
Differences in the microRNAs Levels of Raw Milk from Dairy Cattle Raised under Extensive or Intensive Production Systems

 , , , , ,
, , , , ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. RNA Isolation
2.3. Search for miRNAs Candidates from Sequencing
2.3.1. RNA Sequencing
2.3.2. Identification of Reference miRNAs for qPCR Normalization
2.3.3. Identification of miRNAs Whose Levels Differed between Production Systems
2.4. Validation of Candidate miRNAs Using RT-qPCR
2.4.1. RT-qPCR Analysis
2.4.2. Selection of Stable Reference miRNAs. GeNorm Analysis
2.4.3. miRNAs Levels Normalization and Estimation
2.5. Prediction and Functional Analysis of Genes Targeted by miRNAs
3. Results
3.1. miRNAs Levels in Fat and Cellular Fractions of Milk
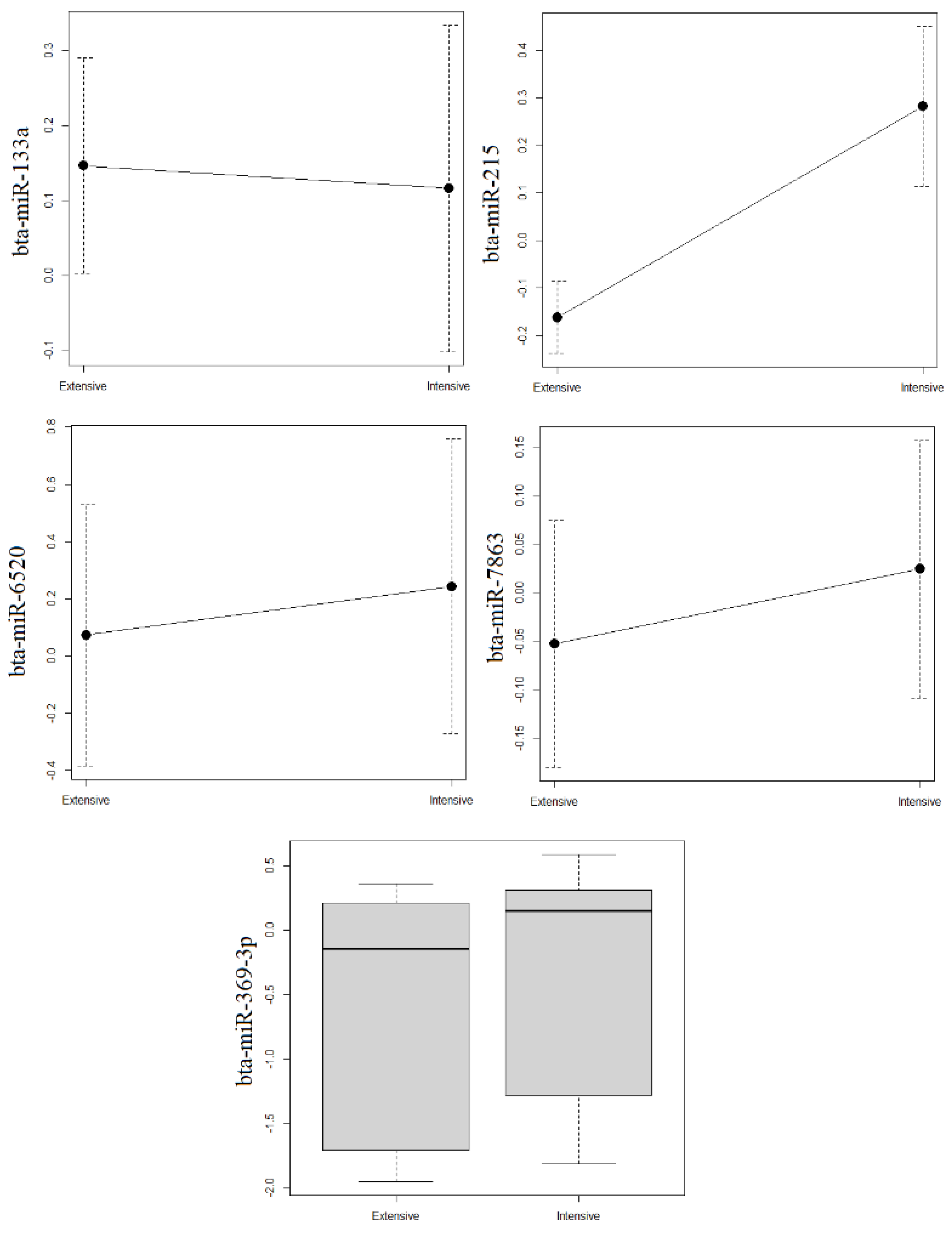
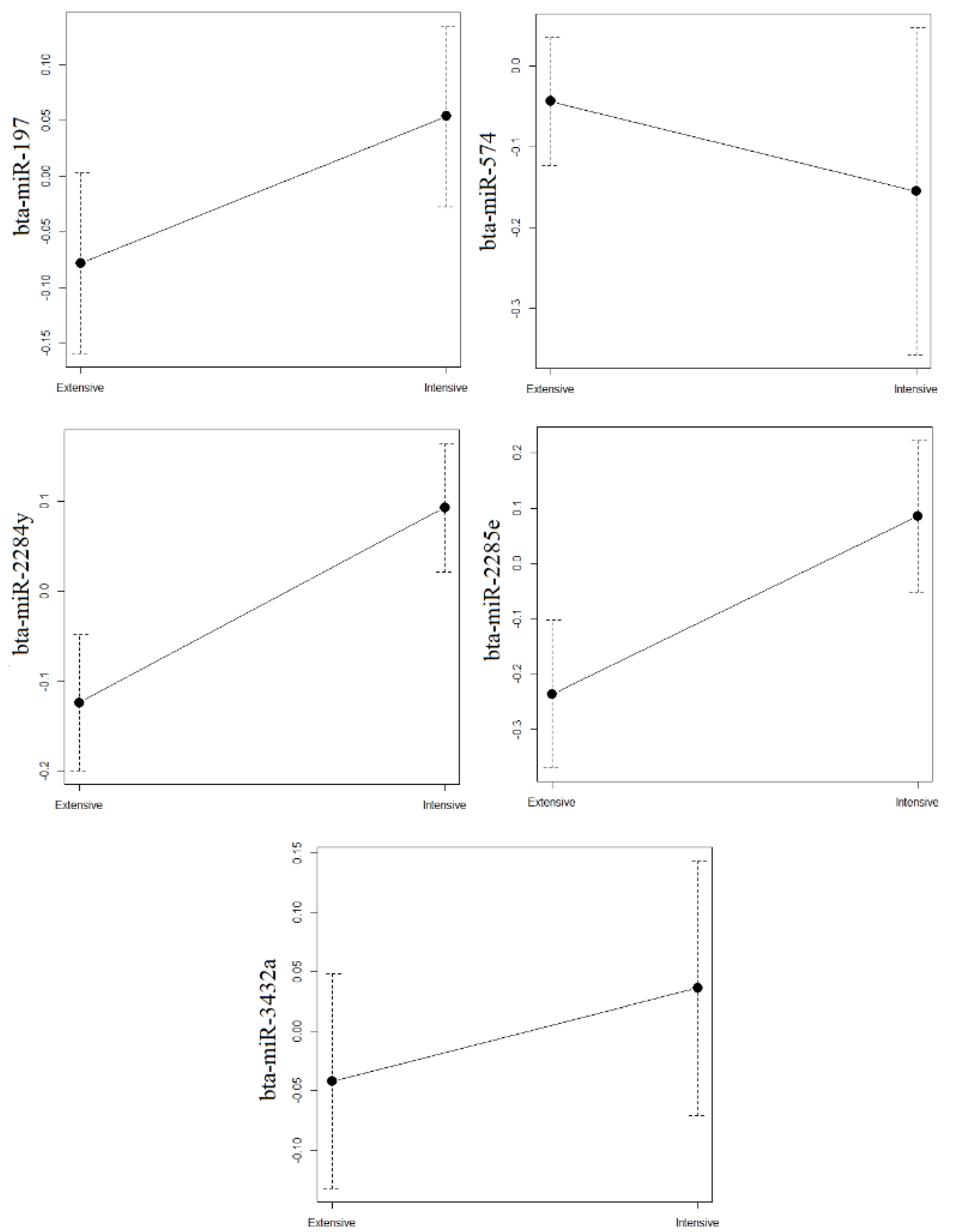
3.2. Validation of miRNAs Whose Levels Differed between Intensive and Extensive Production
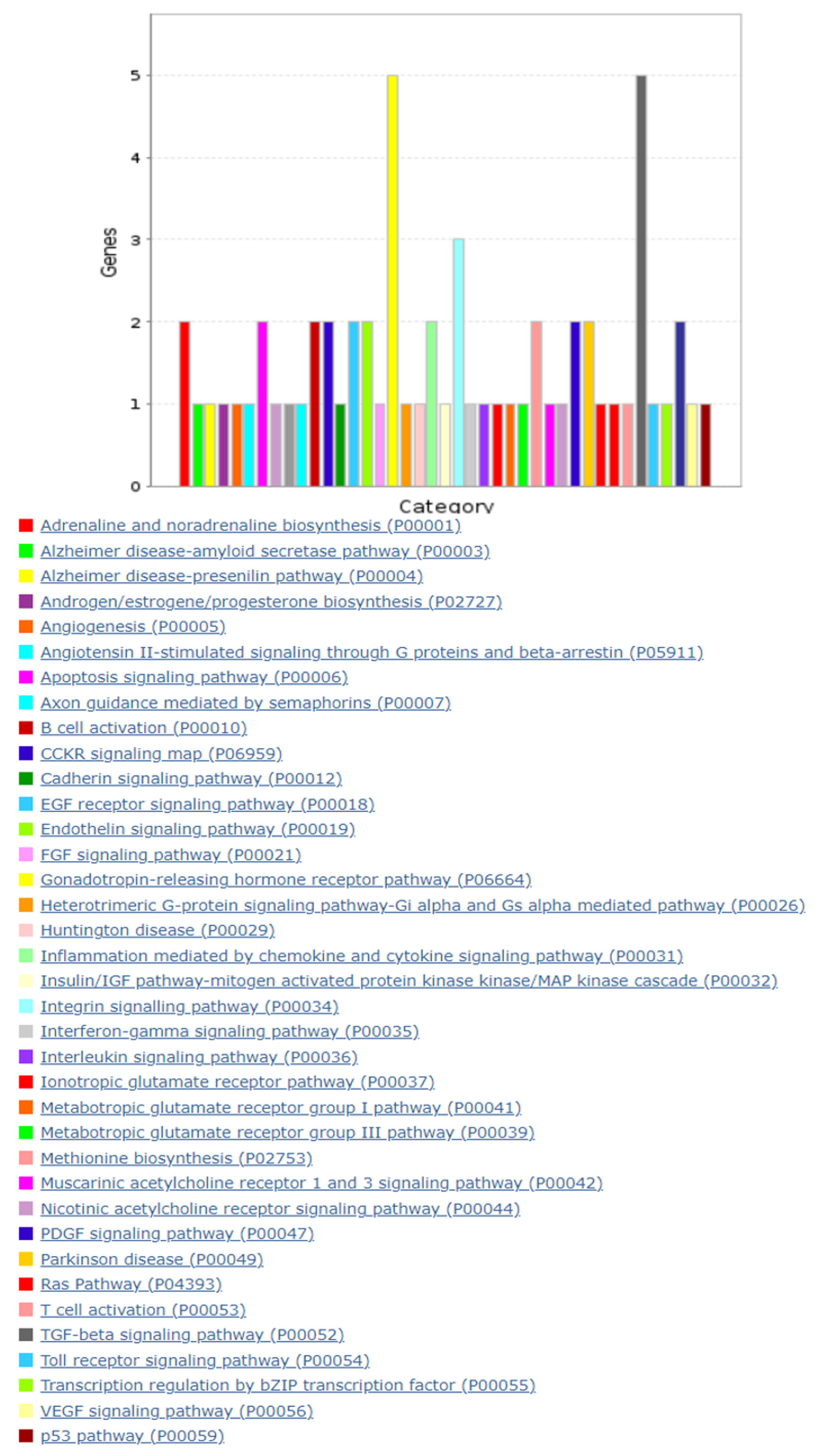
3.3. Putative Target Gene and Pathway Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luykx, D.M.A.M.; van Ruth, S.M. An Overview of Analytical Methods for Determining the Geographical Origin of Food Products. Food Chem. 2008, 107, 897–911. [Google Scholar] [CrossRef]
- Valenti, B.; Martin, B.; Andueza, D.; Leroux, C.; Labonne, C.; Lahalle, F.; Larroque, H.; Brunschwig, P.; Lecomte, C.; Brochard, M.; et al. Infrared Spectroscopic Methods for the Discrimination of Cows’ Milk According to the Feeding System, Cow Breed and Altitude of the Dairy Farm. Int. Dairy J. 2013, 32, 26–32. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a Big Role in Gene Regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Thorning, T.K.; Raben, A.; Tholstrup, T.; Soedamah-Muthu, S.S.; Givens, I.; Astrup, A. Milk and Dairy Products: Good or Bad for Human Health? An Assessment of the Totality of Scientific Evidence. Food Nutr. Res. 2016, 60, 32527. [Google Scholar] [CrossRef]
