
First Report of Isolation and Molecular Characterization of Felid Herpesvirus-1 from Symptomatic Domestic Cats in Egypt



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Considerations
2.2. Sampling
2.3. Virus Isolation
2.4. DNA Extraction and PCR Amplification of Thymidine Kinase (TK) Gene
2.5. DNA Sequencing and Phylogenetic Analysis
3. Results
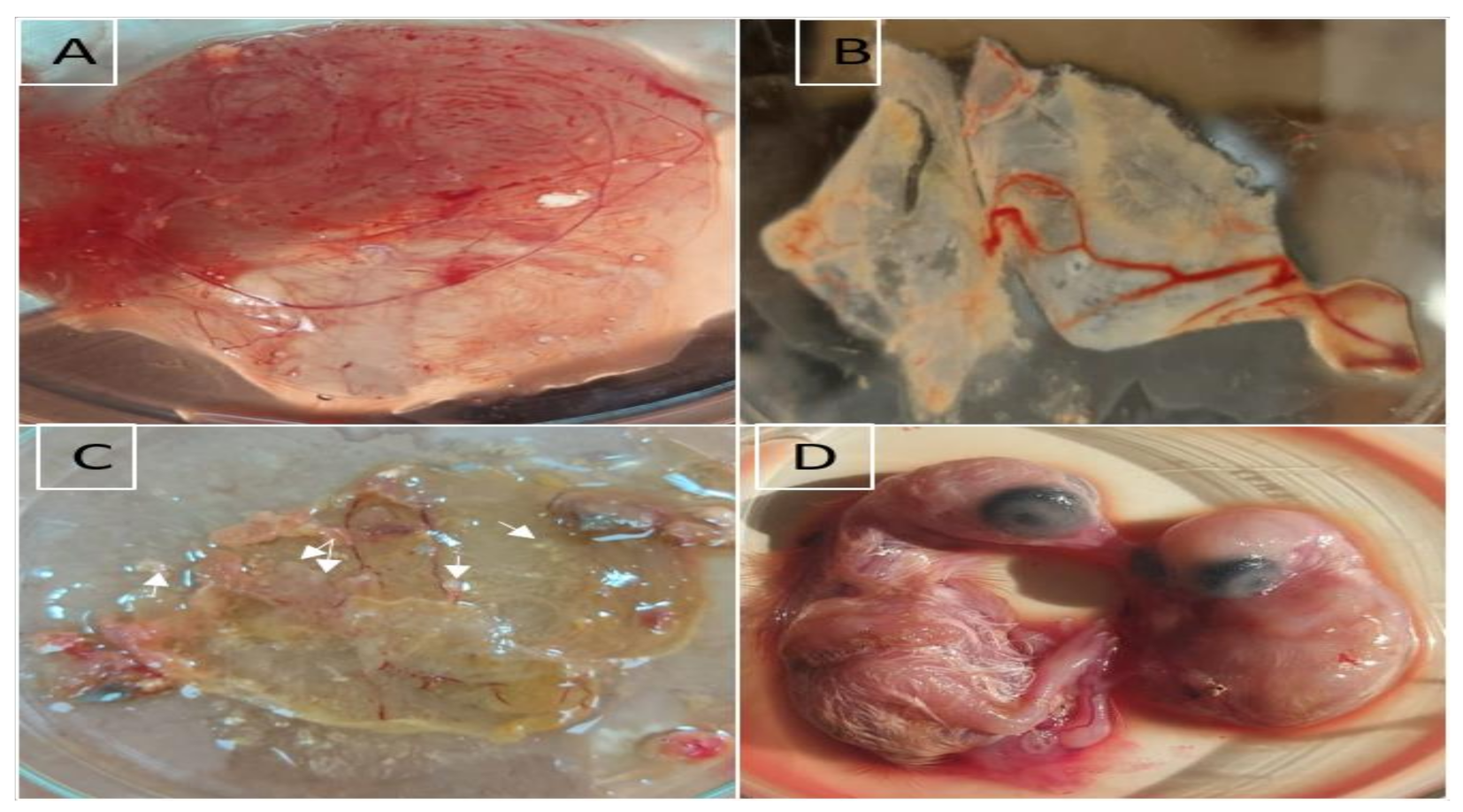
3.1. Virus Isolation
3.2. Molecular Identification by PCR
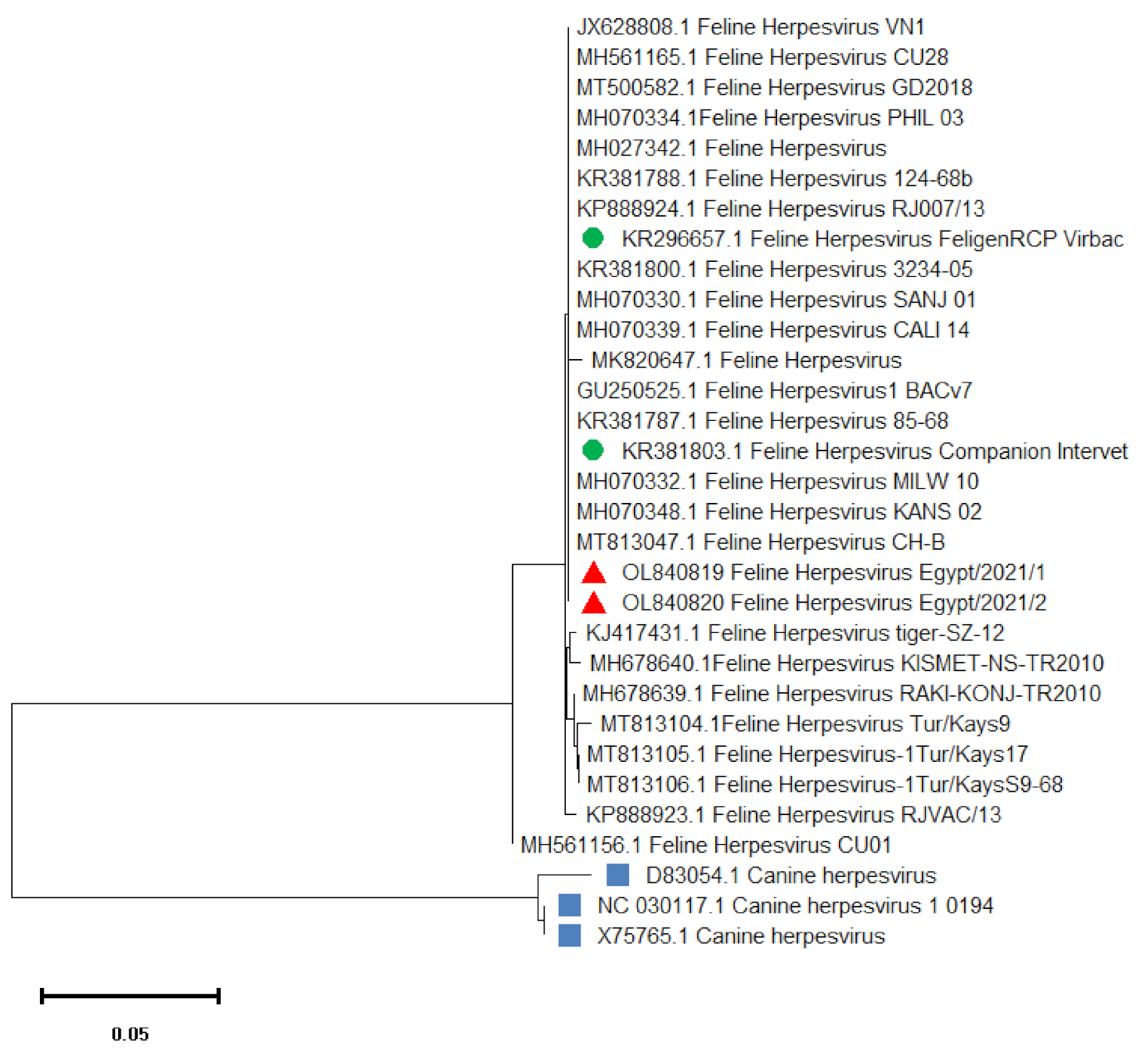
3.3. Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Macías, P.; Navarro, C. Molecular detection of feline herpesvirus by means polymerase chain reaction. Dairy Vet. Sci. J. 2018, 8, 555736. [Google Scholar]
- Maeda, K.; Horimoto, T.; Mikami, T. Properties and functions of feline herpesvirus type 1 glycoproteins. J. Vet. Med. Sci. 1998, 60, 881–888. [Google Scholar] [CrossRef] [Green Version]
- Maggs, D.J.; Clarke, H.E. Relative sensitivity of polymerase chain reaction assays used for detection of feline herpesvirus type 1 DNA in clinical samples and commercial vaccines. Am. J. Vet. Res. 2005, 66, 1550–1555. [Google Scholar] [CrossRef] [PubMed]
- Monne Rodriguez, J.M.; Leeming, G.; Köhler, K.; Kipar, A. Feline herpesvirus pneumonia: Investigations into the pathogenesis. Vet. Pathol. 2017, 54, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Bol, S.; Bunnik, E.M. Lysine supplementation is not effective for the prevention or treatment of feline herpesvirus 1 infection in cats: A systematic review. BMC Vet. Res. 2015, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gould, D. Feline herpesvirus-1: Ocular manifestations, diagnosis and treatment options. J. Feline Med. Surg. 2011, 13, 333–346. [Google Scholar] [CrossRef]
- Thiry, E.; Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A. Feline herpesvirus infection. ABCD guidelines on prevention and management. J. Feline Med. Surg. 2009, 11, 547–555. [Google Scholar] [CrossRef]
- Helps, C.; Reeves, N.; Egan, K.; Howard, P.; Harbour, D. Detection of Chlamydophila Felis and feline herpesvirus by multiplex real-time PCR analysis. J. Clin. Microbiol. 2003, 41, 2734–2736. [Google Scholar] [CrossRef] [Green Version]
- Lewin, A.C.; Kolb, A.W.; McLellan, G.J.; Bentley, E.; Bernard, K.A.; Newbury, S.P.; Brandt, C.R. Genomic, recombinational and phylogenetic characterization of global feline herpesvirus 1 isolates. Virology 2018, 518, 385–397. [Google Scholar] [CrossRef]
