrossRef] [Green Version]
- Tully, T.N.; Dorrestein, G.M.; Jones, A.K.; Cooper, E.J. Handbook of Avian Medicine—University of Missouri Libraries. Available online: http://link.library.missouri.edu/portal/Handbook-of-avian-medicine-edited-by-Thomas-N./c65YkONeUPE/ (accessed on 7 March 2022).
- Ding, H.; Yue, Q.; Chang, L.; Xi, J.; Chen, H.; Li, F.; Wang, D.; Zhou, R. Whole blood gas and biochemical reference intervals for Lohmann Silver layers. Poult. Sci. 2021, 100, 101368. [Google Scholar] [CrossRef]
- Rafiullah; Khan, S.; Khan, R.U.; Ullah, Q. Does the gradual replacement of spent silkworm (Bombyx mori) pupae affect the performance, blood metabolites and gut functions in White Leghorn laying hens? Res. Vet. Sci. 2020, 132, 574–577. [Google Scholar] [CrossRef]
- Borzouie, S.; Rathgeber, B.M.; Stupart, C.M.; MacIsaac, J.; MacLaren, L.A. Effects of Dietary Inclusion of Seaweed, Heat Stress and Genetic Strain on Performance, Plasma Biochemical and Hematological Parameters in Laying Hens. Animals 2020, 10, 1570. [Google Scholar] [CrossRef]
- Ismail, I.E.; Abdelnour, S.A.; Shehata, S.A.; El-Hack, M.E.A.; El-Edel, M.A.; Taha, A.E.; Schiavitto, M.; Tufarelli, V. Effect of Dietary Boswellia serrata Resin on Growth Performance, Blood Biochemistry, and Cecal Microbiota of Growing Rabbits. Front. Vet. Sci. 2019, 6, 471. [Google Scholar] [CrossRef] [Green Version]
- Panaite, T.D.; Saracila, M.; Papuc, C.P.; Predescu, C.N.; Soica, C. Influence of Dietary Supplementation of Salix alba Bark on Performance, Oxidative Stress Parameters in Liver and Gut Microflora of Broilers. Animals 2020, 10, 958. [Google Scholar] [CrossRef]
- Davis, A.K. Effect of handling time and repeated sampling on avian white blood cell counts (wfecto del tiempo de manipulación en muestras repetidas en el conteo de celulas blancas en aves). J. Field Ornithol. 2005, 76, 334–338. [Google Scholar] [CrossRef] [Green Version]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef] [PubMed]
- Gross, W.B.; Siegel, H.S. Evaluation of the Heterophil/Lymphocyte Ratio as a Measure of Stress in Chickens. Avian Dis. 1983, 27, 972. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.W. Avian Hematology and Cytology, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1995; ISBN 978-0-8138-2970-8. [Google Scholar]
- Maxwell, M.H.; Robertson, G.W. The avian heterophil leucocyte: A review. World’s Poult. Sci. J. 1998, 54, 155–178. [Google Scholar] [CrossRef]
- Lentfer, T.L.; Pendl, H.; Gebhardt, S.; Fröhlich, E.K.F.; Von Borell, E. H/L ratio as a measurement of stress in laying hens - methodology and reliability. Br. Poult. Sci. 2015, 56, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Davison, F.; Kaspers, B.; Karel, A. Schat Avian Immunology, Free Online Publishing. Available online: https://authorzilla.com/lx8zn/avian-immunology.html (accessed on 26 January 2022).