- López de Las Hazas, M.C.; Del Pozo-Acebo, L.; Hansen, M.S.; Gil-Zamorano, J.; Mantilla-Escalante, D.C.; Gómez-Coronado, D.; Marín, F.; Garcia-Ruiz, A.; Rasmussen, J.T.; Dávalos, A. Dietary bovine milk miRNAs transported in extracellular vesicles are partially stable during GI digestion, are bioavailable and reach target tissues but need a minimum dose to impact on gene expression. Eur. J. Nutr. 2022, 61, 1043–1056. [Google Scholar] [CrossRef]
- Billa, P.A.; Faulconnier, Y.; Ye, T.; Chervet, M.; Le Provost, F.; Pires, J.A.A.; Leroux, C. Deep RNA-Seq Reveals MiRNome Differences in Mammary Tissue of Lactating Holstein and Montbéliarde Cows. BMC Genom. 2019, 20, 621. [Google Scholar] [CrossRef]
- Le Guillou, S.; Leduc, A.; Laubier, J.; Barbey, S.; Rossignol, M.N.; Lefebvre, R.; Marthey, S.; Laloë, D.; Le Provost, F. Characterization of Holstein and Normande Whole Milk MiRNomes Highlights Breed Specificities. Sci. Rep. 2019, 9, 20345. [Google Scholar] [CrossRef]
- Miretti, S.; Lecchi, C.; Ceciliani, F.; Baratta, M. MicroRNAs as Biomarkers for Animal Health and Welfare in Livestock. Front. Vet. Sci. 2020, 7, 578193. [Google Scholar] [CrossRef]
- Tzelos, T.; Ho, W.; Charmana, V.I.; Lee, S.; Donadeu, F.X. MiRNAs in Milk Can Be Used towards Early Prediction of Mammary Gland Inflammation in Cattle. Sci. Rep. 2022, 12, 131. [Google Scholar] [CrossRef]
- Muroya, S.; Ogasawara, H.; Hojito, M. Grazing Affects Exosomal Circulating MicroRNAs in Cattle. PLoS ONE 2015, 10, e0136475. [Google Scholar] [CrossRef]
- Muroya, S.; Shibata, M.; Hayashi, M.; Oe, M.; Ojima, K. Differences in Circulating MicroRNAs between Grazing and Grain-Fed Wagyu Cattle Are Associated with Altered Expression of Intramuscular MicroRNA, the Potential Target PTEN, and Lipogenic Genes. PLoS ONE 2016, 11, e0162496. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Dudemaine, P.L.; Zhao, X.; Lei, C.; Ibeagha-Awemu, E.M. Comparative Analysis of the MiRNome of Bovine Milk Fat, Whey and Cells. PLoS ONE 2016, 11, e0154129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous Plant MIR168a Specifically Targets Mammalian LDLRAP1: Evidence of Cross-Kingdom Regulation by MicroRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef]