- Holst, B.S.; Berndtsson, L.T.; Englund, L. Isolation of feline herpesvirus-1 and feline calicivirus from healthy cats in Swedish breeding catteries. J. Feline Med. Surg. 2005, 7, 325–331. [Google Scholar] [CrossRef]
- Chvala-Mannsberger, S.; Bago, Z.; Weissenböck, H. Occurrence, morphological characterization and antigen localization of felid herpesvirus-induced pneumonia in cats: A retrospective study (2000–2006). J. Comp. Pathol. 2009, 141, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Zicola, A.; Saegerman, C.; Quatpers, D.; Viandier, J.; Thiry, E. Feline herpesvirus 1 and feline calicivirus infections in a heterogeneous cat population of a rescue shelter. J. Feline Med. Surg. 2009, 11, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.S.; Monteiro, F.L.; Scariot, A.C.A.; Chagas, D.B.; Fischer, G.; Lima, M.d.; Hübner, S.O. High occurrence of felid alphaherpesvirus 1 and feline calicivirus in domestic cats from southern Brazil. Pesquisa Veterinária Brasileira 2020, 40, 685–689. [Google Scholar] [CrossRef]
- Legendre, A.M.; Kuritz, T.; Heidel, R.E.; Baylor, V.M. Polyprenyl immunostimulant in Feline rhinotracheitis: Randomized placebo-controlled experimental and field safety studies. Front. Vet. Sci. 2017, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Vogtlin, A.; Fraefel, C.; Albini, S.; Leutenegger, C.; Schraner, E.; Spiess, B.; Lutz, H.; Ackermann, M. Quantification of feline herpesvirus 1 DNA in ocular fluid samples of clinically diseased cats by real-time TaqMan PCR. J. Clin. Microbiol. 2002, 40, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Maes, R. Felid herpesvirus type 1 infection in cats: A natural host model for alphaherpesvirus pathogenesis. ISRN Vet. Sci. 2012, 2012, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gaskell, R.; Dawson, S.; Radford, A.; Thiry, E. Feline herpesvirus. Vet. Res. 2007, 38, 337–354. [Google Scholar] [CrossRef] [Green Version]
- Tai, S.S.; Niikura, M.; Cheng, H.H.; Kruger, J.M.; Wise, A.G.; Maes, R.K. Complete genomic sequence and an infectious BAC clone of feline herpesvirus-1 (FHV-1). Virology 2010, 401, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.-K.; Kim, H.-H.; Park, Y.-R.; Yoo, J.Y.; Choi, S.-S.; Park, Y.; An, S.; Park, J.; Kim, J.; Kim, H.-J. Isolation and molecular characterization of feline herpesvirus 1 from naturally infected Korean cats. J. Bacteriol. Virol. 2020, 50, 263–272. [Google Scholar] [CrossRef]