- Jugan, M.C.; Rudinsky, A.J.; Parker, V.J.; Gilor, C. Use of probiotics in small animal veterinary medicine. J. Am. Vet. Med. Assoc. 2017, 250, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Bütz, D.E.; Whelan, R.; Naranjo, V.; Arendt, M.K.; Ramuta, M.D.; Yang, X.; Crenshaw, T.D.; Cook, M.E. Effects of dietary methionine plus cysteine levels on growth performance and intestinal antibody production in broilers during Eimeria challenge. Poult. Sci. 2020, 99, 374–384. [Google Scholar] [CrossRef]
- Bi, S.; Zhang, J.; Qu, Y.; Zhou, B.; He, X.; Ni, J. Yeast cell wall product enhanced intestinal IgA response and changed cecum microflora species after oral vaccination in chickens. Poult. Sci. 2020, 99, 6576–6585. [Google Scholar] [CrossRef]
- Ricci, F.G.; Terkelli, L.R.; Venancio, E.J.; Justino, L.; Dos Santos, B.Q.; Baptista, A.A.S.; Oba, A.; de Oliveira Souza, B.D.; Bracarense, A.P.F.R.L.; Hirooka, E.Y.; et al. Tryptophan Attenuates the Effects of OTA on Intestinal Morphology and Local IgA/IgY Production in Broiler Chicks. Toxins 2020, 13, 5. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Values of Nutrients (%/kg of Finisher Diet) | Additives (mg/kg; IU; OTU/kg) | Complementary Feed Composition (%) |
|---|---|---|---|
| Corn; Corn gluten flour; Soybeans meal (* CP 43%); Calcium carbonate; roasted Soybeans; Rice husk; Corn gluten; Wheat bran; Soybean oil | CP, 17%; ** CF, 5%; *** CF, 3.51%; ! Cash, 13.27%; ǂ Ca, 4.02%; ǂǂ P, 0.58%; ≠ NaCl, 0.15%; + Ly, 0.85%; $ Met: 0.33%. | Vitamin A, 9950 IU; Vitamin D3, 2701 IU; Vitamin E, 38 mg; Vitamin K3, 2 mg; Vitamin B1, 1.5 mg; Vitamin B2, 4.5 mg; Vitamin B6, 2.5 mg; Vitamin B12, 0.008 mg; niacin, 35 mg; Ca-D-pantotenate, 10 mg; folic acid, 1 mg; biotin, 0.1 mg; betaine hydrochloride, 250 mg; Cu, 5 mg; anhydrous calcium iodate, 0.50 mg; Mn, 50 mg; Se, 0.075 mg; Zn, 40 mg; Cantaxantine, 2 mg; promoters of digestion 6-phytase, 213 OTU; DL-Methionine, 627 mg | Calcium carbonate, 74.5%; Colloidal silica, 15%; Salix & DE, 5%; Boswellia & DE, 5%; Sodium chloride, 0.35%; Magnesium carbonate, 0.15% |
| Parameters | T | C | p-Value (KWt) |
|---|---|---|---|
| Mean ± DS | Mean ± DS | ||
| Kg/BW 1°P (1–7 week) | 1.0 ± 0.1 | 1.1 ± 0.1 | p > 0.749 |
| Kg/BW 2°P (8–12 week) | 1.5 ± 0.1 | 1.5 ± 0.1 | p > 0.601 |
| Kg/BW 3°P (13–19 week) ” | 1.7 ± 0.0 | 1.8 ± 0.0 | * p < 0.004 |
| FI 1°P (1–7 week) ** | 71.9 ± 18.5 | 70.2 ± 17.6 | p > 0.100 |