- Chen, X.; Gao, C.; Li, H.; Huang, L.; Sun, Q.; Dong, Y.; Tian, C.; Gao, S.; Dong, H.; Guan, D.; et al. Identification and Characterization of MicroRNAs in Raw Milk during Different Periods of Lactation, Commercial Fluid, and Powdered Milk Products. Cell Res. 2010, 20, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Bovine Milk Contains MicroRNA and Messenger RNA That Are Stable under Degradative Conditions. J. Dairy Sci. 2012, 95, 4831–4841. [Google Scholar] [CrossRef]
- Bauman, D.E.; Harvatine, K.J.; Lock, A.L. Nutrigenomics, Rumen-Derived Bioactive Fatty Acids, and the Regulation of Milk Fat Synthesis. Annu. Rev. Nutr. 2011, 31, 299–319. [Google Scholar] [CrossRef] [PubMed]
- Ollier, S.; Leroux, C.; de la Foye, A.; Bernard, L.; Rouel, J.; Chilliard, Y. Whole Intact Rapeseeds or Sunflower Oil in High-Forage or High-Concentrate Diets Affects Milk Yield, Milk Composition, and Mammary Gene Expression Profile in Goats. J. Dairy Sci. 2009, 92, 5544–5560. [Google Scholar] [CrossRef] [PubMed]
- Quan, S.; Nan, X.; Wang, K.; Zhao, Y.; Jiang, L.; Yao, J.; Xiong, B. Replacement of Forage Fiber with Non-Forage Fiber Sources in Dairy Cow Diets Changes Milk Extracellular Vesicle-MiRNA Expression. Food Funct. 2020, 11, 2154–2162. [Google Scholar] [CrossRef] [PubMed]
- Marin, E.C. Sistema de Explotación Ganadera: Notas En Torno a Su Concepto. Lurralde 1996, 19, 89–104. [Google Scholar]
- Ruiz, J.; Herrera, M.P.; Barba, R.; Busqué, J. Definición y Caracterización de la Extensividad en las Explotaciones Ganaderas en España. Situación de la Ganadería Extensiva en España (I); Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente: Madrid, Spain, 2017. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Erlbaum, L., Ed.; Routledge: London, UK, 1988; ISBN 0805802835. [Google Scholar]
- Student. The Probable Error of a Mean. Biometrika 1908, 6, 1–25. [Google Scholar] [CrossRef]
- Pearson, K., VII. Note on Regression and Inheritance in the Case of Two Parents. Proc. R. Soc. Lond. 1895, 58, 240–242. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed]