- Henzel, A.; Brum, M.C.S.; Lautert, C.; Martins, M.; Lovato, L.T.; Weiblen, R. Isolation and identification of feline calicivirus and feline herpesvirus in Southern Brazil. Braz. J. Microbiol. 2012, 43, 560–568. [Google Scholar] [CrossRef] [Green Version]
- Nunberg, J.H.; Wright, D.; Cole, G.; Petrovskis, E.; Post, L.; Compton, T.; Gilbert, J. Identification of the thymidine kinase gene of feline herpesvirus: Use of degenerate oligonucleotides in the polymerase chain reaction to isolate herpesvirus gene homologs. J. Virol. 1989, 63, 3240–3249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townsend, W.M.; Jacobi, S.; Tai, S.-H.; Kiupel, M.; Wise, A.G.; Maes, R.K. Ocular and neural distribution of feline herpesvirus-1 during active and latent experimental infection in cats. BMC Vet. Res. 2013, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Maes, R.K.; Kruger, J.M.; Kiupel, M.; Giessler, K.S.; Soboll Hussey, G. Safety and efficacy of felid herpesvirus-1 deletion mutants in cats. Viruses 2021, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Vaz, P.K.; Job, N.; Horsington, J.; Ficorilli, N.; Studdert, M.J.; Hartley, C.A.; Gilkerson, J.R.; Browning, G.F.; Devlin, J.M. Low genetic diversity among historical and contemporary clinical isolates of felid herpesvirus. BMC Genom. 2016, 17, 704. [Google Scholar] [CrossRef] [Green Version]
- Sandmeyer, L.S.; Waldner, C.L.; Bauer, B.S.; Wen, X.; Bienzle, D. Comparison of polymerase chain reaction tests for diagnosis of feline herpesvirus, Chlamydophila Felis, and Mycoplasma spp. infection in cats with ocular disease in Canada. Can. Vet. J. 2010, 51, 629. [Google Scholar] [PubMed]
- Stiles, J. Feline herpesvirus. Clin. Tech. Small Anim. Pract. 2003, 18, 178–185. [Google Scholar] [CrossRef]
- Maggs, D.J. Update on pathogenesis, diagnosis, and treatment of feline herpesvirus type 1. Clin. Tech. Small Anim. Pract. 2005, 20, 94–101. [Google Scholar] [CrossRef]
- Burgesser, K.M.; Hotaling, S.; Schiebel, A.; Ashbaugh, S.E.; Roberts, S.M.; Collins, J.K. Comparison of PCR, virus isolation, and indirect fluorescent antibody staining in the detection of naturally occurring feline herpesvirus infections. J. Vet. Diagn. Investig. 1999, 11, 122–126. [Google Scholar] [CrossRef]