| FI 2°P (8–12 week) | 115.8 ± 16.8 | 109.5 ± 16.9 | p > 0.077 |
| FI 3°P (13–19 week) | 139.8 ± 8.3 | 139.4 ± 8.9 | p > 0.970 |
| FI (1–19 week) | 108.3 ± 33.2 | 105.3 ± 33.4 | p > 0.516 |
| WI 1°P (1–7 week) *** | 140.1 ± 35.5 | 125.6 ± 38.2 | * p < 0.013 |
| WI 2°P (8–12 week) | 208.5 ± 46.3 | 163.0 ± 46.6 | * p < 0.000 |
| WI 3°P (13–19 week) | 251.4 ± 32.0 | 251.2 ± 34.1 | p > 0.929 |
| WI (1–19 week) | 198.7 ± 61.2 | 181.2 ± 67.7 | * p < 0.013 |
| FCR (1–7 week) ! | 1.7 ± 1.0 | 1.2 ± 0.4 | p > 0.609 |
| Feed Efficiency (8–19 week) £ | 2.8 ± 0.7 | 3.3 ± 1.7 | p > 0.508 |
| Egg mass (2–12 week/laying) # | 50.1 ± 10.7 | 46.3 ± 15.0 | p > 0.921 |
| n° eggs/day/lay (1–12 week/laying) | 46.6 ± 17.9 | 44.4 ± 18.0 | p > 0.246 |
| n° total egg/group (1–12 week/laying) | 3924 | 3559 | p > 0.068 |
| Mean DR% (2–12 week/laying) $ | 83.4 ± 26.6 | 75.3 ± 31.8 | p > 0.418 |
| Egg weigth (n = 20/group) (3–12 week/laying) & | 59.9 ± 2.5 | 61.0 ± 3.4 | * p < 0.000 |
| Serum Analytes | t0 | t1 | t2 | t3 | From Reference § | ||||
|---|---|---|---|---|---|---|---|---|---|
| C | T | C | T | C | T | C | T | ||
| AST (U/L) | 270 ± 40.0 | 243 ± 21.94 | 216 ± 20.4 | 207 ± 12.5 | 221 ± 25.8 | 207 ± 17.7 | 214 ± 16.3 | 217 ± 25.1 | 118–298 [30] |
| ALT (U/L) | 1.25 ± 0.46 | 1.71 ± 0.76 | 1.88 ± 0.64 | 2.50 ± 0.93 | 1.57 ± 0.53 | 1.38 ± 0.52 | 1.63 ± 0.74 | 2.88 ± 1.25 | 25.2 ± 7.77 [31] |
| ALP (U/L) | 1159 ± 310 | 1510 ± 367 | 713 ± 182 | 673 ± 98.5 | 730 ± 167 | 595 ± 209 | 480 ± 206 | 552 ± 236 | 407 ± 39.84 [31] |
| Bilirubin (μmol/L) | 0.17 ± 0.00 | 0.34 ± 0.17 | 0.88 ± 0.68 | 0.34 ± 0.17 | 0.34 ± 0.32 | 0.34 ± 0.17 | 0.51 ± 0.34 | 0.51 ± 0.51 | -- |
| Cholesterol (mmol/L) | 3.42 ± 0.17 | 3.16 ± 0.35 | 2.98 ± 0.17 | 2.92 ± 0.31 | 3.19 ± 0.55 | 3.00 ± 0.71 | 2.98 ± 0.74 | 3.32 ± 0.62 | 3.37 ± 0.35 [31] |
| Triglycerides (g/L) | 1.05 ± 0.49 | 0.90 ± 0.21 | 1.29 ± 0.19 | 1.15 ± 0.10 | 12.6 ± 3.38 | 12.6 ± 4.63 | 12.29 ± 5.94 | 17.36 ± 3.46 | 13.6 ± 3.56 [31] |
| Glucose (mmol/L) | 12.9 ± 1.23 * | 14.3 ± 0.74 * | 8.63 ± 0.79 | 9.35 ± 1.21 | 10.78 ± 0.49 | 11.2 ± 1.32 | 7.20 ± 1.72 | 7.70 ± 1.96 | 9.41–13.6 [30] |
| Total proteins (g/L) | 42.7 ± 3.13 | 39.3 ± 2.92 | 48.4 ± 3.62 | 46.2 ± 4.11 | 48.7 ± 4.85 | 46.8 ± 4.29 | 45.9 ± 3.40 | 48.3 ± 4.09 | 39–70 [30] |
| Albumin (g/L) | 16.3 ± 1.03 * | 15.0 ± 0.80 * | 19.2 ± 1.19 | 18.4 ± 1.46 | 18.7 ± 1.10 * | 17.5 ± 0.75 * | 18.3 ± 0.82 | 18.8 ± 0.84 | 15.0–33.0 [30] |