- Abou el qassim, L. Variaciones de Los Perfiles de MicroARN en la Leche Cruda de Vaca Según el Sistema de Alimentación. Master’s Thesis, University of Zaragoza, Zaragoza, Spain, 2017. [Google Scholar]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. QBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2008, 8, R19. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Barte, D.P. Predicting Effective MicroRNA Target Sites in Mammalian MRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER Version 16: A Revised Family Classification, Tree-Based Classification Tool, Enhancer Regions and Extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef] [PubMed]
- Vicente, F.; Santiago, C.; Jiménez-Calderón, J.D.; Martínez-Fernández, A. Capacity of Milk Composition to Identify the Feeding System Used to Feed Dairy Cows. J. Dairy Res. 2017, 84, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Alsaweed, M.; Lai, C.T.; Hartmann, P.E.; Geddes, D.T.; Kakulas, F. Human Milk MiRNAs Primarily Originate from the Mammary Gland Resulting in Unique MiRNA Profiles of Fractionated Milk. Sci. Rep. 2016, 6, 20680. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C.; Kakulas, F. Milk Exosomes and MicroRNAs: Potential Epigenetic Regulators. In Handbook of Nutrition, Diet, and Epigenetics; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; Volume 3, pp. 1467–1494. ISBN 9783319555300. [Google Scholar]
- Del Pozo-Acebo, L.; de las Hazas, M.C.L.; Tomé-Carneiro, J.; Gil-Cabrerizo, P.; San-Cristobal, R.; Busto, R.; García-Ruiz, A.; Dávalos, A. Bovine Milk-Derived Exosomes as a Drug Delivery Vehicle for Mirna-Based Therapy. Int. J. Mol. Sci. 2021, 22, 1105. [Google Scholar] [CrossRef]
- Knaus, W. Perspectives on Pasture versus Indoor Feeding of Dairy Cows. J. Sci. Food Agric. 2016, 96, 9–17. [Google Scholar] [CrossRef]
- Al-Husseini, W.; Chen, Y.; Gondro, C.; Herd, R.M.; Gibson, J.P.; Arthur, P.F. Characterization and Profiling of Liver MicroRNAs by RNA-Sequencing in Cattle Divergently Selected for Residual Feed Intake. Asian-Australas. J. Anim. Sci. 2016, 29, 1371–1382. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, K.L.; Zheng, X.M.; Li, H.X.; Wang, G.L. Identification and Bioinformatics Analysis of MicroRNAs Associated with Stress and Immune Response in Serum of Heat-Stressed and Normal Holstein Cows. Cell Stress Chaperones 2014, 19, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, Z.; Yang, Z.; Jiang, H.; Chu, S.; Mao, Y.; Li, M.; Chen, Z.; Aboragah, A.; Loor, J.J.; et al. Abundance of Solute Carrier Family 27 Member 6 (: SLC27A6) in the Bovine Mammary Gland Alters Fatty Acid Metabolism. Food Funct. 2021, 12, 4909–4920. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Bello-pérez, E.; Geldsetzer-Mendoza, C.; Cancino-Padilla, N.; Morales, M.S.; Leskinen, H.; Garnsworthy, P.C.; Loor, J.J.; Romero, J. Effects of Dietary Vegetable Oils on Mammary Lipid-Related Genes in Holstein Dairy Cows. Animals 2020, 10, 57. [Google Scholar] [CrossRef]
- Ma, N.; Abaker, J.A.; Wei, G.; Chen, H.; Shen, X.; Chang, G. A High-Concentrate Diet Induces an Inflammatory Response and Oxidative Stress and Depresses Milk Fat Synthesis in the Mammary Gland of Dairy Cows. J. Dairy Sci. 2022, 105, 5493–5505. [Google Scholar] [CrossRef]
- Zhou, Q.; Yang, H.; Pan, H.; Pan, H.; Zhou, J. Exosomes Isolated from the MiR-215-Modified Bone Marrow Mesenchymal Stem Cells Protect H2O2-Induced Rat Myoblasts via the MiR-215/FABP3 Pathway. Exp. Mol. Pathol. 2021, 119, 104608. [Google Scholar] [CrossRef]
- Lu, L.M.; Li, Q.Z.; Huang, J.G.; Gao, X.J. Proteomic and Functional Analyses Reveal MAPK1 Regulates Milk Protein Synthesis. Molecules 2013, 18, 263–275. [Google Scholar] [CrossRef]
- Hemnes, A.; Fessel, J.; Chen, X.; Zhu, S.; West, J.; Talati, M. BMPR2 Dysfunction Impairs Insulin Signaling and Glucose Homeostasis in Cardiomyocytes. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 318, L429–L441. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Production System | Number of Cows | Milk Production (L/Day/Cow) | Grazing (h/Day) | Grass Silage (kg F */Day/Cow) | Maize Silage (kg F */Day/cow) | Hay (kg F */Day/Cow) | Concentrate (kg F */Day/Cow) |
|---|---|---|---|---|---|---|---|
| Intensive (3 farms) | 51 | 30 | 0 | 17 | 10.0 | 2 | 10 |
| 65 | 28 | 0 | 10 | 15.0 | 10 | 11 | |
| 90 | 29 | 0 | 16 | 20.0 | 2 | 12 | |