- Calnek, B.; Barnes, H.; Beard, C.; Yoder, H. Diseases of Poultry; Iowa State University Press: Ames, IA, USA, 1997. [Google Scholar]
- Sun, H.; Li, Y.; Jiao, W.; Liu, C.; Liu, X.; Wang, H.; Hua, F.; Dong, J.; Fan, S.; Yu, Z. Isolation and identification of feline herpesvirus type 1 from a South China tiger in China. Viruses 2014, 6, 1004–1014. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Grahn, B.H.; Sisler, S.; Storey, E. Qualitative tear film and conjunctival goblet cell assessment of cats with corneal sequestra. Vet. Ophthalmol. 2005, 8, 167–170. [Google Scholar] [CrossRef]
- Hara, M.; Fukuyama, M.; Suzuki, Y.; Kisikawa, S.; Ikeda, T.; Kiuchi, A.; Tabuchi, K. Detection of feline herpesvirus 1 DNA by the nested polymerase chain reaction. Vet. Microbiol. 1996, 48, 345–352. [Google Scholar] [CrossRef]
- Maggs, D.J.; Lappin, M.R.; Reif, J.S.; Collins, J.K.; Carman, J.; Dawson, D.A.; Bruns, C. Evaluation of serologic and viral detection methods for diagnosing feline herpesvirus-1 infection in cats with acute respiratory tract or chronic ocular disease. J. Am. Vet. Med. Assoc. 1999, 214, 502–507. [Google Scholar]
- Sykes, J.E.; Allen, J.L.; Studdert, V.P.; Browning, G.F. Detection of feline calicivirus, feline herpesvirus 1 and Chlamydia psittaci mucosal swabs by multiplex RT-PCR/PCR. Vet. Microbiol. 2001, 81, 95–108. [Google Scholar] [CrossRef]
- Fernandez, M.; Manzanilla, E.G.; Lloret, A.; León, M.; Thibault, J.-C. Prevalence of feline herpesvirus-1, feline calicivirus, Chlamydophila Felis and Mycoplasma Felis DNA and associated risk factors in cats in Spain with upper respiratory tract disease, conjunctivitis and/or gingivostomatitis. J. Feline Med. Surg. 2017, 19, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Low, H.C.; Powell, C.C.; Veir, J.K.; Hawley, J.R.; Lappin, M.R. Prevalence of feline herpesvirus 1, Chlamydophila Felis, and Mycoplasma spp DNA in conjunctival cells collected from cats with and without conjunctivitis. Am. J. Vet. Res. 2007, 68, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.-T.; Park, H.-M. Prevalence of feline herpesvirus 1, feline calicivirus and Chlamydophila Felis in clinically normal cats at a Korean animal shelter. J. Vet. Sci. 2008, 9, 207–209. [Google Scholar] [CrossRef]