| Globulins (g/L) | 25.8 ± 1.85 | 26.2 ± 4.92 | 29.2 ± 2.58 | 27.8 ± 3.17 | 30.0 ± 4.01 | 29.2 ± 4.15 | 27.7 ± 2.70 | 29.5 ± 3.34 | 16.0–43.0 [30] |
| Albumin/ Globulins | 0.65 ± 0.06 | 0.61 ± 0.05 | 0.66 ± 0.03 | 0.67 ± 0.07 | 0.63 ± 0.06 | 0.61 ± 0.07 | 0.67 ± 0.05 | 0.64 ± 0.05 | -- |
| P (mmol/L) | 1.78 ± 0.32 | 1.53 ± 0.19 | 2.01 ± 0.18 | 1.92 ± 0.23 | 2.01 ± 0.17 | 2.07 ± 0.21 | 2.17 ± 0.37 | 2.33± 0.42 | 0.52- 2.33 [30] |
| Ca (mmol/L) | 2.65 ± 0.34 | 2.98 ± 0.20 | 2.70 ± 0.21 | 2.46 ± 0.43 | 6.10 ± 0.41 | 6.40 ± 0.63 | 6.75 ± 0.10 | 7.25 ± 0.64 | > 2.73 [30] |
| Uric acid (mmol/L) | 0.30 ± 0.06 | 0.28 ± 0.06 | 0.42 ± 0.08 | 0.39 ± 0.08 | 0.37 ± 0.11 | 0.33 ± 0.09 | 0.39 ± 0.12 | 0.30 ± 0.12 | 0.05–0.53 [30] |
| IgA (g/L) | 0.72 ± 0.22 | 0.83 ± 0.09 | 0.70 ± 0.01 | 0.71 ± 0.08 | 0.70 ± 0.00 | 0.65 ± 0.03 | 0.63 ± 0.00 | 0.51 ± 0.25 | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrini, A.; Dalmonte, T.; Lupini, C.; Andreani, G.; Salaroli, R.; Quaglia, G.; Zannoni, A.; Scozzoli, M.; Forni, M.; Isani, G. Influence of Dietary Supplementation with Boswellia serrata and Salix alba on Performance and Blood Biochemistry in Free-Range Leghorn Laying Hens. Vet. Sci. 2022, 9, 182. https://doi.org/10.3390/vetsci9040182
Guerrini A, Dalmonte T, Lupini C, Andreani G, Salaroli R, Quaglia G, Zannoni A, Scozzoli M, Forni M, Isani G. Influence of Dietary Supplementation with Boswellia serrata and Salix alba on Performance and Blood Biochemistry in Free-Range Leghorn Laying Hens. Veterinary Sciences. 2022; 9(4):182. https://doi.org/10.3390/vetsci9040182
Chicago/Turabian StyleGuerrini, Alessandro, Thomas Dalmonte, Caterina Lupini, Giulia Andreani, Roberta Salaroli, Giulia Quaglia, Augusta Zannoni, Maurizio Scozzoli, Monica Forni, and Gloria Isani. 2022. "Influence of Dietary Supplementation with Boswellia serrata and Salix alba on Performance and Blood Biochemistry in Free-Range Leghorn Laying Hens" Veterinary Sciences 9, no. 4: 182. https://doi.org/10.3390/vetsci9040182
APA StyleGuerrini, A., Dalmonte, T., Lupini, C., Andreani, G., Salaroli, R., Quaglia, G., Zannoni, A., Scozzoli, M., Forni, M., & Isani, G. (2022). Influence of Dietary Supplementation with Boswellia serrata and Salix alba on Performance and Blood Biochemistry in Free-Range Leghorn Laying Hens. Veterinary Sciences, 9(4), 182. https://doi.org/10.3390/vetsci9040182