| Extensive (3 farms) | 24 | 21 | 20 | 10 | 0.0 | 2 | 7 |
| 14 | 31 | >12 | 14 | 0.0 | 6 | 6 | |
| 15 | 29 | 18 | 15 | 0.0 | 3 | 6 |
| Production System | Number of Cows | Milk Production (L/Day Cow) | Grazing (h/Day) | Grass Silage (kg F */Day/Cow) | Maize Silage (kg F */Day/Cow) | Hay (kg F */Day/Cow) | Concentrate (kg F */Day/Cow) |
|---|---|---|---|---|---|---|---|
| Intensive (10 farms) | 124 | 37.4 | 0 | 8.0 | 30.0 | 0.8 | 11.5 |
| 116 | 37.0 | 0 | 10.0 | 30.0 | 0.0 | 10.5 | |
| 90 | 29.0 | 0 | 16.0 | 20.0 | 3.0 | 12.0 | |
| 240 | 36.0 | 0 | 10.0 | 16.0 | 2.5 | 12.0 | |
| 250 | 38.0 | 0 | 12.0 | 30.0 | 0.9 | 12.3 | |
| 37 | 27.0 | 0 | 14.0 | 28.0 | 0.0 | 10.5 | |
| 110 | 30.0 | 0 | 16.0 | 20.0 | 0.0 | 11.0 | |
| 72 | 28.0 | 0 | 15.0 | 20.0 | 2.5 | 12.0 | |
| 118 | 36.0 | 0 | 16.0 | 22.0 | 0.0 | 10.5 | |
| 124 | 37.0 | 0 | 11.0 | 20.0 | 4.5 | 12.0 | |
| Extensive (10 farms) | 20 | 21.0 | 6 | 10.0 | 0.0 | 6.8 | 6.5 |
| 24 | 21.0 | 20 | 12.0 | 0.0 | 0.0 | 7.0 | |
| 12 | 26.2 | 22 | 0.0 | 0.0 | 6.0 | 4.0 | |
| 8 | 18.8 | 21 | 0.0 | 0.0 | 4.6 | 4.1 | |
| 35 | 19.5 | 20 | 14.0 | 0.0 | 0.0 | 4.7 | |
| 15 | 29.0 | 18 | 33.0 | 0.0 | 0.7 | 6.2 | |
| 30 | 27.0 | 18 | 0.0 | 0.0 | 4.0 | 8.0 | |
| 7 | 20.0 | 22 | 6.0 | 0.0 | 0.0 | 5.0 | |
| 16 | 23.0 | 21 | 0.0 | 0.0 | 0.0 | 5.0 | |
| 22 | 24.0 | 20 | 0.0 | 0.0 | 0.0 | 6.0 |
| Total Reads | Small RNA Reads | Small RNAs (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total | rRNA | snRNA | snoRNA | tRNA | miRNA | ncRNA | ||||
| Cellular fraction | Intensive Farms | 32,560,190 | 16,693,358 | 0.5 | 12.7 | 3.3 | 5.7 | 45.0 | 2.2 | 31.1 |
| 26,678,803 | 13,626,331 | 0.5 | 8.3 | 2.0 | 4.7 | 64.3 | 0.9 | 19.8 | ||
| 23,616,528 | 11,871,084 | 0.5 | 16.5 | 3.7 | 5.5 | 37.6 | 2.0 | 34.8 | ||
| Extensive Farms | 19,944,125 | 9,753,050 | 0.5 | 13.6 | 3.1 | 5.6 | 51.5 | 1.4 | 24.7 | |
| 21,166,161 | 9,650,109 | 0.5 | 14.9 | 3.9 | 6.6 | 45.4 | 1.4 | 27.8 | ||
| 20,137,934 | 10,053,989 | 0.5 | 15.9 | 3.9 | 6.1 | 36.8 | 2.3 | 34.9 | ||
| Fat fraction | Intensive Farms | 7,620,977 | 3,867,636 | 0.5 | 20.6 | 3.4 | 6.1 | 29.8 | 1.7 | 38.4 |