- Wieliczko, A.; Płoneczka-Janeczko, K. Feline herpesvirus 1 and Chlamydophila Felis prevalence in cats with chronic conjunctivitis. Pol. J. Vet. Sci. 2010, 13, 381–383. [Google Scholar]
- Sjödahl-Essén, T.; Tidholm, A.; Thorén, P.; Persson-Wadman, A.; Bölske, G.; Aspán, A.; Treiberg Berndtsson, L. Evaluation of different sampling methods and results of real-time PCR for detection of feline herpes virus-1, Chlamydophila Felis and Mycoplasma Felis in cats. Vet. Ophthalmol. 2008, 11, 375–380. [Google Scholar] [CrossRef]
- Sykes, J.E.; Anderson, G.A.; Studdert, V.P.; Browning, G.F. Prevalence of feline Chlamydia psittaci and feline herpesvirus 1 in cats with upper respiratory tract disease. J. Vet. Intern. Med. 1999, 13, 153–162. [Google Scholar] [CrossRef]
- Zirofsky, D.; Rekers, W.; Powell, C.; Hawley, J.; Veir, J.; Lappin, M. Feline herpesvirus 1 and Mycoplasma spp. conventional PCR assay results from conjunctival samples from cats in shelters with suspected acute ocular infections. Top. Companion Anim. Med. 2018, 33, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Stiles, J.; McDermott, M.; Willis, M.; Roberts, W.; Greene, C. Comparison of nested polymerase chain reaction, virus isolation, and fluorescent antibody testing for identifying feline herpesvirus in cats with conjunctivitis. Am. J. Vet. Res. 1997, 58, 804–807. [Google Scholar] [PubMed]
- Westermeyer, H.D.; Kado-Fong, H.; Maggs, D.J. Effects of sampling instrument and processing technique on DNA yield and detection rate for feline herpesvirus-1 via polymerase chain reaction assay. Am. J. Vet. Res. 2008, 69, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Volopich, S.; Benetka, V.; Schwendenwein, I.; Möstl, K. Sommerfeldinstrument and processing technique on DNA yield and detectrus DNA and Chlamydophila Felis antigen detection rates in normal cats and cats with conjunctival and corneal lesions. Vet. Ophthalmol. 2005, 8, 25–32. [Google Scholar] [CrossRef]
- Horimoto, T.; Limcumpao, J.; Xuan, X.; Ono, M.; Maeda, K.; Kawaguchi, Y.; Kai, C.; Takahashi, E.; Mikami, T. Heterogeneity of feline herpesvirus type 1 strains. Arch. Virol. 1992, 126, 283–292. [Google Scholar] [CrossRef]
- Hamano, M.; Maeda, K.; Mizukoshi, F.; Mochizuki, M.; Tohya, Y.; Akashi, H.; Kai, K. Genetic rearrangements in the gC gene of the feline herpesvirus type 1. Virus Genes 2004, 28, 55–60. [Google Scholar] [CrossRef]
- Hamano, M.; Maeda, K.; Kai, K.; Mochizuki, M.; Tohya, Y.; Akashi, H. A novel genetic marker to differentiate feline herpesvirus type 1 field isolates. Vet. Microbiol. 2005, 106, 195–200. [Google Scholar] [CrossRef]
- Leeming, G.; Meli, M.L.; Cripps, P.; Vaughan-Thomas, A.; Lutz, H.; Gaskell, R.; Kipar, A. Tracheal organ cultures as a useful tool to study Felid herpesvirus 1 infection in respiratory epithelium. J. Virol. Methods 2006, 138, 191–195. [Google Scholar] [CrossRef]
- Madbouly, H.; Tamam, S.; Abd-El-Gaid, A. Isolation and identification of bovine herpes virus-1 (BHV-1) from semen of foreign breeds bulls. J. Vet. Med. Res. 2008, 18, 22–27. [Google Scholar] [CrossRef]
- Magouz, A.; Medhat, S.; Abou Asa, S.; Desouky, A. Detection of infectious laryngotracheitis virus (Gallid herpesvirus-1) from clinically infected chickens in Egypt by different diagnostic methods. Iran. J. Vet. Res. 2018, 19, 194. [Google Scholar]
- El-sayyad, M.I.; Mandour, M.F.; Shahein, M.A.; Abd Eldaim, M.M.; Abdelwahab, S.A.; El-Tarabili, M.M. Detection of equine herpes virus-1 in Arabian horses from different localities in Egypt. Anim. Health Res. J. 2015, 3, 15–30. [Google Scholar]
- Gagnon, C.A.; Tremblay, J.; Larochelle, D.; Music, N.; Tremblay, D. Identification of a novel herpesvirus associated with cutaneous ulcers in a fisher (Martes pennanti). J. Vet. Diagn. Investig. 2011, 23, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Akter, T.; Tabassum, S.; Jahan, M.; Nessa, A.; Islam, M.; Giasuddin, M. Isolation of herpes simplex viruses by chick embryo culture. Mymensingh Med. J. 2013, 22, 365–369. [Google Scholar] [PubMed]
- Bergmann, M.; Speck, S.; Rieger, A.; Truyen, U.; Hartmann, K. Antibody response to feline herpesvirus-1 vaccination in healthy adult cats. J. Feline Med. Surg. 2020, 22, 329–338. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Target Gene | Amplified Product | Reference |
|---|---|---|---|---|
| FHV-F | GACGTGGTGAATTATCAGC | TK gene | 293 bp | [30] |
| FHV-R | CAACTAGATTTCCACCAGGA | TK gene |
| % Diversity | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| % Identity | 1-FHV 1 Egypt/2021/1 | - | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 26 | 25 |