| 7,209,138 | 3,139,312 | 0.4 | 21.9 | 3.7 | 6.6 | 25.3 | 1.9 | 40.6 | ||
| 6,180,232 | 3,115,110 | 0.5 | 26.0 | 3.8 | 6.0 | 15.8 | 1.6 | 46.9 | ||
| Extensive Farms | 7,056,743 | 2,599,438 | 0.4 | 19.4 | 3.7 | 8.4 | 24.8 | 2.2 | 41.5 | |
| 6,682,264 | 3,279,814 | 0.5 | 25.0 | 4.1 | 7.1 | 16.9 | 2.4 | 44.5 | ||
| 7,035,380 | 3,854,280 | 0.6 | 25.9 | 4.3 | 7.0 | 19.2 | 2.3 | 41.4 | ||
| Small RNA Class | p-Value Based on Student’s t Test | ||
|---|---|---|---|
| Cellular vs. Fat Fraction * | Cellular Fraction: Extensive vs. Intensive | Fat Fraction: Extensive vs. Intensive | |
| rRNA | 0.004 | 0.513 | 0.827 |
| snRNA | 0.196 | 0.268 | 0.184 |
| snoRNA | 0.024 | 0.127 | 0.050 |
| tRNA | 0.004 | 0.827 | 0.513 |
| miRNA | 0.260 | 0.825 | 0.040 |
| Non-coding RNA | 0.004 | 0.827 | 0.513 |
| miRNA | Result for | Ranking According to | Average Ranking | ||||
|---|---|---|---|---|---|---|---|
| Test 1 a | Test 2 b | Test 3 c | Test 1 | Test 2 | Test 3 | ||
| bta-miR-215 * | 3.180 | 0.010 | 0.960 | 1 | 1 | 1 | 1.0 |
| bta-miR-369-3p * | 1.760 | 0.020 | 0.900 | 3 | 2 | 2 | 2.3 |
| bta-miR-6520 * | 1.360 | 0.030 | 0.850 | 4 | 3 | 4 | 3.7 |
| bta-miR-7863 * | 1.970 | 0.080 | 0.860 | 2 | 7 | 3 | 4.0 |
| bta-miR-133a * | 1.300 | 0.040 | 0.840 | 5 | 5 | 5 | 5.0 |
| bta-miR-532 | 1.260 | 0.040 | 0.840 | 6 | 4 | 6 | 5.3 |
| bta-miR-148a | 1.210 | 0.120 | 0.780 | 7 | 13 | 7 | 9.0 |
| bta-miR-138 | 1.000 | 0.070 | 0.770 | 22 | 6 | 8 | 12.0 |
| bta-miR-450a | 1.190 | 0.140 | 0.760 | 8 | 18 | 10 | 12.0 |
| bta-miR-6527 | 1.010 | 0.090 | 0.770 | 21 | 8 | 9 | 12.7 |
| miRNA | Result for | Ranking According to | Average Ranking | ||||
|---|---|---|---|---|---|---|---|
| Test 1 a | Test 2 b | Test 3 c | Test 1 | Test 2 | Test 3 | ||
| bta-miR-574 * | 5.770 | 0.000 | 0.990 | 1 | 1 | 1 | 1.0 |
| bta-miR-3432a * | 5.520 | 0.010 | 0.980 | 2 | 3 | 2 | 2.3 |
| bta-miR-2285e * | 2.540 | 0.010 | 0.950 | 5 | 2 | 3 | 3.3 |
| bta-miR-197 * | 1.970 | 0.010 | 0.920 | 6 | 4 | 5 | 5.0 |
| bta-miR-2284y * | 2.750 | 0.020 | 0.940 | 3 | 8 | 4 | 5.0 |
| bta-miR-219 | 1.740 | 0.010 | 0.910 | 9 | 5 | 7 | 7.0 |