| 2-FHV 1 Egypt/2021/2 | 100 | - | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 26 | 25 | |
| 3-FHV 11Tur/KaysS9-68 | 99 | 99 | - | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 32 | 31 | |
| 4-FHV 1 KANS_02 | 100 | 100 | 99 | - | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 26 | 25 | |
| 5-FHV 1 PHIL_03 | 100 | 100 | 99 | 100 | - | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 26 | 25 | |
| 6-FHV 1 FeligenRCP Virbac | 100 | 100 | 99 | 100 | 100 | - | 0 | 1 | 1 | 0 | 0 | 0 | 26 | 25 | |
| 7-FHV1Companion Intervet | 100 | 100 | 99 | 100 | 100 | 100 | - | 1 | 1 | 0 | 0 | 0 | 26 | 25 | |
| 8-FHV 1 RJVAC/13 | 99 | 99 | 99 | 99 | 99 | 99 | 99 | - | 1 | 1 | 1 | 1 | 26 | 25 | |
| 9-FHV 1 tiger-SZ-12 | 99 | 99 | 99 | 99 | 99 | 99 | 99 | 99 | - | 1 | 1 | 1 | 26 | 25 | |
| 10-FHV 1 BACv7 | 100 | 100 | 99 | 100 | 100 | 100 | 100 | 99 | 99 | - | 0 | 0 | 31 | 30 | |
| 11-FHV 1 VN1 | 100 | 100 | 99 | 100 | 100 | 100 | 100 | 99 | 99 | 100 | - | 0 | 26 | 25 | |
| 12-FHV 1 CH-B | 100 | 100 | 99 | 100 | 100 | 100 | 100 | 99 | 99 | 100 | 100 | - | 26 | 25 | |
| 13-Canine herpesvirus 0194 | 74 | 74 | 68 | 74 | 74 | 74 | 74 | 74 | 74 | 69 | 74 | 74 | - | 2 | |
| 14-D83054.1Canine herpesvirus | 75 | 75 | 69 | 75 | 75 | 75 | 75 | 75 | 75 | 70 | 75 | 75 | 98 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magouz, A.; Lokman, M.S.; Albrakati, A.; Elmahallawy, E.K. First Report of Isolation and Molecular Characterization of Felid Herpesvirus-1 from Symptomatic Domestic Cats in Egypt. Vet. Sci. 2022, 9, 81. https://doi.org/10.3390/vetsci9020081
Magouz A, Lokman MS, Albrakati A, Elmahallawy EK. First Report of Isolation and Molecular Characterization of Felid Herpesvirus-1 from Symptomatic Domestic Cats in Egypt. Veterinary Sciences. 2022; 9(2):81. https://doi.org/10.3390/vetsci9020081
Chicago/Turabian StyleMagouz, Asmaa, Maha S. Lokman, Ashraf Albrakati, and Ehab Kotb Elmahallawy. 2022. "First Report of Isolation and Molecular Characterization of Felid Herpesvirus-1 from Symptomatic Domestic Cats in Egypt" Veterinary Sciences 9, no. 2: 81. https://doi.org/10.3390/vetsci9020081
APA StyleMagouz, A., Lokman, M. S., Albrakati, A., & Elmahallawy, E. K. (2022). First Report of Isolation and Molecular Characterization of Felid Herpesvirus-1 from Symptomatic Domestic Cats in Egypt. Veterinary Sciences, 9(2), 81. https://doi.org/10.3390/vetsci9020081