| bta-miR-2397-3p | 1.770 | 0.020 | 0.900 | 8 | 7 | 8 | 7.7 |
| bta-miR-2308 | 2.560 | 0.050 | 0.910 | 4 | 14 | 6 | 8.0 |
| bta-miR-2419-5p | 1.620 | 0.020 | 0.890 | 11 | 6 | 9 | 8.7 |
| bta-miR-2409 | 1.790 | 0.040 | 0.890 | 7 | 12 | 10 | 9.7 |
| Milk Fraction | miRNA | Coefficient of Variation |
|---|---|---|
| Fat | bta-miR-532 | 0.060 |
| bta-miR-151-3p | 0.070 | |
| bta-miR-27b | 0.090 | |
| bta-miR-103 | 0.090 | |
| bta-miR-30a-5p | 0.090 | |
| bta-miR-99a-3p | 0.090 | |
| Cellular | bta-miR-103 | 0.080 |
| bta-miR-107 | 0.090 | |
| bta-miR-181a | 0.090 | |
| bta-miR-28 | 0.100 | |
| bta-miR-345-3p | 0.100 | |
| bta-miR-28342 | 0.100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou el qassim, L.; Alonso, J.; Zhao, K.; Le Guillou, S.; Diez, J.; Vicente, F.; Fernández-Sanjurjo, M.; Iglesias-Gutiérrez, E.; Guan, L.; Royo, L.J. Differences in the microRNAs Levels of Raw Milk from Dairy Cattle Raised under Extensive or Intensive Production Systems. Vet. Sci. 2022, 9, 661. https://doi.org/10.3390/vetsci9120661
Abou el qassim L, Alonso J, Zhao K, Le Guillou S, Diez J, Vicente F, Fernández-Sanjurjo M, Iglesias-Gutiérrez E, Guan L, Royo LJ. Differences in the microRNAs Levels of Raw Milk from Dairy Cattle Raised under Extensive or Intensive Production Systems. Veterinary Sciences. 2022; 9(12):661. https://doi.org/10.3390/vetsci9120661
Chicago/Turabian StyleAbou el qassim, Loubna, Jaime Alonso, Ke Zhao, Sandrine Le Guillou, Jorge Diez, Fernando Vicente, Manuel Fernández-Sanjurjo, Eduardo Iglesias-Gutiérrez, Leluo Guan, and Luis J. Royo. 2022. "Differences in the microRNAs Levels of Raw Milk from Dairy Cattle Raised under Extensive or Intensive Production Systems" Veterinary Sciences 9, no. 12: 661. https://doi.org/10.3390/vetsci9120661
APA StyleAbou el qassim, L., Alonso, J., Zhao, K., Le Guillou, S., Diez, J., Vicente, F., Fernández-Sanjurjo, M., Iglesias-Gutiérrez, E., Guan, L., & Royo, L. J. (2022). Differences in the microRNAs Levels of Raw Milk from Dairy Cattle Raised under Extensive or Intensive Production Systems. Veterinary Sciences, 9(12), 661. https://doi.org/10.3390/vetsci9120661